
Abstract
The processes that govern the formation of stromatolites—structures that may represent macroscopic manifestation of microbial processes and a clear target for astrobiological investigation—occur at various scales (local versus regional), yet determining their relative importance remains a challenge, particularly for ancient deposits and/or if similar deposits are discovered elsewhere in the Solar System. We build upon the traditional multiscale level approach of investigation (micro-, meso-, macro-, mega-) by including a lateral comparative investigational component of fine- to large-scale features to determine the relative significance of local and/or nonlocal controls on stromatolite morphology, and in the process, help constrain the dominant influences on microbialite formation.
In one example of lateral comparative investigation, lacustrine microbialites from the Miocene Barstow Formation (California) display two main mesofabrics: (1) micritic bands that drastically change in thickness and cannot directly be traced between adjacent decimeter-scale subunits and (2) sparry fibrous layers that are strikingly consistent across subunits, suggesting the formation of sparry fibrous layers was influenced by a process larger than the length scale between the subunits (likely lake chemistry).
Microbialites from the uppermost Triassic Cotham Member, United Kingdom, occur as meter-scale mounds and contain a characteristic succession of laminated and dendrolitic mesofabrics. The same succession of laminated/dendrolitic couplets can be traced, not only from mound to mound, but over 100 km, indicating a regional-scale influence on very small structures (microns to centimeters) that would otherwise not be apparent without the lateral comparative approach, and demonstrating that the scale of the feature does not necessarily scale with the scope of the process. Thus, the combination of lateral comparative investigations and multiscale analyses can provide an effective approach for evaluating the dominant controls on stromatolite texture and morphology throughout the rock record and potentially on other planets via rover-scale analyses (e.g., Mars). Key Words: Scale—Stromatolite—Continuity—Laminae—Microbialite. Astrobiology 16, 271–281.
1. Introduction
L
With improved technology and geochemical capabilities, there has been increasing interest in sub-meter-scale features, with a particular rise in microscopic, high-resolution investigations (e.g., Reitner et al., 2011). Focus on sub-meter-scale features (e.g., laminae, centimeter-scale shapes, porosity, branching, microfossils) has preferentially drawn attention to sub-meter-scale local controls (e.g., the role of microbes, diagenetic alteration, local hydrodynamics, high-resolution geochemical changes), potentially masking the role of large-scale, nonlocal controls on the textures in question and possibly resulting in misleading or partial interpretations.
To investigate the under-exploited scale-of-process questions in ancient stromatolites, in this paper we first build upon the traditional multiscale approach (Shapiro, 2000) defined as mega- (meter/outcrop scale), macro- (decimeters), meso- (centimeters), and microscale (millimeters or less) by incorporating a lateral comparative component (Fig. 1). The length scale of the lateral comparative investigation is site-specific and generally defined by how far time-equivalent units can be traced laterally, emphasizing the relative importance of scale and dominant controls that may contribute to stromatolite morphogenesis. We then present examples of a multiscale high-resolution, lateral comparative analysis of stromatolite textures from the Miocene Barstow Formation lacustrine stromatolites and marginal marine stromatolites from the uppermost Triassic Cotham Member of the southwestern United Kingdom. We focus on banded deposits as they are (1) most representative of deposits found throughout most of Earth history (Riding, 2011) and (2) deposits that are increasingly difficult to reproduce in the laboratory and are rarely found lithifying in modern environments, warranting alternate, complementary methods of investigation. We present the lateral comparative analysis as the next step in the now widely used hierarchical scheme presented by Shapiro (2000).
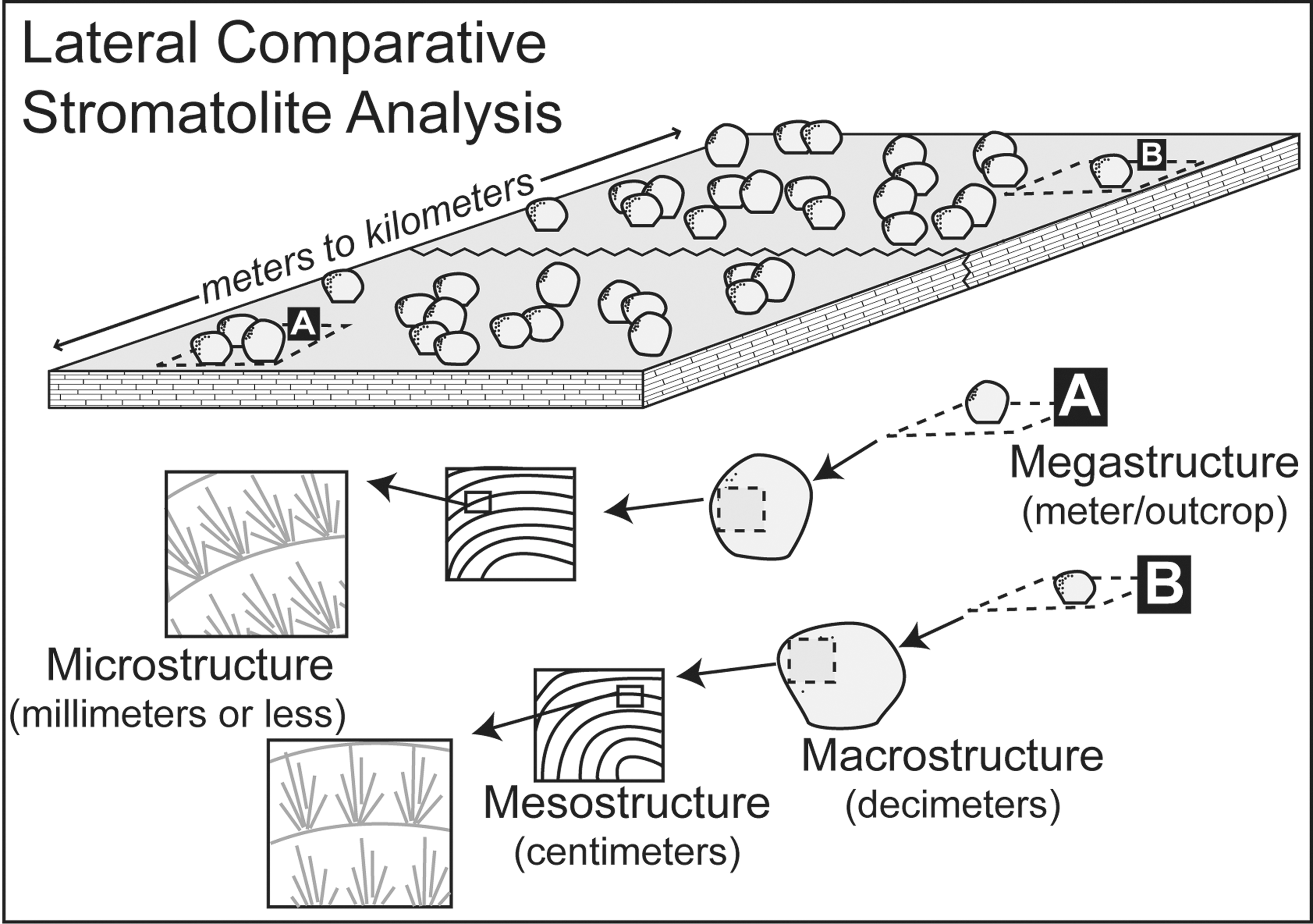
2. Scales of Control and Lateral Continuity
In most depositional systems, morphological attributes of stromatolites are controlled by changes in organic, inorganic, and potentially postdepositional processes, each of which may act at various scales and exert varying degrees of control on the fabric (Table 1). Here, we define a local process as something that operates at the scale of the stromatolitic structure or smaller (thus, most local processes would be meter-scale or smaller; an example might be the microbes that influence stromatolite texture, or local waves that might shape the morphology). Nonlocal, on the other hand, would refer to a process that operates at a scale greater than the stromatolite in question, typically hundreds of meters or greater. Climate or seasonal changes are examples of a nonlocal process that may exert some control on stromatolite morphogenesis.
Microscale processes that can impart local textural changes can include crystal growth/sedimentation, microbial processes that induce crystal growth and/or passively alter the microtexture (Dupraz et al., 2009), and diagenesis. Microscopic processes are usually hierarchical in nature. That is, most microscale processes can be scaled to the millimeter and centimeter scale (e.g., microorganisms can form colonies and biofilms to increase their influence on the accreting substrate, and similarly, although crystals nucleate at the microscale to sub-microscale, they may grow to the centimeter scale or larger). Observations made at the microscale can thus benefit from petrographic analyses that expand the field of view to the millimeter and centimeter scale (e.g., photomosaic petrography) to reveal millimeter- and centimeter-scale textural relationships with highly resolved microscopic elements. Millimeter-scale processes may constitute grain incorporation into the fabric (e.g., trapping of grains), crystal growth, consortia of microbes and biofilms. Similarly, diagenetic processes like dissolution, neomorphism, and weathering may alter the fabric at various scales and additionally contain a critical temporal component, potentially acting upon stromatolite micro- to macroscale features at any time after deposition.
Relatively larger-scale processes, meter-scale or greater, may also contribute to stromatolite fabrics (Table 1). Like local processes, nonlocal processes vary strongly by setting. In lakes and marine environments, for example, currents and wave activity, usually dominant at the scale of meters to tens of meters, may alter stromatolite laminae and cause the structures to be deformed before they are fully lithified. These meter-scale processes are also likely responsible for the meter-scale shapes of biostromes, possibly controlling their meter-scale spatial distribution (e.g., Hoffman, 1976; Andres and Reid, 2006; Bosak et al., 2013; Mariotti et al., 2014). In spring settings, however, the morphology of stromatolites and other associated precipitates is dominantly controlled by flow regime patterns that cause sub-meter-scale lateral variations in temperature, nutrients, turbulence, and so on, resulting in highly distinct and diverse morphologies between adjacent deposits (Fouke et al., 2000).
There are also processes that can occur on the scale of hundreds of meters to kilometers that can be captured by stromatolites and ultimately influence sub-meter-scale textures, like changes in climate and seasonal changes in water chemistry that may cause biogeochemical changes over vast distances. It is well recognized, for example, that in environments where the local controls may be subdued (e.g., closed-basin lakes), given enough external control, stromatolites can record high-frequency climatic changes capturing region-wide, if not global, signals (Benson et al., 1990; Berelson et al., 2009; Petryshyn et al., 2012; Frantz et al., 2014). To establish a clear idea of the scope of the significance of specific sub-meter-scale features, however, it is important to get a relative sense, if not determine, which processes (local or nonlocal) employ the greatest degree of control on the textures being investigated.
Although the formation of stromatolite laminae is the result of a complex mix of various processes (Table 1), it may be possible to dissect the dominant scales of control on stromatolite textures by measuring the degree to which various textures vary laterally along strike. Ecological interactions have been shown to decrease exponentially with increasing geographical distance (Nekola and White, 1999), a process known as distance decay. Similar observations might be expected for two stromatolite deposits forming at the same time in a shared depositional system (Fig. 2), such that the greater the distance between two deposits, the higher the likelihood of physical, chemical, or biological chaos (noise or randomness) between the two, resulting in a higher likelihood for morphological differences (e.g., Pope and Grotzinger, 2000). We know, however, that there are stromatolite-forming systems in which the opposite is observed and immediately adjacent deposits can contain vastly different morphologies (Logan et al., 1964), and likewise, distant deposits can exhibit nearly identical textures (Horodyski, 1983; Sumner, 1997). In cases where lateral continuity is reduced between adjacent deposits, we can assume that local controls are dominant (Table 1). Conversely, in instances where lateral continuity of textures can easily be traced, it is likely that those traceable features are the product of processes whose scale of control extended for at least the minimum distance between the two deposits. We explore the idea of scale and lateral comparisons in the following case studies.
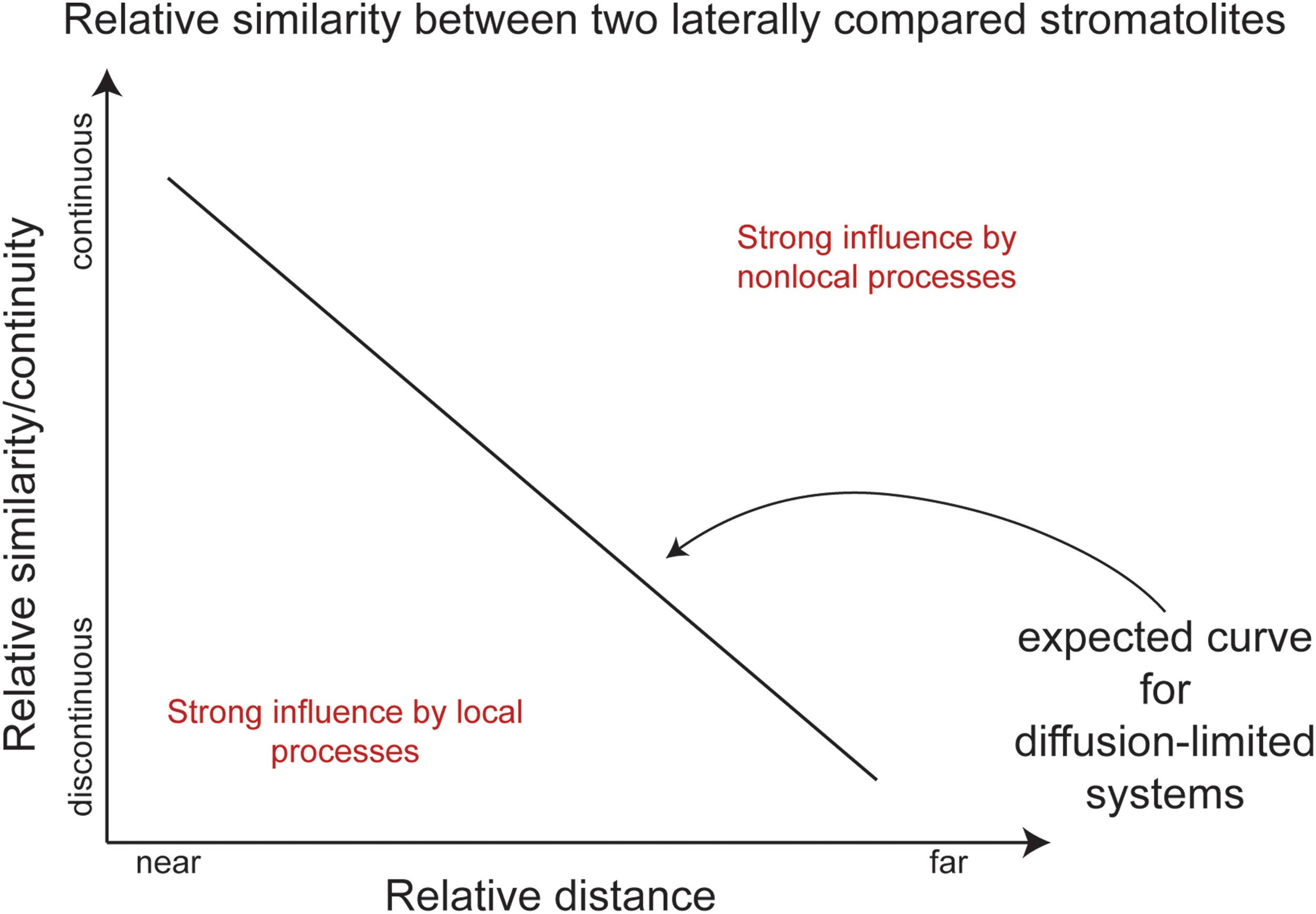
Expected similarity/continuity trend for two laterally compared stromatolites according to distance decay theory. Most environmental systems generally follow this linear trend, except when local or nonlocal processes are dominant (space in red). After Nekola and White (1999). (Color graphics available at
3. Case Study Results
3.1. Miocene Barstow Formation lacustrine tufa
The Barstow Formation outcrops approximately 13 km north of Barstow in the Central Mojave Desert San Bernardino County, California (Fig. 3). It is approximately 1000 m thick and consists of a fining upward succession of alluvial, fluvial, and lacustrine origin (Dibblee, 1968; Woodburne et al., 1990). The original lake system, now tectonically dissected, was thought to be ∼9 km2 (Woodburne et al., 1990). The middle and upper members contain localized calcium carbonate mounds referred to as “tufa” among predominantly sandstone and claystone layers. Three tufa-bearing localities have been described from this region, spanning distances of up to ∼1.5 km apart (Cole et al., 2004). Individual meter-scale mounds are usually domal in shape, laterally discontinuous, and spaced approximately three to tens of meters or more apart. Here, we focus on decimeter-scale stromatolitic subunits (Fig. 4) of a tufa mound (∼2 to 1.5 m thick) that occurs at the base of the Middle Member in the south limb of the Owl Canyon Campground locality (Fig. 3). Tufa mounds at this locality overlie carbonate-cemented vertical pipes that display vertical and concentric growth (Cole et al., 2004). Stromatolitic subunits are cylindrical/nodular in shape and precipitated surface normal relative to the growth surface (Fig. 4). Some of the tufa samples exhibit thin (∼2 mm diameter), hollow, and secondarily infilled tubular internal molds. Vertical cross sections of samples illustrate two dominant mesoscopic textures: dense banded fibrous calcite and porous weakly laminated calcite (Fig. 5). Microscopically, the banded calcite is composed of fibrous spar layers (∼100–500 μm thick) separated by micrite layers (∼50 μm thick). Fibrous spar layers are laterally persistent across subunits and maintain a constant thickness, whereas the micrite layers vary in thickness along individual laminae (Fig. 6).
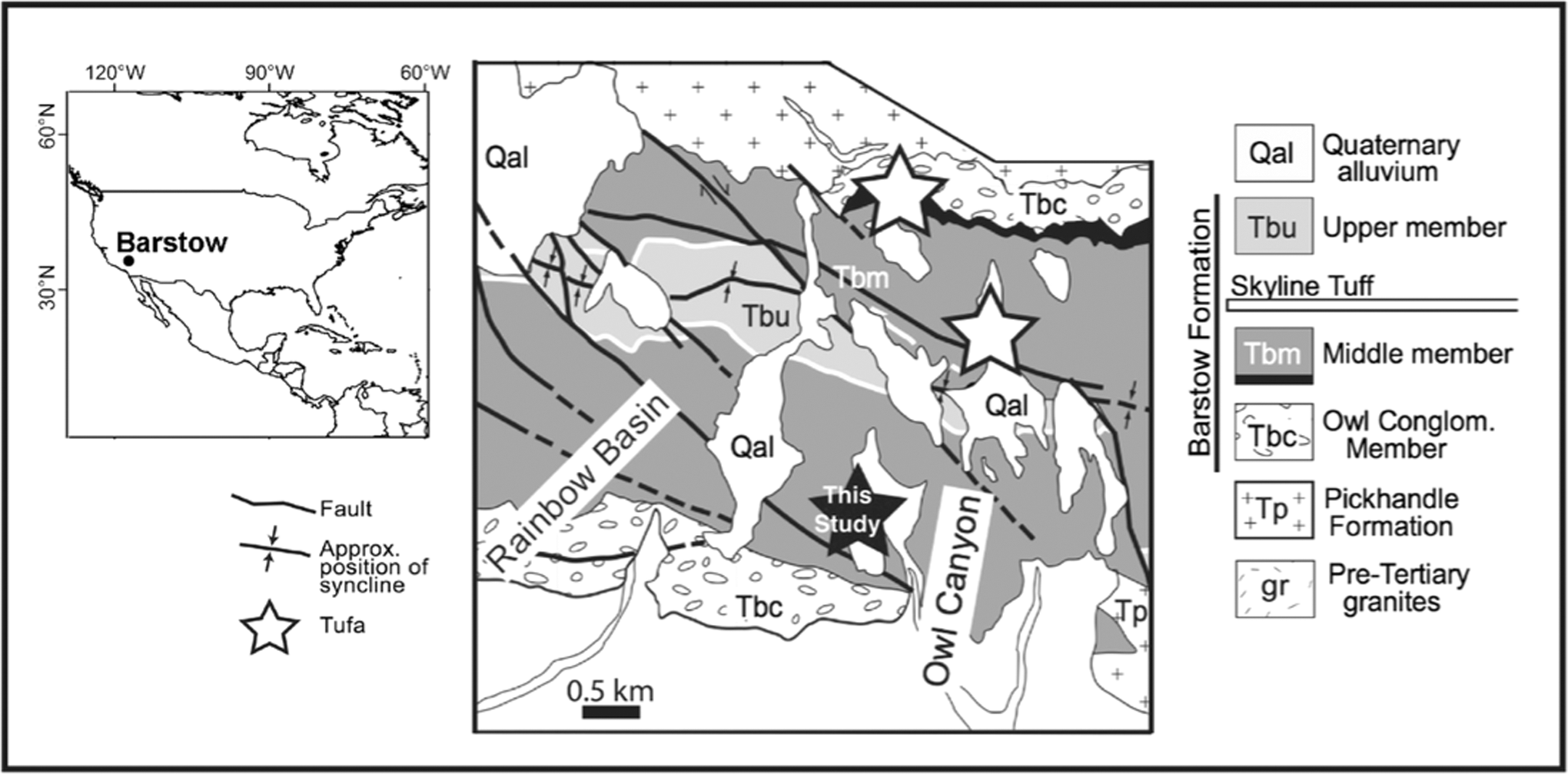
Geologic map of the Barstow Formation, Southern California, denoting tufa locations. After Cole et al. (2005).

In situ macro and mesostructure of Barstow Formation tufa. (

Mesostructure of several Barstow Formation subunits. (

Thin section photomicrographs of Barstow Formation subunits. (
3.2. Uppermost Triassic “Cotham Marble” carbonate microbialites
The uppermost Triassic Cotham Member microbialites, known regionally as the “Cotham Marble,” although they are composed of carbonate (Hamilton, 1961; Wright and Mayall, 1981), are ∼20 cm thick deposits that occur as meter-scale mounds and extend for over 2000 km2 across the southwestern United Kingdom (Fig. 7A) (Ibarra et al., 2014). The deposits formed during the Triassic-Jurassic transition, a mass extinction event and a time of significant biotic and environmental change (Raup and Sepkoski, 1982; Greene et al., 2012). Detailed multiscale investigations of the Cotham Marble from several locations were presented by Ibarra et al. (2014) and revealed a cyclic pattern of sedimentation composed of alternating laminated and dendrolitic mesofabrics (Fig. 7B). The distinct laminated or dendrolitic layers can be traced not only from mound to mound but across distances of over 100 km (Ibarra et al., 2015). Dendrolitic layers are composed of centimeter-scale “bush-like” dendrolites, relatively evenly spaced and surrounded by micrite (Ibarra et al., 2014). The dendrolites are micritic and contain a distinct dark outline. Pyrite-coated filamentous putative microfossils are found restricted to the dendrolitic fabric, suggesting a biogenic origin for the dendrolites. Individual dendrolite bushes are unique from one another in height, width, and branching pattern (Fig. 7B). Microscopically, laminated layers are dominantly micritic, and individual submillimeter laminae in some of the laminated layers can be traced from site to site over distances of at least ∼100 km (Ibarra et al., 2015; Fig. 7C–7E).

Uppermost Triassic “Cotham Marble” microbialites. (
4. Interpretation
At the meter scale, tufa mounds of the Barstow Formation formed when Ca-rich groundwater entered the alkaline saline lake via subaqueous springs. The bicarbonate-rich lake water would have fostered localized carbonate formation at sites where groundwater entered the lake (Park, 1995; Becker et al., 2001; Cole et al., 2004). Localization of stromatolitic tufa deposits around springs is a common formation mechanism observed in many modern and ancient carbonate deposits (Scholl, 1960; Bischoff et al., 1993; Arp et al., 1999), suggesting that the discontinuity of individual mounds was strongly controlled by the presence of spring vents imparting strong macroscopic control. The occurrence of tubular voids and secondarily filled molds suggests aquatic plants served as sites for calcium carbonate nucleation as recognized by Pedone and Caceres (2002). Nucleation around aquatic plants is common in other lacustrine carbonates (Riding, 1979; Arenas et al., 2000). Mesoscopic, laterally continuous dense bands of constant thickness reflect uniform rates of carbonate precipitation controlled at the scale of the meter-scale mound or greater (e.g., reflecting the scales of homogeneity in spring activity, lake chemistry, and/or climate) and would thus plot on the relatively continuous end of the lateral comparative investigation diagram (Fig. 2). On the other hand, uneven micrite laminations would plot on the relatively discontinuous end of the continuity diagram as they are more strongly controlled by local mechanisms (e.g., microbial activity, cement growth) (Figs. 5–6). Thus, a lateral comparison of features across the depositional system gives insight into which features are locally controlled versus more regionally controlled.
Multiscale observations on the Cotham Marble microbialites indicate the decimeter- to meter-scale distribution of the microbialite domes varies at the scale of each mound, suggesting that processes on the scale of about a meter control the macroscopic spatial distribution of the mounds. For example, meter-scale processes (e.g., channels or currents) may have shaped the distances between adjacent mounds (e.g., Hoffman, 1976; Mariotti et al., 2014). Mesoscopically, the changes from laminated to dendrolitic fabrics can be traced over distances on the order of tens of kilometers, reflecting widespread cyclic, nonlocal controls (changes in climate, water chemistry, or seasonality across the various locations). At the submillimeter level, laminated layers reflect influence at the megascale or greater due to remarkable fine-scale continuity from mound to mound and across the various locations, and would thus plot on the relatively continuous and distal region of the continuity diagram (Fig. 2). Instances where individual laminae are truncated or vary in thickness along a single band indicate localized controls on the texture, likely from currents of some kind.
The dendrolite layers reflect a unique interplay between local and nonlocal controls. On the one hand, dendrolitic layers can be traced from mound to mound laterally for tens of kilometers, reflecting regional controls on their distribution. On the other hand, each dendrolitic structure is morphologically unique despite displaying similar dendrolitic and evenly spaced branching patterns throughout the sites investigated, thus highlighting the strong role of local controls on microscale variability. In this unique case, given the excellent preservation of microbial mat textures and putative microfossils, we attribute construction of each dendrolite bush to reflect the branching patterns of microbial mats, under local control. The centimeter-scale spacing of the dendrolite bushes may result from a nearest-neighbor effect, such as diffusion or some limiting nutrient or light limitation. The dendrolite layer can be traced from mound to mound for tens of kilometers, implying some larger-scale regional control. Therefore, while microbial mats can control the resulting sub-centimeter-scale fabrics, their widespread and extraordinarily persistent distribution of similar microscopic textures implies nonlocal processes were the main drivers of formation (Table 1).
5. Discussion
5.1. Significance for scales of control
Previous investigators have utilized the concept of “lateral linkage” between stromatolites and lateral continuity of structures across large and small distances (Logan et al., 1964; Sumner, 1997; Pope and Grotzinger, 2000; Allwood et al., 2007; Kah et al., 2009; Van Kranendonk, 2011) to infer local or environmental controls on morphology. In particular, lateral continuity of stromatolitic domes has been applied to studies of basin analyses (Surdam and Wray, 1976). Additionally, extensive studies on the role of scale-centric investigations across various modern (Rankey, 2002) and ancient sedimentary environments (Drummond and Wilkinson, 1993; Adams and Grotzinger, 1996) have been shown to be useful in providing qualitative and quantitative data on relative controls on geomorphology. However, the use of lateral comparative investigations to determine possible dominant controls on stromatolites has not been explicitly presented, nor has this method been combined with multiscale investigations. Although this method does not reveal explicit processes of control, it can be used to constrain the scale of the principal processes of control and in turn inform our understanding of the environmental scope of the textures under investigation.
Lacustrine and marine stromatolite examples from this study contain banded textures that were common in marine environments for over 2 billion years of Earth's history and continue to be prevalent in modern lacustrine and spring settings today (Riding, 2011). Analyses of lateral continuity of sparry, micritic, and dendrolitic mesofabrics in the examples above demonstrate the utility of the lateral continuity approach in analyses of the relative controls on ancient stromatolites. While the use of observations and analyses that focus on sub-meter-scale features is important for assessing the relative control of local processes (Table 1), considering the potential role of nonlocal processes may ultimately reveal the degree to which the textures under investigation reflect local versus nonlocal processes. Specifically, this study presents the following observations:
(1) Relatively high-frequency changes of dominant controls can occur at small (centimeter-scale) scales (Fig. 5). For instance, lateral variability of textures for the Barstow tufa change at least three times over the scale of about 5 cm (Fig. 5), reflecting relatively high-resolution textural changes in the dominant processes of control, highlighting the importance of high-resolution analyses in paleoenvironmental reconstruction.
(2) The size of the features observed does not necessarily scale with the extent of the dominant processes that control those features. For example, one of the most striking observations of the Barstow tufa subunits is the degree to which mesoscopic textures can be traced laterally (Fig. 5) despite the unique macromorphology of each subunit (Fig. 4). However, at the mesoscale, the process responsible for their overall macroscopic shape is nucleation around aquatic plants, a highly localized phenomenon that leaves a conspicuous imprint on the successive fabric. Conversely, the sparry laminae of the Barstow tufa—and similarly the micritic laminae of the Cotham Marble—are relatively small, on the order of 1 mm or less; yet the dominant processes that influence their formation are large—on the scale of meters, at minimum, for the Barstow system and ∼100 km for the Cotham Marble. This observation highlights that large-scale processes do not necessarily translate into large, conspicuous textures and may consequently go unnoticed when investigating a single subunit or at coarse resolution.
(3) This study presents an example of how lateral comparative investigations can help address how processes at various scales interact to produce the observed features/textures in stromatolites. The dendrolitic layers of the Cotham Marble resulted from the interaction of highly localized and nonlocalized controls. Without the lateral comparative analyses and given the presence of putative microfossils within the dendrolitic fabric (Ibarra et al., 2014), local evidence would suggest that microbial controls are the dominant process in control of the dendrolitic fabric. However, the expansive continuity of the same dendrolitic fabrics over a span of more than 100 km indicates that nonlocal processes (e.g., water chemistry, depth, climate) were the main drivers of their widespread occurrence. The dendrolitic fabric—interpreted as microbial in origin—is in fact embedded within a larger, wide-reaching variable of control that ultimately determines its development.
(4) It is clear that the ability to assess lateral continuity will be specific to each system. A potential setback associated with this method is the fact that laterally continuous sections may not always be available due to limitations in outcrop extent, problems correlating beds from one area to another in a time-correlative (vs. lithostratigraphic) framework, or other postdepositional processes. That being said, developing an awareness of the potential for lateral investigation may lead to a more complete understanding of stromatolite morphogenesis, even when strict application of the method is not available because of outcrop and other eccentricities.
Combining lateral comparative investigation and multiscale observations on stromatolites may allow for more robust estimates of dominant controls. Scaling issues are widely recognized in studies of processes and the development of geomorphic landscapes, and it is understood that they are governed by a range of processes that operate at a range of scales (de Boer, 1992). Unlike studies of modern biological systems that utilize spatial relationships to examine hierarchical patterns of control (e.g., Nekola and White, 1999), the study of stromatolites, which by definition are lithified structures, involves retained morphological attributes of those controls, many of which could be preserved for billions of years (e.g., Allwood et al., 2009). Therefore, lateral investigations may allow for characterization of the nature of spatial variation to more precisely recognize the significance of measurements made at various scales.
As observations on stromatolite textures continue toward increasingly localized and microscopic techniques, it is important to be able to upscale fine-scale measurements and extrapolate the full scope of their potential environmental significance (cf. Andres and Reid, 2006). Relating fine-scale stromatolite patterns (which we are arguably the most familiar with) to broad-scale phenomena and controls wherever applicable remains a critical challenge, yet it is essential to being able to understand their potential as paleoecological archives and astrobiological targets (Cady et al., 2003). Furthermore, a general understanding of regional spatial controls using high-resolution lateral comparative investigations may help reduce the number of factors that must be investigated and also inform at which scales measurements should be done (e.g., Andres and Reid, 2006; Planavsky and Ginsburg, 2009).
Finally, this study shows that, when investigated under the framework of combined lateral comparative and multiscale observations, multiple processes (local vs. nonlocal) may be responsible for controls at various scales, and in most instances determining which specific controls are dominant is not straightforward (Awramik and Grey, 2005). Most ancient stromatolites have experienced varying degrees of diagenesis, which makes it difficult to decipher the relative contribution of primary versus altered fabrics (Grotzinger and Knoll, 1999). Therefore, while lateral comparative analyses may be used to infer the relative dominance of local versus nonlocal controls, they are not necessarily a means of assessing specific growth variables. If a specific process is invoked, other more conclusive methods should be used to assess controls (e.g., Allwood et al., 2009). For instance, high variability between adjacent stromatolites or the observation of wrinkly laminae at the submillimeter scale does not necessarily invoke biogenicity but rather more broadly reflects highly localized controls on variability (Table 1).
5.2. Astrobiological implications
Stromatolites and similar microbialites constitute a key target in the search for life on other planets (Cady et al., 2003), yet our methods for determining the role of life, if any, in the formation of ancient deposits remain equivocal. Conducting a lateral comparative assessment of structures that resemble stromatolites and potential microbialites, if discovered on other planets, will help narrow down the processes of control and therefore constrain their potential to serve as biosignatures. It is clear, however, that to be certain about putative extraterrestrial life, microscopic investigations will be necessary and will likely require sample return. Nonetheless, the degree of lateral variability will allow for an evaluation of the dominant scale of controls on morphological patterns of the structures in question to better assess their potential biogenicity.
Lateral comparative assessments of centimeter- to meter-scale deposits are already feasible on Mars with the present rover capabilities. In a similar method, other studies have looked at image analyses from Opportunity at the outcrop scale to try to discern biogenic versus abiogenic features (Wagstaff and Corsetti, 2010). In instances where kilometer-scale controls can be assessed, this method can be complemented by satellite imagery of laterally continuous beds. In this respect, lateral comparative assessments can prove to be a valuable method that is feasible with rover-scale imagery and integral as a step for assessing sample return priority in the future.
6. Conclusions
We present a combined multiscale and lateral comparative analysis for banded lacustrine stromatolites and marginal marine stromatolites to assess the potential relationships between growth variables and multiscale stromatolite fabrics. Relatively high-resolution analyses of laterally continuous features reveal that (1) lateral patterns are strongly dependent on the nature of the depositional system, (2) some features are controlled by local processes, whereas others are primarily controlled by regional processes, and (3) in some instances, the scale of the feature does not necessarily scale with the scope of the process. These results have implications regarding one of the most longstanding enigmas in the study of ancient stromatolites—determining the relative role of various processes on stromatolite laminae and their overall morphology. The combined use of lateral comparative and multiscale observations thus far remains an underutilized tool for assessing the relative importance of stromatolite textures and for assisting in determining the significance of stromatolites through space and time.
Footnotes
Acknowledgments
We thank two anonymous reviewers for their constructive comments. We also thank Sarah Greene, Carie Frantz, Victoria Petryshyn, David Bottjer, and Will Berelson for valuable discussions. This research was supported by the National Science Foundation (Earth Life Transitions Grant EAR-1338329) and the American Philosophical Society Lewis and Clark Fund for Exploration and Field Research in Astrobiology to Y.I.
Author Disclosure Statement
No competing financial interests exist.