
Abstract
Impact-generated lithologies have recently been identified as viable and important microbial habitats, especially within cold and arid regions such as the polar deserts on Earth. These unique habitats provide protection from environmental stressors, such as freeze-thaw events, desiccation, and UV radiation, and act to trap aerially deposited detritus within the fissures and pore spaces, providing necessary nutrients for endoliths. This study provides the first culture-independent analysis of the microbial community structure within impact-generated lithologies in a Mars analog environment, involving the analysis of 44,534 16S rRNA sequences from an assemblage of 21 rock samples that comprises three shock metamorphism categories. We find that species diversity increases (H = 2.4–4.6) with exposure to higher shock pressures, which leads to the development of three distinct populations. In each population, Actinobacteria were the most abundant (41%, 65%, and 59%), and the dominant phototrophic taxa came from the Chloroflexi. Calculated porosity (a function of shock metamorphism) for these samples correlates (R 2 = 0.62) with inverse Simpson indices, accounting for overlap in populations in the higher shock levels. The results of our study show that microbial diversity is tied to the amount of porosity in the target substrate (as a function of shock metamorphism), resulting in the formation of distinct microbial populations. Key Words: Microbial diversity—Endoliths—Impact melt-rocks—Mars—Astrobiology. Astrobiology 16, 775–786.
1. Introduction
T
Since the initial survey in the late 1970s, much work has also been done to attempt to characterize the microbial diversity of endolithic communities in polar and nonpolar deserts around the world (e.g., Friedmann, 1980, 1982; Friedmann and Kibler, 1980; Friedmann et al., 1993; Wierzchos and Ascaso, 2001; de la Torre et al., 2003; Wierzchos et al., 2006, 2011; Omelon, 2008; Pointing et al., 2009). In a review by Walker and Pace (2007) that focuses on rRNA gene-based studies, the authors point out that these communities are extremely simple in terms of diversity and share compositional similarities with other desert environments. One such example can be found in the genus Chroococcidiopsis, a cyanobacterium that is common in endolithic communities worldwide (Friedmann, 1980). Other members of the Cyanobacteria are also prevalent in both Arctic and Antarctic endolithic ecosystems, such as Leptolyngbya, Gloeocapsa, Trebouxia, Oscillatoria, and Nostoc spp. (Omelon et al., 2006; Pointing et al., 2009; Jungblut et al., 2010). Given these similarities, it is plausible that the physical and chemical conditions of the rock—such as porosity, translucency, hygroscopic properties, and rock type—have a higher influence on the composition of the microbial community than geography (Walker and Pace, 2007; Fierer and Lennon, 2011).
In the last decade, a new habitat has been added to the list of substrates capable of hosting endolithic communities, in the form of impact-generated lithologies. Meteorite impacts are a fundamental, though often overlooked, geological process that affects all rocky and icy planetary bodies. The extreme heat and pressure generated by meteorite impact events results in the shock metamorphism of the rock on both a micro (i.e., mineral) and macro (i.e., whole-rock) scale, the formation of microfractures, changes in density, as well as partial to complete melting of the rock and the generation of glass phases (Kieffer et al., 1976). A subsequent increase in the transmission of light through the substrate due to increased porosity is also seen (Cockell et al., 2002; Pontefract et al., 2014). Of particular interest is the impact generation of endolithic habitats in crystalline rocks, specifically gneisses, which before the impact were not capable of being internally colonized (Cockell et al., 2002, 2005). The impacting process can further result in the generation of transient hydrothermal systems that can lead to the formation of various types of evaporites, which can also provide a viable substrate for microbial colonization (e.g., Pellerin et al., 2009; Rhind et al., 2014). Given that microorganisms capable of endolithic growth have developed this strategy as an adaptation to harsh environmental conditions here on Earth, similar microhabitats may be good places to search for life on Mars, if it had adapted to similarly extreme conditions in the martian environment after 3.8 Ga. The extensive impact cratering that Mars experienced, both during the Late Heavy Bombardment (Kring and Cohen, 2002) and afterward, reveals a planet where the dominant lithology is both shocked and brecciated, creating an excellent environment for microbial colonization (Newsom, 1980; Schultz and Mustard, 2004; Cannon and Mustard, 2015).
Early work on these types of substrates showed the prevalence of endolithic communities in shocked gneisses from the Haughton impact structure, Canada (Cockell et al., 2002, 2005; Cockell, 2004). Similarly, work specifically on impact glasses from the Ries impact structure, Germany, has revealed the presence of putative microtubules, which are likely related to earlier hydrothermal activity within the crater (Sapers et al., 2014). Pontefract et al. (2014) showed that biomass levels increase with increasing shock level as a function of porosity, with the unshocked rocks (crystalline)—mainly hosting an epilithic community—having intact cell counts at ∼106 cells/g of rock, whereas moderately (∼30 GPa) and highly shocked samples (largely amorphous glass, >50 GPa) reached levels of ∼108 and ∼109 cells/g, respectively. The biodiversity of these impact-created habitats, however, has yet to be characterized fully. Fike et al. (2003) performed a preliminary study of the heterotrophic organisms inhabiting shocked versus unshocked lithologies based on rock isolates and showed that most of the organisms within this habitat shared similar metabolic strategies. Community complexity increased in the shocked gneisses, relative to host impact breccias from the same area, which were dominated by three genera: Arthrobacter, Pseudomonas, and Stenotrophomonas. Alternatively, shocked samples contained organisms from the bacterial genera Arthrobacter, Pseudomonas, Bacillus, Stenotrophomonas, Planococcus, Caulobacter, and Janthinobacter with approximately equal levels of abundance. This initial research suggested that the impact event created an environment with its own unique selectional pressures, affecting the species abundance and distribution of microorganisms within the endolithic community. In this study, we show that shock metamorphism affects the composition of the endolithic community, especially with reference to phototrophic bacteria. Here, the first culture-independent analysis of microbial diversity within shocked gneisses from the Haughton impact structure is presented, expanding on a new type of environment within the Arctic tundra as well as elucidating the selective pressures related to shock metamorphism and the importance of such an environment with respect to the search for life on Mars.
2. Methods
2.1. Field site
Samples were collected from multiple sites within the Haughton impact structure (Fig. 1), located in the northwestern region of Devon Island, Nunavut (75°08′N, 87°51′W), in the Canadian High Arctic archipelago (Osinski et al., 2005). (See Supplementary Table S1 for sample locations, where samples with the HMP tag were collected in July/August 2000, DI08 tags were collected in July/August 2008, and all other samples were collected in July/August 2010. Supplementary Data are available online at
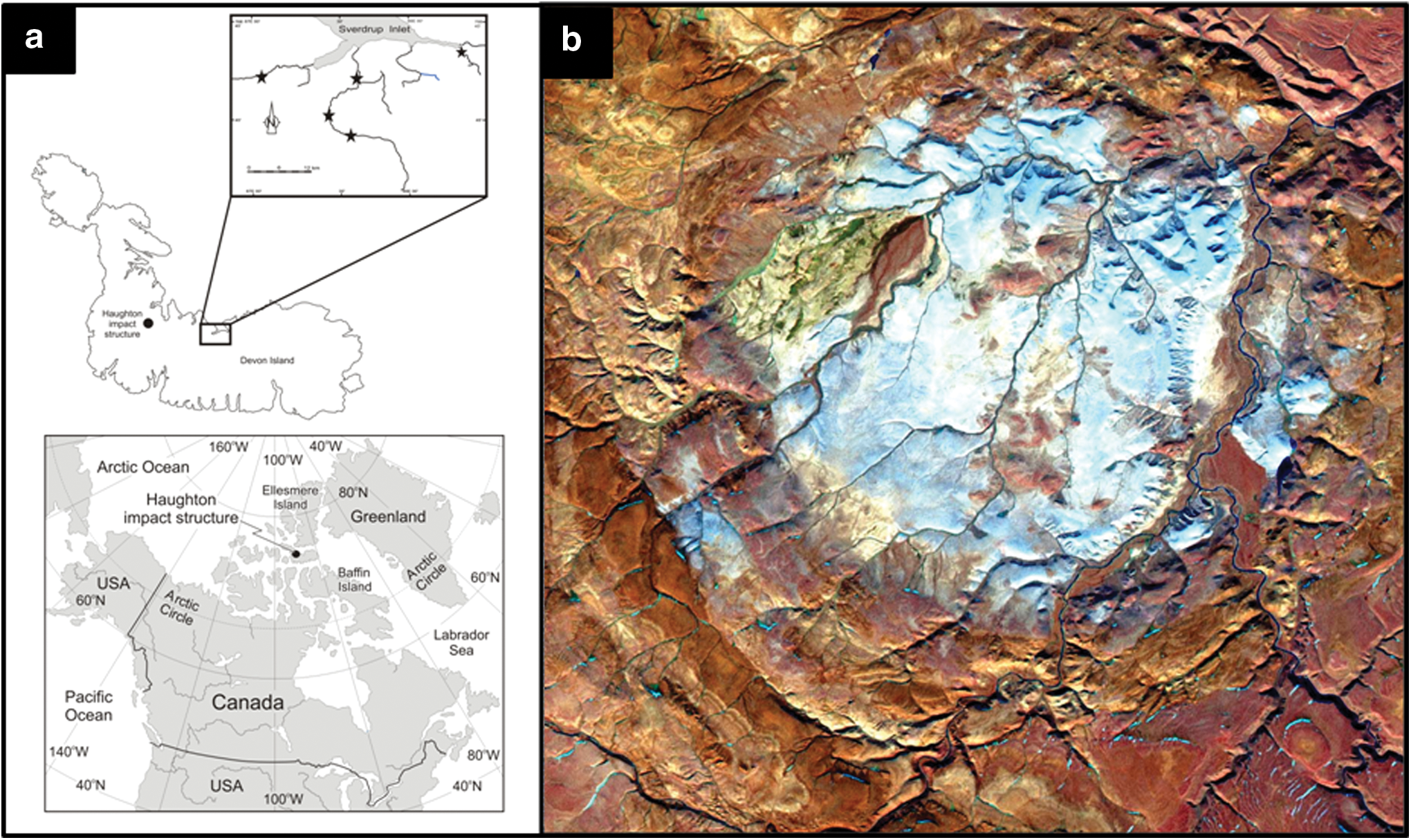
(
2.2. DNA extraction, sequencing, and statistical analysis
An assemblage of 21 samples—divided into three populations—that comprised unshocked (0), moderate shock (levels 3 and 4), and high shock (levels 6 and 7) were examined for bacterial 16S rRNA gene analysis; three samples (1 per population) were examined for fungal and eukaryal SSU and 23S rRNA gene analysis. Ten grams of sample was weighed out and powdered with a sterile alumina mortar and pestle in a laminar flow hood. DNA extraction was then performed using the PowerMax Soil DNA Isolation Kit (MoBio, Carlsbad, CA, USA) according to the manufacturer's instructions. Samples were sequenced using the Roche 454 FLX and Genome Sequencer with TITANIUM sequencing chemistry at Research and Testing Laboratories (Austin, TX, USA). Primer sets used by Research and Testing were as follows: Bacterium-based primers 28F: 5′–GAG TTT GAT CNT GGC TCA G–3′ and 519R: 5′–GTN TTA CNG CGG CKG CTG–3′ (Fan et al., 2012); Algal-based primers (23S plastid, V5) p23SrV_f1: 5′–GGA CAG AAA GAC CCT ATG AA–3′ and p23SrV_r1: 5′–TCA GCC TGT TAT CCC TAG AG–3′ (Sherwood and Presting, 2007); and Fungal-based primers SSUf: 5′–TGG AGG GCA AGT CTG GTG–3′ and SSUr: 5′–TCG GCA TAG TTT ATG GTT AA–3′ (Hume et al., 2012).
The MOTHUR software suite v.1.34.4 was used for the analysis of the 454 data (Schloss et al., 2009), and standard operating procedures were followed with regard to sequence trimming, sequencing error removal, chimera checking, and generation of operational taxonomic units (OTUs), with a 97% cutoff. OTU data were normalized to 541 for each sample, corresponding to the smallest sample size. At this stage, sample SI001A was discarded from further analysis as it was well below the cutoff, containing only 30 OTUs. Alpha diversity indices were calculated for the remaining 20 samples: Shannon's index, Simpson's index, and Chao's index. Good's coverage values were calculated for OTUs at the 0.03 level of definition (Good, 1953). MOTHUR was also used to run an analysis of molecular variance (AMOVA) and to generate a nonmetric multidimensional scaling (NMDS) plot using Bray-Curtis similarities.
2.3. Culturing and microscopy
To determine the viability of the phototrophic endolithic community, small, 1 cm3 rocks from varying shock levels were placed in BG-11 media for enrichment at 24°C for 1 month using a 16 h daylight: 8 h dark cycle. Enrichments were then plated on BG-11 agar to isolate species, which were imaged with a Nikon Eclipse LV100POL compound microscope. Identification of isolates was made in accordance with the work of Castenholz and Waterbury (1989) or Wehr (2003). Isolates were also plated on NO3-deficient agar plates to identify nitrogen fixers within the community [for a list of all nitrogen fixers, see Raymond et al. (2004)]. For visualization of in situ colonization, samples of shocked gneiss were either fractured into small pieces by using a sterile technique and imaged directly with secondary electrons or were embedded in plastic for back-scattered electron microscopy (SEM-BSE) and energy-dispersive X-ray spectroscopy (EDX). Fractured pieces were fixed in glutaraldehyde, dehydrated in ethanol and critical-point dried to preserve cell structure, platinum-coated by a Denton Vacuum Desk II sputter coater at 12 mA for 150 s, and imaged. For embedded samples, fresh faces were fixed in 2% glutaraldehyde, stained with a 0.1 M OsO4-cacodylate buffer, and embedded with EmBed following the protocol from Electron Microscopy Sciences, Hatfield, Pennsylvania (Dykstra and Reuss, 2003).
2.4. Density and porosity
Of the 21 samples used for DNA extraction, 15 were chosen for porosity analysis. Grain density was determined by helium pycnometry (Wilkison et al., 2003; Tamari, 2004) and bulk density determined from volume analyses by X-ray microcomputed tomography 3-D images (McCausland et al., 2010), which could then be used to calculate sample porosity. The advantage of these methods is that they are nondestructive, so the measured properties most directly represent the rock porosity encountered by the microbial community. A Quantachrome Multipycnometer was used to obtain the grain volume for each sample via the ideal gas law through a minimum of five repeat measurements (Tamari, 2004). Each selected sample was also imaged with X-ray microcomputed tomography using a GE eXplore speCZT scanner at the Robarts Research Institute, Western University (London, Canada). The scanner operated with a peak X-ray voltage of 90 kV and a current of 40 mA. In an angular increment of 0.4 degrees, 900 radiograph views were acquired around the sample in about 5 min. These data were reconstructed into a 3-D image volume with an isotropic voxel size of 50 μm. Using the image grayscale values of air and water, we calibrated all the image data into standard Hounsfield units. CT images were analyzed with GE Microview software to obtain sample bulk volumes using a thresholding method (McCausland et al., 2010). Sample porosity was calculated from the grain and bulk densities rather than from the volumes because of slight changes in sample mass between the applications of the methods, especially for friable samples.
3. Results
3.1. Culturing and microscopy
Samples of shocked gneiss placed in BG-11 nutrient media showed considerable growth after 1 month. In most cases, growth of cyanobacteria and algae within the flasks completely engulfed the rock, which was frequently no longer visible. Culturing methods revealed several species of cyanobacteria, which could be determined to the genus level: Oscillatoria spp., Nostoc spp., Anabaena spp. (Fig. 2a), Gloeocapsa spp., Chroococcidiopsis spp., Leptolyngbya spp., and one species of alga, Chlorella sp., which was also detected through pyrosequencing (Fig. 2b). Analysis of 454 sequence reads showed 22 species present within the populations belonging to genera that are known to have N2-fixing representatives, such as Rhizobium, Chroococales, Nostoc, Oscillatoria, Calothrix, Prochlorales, Rhodospirillales, Burkholderiales, and Chromatiales. Isolated colonies from the rocks grown in BG-11 were all plated on nitrogen-deficient BG-11 agar and grown for 3 weeks. Growth of Leptolyngbya and Anabaena were both observed, but no other isolate showed growth, indicating that the species of Oscillatoria in culture was not capable of N2 fixation.
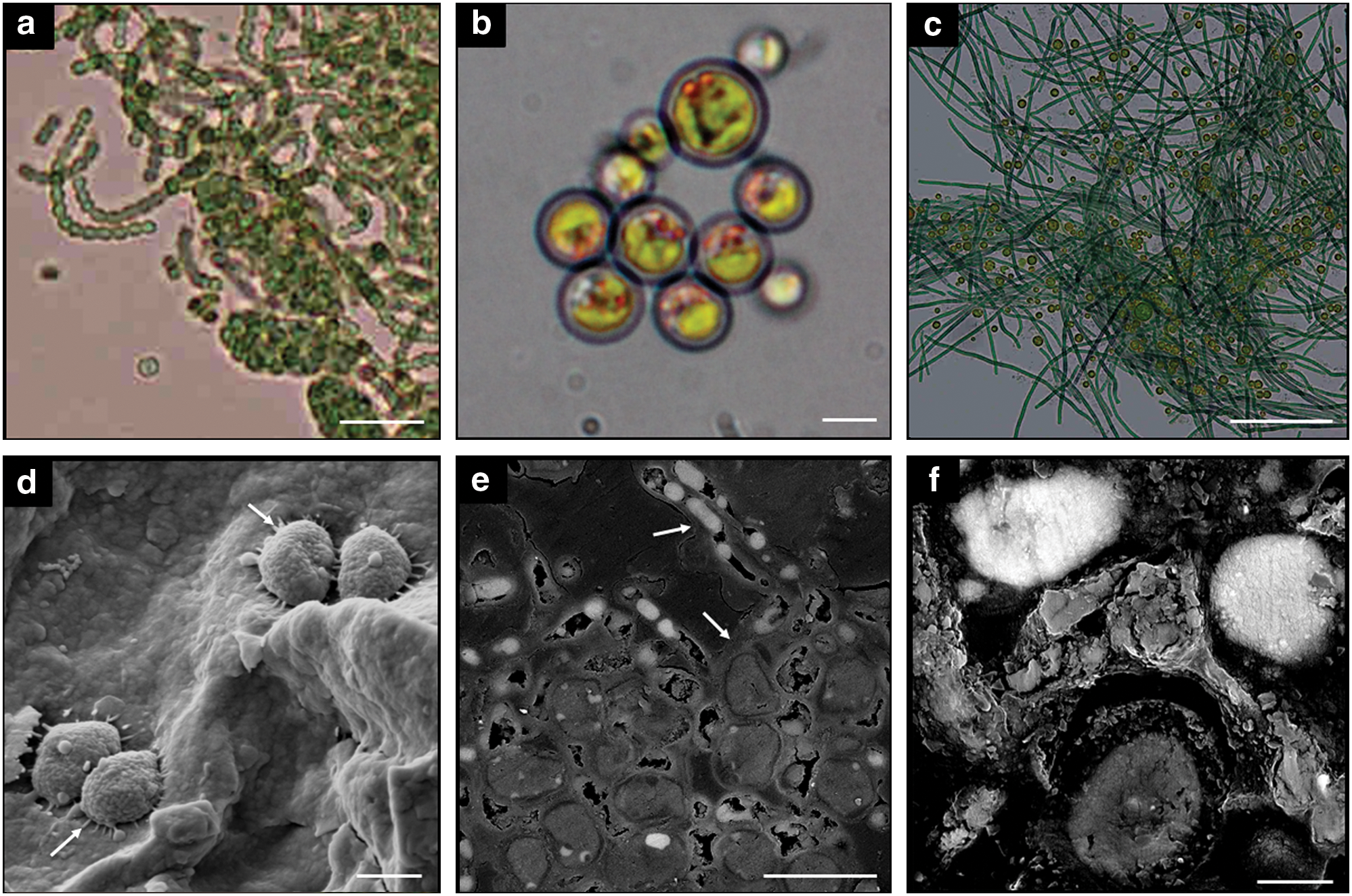
Micrographs of cultured isolates from shocked gneisses (upper panels) and SEM micrographs of in situ colonization within highly shocked gneisses (lower panels). (
Scanning electron microscopy imaging of field samples revealed extensive endolithic colonization, associated with pore spaces within the rocks. Microbes were seen either directly bound to the substrate (Fig. 2d), as a massive colony contained within exopolymeric substance (EPS) (Fig. 2e), or fossilized within calcite (Fig. 2f), a prevalent depositional material within the Haughton impact structure (Osinski et al., 2005). See Supplementary Figs. S1 and S2 for EDX data.
3.2. 16S rRNA analysis and diversity calculations
A total of 2,795 OTUs were generated from 44,534 sequences, with a mean length of 244 nucleobases and a Good's coverage > 90%. Analyses of the total bacterial 16S rRNA showed marked statistical differences between the unshocked and shocked populations, revealing some general trends. A total of 45% of OTUs belonged to the high shock community, 36% to the moderate shock community, and 18% to the unshocked community (see Supplementary Fig. S3 for a Venn diagram of un-subsampled sequences). The unshocked samples consisted primarily of three classes of bacteria: Actinobacteria (40%), Gammaproteobacteria (29%), and Alphaproteobacteria (21%), with a small contribution from the Clostridia and the Bacilli. Among the different shock levels, Actinobacter and Fusobacteria classes were ubiquitous throughout, with the Actinobacteria having the largest population among all communities (as high as 65%). A small number of OTUs (65) were shared between all three communities, and in general, species that were largely present in the unshocked communities were poorly represented within the shocked samples and vice versa. For example, with the exception of a few OTUs, both the Alphaproteobacteria and Gammaproteobacteria decrease by almost an order of magnitude with increasing shock (i.e., 21% and 29%, down to 5% and 2.5%, respectively), whereas the Betaproteobacteria are largely present only within the shocked samples (Fig. 3a). Algal and fungal sequencing was completed for only three samples (one from each shock community) to serve as an indicator of species present (Fig. 3b, 3c). Within the algae, only the moderate and high shock samples could be amplified (BE009A13 and AH003A, respectively), and both low and moderate shock samples failed to generate amplicons for fungal analysis. The moderately shocked sample analyzed for algae was composed entirely by an OTU belonging to the Chlorophyta, most closely affiliated (97%) with Rosenvingiella radicans. The algal community in the highly shocked sample showed more taxonomic diversity, consisting of 9 OTUs within the Chlorophyta (though still dominated by R. radicans) and 1 of Streptophyta: Klebsormidium flaccidum, a filamentous green algae. The fungal OTUs (Fig. 3b) from sample AH003A were dominated by the Ascomycota, consisting of sequences most closely affiliated with Thelidium decipiens and Placidiopsis cinerascens, and the Microsporidia (Systenostrema alba), a parasitic fungus of animals, with a small contribution from the Basidiomycota in the form of mushroom-forming spores such as Cortinarius sodagnitus. Focusing solely on the bacterial phototrophic community, the largest contributor within the rocks came from the Chloroflexi (class), which represented 2% of sequence reads in the unshocked lithologies, increasing to 16% in the high shock samples (Fig. 4). The dominant order at all shock levels was Chloroflexalus, with small contributions from the Caldilineales, a strictly anaerobic order (Yamada et al., 2006). The Cyanobacteria were not largely present (maximum of 6% of sequences in the highly shocked samples, though counts did increase by an order of magnitude between each of the unshocked, moderately shocked, and highly shocked samples), having a minor contribution from the Chroococcales and Oscillatoriales, the latter of which was only present within the shocked lithologies.

(

Percentage abundance of the phototrophic bacterial communities from the unshocked, moderately shocked, and highly shocked gneiss pools, presented at the class level as a function of the percentage of the total bacterial community. Chloroflexi classes have been grouped for the sake of clarity, as all Chloroflexi classes from the sequence reads (other than the class Chloroflexi) had only a small contribution. (Color graphics available at
Alpha diversity analyses using the inverse Simpson index (D), Shannon-Weiner index (H), and Chao index all showed an increase both in species richness and diversity with increasing shock (see Supplementary Table S3 for confidence intervals). H increased from 2.4 to 4.6, and the inverse Simpson gave a diversity coefficient of 7.4 in unshocked samples, to 29.6 in moderate shock and 48.6 for the high shock community (Table 1). An AMOVA was conducted to test whether the centers of the clouds for each population were statistically separated. All comparisons, except for that of high shock–moderate shock revealed a statistically separate population, where high shock versus unshocked had a p value of 0.01, moderate shock versus unshocked had p < 0.005, and a three-way comparison of all communities yielded a p value of 0.002. The NMDS plot was conducted using three dimensions in order to reduce stress on the system and had a final stress value of 0.24 and an R 2 of 0.46. The plot (Fig. 5) only showed clear grouping between the unshocked and shocked populations, with the moderately shocked group clustering within the highly shocked group, though having its own distinct center. Samples of the same shock level group, but from different years and different storage conditions, yielded comparable numbers of sequence reads, while also maintaining similar levels of observed species diversity. For example, within the unshocked sample group, the average number of sequence reads was 2395 for samples collected in 2000 (D = 8.84) versus 2465 for samples collected in 2010 (D = 5.2), a difference of 2.9%, indicating that little to no sample degradation occurred.

Nonmetric multidimensional scaling (NMDS) plot for bacterial 16S rRNA from unshocked, moderately shocked, and highly shocked populations (AMOVA, n = 20, p < 0.01 for division in a pairwise comparison between unshocked and highly shocked populations, and p < 0.005 for pairwise between unshocked and moderate shock). High shock samples do appear to show some grouping distinct from the unshocked communities but plot very closely with moderate shock samples, though the centers are different. (Color graphics available at
Standard deviations are provided for all diversity indices.
Alpha indices calculated only for the bacterial 16S rRNA genes.
3.3. Porosity measurements
Porosity measurements for this study represent a subset of a much larger data set and follow a clear exponential trend with increasing shock level (see Pontefract et al., 2014). Values for the crystalline lithologies typically range from between 0% (for unshocked) and up to 70% for highly shocked samples. For the samples used in this study (see Supplementary Tables S1 and S2), unshocked lithology porosities range from 0% to 4% of rock volume, whereas the high shock samples range from 34% to 62% porosity. Porosity values typically show a high degree of variance within each shock level demarcation (e.g., Singleton et al., 2011), and a Pearson correlation analysis of porosity with diversity (inverse Simpson) shows a high degree of correlation, R 2 = 0.79. When plotted against each other, a simple linear regression gives an R 2 of 0.62 (Fig. 6).

(
4. Discussion
This study represents the first comprehensive assessment of a microbial community within impact-generated lithologies and represents one of the few culture-independent studies of land-based high-Arctic microbial communities (e.g., Nemergut et al., 2005; Wilhelm et al., 2011; Deslippe et al., 2012; Ziolkowski et al., 2013; Rhind et al., 2014).
Similar to Arctic permafrost communities, the Actinobacteria were the largest contributors to the endolithic community at all three shock levels, though very few OTUs were shared between groups; only 1.2% of Actinobacteria sequences were common to all three shock levels, generally belonging to the order Actinomycetales (77%) and Acidimicrobiales (23%). The unshocked lithologies contained a community structure close to that of an Arctic soil study based in Alaska (Deslippe et al., 2012), having abundant Proteobacteria (specifically within the Alphaproteobacteria and Gammaproteobacteria—both of which contain N2-fixing species) and low (<3%) levels of the Firmicutes, Bacteroidetes, and Chloroflexi. Interestingly, the Gemmatimonadetes (Gram-negative aerobic bacteria), which are generally quite prevalent in Arctic soils, were found only within the high shock level samples. The higher shocked samples had microbial compositions similar to that of Fike et al. (2003) and Pellerin et al. (2009)—though their study did not identify any phototrophic contribution—and the permafrost layer communities described by Steven et al. (2007) and Wilhelm et al. (2011). Within endolithic communities, it has been suggested that nitrogen fixation is generally rare (Friedmann, 1982). We do not find this to be the case in our study, although the N2-fixing species, such as Rhizobium, were found only within sequence reads from shocked lithologies and were also accompanied by an increase in the phototrophic communities, several species of which are diazatrophs. Presumably, some nitrogen input for the community was also likely provided through atmospheric fixation and deposition (Walker and Pace, 2007).
4.1. The phototrophic community
The most profound effects of shock on the microbial community were seen within the phototrophic organisms, spanning four phyla within the bacteria. The Chloroflexi (green nonsulfur bacteria) were the dominant phototroph in each shock level (2%, 9%, and 16%, respectively), over the Cyanophyta, which, even in the high shock level samples, only reached an abundance of 6%. In addition, there was a very small addition to the phototrophic community in the Rhodospirillaceae, a photoorganoheterotroph also from the green nonsulfur bacteria. Two previous studies mention the presence of Chloroflexi within soils or endolithic communities (Deslippe et al., 2012; Rhind et al., 2014). This is in contrast, however, to other literature on Antarctic and Arctic endolithic communities that have consistently found cyanobacteria to be the dominant phylotype—up to 56% (e.g., de la Torre et al., 2003; Pointing et al., 2009). In a study by Ziolkowski et al. (2013), who examined microbial community composition in gypsum crusts on Axel Heiberg, the same primer sets were used as in the present study, yet cyanobacteria were found to be the dominant organism in the system. This study shows that, even if bias exists, this primer set can satisfactorily resolve cyanobacterial taxa within samples. A few studies have indicated anomalously high numbers of Chloroflexi sequences in their data sets (e.g., Costello and Schmidt, 2006; Wilhelm et al., 2011), but the reason behind the occurrence of such high abundances in these locations remains unclear. Within the shocked gneiss community, the cyanobacteria present consisted of Chroococcales, Nostocales, Oscillatoriales, few of which were cultured in the lab. Cyanobacteria in endolithic communities from Antarctica appear to be much more diverse, containing 112 distinct species, many of which are unknown (Pointing et al., 2009), compared to 62 found in the shocked gneisses. This is unusual considering the fact that the Arctic environment is considerably less extreme than that of the Antarctic, receiving much larger amounts of precipitation throughout the year and experiencing warmer temperatures, resulting in less stringent selective pressures (Omelon et al., 2006). In addition, the sandstones of Antarctica have a porosity of between 2.3% and 13.0% (Cockell et al., 2003), compared to the shocked gneisses, which can reach porosities of over 60% (Singleton et al., 2011). It is clear, however, that the level of porosity does affect the colonization of shocked gneisses by phototrophs (both Cyanobacteria and Chloroflexi), or it is a main factor, evidenced both by the marked increase in bacterial diversity and species richness with increasing shock (Table 1) and by the presence of eukaryotic algae and large filamentous cyanobacteria, such as Oscillatoriales. In this instance, the increased porosity would allow for up to 100% transmission of light in some areas (Pontefract et al., 2014) and provide sufficient space previously unavailable for such relatively large organisms.
Neither the algal nor fungal species within these samples appeared to be strong contributors to the overall community, existing at very low abundances, which is in keeping with previous microscopic observations by Cockell et al. (2003) and Omelon et al. (2006), though work on a larger sample size must be completed. As a whole, heterotrophic bacteria dominated the community, with the phototrophs supplying, at most, 12% of the sequence reads. This suggests that the organisms must, in addition to using carbon sourced from the primary producers, rely on an exogenous form of carbon input from the environment, likely along with other nutrients such as phosphorus and nitrogen. Without knowing the composition of the active community, it is difficult to surmise the metabolic diversity of these Arctic endolithic communities, and an investigation of the active community should be completed. The presence of so many bacterial species in such a harsh environment, as well as a plausible reliance on external sources of nutrients, indicates that few species might occupy metabolic niches, with the remainder of the population potentially using similar metabolic pathways (Fierer and Lennon, 2011). The dominant presence of Actinobacteria within all the samples does show that a significant amount of carbon turnover is possible within these communities, made more likely by the wetter conditions experienced in the Arctic.
4.2. Impact-generated diversity
Overall, a significant change in microbial community composition with increasing shock pressure in the gneisses was observed, revealing the presence of distinct populations, likely based on the level of created porosity and light transmission within the rock. AMOVA was able to show that there is a statistical difference in community composition between the unshocked and shocked populations; however, due to the high variability inherent in the samples, the moderately and highly shocked communities showed no significant difference from each other. This is likewise reflected in the NMDS plot (Fig. 5), which, aside from showing a more distinct difference between unshocked and shocked populations, is not able to visually resolve any further patterns related to host-rock shock metamorphism. The high stress level (0.24) also indicates that the 3-D plot is not an optimum representation of the data. That porosity is highly variable within each shock level and that it correlates with species diversity accounts for a large amount of the variation seen in the NMDS plot, as well as in the standard deviations for the diversity indices in the high shock level communities. Other factors that affect diversity may have to do with placement along flat-lying or inclined areas within the crater, which would influence drainage, the amount of sun exposure experienced, and the amount of airborne material received. Geographical location did not appear to be a determining factor, as some samples collected from opposite regions within the crater plot together, while samples collected near each other along the same escarpment do not. Future work in this area requires a detailed ecological study of the crater region so as to determine the effects of variable light incidence and precipitation on shocked lithologies and how these variables may interact to influence community composition.
4.3. Astrobiological implications
A wide range of studies spanning polar and nonpolar desert endolithic communities has shown that such populations are highly specialized and, in many cases, ideally adapted to surviving such harsh conditions (Friedmann et al., 1993; de la Torre et al., 2003; Wierzchos et al., 2006; Walker and Pace, 2007; Omelon, 2008). As a result of this, these organisms tend to be remarkably persistent over long periods of geological time (Pointing et al., 2009). What does this mean for potential life elsewhere in our solar system? On Mars for example—which is essentially a polar desert, albeit an extreme one—impact cratering and the generation of shocked lithologies as well as transient hydrothermal systems (Osinski et al., 2013) is an important geological process. If life existed on Mars in the early Noachian, it is plausible that, as conditions began to deteriorate and Mars transitioned into a cold, dry planet, life may have survived in the abundant endolithic environments afforded by impact craters. Early work by Newsom (1980) posited the presence of widespread deposits of impact melt on Mars, which was confirmed in a recent study by Cannon and Mustard (2015), in which extensive deposits of glass-rich impactites were revealed. Not only do surficial brecciated deposits provide attractive targets for life-detection missions, but endolithic environments could likely have formed deep within the fractured subsurface of these craters (Boston et al., 1992; Ascaso and Wierzchos, 2002; Cockell et al., 2012). Future infilling of these deposits may then have preserved these organisms, making these brecciated, impact-melt lithologies an attractive target for future life-detection missions as well as for Mars sample return.
5. Concluding Remarks
This culture-independent study improves upon previous culture work on impact-generated lithologies and provides a clearer understanding of how microbial populations scale within different shock regimes. We found a dominant heterotrophic community at all shock levels, with a relatively small phototrophic community, largely dominated by the green nonsulfur bacteria. Altogether, the impact-created endolithic environment is a beneficial habitat within polar deserts, which has important implications for the potential for viable habitats on other terrestrial bodies in our solar system—especially in instances where impact cratering may be one of the only active geological processes.
Footnotes
Acknowledgments
This work was funded by the NSERC Discovery Grant, the Canadian Astrobiology Training Program and the Northern Scientific Training Program, with logistical support provided by the Polar Continental Shelf Program. Thank you to Pat Schloss, Jacqueline Goordial, Jen Ronholm, Chris Carr, Zoe Lindo and the three anonymous reviewers for their valuable commentary and contributions to this work. Project sequencing data sets have been deposited under the NCBI Sequence Read Archive (SRA) under project PRJNA308985.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.