
Abstract
Fluid inclusions trapped in ancient halite can contain a community of halophilic prokaryotes and eukaryotes that inhabited the surface brines from which the halite formed. Long-term survival of bacteria and archaea and preservation of DNA have been reported from halite, but little is known about the distribution of microbes in buried evaporites. Here we report the discovery of prokaryotes and single-celled algae in fluid inclusions in Pleistocene halite, up to 2.26 Ma in age, from the Qaidam Basin, China. We show that water activity (a w), a measure of water availability and an environmental control on biological habitability in surface brines, is also related to microbe entrapment in fluid inclusions. The a w of Qaidam Basin brines progressively decreased over the last ∼1 million years, driven by aridification of the Asian interior, which led to decreased precipitation and water inflow and heightened evaporation rates. These changes in water balance produced highly concentrated brines, which reduced the habitability of surface lakes and decreased the number of microbes trapped in halite. By 0.13 Ma, the a w of surface brines approached the limits tolerated by halophilic prokaryotes and algae. These results show the response of microbial ecosystems to climate change in an extreme environment, which will guide future studies exploring deep life on Earth and elsewhere in the Solar System. Key Words: Halite fluid inclusions—Ancient microbes—Water activity—Qaidam Basin—Pleistocene aridification. Astrobiology 16, 379–388.
1. Introduction
B
Earlier studies showed that the distribution of halophilic prokaryotes and single-celled algae in ancient halite is controlled by the habitability of the surface environments in which the halite formed (Schubert et al., 2009a, 2010b; Lowenstein et al., 2011). Microorganisms occur in fluid inclusions in Pleistocene halite from Death Valley and Saline Valley, California, that originally crystallized in perennial hypersaline lakes, but not in halite formed in drier, ephemeral saline pan environments (Schubert et al., 2009a, 2009b; Lowenstein et al., 2011). Here we document prokaryotes and single-celled Dunaliella-like algae in older, Pleistocene halite, up to 2.26 Ma in age, from the Qaidam Basin, China, and show that a second environmental factor, water activity (a w), is critical for the entrapment and preservation of microorganisms in fluid inclusions. The distribution of microorganisms in subsurface halite from the Qaidam Basin shows a strong correlation with the chemical composition of paleobrines and a w, the latter a measure of water availability and salinity. Brine chemistry in the Qaidam Basin was, in turn, controlled by progressive aridification of the Asian interior during the Pleistocene.
2. Geological Background
The Qaidam Basin is an intermontane basin (120,000 km2) on the northeastern Tibetan Plateau, covered by dry salt pans and sparse hypersaline lakes (Kezao and Bowler, 1986). It is part of the arid Asian interior, which extends from the Chinese Loess Plateau in eastern Asia to the Caspian Sea in central Asia (Fig. 1). The Asian interior has undergone long-term aridification for >20 million years, linked to uplift of the Tibetan Plateau and Northern Hemisphere glaciation (An et al., 2001; Guo et al., 2002; Ding et al., 2005). Evaporites have accumulated in the Qaidam Basin for ∼4 million years (Wang et al., 2013). A 939 m long salt core (SG-1) through the Chahansilatu salt flat, western Qaidam Basin, is composed of bedded halite and mud with gypsum (CaSO4·2H2O), glauberite (Na2SO4·CaSO4), thenardite (Na2SO4), bloedite (Na2SO4·MgSO4·4H2O), and polyhalite (K2SO4·MgSO4·2CaSO4·2H2O) (Fig. 1). Paleomagnetism and spectral analysis of magnetic susceptibility indicate deposition from 2.69 to 0.08 Ma (Zhang et al., 2012; Herb et al., 2015).
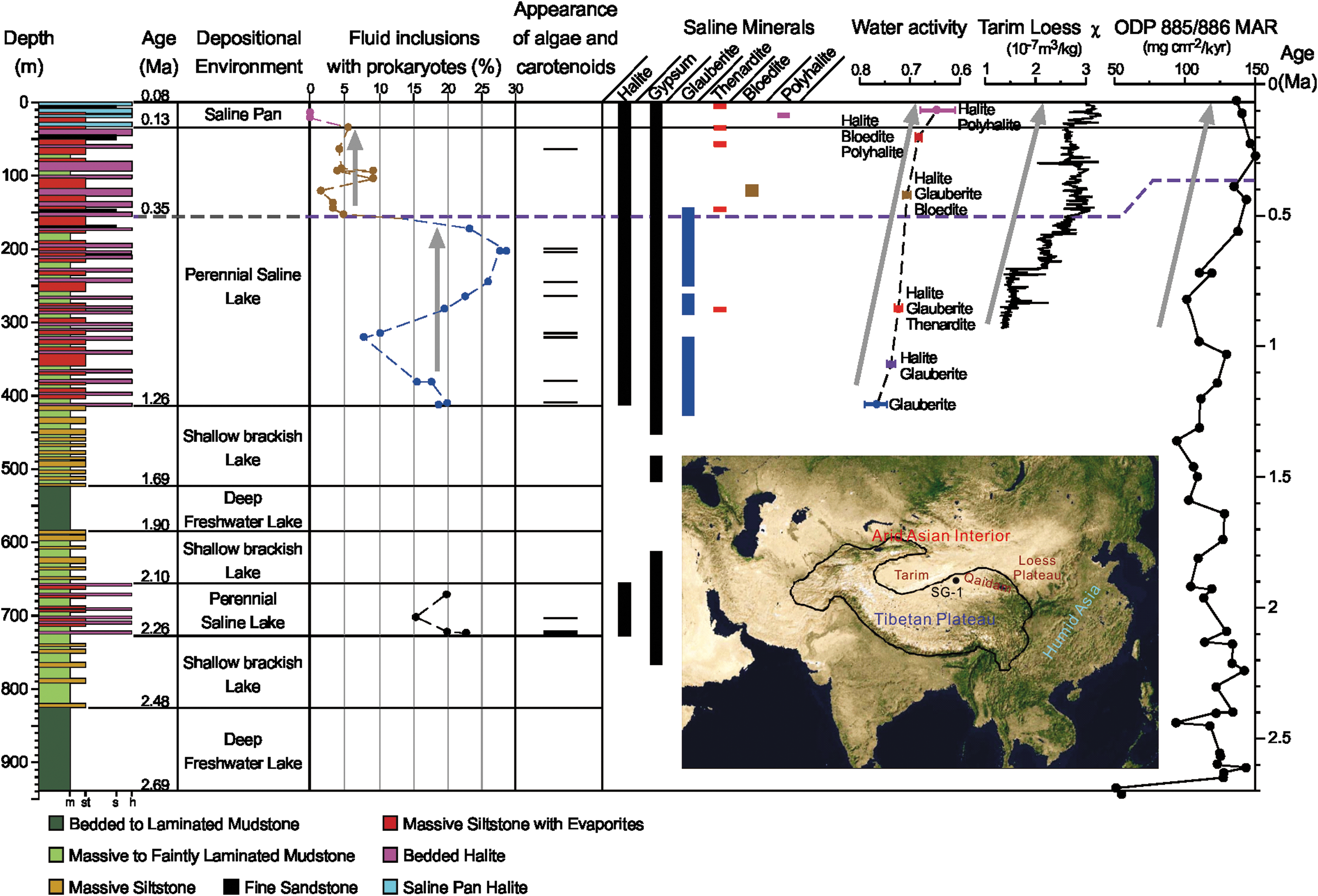
Stratigraphy, ages calculated from orbitally tuned analysis (5 kyr resolution) of magnetic susceptibility (Herb et al., 2015), and depositional environments (Wang et al., 2012), core SG-1, Qaidam Basin, China; m = mudstone, st = siltstone, s = sandstone, h = halite. Distribution of prokaryotes, algae, and carotenoids in core SG-1 obtained from microscopic analysis (Table 1). Vertical gray arrows show large number of prokaryotes in fluid inclusions (1.26–0.38 Ma) and then generally lower numbers of prokaryotes (0.35–0.13 Ma). Inset map shows arid Asian interior, Tibetan Plateau, Qaidam Basin, and location of core SG-1. Saline mineral distribution modified from Li et al. (2010); halite—NaCl; gypsum—CaSO4·2H2O; glauberite—Na2SO4·CaSO4; thenardite—Na2SO4; bloedite—Na2SO4·MgSO4·4H2O; polyhalite—K2SO4·MgSO4·2CaSO4·2H2O. Water activity was calculated from mineral assemblages in core SG-1 and evaporation simulations of groundwater brines from Chahansilatu salt flat (Table 2), using the computer program EQL/EVP. Gray arrow shows progressive decrease in water activity (increase in brine concentration) from ∼1.26 to 0.08 Ma. Loess magnetic susceptibility from sediment core in nearby Tarim Basin shows stepwise drying of central Asia over the past ∼0.9 million years (gray arrow) (Zan et al., 2010). MAR (mass accumulation rate) of eolian dust calculated from ODP 885/886 core, North Pacific Ocean (Rea et al., 1998), shows increasing dust over the last ∼0.8 million years (gray arrow). This dust, derived from the Asian interior, is an indicator of aridification there.
Paleoenvironments at Chahansilatu salt flat, interpreted from core SG-1, indicate progressive drying of the western Qaidam Basin over the last 2.7 million years (Wang et al., 2012). Bedded and finely laminated detrital mudstones, 2.69–2.48 and 1.90–1.69 Ma in age, are diagnostic of accumulation in deep freshwater lakes (Fig. 1). Mudstones, siltstones, and fine sandstones, 2.48–2.26, 2.10–1.90, and 1.69–1.26 Ma in age, deposited in shallow brackish lakes, contain plant relicts, burrows, sand lenses, wave ripples, and displacive gypsum (Wang et al., 2012). Bedded halite with crystalline growth textures, 2.26–2.10 and 1.26–0.13 Ma in age, formed in perennial hypersaline lakes (Fig. 1). The youngest halites, 0.13–0.08 Ma in age, contain dissolution features diagnostic of deposition in saline pans and ephemeral saline lakes, similar to the modern desiccated Qaidam Basin (Fig. 1).
3. Materials and Methods
3.1. Halite and microorganism microscopy
Large-format polished thin sections (5 × 7.5 cm) of Qaidam Basin halite were examined with a Leica Wild M3Z stereomicroscope. Plan view photographs of thin sections were taken with a Nikon D80 camera. Halophilic microorganisms were identified in fluid inclusions using a Zeiss compound microscope with an oil immersion objective (Plan Apo 100×/1.4 oil). Fluorescent characteristics of halophiles were studied using an HBO 100 mercury lamp and Chroma Technology Corp. filters [DAPI: exciter D350/50 nm, emitter D460/50 nm, beamsplitter 400dclp; green fluorescent protein (GFP): exciter HQ450/50 nm, emitter HQ510/50 nm, beamsplitter Q480lp; and TRITC: exciter HQ545/30 nm, emitter HQ620/60 nm, beamsplitter Q570lp].
In situ direct counts of microorganisms trapped in fluid inclusions from halite were used to evaluate the distribution of halophilic microorganisms in core SG-1 (Fig. 1). Percentages of fluid inclusions with prokaryotes were determined microscopically using a 100× oil immersion objective. Five thousand one hundred fluid inclusions in thin sections of halite from 28 stratigraphic intervals were examined for prokaryotes and algae along random transects.
3.2. Modern brine evaporation simulation
The EQL/EVP computer program (Risacher and Clement, 2001) was used to simulate the saline mineral sequence produced during the evaporation of brine under equilibrium conditions. Simulations were carried out at 298 K, 1 atm pressure, and CO2 partial pressure of 10−3.4, using a brine with the composition of groundwater from the core SG-1 drill site. At each evaporation step, the chemical composition of the brine and the masses of precipitated minerals were calculated using the Pitzer ion interaction model and the Newton Raphson method for solving the set of mass balance and mass action equations describing the system (Risacher and Clement, 2001). Simulations were run under open conditions, which remove minerals from the system as they precipitate (Risacher and Clement, 2001). The activity of water (a w), a function of the concentration of all the components, was calculated at each evaporation step (Risacher and Clement, 2001). Comparisons between brine chemistries calculated using EQL/EVP and the chemical composition of natural waters show that the computer program accurately simulates brine evolution in natural systems (Risacher and Clement, 2001; Ma et al., 2010).
The mineral sequence produced during progressive evaporation of brines from the Qaidam Basin, SG-1 drill site, Chahansilatu salt flat, was glauberite, halite + glauberite, halite + glauberite + thenardite, halite + glauberite + bloedite, halite + bloedite + polyhalite, and halite + polyhalite, which matches the vertical sequence of salts observed in core SG-1 (Fig. 1). These saline mineral assemblages define specific water activities that decrease during each step of progressive evaporation. In order to reproduce the observed saline mineral sequence found in core SG-1, the chemical composition of modern brine from the Chahansilatu salt flat was modified slightly (addition of 100 mmol per liter SO4 2− and removal of 100 mmol per liter Mg2+).
4. Results
4.1. Depositional environments of halite
Diagnostic sedimentary textures and fabrics of halite in core SG-1, Chahansilatu salt flat, were used to interpret depositional environments in the Qaidam Basin over the past 2.26 million years. Three types of halite occur in core SG-1: (1) Chevron halite (Fig. 2a) consists of crystals with dense fluid inclusions in the cores and outer zones containing few fluid inclusions. Chevrons form by rapid crystal growth, and clear rims form during later slow crystallization at the brine bottom (Arthurton, 1973; Lowenstein and Hardie, 1985). These textures are typical of shallow saline lake deposits (Wardlaw and Schwerdtner, 1966; Lowenstein and Hardie, 1985). (2) Clear bottom-growth halite (Fig. 2b) contains clear crystals, with sparse fluid inclusions and no chevron bands, suggesting slow growth of halite at the bottom of a perennial saline lake. (3) Saline pan halite (Fig. 2c) is rare and only occurs above depths of 30 m. It contains vertical dissolution pipes formed during surface flooding by dilute waters; pipes are later filled with clear halite cements.

Thin section photomicrographs of halite, core SG-1, Qaidam basin. (
Depositional environments in which halite was deposited in core SG-1 were interpreted from petrographic observations combined with previous lithofacies analysis (Wang et al., 2012) (Fig. 2): (1) 2.26–2.10 Ma, predominantly clear bottom-growth halite crystallized on the floor of a perennial hypersaline lake (Fig. 2b). (2) 1.26–0.13 Ma, composed of clear bottom-growth halite, chevron halite, and cumulates (small halite crystals precipitated at the air-brine interface), deposited in a perennial hypersaline lake (Fig. 2a). (3) 0.13–0.08 Ma, contains saline pan halite (Fig. 2c), which marks the desiccation of earlier, long-lived perennial saline lakes.
4.2. Identification of prokaryotes and algae
Bedded halite from core SG-1 contains arrays of fluid inclusions oriented in bands parallel to crystal growth faces, named chevrons (Fig. 3a). Larger, less densely spaced fluid inclusions in crystals (clear bottom-growth halite) (Fig. 3b) contain coccoid-shaped prokaryote cells less than 1 μm in size (Fig. 3c). These cells commonly autofluoresce when using epifluorescence microscopy (Fig. 3d–3f). Prokaryote cells display random zigzag Brownian motion in fluid inclusions (Supplementary Movie 1; Supplementary Material is available online at

Photomicrographs of prokaryotes in fluid inclusions in ancient halite, core SG-1. (
A second type of microorganism, 10–20 μm in diameter, with spherical or ellipsoidal shape and green, orange, and red color, occurs in close association with smaller prokaryotes in fluid inclusions (Fig. 4a–4b). Cells are similar in size and structure to the green algae Dunaliella, an important photosynthetic autotroph in modern hypersaline environments (Oren, 2005). Many cells are coated with red-brown or orange crystalline material (Fig. 4a–4b) that is birefringent under cross-polarized light (Fig. 4c) and has been identified as carotenoid by laser Raman spectroscopy (Winters et al., 2013). Carotenoids are produced by modern Dunaliella cells in response to elevated salinity, high light intensity, or low concentrations of nutrients (Ben-Amotz et al., 1982; Loeblich, 1982; Borowitzka and Borowitzka, 1989). Green and orange cells autofluoresce brightly using epifluorescence microscopy (Fig. 4d), suggesting they may also contain chlorophyll.
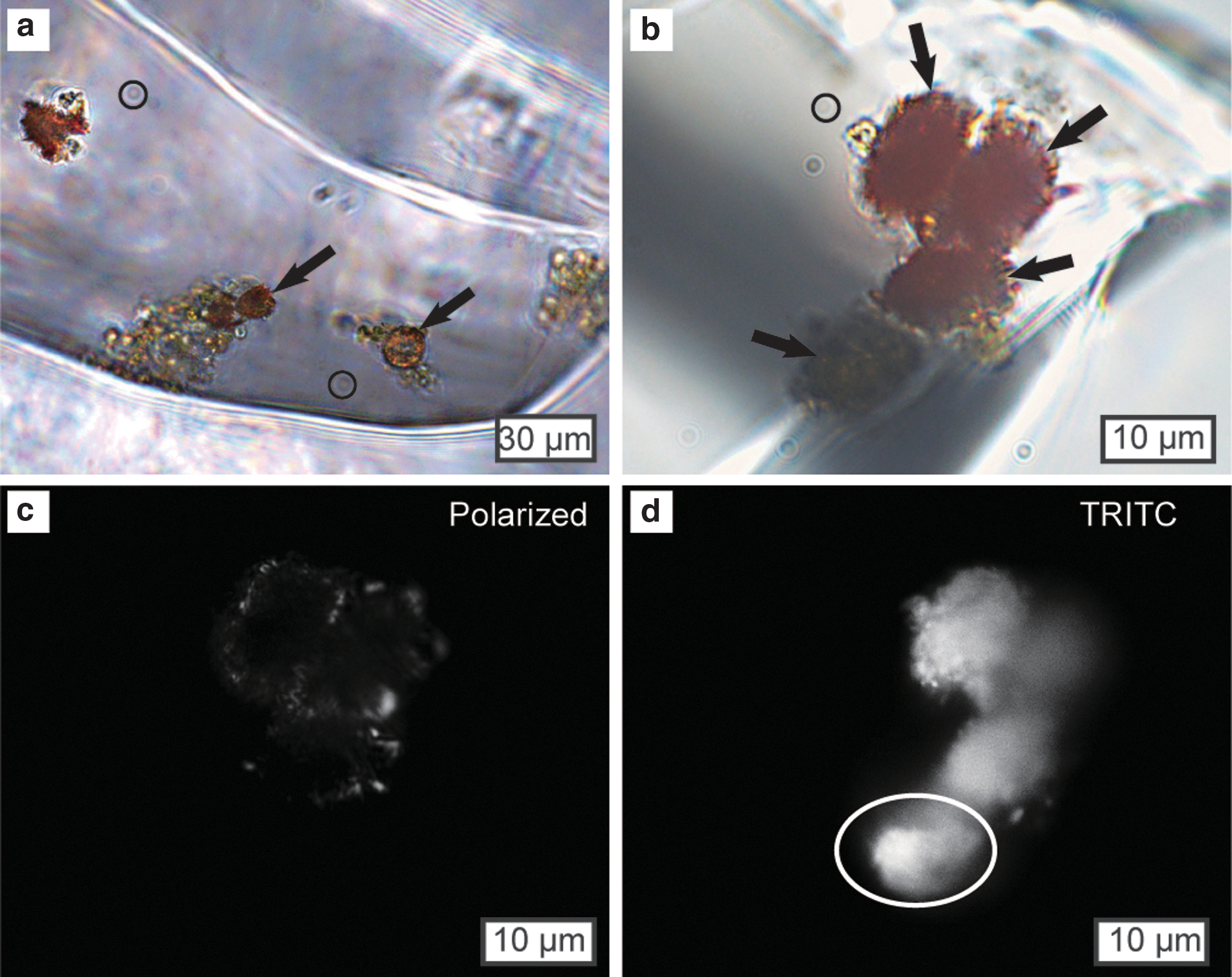
Photomicrographs of single-celled algae and carotenoids in fluid inclusions in halite from core SG-1. (
4.3. Distribution of prokaryotes and algae
Direct counts of prokaryotes trapped in more than 5000 fluid inclusions from 28 halite intervals were used to evaluate the distribution of halophilic microorganisms in core SG-1 (Fig. 1 and Table 1). Nearly all prokaryotes occur in fluid inclusions from “clear” (inclusion-poor) bottom-growth halite, interpreted to have crystallized slowly on the floor of perennial hypersaline lakes in the Qaidam Basin (Fig. 3b). Slow crystallization allowed prokaryotes living in the water column to become trapped within halite crystals at the brine bottom (Schubert et al., 2009b).
Relatively large percentages of fluid inclusions in halite (15–23%) from the lower perennial saline lake interval (723–671 m, 2.26–2.1 Ma) contain prokaryotes (Fig. 1 and Table 1). Similarly abundant prokaryotes occur in fluid inclusions from the thick upper perennial saline lake halite (8–29% in 12 intervals, 413.6–171.0 m, 1.26–0.38 Ma). The occurrence of algal cells in fluid inclusions parallels the distribution of prokaryotes: before 0.38 Ma (below 171 m), nearly all halite intervals contain Dunaliella-like algal cells. Abundant microorganisms in fluid inclusions from clear bottom-growth halite indicate that biologically productive saline lakes, with large numbers of halophilic prokaryotes and Dunaliella, existed in the Qaidam Basin between 2.26 and 0.38 Ma.
There is a sharp drop in the number of prokaryote cells in halite from the upper perennial saline lake interval, above 154 m (0.35 Ma) (Fig. 1 and Table 1). In that interval (154–30.8 m, 0.35–0.13 Ma), the percentage of fluid inclusions with prokaryotes varies from 1.5% to 9.2% and averages ∼5%. Fluid inclusions from only one halite sample in that interval (62.3 m, 0.18 Ma) contain algal cells. These data indicate a modification in the microbial community at 0.35 Ma, changing from earlier saline lakes in which microorganisms thrived (413.6–171.0 m, 1.26–0.38 Ma) to later, less biologically productive saline lakes (154–30.8 m, 0.35–0.13 Ma). The thick section of halite with relatively few microorganisms (154–30.8 m) suggests that although the depositional environment (perennial saline lake) was favorable for microbial activity (Schubert et al., 2009a, 2009b; Lowenstein et al., 2011), some other factor limited the productivity of the halophilic microbial community.
No halophilic microorganisms were identified in fluid inclusions in saline pan halite from the top 31 m of core SG-1 (0.13–0.08 Ma) (Fig. 1 and Table 1). This is consistent with previous observations that showed that halite from modern and ancient saline pans contained relatively few microorganisms in fluid inclusions (Schubert et al., 2009a). Saline pans are characterized by shallow, ephemeral lakes that are too short-lived for robust halophile communities to develop, or rapid crystallization of halite during brief saline lake periods may not have coincided with periods of halophilic microbial activity (Schubert et al., 2009a).
4.4. Modern brine simulation
Dissolved salts present special challenges to life in hypersaline environments because organisms need mechanisms to prevent the loss of water by osmosis (Grant, 2004). Microorganisms have evolved to accumulate KCl or to produce organic compatible solutes, such as glycerol, both of which can counterbalance the osmotic stresses of hypersaline waters (Grant, 2004). Water availability in hypersaline environments can be expressed by salinity, or by the thermodynamic parameter, activity of water, a
w. The activity of a constituent, i, in a solution is given by the ratio of fugacity, fi
, in a given state, to the fugacity, fi
o, in some standard state at the same temperature (Klotz and Rosenberg, 1974):
At 1 atm total pressure, water vapor may be considered an ideal gas, so the fugacities may be replaced by the partial pressure of H2O, and
where the standard state is pure H2O at 1 atm and the temperature of interest (Klotz and Rosenberg, 1974). Any aqueous solution with a finite amount of dissolved constituents has a w less than 1.
Halophilic archaea and bacteria can grow at a w < 0.75 and in extreme cases a w as low as 0.635 (Stevenson et al., 2015). Halophilic algae, such as Dunaliella, can grow at a w < 0.75, but not < 0.70 (Stevenson et al., 2015). Salinities in natural waters can approach and exceed the water activity limits tolerated by halophilic archaea, bacteria, and eukaryotes.
The activity of water in Qaidam Basin brines over the past 2.26 million years was calculated from the assemblage of saline minerals in core SG-1 and evaporation simulations of modern subsurface brines from Chahansilatu salt flat using the computer program EQL/EVP (Risacher and Clement, 2001). Progressive evaporative concentration of Chahansilatu brine produces the mineral sequence glauberite (a w = 0.79–0.74), halite + glauberite (a w = 0.74–0.73), halite + glauberite + thenardite (a w = 0.73–0.72), halite + glauberite + bloedite (a w = 0.72–0.71), halite + bloedite + polyhalite (a w = 0.69–0.68), and finally halite + polyhalite (a w = 0.68–0.61) (Table 2). This mineral sequence matches that observed in core SG-1, which allows calculation of a w in Qaidam Basin paleobrines (Fig. 1). Halite, gypsum, and glauberite occur at depths 723–154 m (2.26–0.35 Ma), which defines a w > 0.73 and coincides with relatively abundant halophilic prokaryotes and algal cells in fluid inclusions in halite formed in perennial hypersaline lakes before 0.35 Ma (Fig. 1). Halite and gypsum plus thenardite, then bloedite, and finally polyhalite, occur successively at depths above 154 m (0.35 Ma), which indicates that a w progressively decreased from 0.73 to as low as 0.61 over this interval (Fig. 1 and Table 2). Over the same interval, the number of prokaryote cells in halite drops sharply (upper perennial saline lake interval of Fig. 1). The upper saline pan interval, with polyhalite and a w = 0.68–0.61, is completely devoid of halophilic microorganisms (Fig. 1). There is thus a strong correlation between decreasing water activities in surface brines in the Qaidam Basin paleolakes and low microbial abundances in fluid inclusions in halite. The lack of preserved halophilic prokaryotes and algae in fluid inclusions in halite from above ∼30 m (0.13 Ma) suggests that water activity was < ∼0.70 (halite + bloedite and polyhalite) and approached the limit tolerated by halophilic microbes inhabiting the Pleistocene hypersaline lakes of the Qaidam Basin.
The precipitation sequence glauberite (Na2SO4·CaSO4), halite (NaCl), thenardite (Na2SO4), bloedite (Na2SO4·MgSO4·4H2O), and polyhalite (K2SO4·MgSO4·2CaSO4·2H2O) matches the saline minerals observed in core SG-1 (Fig. 1).
5. Discussion and Conclusion
Elevated salinity and falling a w in brines of the western Qaidam Basin over the past 2.26 million years coincide with stepwise aridification of the Asian interior during the Pleistocene (An et al., 2001; Guo et al., 2002; Ding et al., 2005). Loess and red clays from the Chinese Loess Plateau document expansion of deserts in central Asia during the Plio-Pleistocene, possibly linked to uplift of the Tibetan Plateau and Northern Hemisphere ice volume changes (Ding et al., 2005). Cores from the North Pacific show increased accumulation of eolian dust, derived from the Asian interior, for the past 2.7 million years, especially during the past 0.8 million years (Rea et al., 1998) (Fig. 1). Magnetic susceptibility from cores composed of loess from the Tarim Basin (Fig. 1) also document stepwise drying of central Asia over the past million years, especially at 0.33 and 0.13 Ma (Zan et al., 2010).
The above evidence for increasing aridity of the Asian interior coincides with the trend toward increased salinities and decreased a w in surface brines of the Qaidam Basin over the past 2.26 million years and the shift to lower halophile abundances in core SG-1 over the same period. Progressively increasing aridity led to decreased precipitation and water inflow to the western Qaidam Basin, and heightened evaporation rates. These climate changes produced more concentrated surface brines, which by 0.35 Ma reduced the habitability of surface lakes. By 0.13 Ma, saline lakes desiccated, and a w of ephemeral surface waters dropped <0.7, which approached the limits tolerated by halophilic prokaryotes and algae (Stevenson et al., 2015).
We conclude that climate change (aridification of the Asian interior) led to progressive increases in brine salinity in the Qaidam Basin, which reduced the habitability of saline lakes there. The a w, a thermodynamic parameter used to define the limits of life in the laboratory, can also be used to explain the response of microbial ecosystems to environmental changes in extreme natural environments on Earth and other parts of the Solar System (Tosca et al., 2008).
Footnotes
Acknowledgments
We thank our group members for input during drilling and lab work, and Robert Demicco for providing a modified EQL/EVP program. Research funding comes from the NSFC grant (41302133), National Basic Research Program of China (2011CB403007), the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB03020401), NSF Geobiology and Low Temperature Geochemistry grant EAR 1024692, China Postdoctoral Science Foundation (2013M540150), and German Research Foundation under the priority programme TiP (SPP 1371).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.