
Abstract
Dehydrated Antarctic cryptoendolithic communities and colonies of the rock inhabitant black fungi Cryomyces antarcticus (CCFEE 515) and Cryomyces minteri (CCFEE 5187) were exposed as part of the Lichens and Fungi Experiment (LIFE) for 18 months in the European Space Agency's EXPOSE-E facility to simulated martian conditions aboard the International Space Station (ISS). Upon sample retrieval, survival was proved by testing colony-forming ability, and viability of cells (as integrity of cell membrane) was determined by the propidium monoazide (PMA) assay coupled with quantitative PCR tests. Although less than 10% of the samples exposed to simulated martian conditions were able to proliferate and form colonies, the PMA assay indicated that more than 60% of the cells and rock communities had remained intact after the “Mars exposure.” Furthermore, a high stability of the DNA in the cells was demonstrated. The results contribute to assessing the stability of resistant microorganisms and biosignatures on the surface of Mars, data that are valuable information for further search-for-life experiments on Mars. Key Words: Endoliths—Eukaryotes—Extremophilic microorganisms—Mars—Radiation resistance. Astrobiology 15, 1052–1059.
1. Introduction
W
The current surface climate of Mars is characterized by (i) high temperature variations, depending on the time of day, season, and geographical location—the Mars Science Laboratory rover, for example, measured diurnal variations of up to 80°C, from about −70°C at night to up to 10°C at noon (Gómez-Elvira et al., 2014)—(ii) an anoxic atmosphere of 95% CO2 at a pressure that varies, depending on season and time of day, between 600 and 900 Pa (Gómez-Elvira et al., 2014); (iii) cosmic radiation at a mean dose rate of 0.2 mGy/day (Hassler et al., 2014); (iv) and intense solar electromagnetic radiation at wavelength λ > 200 nm at an UV irradiance up to 20 W/m2 at midday (Kuhn and Atreya, 1979; Cockell et al., 2000; Gómez-Elvira et al., 2014). It has been suggested that these harsh conditions could rarely support microbial growth or even survival over extended periods of time (Horneck, 2000; Cockell et al., 2005; Kminek et al., 2010; Rummel et al., 2014).
In preparation of life-detection experiments on Mars (Parnell et al., 2007), terrestrial analogues of putative habitats on Mars have been identified, and the tolerance limits of their microbial communities have been studied. Due to its cold, arid climate and seasonally enhanced UV radiation, Antarctica has been considered an ideal terrestrial model in the quest for life on Mars (Wynn-Williams and Edwards, 2000). In this hostile environment, fungi and cyanobacteria have adopted a strategy to escape most of the stress parameters by colonizing the inside of rocks (Friedmann, 1982). We have selected the Antarctic rock-inhabiting meristematic fungi Cryomyces antarcticus and C. minteri as well as fragments of rocks colonized by the Antarctic cryptoendolithic community (Selbmann et al., 2005, 2011) to test their tolerance under simulated martian conditions, as provided by the EXPOSE-E mission on board the International Space Station (ISS) (Rabbow et al., 2012). Cryomyces antarcticus and C. minteri demonstrated resistance to extreme desiccation under 10−5 Pa vacuum and UV radiation (200–400 nm) during preparatory ground-based tests (Experiment Verification Tests, EVT) for this space experiment (Onofri et al., 2008); they also survived space conditions in low-Earth orbit in the frame of the Lichens and Fungi Experiment (LIFE) on board the ISS (Onofri et al., 2012). These black yeast strains that were exposed to lethal doses of UVC survived, while Saccharomyces pastorianus died (Selbmann et al., 2011).
2. Material and Methods
2.1. Spaceflight data of LIFE
In LIFE, Antarctic black fungi were exposed for 1.5 years to simulated martian environmental conditions during the orbital flight of the EXPOSE-E facility of the European Space Agency (ESA) (Rabbow et al., 2009, 2012). EXPOSE-E was launched on 7 February 2008 with Space Shuttle STS-122 to the ISS. It was mounted on 15 February 2008 by extravehicular activity (EVA) to the balcony of the Columbus module of the ISS as part of the European Technology Exposure Facility (EuTEF), and samples were exposed to a simulated martian environment (Table 1). EXPOSE-E was decommissioned on 1 September 2009, retrieved by EVA on 2 September 2009, and returned to Earth on 12 September 2009 with STS-128. During the 1.5-year mission, the samples were kept in a pressurized, simulated martian atmosphere (1.6% argon, 0.15% oxygen, 2.7% nitrogen, 370 ppm H2O, in CO2 at a pressure of 103 Pa) and exposed to simulated martian UV radiation (Table 1) such that the spectrum of solar extraterrestrial electromagnetic radiation was cut off with optical filters at a wavelength of λ = 200 nm (Rabbow et al., 2012). Some samples were insolated with a reduced irradiance by 3 orders of magnitude by neutral density filters. In addition, dark flight samples were located beneath the insolated ones. Cosmic radiation dose and temperature were recorded as provided under the conditions of the EXPOSE-E mission (Table 1).
2.2. Biological test systems of LIFE
The psychrophilic, UV-resistant, and desiccation-tolerant black meristematic fungi Cryomyces antarcticus CCFEE 515 and C. minteri CCFEE 5187 are known to be good eukaryotic models for astrobiological investigations (Onofri et al., 2004, 2012). They were isolated from the McMurdo Dry Valleys of Antarctica (Southern Victoria Land), which is one of the most hostile environments on Earth (Selbmann et al., 2005) and considered the closest terrestrial analogue for Mars. These fungi excel by an extraordinary ability to survive a number of stresses, such as extreme temperature and high radiation levels, especially when dehydrated (Onofri et al., 2008; Selbmann et al., 2011, 2014).
Two test systems were used in LIFE, as follows:
The first is comprised of dried sandstone fragments that were colonized by a stratified cryptoendolithic microbial community and collected in January 2004 at Battleship Promontory (76°54′37.6″S, 160°55′27.5″E), Southern Victoria Land, Antarctica. The community develops within 10 mm depth below the rock crust and is organized in different colored and biologically distinct bands (Fig. 1). Fungi occupy the dark and white zones of the community.

(
The second test system consists of dried colonies of the microcolonial black yeastlike fungi Cryomyces antarcticus CCFEE 515 and C. minteri CCFEE 5187, both of which dwell cryptoendolithically. They were isolated from sandstone collected in the McMurdo Dry Valleys, Antarctica (Selbmann et al., 2005) and cultivated on malt extract agar. Colonies were nearly spherical with a diameter of approximately 1 mm. Single cells are 10 μm in diameter on average, with a thick black melanized outer wall (Fig. 1). Colonies and colonized rock fragments were obtained and dehydrated at room temperature in silica-gel desiccator according to Onofri et al. (2008, 2012).
2.3. Viability assays of the LIFE biological test systems
After flight and before performing viability assays, colonies were rehydrated for 72 h in 1.5 mL tubes containing 1 mL of 0.9% NaCl solution, at 5°C. For growth tests, the suspension of a single colony was opportunely diluted to reach a concentration of 5 × 104 cells/mL. A 100 μL portion of the suspension, containing 5000 cells, was seeded on malt agar Petri dishes (five replicates) and incubated at 15°C for 2 months, and the colonies formed were counted.
For comparison, the same test was also performed on dehydrated, untreated colonies that were kept in the dark in the laboratory (Dark control, Fig. 2). Viability was expressed as percentage of colony-forming units (CFU). Propidium monoazide (PMA) assay (Mohapatra and La Duc, 2012) was used for evaluating percentage of cells with undamaged cell membranes by quantifying fungal DNA extracted from colonies of C. antarcticus and C. minteri or colonized sandstone fragments. This was performed by adding PMA (Biotium, Hayward, CA), at a final concentration of 200 μM, to both fungal colonies, rehydrated as above, and powdered rock suspensions in phosphate-buffered saline (PBS) solution. PMA penetrates only damaged cell membranes, cross-linking to DNA after light exposure and thereby preventing PCR. Following DNA extraction and purification (Maxwell 16 automatic DNA extraction instrument, Promega, Madison, WI), quantitative polymerase chain reaction (qPCR; BioRad CFX96 real-time PCR detection system) was employed to quantify the number of fungal ribosomal DNA fragments present in samples either treated or not treated with PMA. Genomic DNA was added at a concentration of 0.1 ng to 23 μL of PCR cocktail containing 1X Power Sybr-Green PCR Master Mix (Applied Bios, Foster City, CA), and 1 μL of NS91 forward (5′-GTC CCT GCC CTT TGT ACA CAC-3′) and ITS51 reverse (5′-ACC TTG TTA CGA CTT TTA CTT CCT C-3′) primers, each at 0.02 M final concentration. These primers amplify a 203 bp product spanning the 18S/ITS1 region of rRNA encoding genes.

Viability of C. antarcticus and C. minteri expressed as percentage of CFU. Test was performed in five replicates. Statistical significance was calculated by using the Tukey test comparing dataset from the same organism. ** and * indicate that values are or are not significantly different, respectively, according to the Tukey test (P < 0.05).
A standard qPCR cycling protocol consisting of a hold at 95°C for 10 min, followed by 40 cycles of denaturing at 95°C for 15 s, annealing at 58°C for 20 s, and elongation at 72°C for 15 s was followed. Fluorescence measurements were recorded at the end of each annealing step. At the conclusion of the 40th cycle, a melt curve analysis was performed by recording changes in fluorescence as a function of raising the temperature from 60°C to 95°C in 0.5°C per 5 s increments. All tests were performed in triplicate. Statistical analyses were performed by one-way analysis of variance (ANOVA) and pairwise multiple comparison procedure (Tukey test), carried out by using the statistical software SigmaPlot.
3. Results
After a stay of 1.5 years in space under simulated martian conditions, the samples did not change in shape or color compared to their preflight appearance. Viability of the black meristematic fungi C. antarcticus and C. minteri and the endolithic fungal communities was tested via colony-forming ability test (Fig. 2) and PMA assay (Fig. 3). Cryomyces antarcticus and C. minteri, even if kept in the dark, showed a very low survival (as CFU%) after exposure to the simulated martian atmosphere (gas composition and pressure) with 1.48 ± 0.26% and 0.08 ± 0.06% CFU, respectively (Fig. 2); the PMA assay gave 66.47 ± 6.15% of cells with undamaged membranes for C. antarcticus and 17.29 ± 4.85% for C. minteri (Fig. 3).
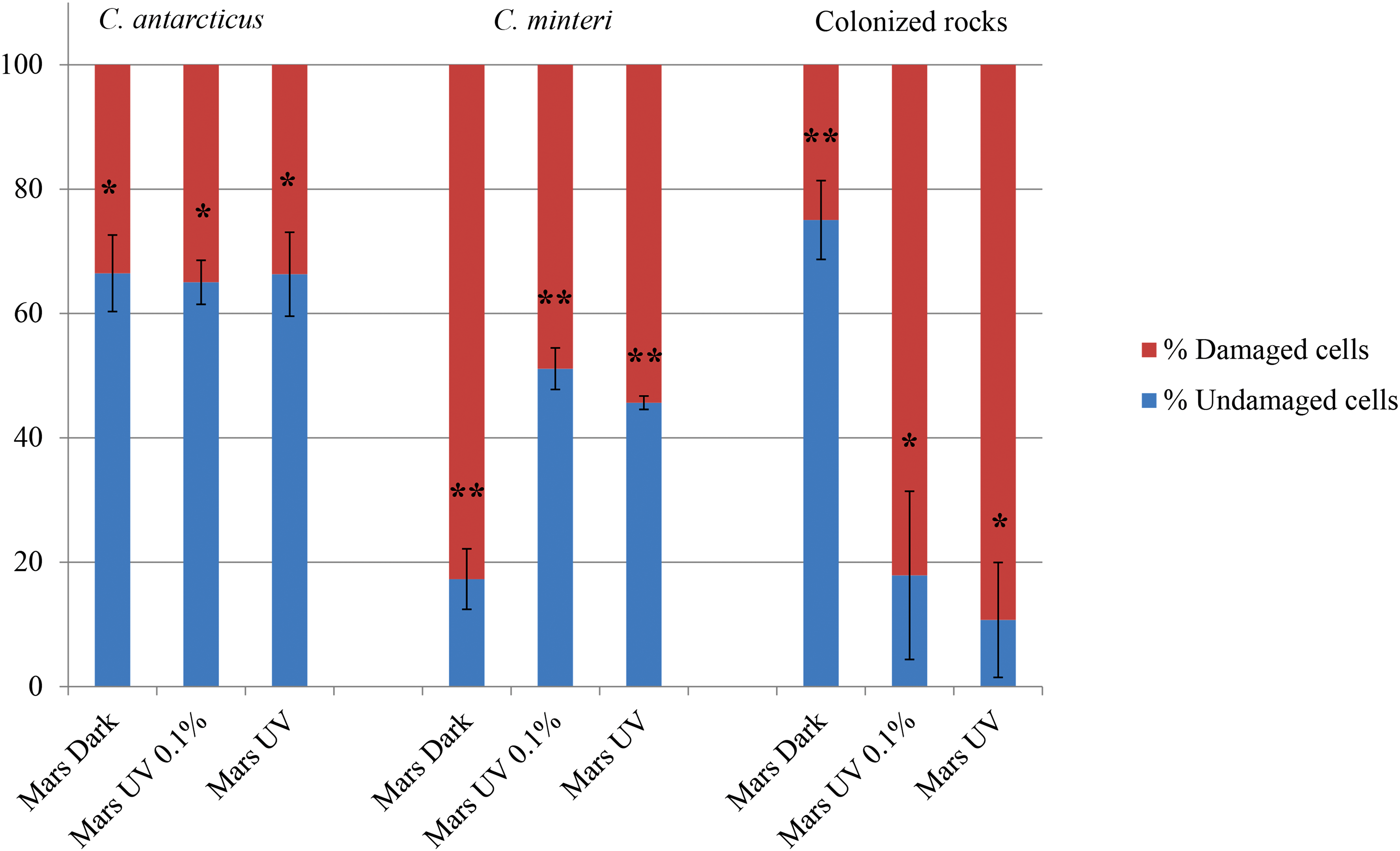
Percentage of intact and damaged cells measured with PMA coupled with qPCR. Test was performed in three replicates. Statistical significance was calculated by using the Tukey test comparing dataset from the same organism or from rock. ** and * indicate that values are or are not significantly different, respectively, according to the Tukey test (P < 0.05).
Different results were obtained for irradiated samples that were exposed to the simulated (full or attenuated) martian UV radiation spectrum (λ > 200 nm).
Survival of fungi, which were exposed beneath neutral density filters (0.1% transmission), was more than 4–5 times higher than that of the dark samples (CFU% of 8.40 ± 1.65 for C. antarcticus and 2.07 ± 0.33 for C. minteri; Fig. 2), while the PMA assay gave a survival of about 3 times higher for C. minteri (51.12 ± 3.34) and nearly identical for C. antarcticus (65.02 ± 3.54; Fig. 3).
Without attenuation (100% solar electromagnetic radiation at λ > 200 nm), survival was in the range of the dark samples: CFU% of 0.87 ± 0.18 for C. antarcticus and of 0.30 ± 0.02 for C. minteri (Fig. 2). In the PMA assay, amplified DNA from cells with undamaged membranes was 66.32 ± 6.75% and 45.66 ± 1.07% for C. antarcticus and C. minteri, respectively (Fig. 3); these values did not significantly differ from those of samples that received 1000 times less solar irradiance.
Colonized sandstone exposed to the simulated martian atmosphere in the dark yielded the highest amount of fungal DNA amplified from intact cells, 75.3 ± 6.40%, while a strong decrease was recorded for insolated samples, for which 17.90 ± 13.52% and 10.72 ± 9.24% were measured for samples exposed to attenuated and full radiation, respectively (Fig. 3).
Survival of the controls was much higher, though it was not 100%; cultivation tests from dried colonies, kept in the dark in the laboratory (Dark controls, Fig. 2), gave 46.50 ± 7.89 and 16.76 ± 2.78 CFU in C. antarcticus and C. minteri, respectively.
4. Discussion
The EXPOSE-E mission on board the ISS provided, for the first time during a space mission, environmental conditions that mimicked most of the parameters of the martian surface: (i) Atmospheric composition and pressure. (ii) Cosmic radiation (maximum dose rate at the sample site: 368 ± 27 μGy/d according to Berger et al., 2012). This dose rate was slightly higher than the 210 ± 40 μGy/d measured by Hassler et al. (2014) in Gale Crater on Mars during the Mars Science Laboratory mission. In addition, the spectra are different; whereas on Mars galactic cosmic rays prevail, protons and electrons of the radiation belts must be added to the galactic radiation source in the orbit of the ISS (Dachev et al., 2012). (iii) UV radiation (maximum 1572 solar constant hours of solar electromagnetic radiation at λ > 200 nm, resulting in a fluence at the sample site of 475 MJ/m2 for 200 nm < λ < 400 nm UV, or 630 kJ/m2 beneath a neutral density filter of 0.1% transmission). Because the martian solar constant amounts to 45% of Earth's solar constant, the applied radiation would be equal to 3493 Mars solar constant hours. (iv) Long-term exposure (559 days in operation, total time in space 583 days). This period corresponds to nearly 1 martian year (687 days).
Temperature was not actively controlled and oscillated between −21.7°C and +42.9°C with a one-time peak at 62°C for a few hours (Rabbow et al., 2012). This range differed substantially from the temperatures on the surface of Mars, which can reach about 20°C as a maximum at noon at the equator and −153°C as a minimum at the poles.
It should be noted that the technical conditions of the ISS did not allow simulating diurnal changes of the environmental parameters as they prevail on Mars. Temperature and insolation varied with the orbit of the ISS, and there was interim shadowing by parts of the ISS, especially the solar panels. In spite of these technical restraints, the EXPOSE-E mission provided a workable tool with which to assess the limits of microbes and microbial communities as well as the stability of biomolecules at the surface of Mars, especially in preparation of future search-for-life ventures.
This Lichens and Fungi Experiment of the EXPOSE-E mission provided, for the first time, data on the viability of rock-dwelling organisms and microbial communities after long-term exposure to martian conditions simulated in space. Even if the “Mars exposed” black fungi Cryomyces antarcticus CCFEE 515 and C. minteri CCFEE 5187 showed fewer than 10% survivors in growth tests, their colony-forming ability was maintained to a certain extent in both strains. This demonstrates that test organisms survived the simulated martian conditions that were applied for 1.5 years and were later able to propagate. In the same experiment, Scalzi et al. (2012) reported that they grew one green alga and one fungus from colonized rock samples after exposure to simulated martian surface conditions. These organisms were identified as representatives of Stichococcus sp. and the lichenized genus Acarospora. In another experiment during the EXPOSE-E mission with Bacillus subtilis spores that were exposed to simulated martian conditions, postflight analysis showed that up to 20% of the spores were able to germinate, grow out, and form colonies, if arranged in multilayers (Horneck et al., 2012).
Although the majority of the “Mars-exposed black fungi” were not able to form colonies, the resulting high quantities of DNA, originating only from cells with undamaged membranes, suggested that cellular integrity was not completely destroyed by the treatments. A similar high fraction of intact cells was found by Brandt et al. (2015) for the lichen Xanthoria elegans, which was exposed to simulated martian conditions in LIFE; after LIVE/DEAD staining, more than 80% of metabolically active mycobiont cells (fungi) were observed, and more than 60% metabolically active photobiont cells (algae).
Survival rates in the growth tests were surprisingly higher in both strains exposed to 0.1% of the radiation than in samples kept in the dark. This is difficult to explain because the increase of 3 orders of magnitude of the radiation (100%) results in a net decrease of survival, as expected (Fig. 2); yet the same result was obtained from the same strains exposed to solar UV radiation under full space conditions in the same experiment LIFE (Onofri et al., 2012).
Remarkably, differences in vitality between C. antarcticus and C. minteri are very similar in all the conditions tested, including control dried colonies that were not exposed.
There was a substantial discrepancy in the viability data obtained by PMA assay or colony test. The PMA assay gave nearly identical values, around 65% cells with undamaged membrane, for all samples exposed to simulated martian conditions in C. antarcticus, regardless of whether they were insolated or not. Similar high values—around 50% cells with undamaged membrane—were found for insolated samples of C. minteri, whereas more than 80% of the dark samples were damaged. Significantly lower was the survival of samples exposed to simulated martian conditions based on cultivation tests where all values remained below 10% of CFU, regardless of whether they were insolated or kept in the dark (Figs. 2 and 3). The differences between the survival rates, measured by the growth test, and the damage to the membranes, measured by PMA test, may be due to the lower sensitivity of the membranes to UV radiation, compared to the ability to multiply; possibly cosmic radiation could be involved in resulting differences.
Propidium monoazide assay, applied to the colonized sandstones, revealed a significant increase in cell membrane damage in treated samples, yet a comparable damage was recorded both in screened and fully exposed samples. This seems to indicate that sandstone does not represent a significant protection to lithobionts. Besides, the rock is mainly composed of quartz, which is the component of the Suprasil filters (>200 nm) used in the experiment; therefore, it is possible that the same wavelengths penetrate the screen and the rock crystals. Furthermore, it should be taken into account that data from rock samples include responses of the whole fungal community and not just the black fungi, which are much more protected by melanins.
The qPCR analyses (performed with the PMA assay) revealed that biomolecules such as DNA are still detectable after 1.5 years of exposure to simulated martian conditions. These results may qualify this biomolecule as a biosignature for future search-for-life extraterrestrial missions; moreover, photoproduct analysis by high-performance liquid chromatography coupled with electrospray ionization–tandem mass spectrometry of DNA extracted from Bacillus subtilis spores flown on EXPOSE-E to simulated martian conditions revealed the presence of a high amount of the 5,6-dihydro-5(α-thyminyl)thymine as the only DNA photoproduct found (Panitz et al., 2015).
It was recently observed that actively growing C. antarcticus (strain CCFEE 534) that was exposed for a short period of time to thermophysical Mars-like conditions responded with a decrease in protein number but recovered the normal metabolic activity within 1 week only (Zakharova et al., 2014).
Therefore, Cryomyces is able to withstand short-term, Mars-simulated ground-based exposition when actively growing and long-term ground-based exposition (up to 1.5 years) when dehydrated.
5. Conclusions and Outlook
The European Space Agency's EXPOSE facility on board the ISS has proven to be an ideal platform for astrobiology investigations in low-Earth orbit. Research areas studied include • chemical evolution of organics in space (Cottin et al., 2012, 2015; Carrasco et al., 2015), • stability of biomolecules in space (Bertrand et al., 2012, 2015), • stability of biomolecules and biosignatures under Mars-like conditions (Noblet et al., 2012; Vergne et al., 2015; and this article), • likelihood of the lithopanspermia hypothesis (Onofri et al., 2012; Tepfer et al., 2012; Wassmann et al., 2012; Panitz et al., 2015), • survival of microorganisms under present-day conditions of Mars (Horneck et al., 2012; Baqué et al., 2013a; Brandt et al., 2015; and this article), • planetary protection requirements (Horneck et al., 2012).
With EXPOSE-R2, the third EXPOSE mission is currently underway on the ISS (ESA, 2014). One of its goals is to extend the current studies by determining the protective role of “martian regolith” on the stability of resistant microorganisms and biosignatures under simulated martian conditions (de Vera et al., 2012; Baqué et al., 2013b).
Footnotes
Acknowledgments
The European Space Agency is acknowledged for the provision and operations of the EXPOSE-E facility. We also thank the Italian National Program of Antarctic Researches (PNRA) and Italian National Antarctic Museum for funding the collection of Antarctic samples, the preservation of the Culture Collection of Fungi from Extreme Environments (CCFEE), and sample analyses. Special thanks are also due to the German Ministry of Economy and Technology BMWi as well as to the HGF-Foundation in the frame of the Helmholtz-Alliance “Planetary Evolution and Life,” which have partly supported the German Co-Is of this work.
Disclosure Statement
No competing financial interests exist for Silvano Onofri, Jean-Pierre de Vera, Laura Zucconi, Laura Selbmann, Giuliano Scalzi, Kasthuri J. Venkateswaran, Elke Rabbow, Rosa de la Torre, and Gerda Horneck.