
Abstract
The thermodynamic availability of water (water activity) strictly limits microbial propagation on Earth, particularly in hypersaline environments. A considerable body of evidence indicates the existence of hypersaline surface waters throughout the history of Mars; therefore it is assumed that, as on Earth, water activity is a major limiting factor for martian habitability. However, the differing geological histories of Earth and Mars have driven variations in their respective aqueous geochemistry, with as-yet-unknown implications for habitability. Using a microbial community enrichment approach, we investigated microbial habitability for a suite of simulated martian brines. While the habitability of some martian brines was consistent with predictions made from water activity, others were uninhabitable even when the water activity was biologically permissive. We demonstrate experimentally that high ionic strength, driven to extremes on Mars by the ubiquitous occurrence of multivalent ions, renders these environments uninhabitable despite the presence of biologically available water. These findings show how the respective geological histories of Earth and Mars, which have produced differences in the planets' dominant water chemistries, have resulted in different physicochemical extremes which define the boundary space for microbial habitability. Key Words: Habitability—Mars—Salts—Water activity—Life in extreme environments. Astrobiology 16, 427–442.
1. Introduction
A
Historically, our knowledge of life in brines (where salinities exceed that found in seawater) has been derived from studies of terrestrial sodium- and chloride-rich environments that, even at saturation, are permissive for the biotic activity of some halophiles and are accordingly populated by dense microbial communities (Oren, 2008). In brine environments on Earth, microbial life is primarily limited by the thermodynamic availability of water (water activity) (Stevenson et al., 2015a, 2015b). The currently accepted limit to life in high salt environments is reached at a water activity of 0.611 (Stevenson et al., 2015b), close to the absolute limit for any cellular growth at a water activity of approximately 0.605 (Williams and Hallsworth, 2009). By extrapolation, this parameter has been considered to be the major limiting factor for habitability in martian brines (Tosca et al., 2008). Water activity is considered by the Committee on Space Research (COSPAR) and the NASA Mars Exploration Program Analysis Group (MEPAG) as a defining parameter for “Special Regions” on Mars (those regions where multiplication of known microbes could plausibly take place) (Rummel et al., 2014) and thus plays a central role in shaping planetary protection policy and Solar System exploration missions.
Planetary geological evolution can, however, result in different water chemistries, with undetermined implications for habitability. Investigations of terrestrial brine environments with chemistries that differ significantly from the dominant brine type on Earth are relatively few but often reveal salt-induced stresses that are otherwise lacking in NaCl brines. For example, MgCl2-rich brine lakes in the deep Mediterranean exhibit high chaotropicity (solute-induced macromolecule-disordering activity) alongside extremely low water activity, exacerbating their hostility and defining the limits of colonization in the brine-seawater interface (Hallsworth et al., 2007; Yakimov et al., 2015). Furthermore, previous studies on salt stress have highlighted adverse effects caused by salt ions that cannot be explained by osmotic stress or low water activity (Lloret et al., 1995; Alves et al., 2015).
The surface evolution of Mars has given rise to significantly different water chemistries; notably the widespread production of waters with high Mg2+, Fe2/3+, and SO4 2- contents (Catling, 1999; Bullock et al., 2004; Knoll et al., 2005; Carr and Head, 2010; Tosca et al., 2011). Due to high divalent:monovalent ratios (Fig. 1), such waters form brines with a high charge density (ionic strength) even at relatively clement water activities. Brine environments on Earth that contain elevated levels of divalent ions, such as the Mg2+-rich Dead Sea, and MgCl2 brines in the deep Mediterranean, commonly contain Cl- as the dominant anion (Grant et al., 1999; Wallmann et al., 2002), and therefore their divalent:monovalent ratios rarely exceed 1 (Fig. 1). A notable exception is the Basque Lakes, in British Columbia, which are rich in magnesium sulfate salts (Eugster and Hardie, 1978). Here, the divalent content far exceeds that found in the Dead Sea and other brines considered as divalent-rich, and it approaches that of some martian brines (Fig. 1).
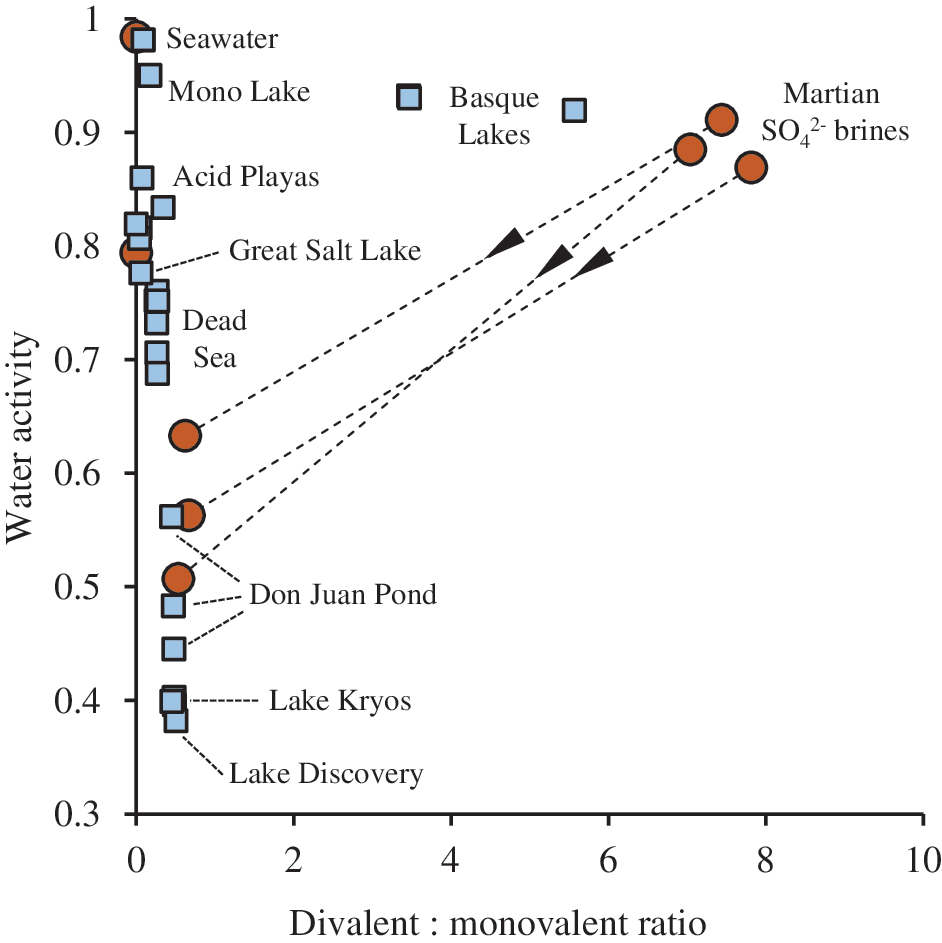
Divalent:monovalent ratios plotted against water activity of modeled martian brines (circles) and terrestrial brine environments (squares). Arrows indicate modelled evaporative concentration (Tosca et al., 2011). For details of terrestrial brine calculations and sources, see Materials and Methods and Table 4.
Due to a complex dependency on charge interactions in biological molecules, high ionic strength can perturb native structure and function. High charge density is capable of inducing deformations in molecules such as nucleic acids and proteins (Baldwin, 1996; Kunz et al., 2004). Many adverse ion–biomolecule interactions are exacerbated in the presence of di- or multivalent ions, including water-activity reduction, chaotropicity, and kosmotropicity as well as associated aggregating/denaturing phenomena (Hofmeister effects), protein and nucleic acid destabilization and lipid bilayer disruption (Kirkwood 1943; Green and Hughes, 1955; Baumann et al., 1997; Dominy et al., 2002; Collins, 2004; Cray et al., 2013; Ball and Hallsworth, 2015). We therefore hypothesized that the elevated divalent:monovalent ratios in martian waters, compared to the majority of waters on Earth (Fig. 1), cause ionic strength to play a role in defining the window for habitability, even when water activity is permissive.
As well as containing high levels of divalent ions, martian brines exert multiple physicochemical extremes, including low pH, low water activity, and high levels of dissolved iron (depending on brine composition). In this study, we carry out the first systematic assessment of the microbial habitability of laboratory-synthesized martian brines. In contrast to chloride-dominated brines on Earth in which microbial propagation is primarily limited by water activity, the results presented here show that high ionic strength in martian brines constrains their habitability to a smaller window than current paradigms predict.
2. Materials and Methods
2.1. Simulated martian brines
Naturally occurring saline environments on Earth with compositions matching those modeled for martian environments have not been reported (Fig. 1). Therefore we synthesized martian brines based on computational reconstructions of evaporative brine formation on the martian surface (Tosca et al., 2011). Brine compositions are known to change significantly as evaporation proceeds (Eugster and Hardie, 1978), and the computational approach employed by these authors produced two stages of concentration for each brine (Stages [a] and [b]), allowing us to probe the effects that natural evaporative concentration can have on habitability. For information on the computational approach used to predict this evaporation and generate these two stages, see Tosca et al. (2011).
The martian brines considered for this work were grouped into three types/classes, representative of diverse saline environments on Mars. These were alkaline carbonate-chloride brines (Type I), which during their more dilute phase are analogous to brackish fluids that persisted at the Curiosity rover's landing site in Gale Crater approximately 3.7 billion years ago (Léveillé et al., 2014). Upon simulated concentration, Type I brines evolved a concentrated K-Na-HCO3-Cl composition similar to fluids that interacted with the Nakhla martian meteorite (Bridges and Schwenzer, 2012). Type II brines were Mg-SO4-Cl-dominated, with comparatively low Na and K concentrations, and are characteristic of widespread large-scale Hesperian-aged salt (evaporite) deposits on Mars, such as those investigated by the Mars Exploration Rover Opportunity at Meridiani Planum (Knoll et al., 2005). Type III brines were similar in composition to Type II brines but contained higher levels of dissolved iron, resulting in brines that were extremely acidic at both stages of simulated concentration. In both Types II and III martian brines, initially high divalent:monovalent ion ratios decreased dramatically following simulated evapoconcentration due to the relative solubility of chlorides (Fig. 1). Both Type II and Type III brines were characterized by high levels of sulfates, which, as well as forming the dominant salt type in many evaporite deposits on Mars, are an abundant component of the globally distributed martian dust (Vaniman et al., 2004; Karunatillake et al., 2014). Type I and II brines were each represented by one evaporation pathway, whereas two evaporation pathways were investigated for Type III brines to capture the compositional and physicochemical diversity possible in their evolution.
Brine compositions for both stages of concentration were taken from Tosca et al. (2011) (Table 1). Salts were dissolved in deionized water, supplemented with 4 g L−1 yeast extract (Oxoid), and the solutions were stirred continuously for approximately 3 h to ensure maximum dissolution. Yeast extract was selected as a carbon source as it provides an extensive inventory of proteins, amino acids, and sugars. Preliminary enrichments in Type I and Type II Stage [a] brines supplemented with peptone, casamino acids, and glucose generally yielded less biomass than did yeast-extract enrichments (data not shown). Due to saturating concentrations of salts in some solutions, brines were left at 30°C for 5 days to allow full equilibration of solid and liquid phases. Simulated martian brine solutions were not buffered; pH was left to vary with the salt component to simulate natural brine conditions. Solutions were then split into equal volumes for aerobic and anaerobic culture and filter-sterilized (0.22 μm diameter pores) into pre-autoclaved culture vessels; anaerobic brines were purged with N2 to remove oxygen and sealed in sterilized 100 mL serum bottles with butyl rubber stoppers to maintain anaerobic conditions. L-cysteine-HCl was added to the anaerobic brines to a final concentration of 0.8 mM from sterile anoxic stocks. An equivalent volume (0.1% v/v) of sterile distilled water was added to aerobic brines. Finally, samples were taken for quantification of water activity, pH, chao/kosmotropic activity, and ionic analyses (see below). Analysis of a pure 4 g L−1 yeast extract solution revealed that ionic strength was increased in all fluids in the current study by <0.004 mol L−1 as a consequence of yeast extract supplementation.
2.2. Environmental inoculum sources
To maximize our chances of obtaining organisms capable of colonizing the brines, we sampled a range of environmental microbial habitats. All sampling was carried out using presterilized sample bags and/or centrifuge tubes. Where possible, samples were obtained from ≥5 cm sediment depths to increase chances of sampling anaerobic organisms as well as aerobes. Samples were stored at 4°C until use. A composite inoculum, made up of two environmental samples that were each added at approximately 1% (v/v) to prepared volumes of brine, was used to screen all brines for evidence of microbial growth. The first—local soil in Edinburgh (UK)—was selected because it has been previously shown that the physicochemical, temporal, and spatial variability within topsoils have selected for organisms that are tolerant of a range of extremes (Young et al., 2008). Preliminary community analysis via 454 pyrosequencing of the Edinburgh soil revealed a typically high diversity of metabolically diverse taxa (Shannon's H = 6.007 ± 0.044, Good's coverage = 92.65% at 97% OTU similarity). The top layers (approximately 5 cm) of this soil cycle between hydration and complete desiccation, driving extreme transitions in solute concentration(s) on a submillimeter scale. As such, these soils represented a source of both high microbial diversity and physicochemical heterogeneity. A sample comprised of a mixture of brine and brine-saturated sediment from a 1.1 km deep subsurface evaporite deposit (Boulby International Subsurface Astrobiology Laboratory, Boulby Mine, Whitby, North Yorkshire, UK) formed the other half of the composite inoculum. Water pH at time of sampling was approximately 7 (Payler, unpublished). Chemical analyses showed this brine to be dominated by NaCl close to saturation, and it is known to support an active community of halophilic microorganisms (Payler, unpublished).
Where the composite inoculum failed to produce growth, additional inoculum sources were: (1) marginal mud from an acidic hydrothermal pool at Kverkfjöll Volcano, Iceland (64°41.205′N, 16°40.502′W) (Cousins et al., 2013). The pool water contained high levels of dissolved iron (130 mM), sulfate (19.3 g L−1), and extremely low pH (1.75) at the time of sampling, values typical of those found in acid mine drainage sites such as Río Tinto (Fernández-Remolar et al., 2004). (2) Brine and sediments from the MgSO4-brine Basque Lakes on the Cariboo Plateau, British Columbia (50°35.596′N, 121°20.934′W). These are some of the only known hypersaline environments on Earth where sulfate forms the dominant anion (Nesbitt, 1990), and divalent:monovalent ratios reach values much greater than 1. As such, they represent perhaps the best terrestrial analogue for divalent-rich martian brines. Lake waters are known to fluctuate in concentration dramatically depending on season (Nesbitt, 1990), and at time of sampling (February 2015), the lake water was in a relatively dilute phase, containing 252 mM Mg, 243 mM sulfate, 71 mM Na, and <5 mM Cl. Lake water pH was 5.80, the sulfate:chloride ratio was 33.3, and the divalent:monovalent ratio was 5.56 (Fig. 1).
Any environmental inoculum contains a finite number of organisms. Thus for any brine that failed to support colonization by the environmental inocula and based on the rationale that “everything is everywhere, but the environment selects” (Baas Becking, 1934), we also placed 100 mL volumes outdoors, open to the atmosphere under a rain cover for 1 month to allow colonization by airborne microbes. The rain cover was a slanted plastic ceiling placed approximately 30 cm above the vessels' openings.
Together, these samples provided a high probability of enrichment for organisms that tolerate the unique combination of stresses present in martian brines. To confirm this, we designed a suite of control brines (Control-1 to Control-6) that systematically validated the tolerance of these inocula to physicochemical extremes of relevance to our experiments (Tables 2 and 3). These were prepared and inoculated with the environmental samples (2% v/v) in triplicate both aerobically and anaerobically in an identical manner to the Mars-relevant brines described above, and were designed to exhibit low water activity (Control-1), low pH (Control-2), combined low pH/low water activity (Control-3), and high levels of dissolved iron (Control-4). Control-5 and Control-6 were designed to exhibit high ionic strength, neutral pH, and permissive water activity (Table 3).
These were designed to test the tolerance of our inoculum communities to low water activity (Control-1), low pH (Control-2), combined low water activity/low pH (Control-3), combined high iron concentration/low pH (Control-4), and high ionic strength (Control-5 and Control-6). Concentrations are in mol L−1. All brines were also supplemented with 4 g L−1 yeast extract.
Concentrations are in mol L−1
Iron concentration and resulting ionic strength taken as average measured iron concentration over incubation period. See Materials and Methods and Fig. S1.
2.3. Incubation
Coping with osmotic stress induced by high levels of salts is energetically expensive (Oren, 2011). Previous analyses of growth data for 241 isolated strains revealed that aerobic organisms and anaerobic organisms that use organics as a terminal electron acceptor were tolerant of a broader range of extremes, including salinity, than anaerobic organisms that utilize inorganic electron acceptors (Harrison et al., 2015). By supplying a rich, complex source of organic carbon (4 g L−1 yeast extract) and a temperature of 30°C, we therefore expected to increase the energetic favorability of respiratory metabolisms and thus the capabilities of microorganisms to deal with the stresses induced by our brines (Oren, 2011). This ensured that apart from the extremes of the brines, the organisms had optimum growth conditions with respect to temperature, energy, and nutrient availability. Our experiment was focused on determining whether the martian brine chemistries alone are limiting to life.
All brines were inoculated in triplicate (2% v/v) and incubated at 30°C for 60 days, then transferred (1% v/v) to fresh, sterile brine media. Further transfers were carried out at appropriate time points, which differed by brine and community. Brines that had been exposed to the atmosphere for 1 month were incubated at 30°C for a further 30 days before also being transferred (1% v/v) to fresh, sterile brine media. For brines that did not contain solid salt precipitate or dissolved iron, growth was quantified as an increase in optical density at 600 nm. In saturated brines and those containing dissolved iron, cells were enumerated by direct counts following SYBR gold or DAPI staining (see below). After three transfers, when cell densities reached approximate maxima, cells were harvested by filtration onto sterile 25 mm polycarbonate filters (Merck Millipore) for DNA extraction. Initial enrichment-stage brines that did not support growth after 60 days were incubated alongside the transfers and monitored at regular intervals for the remainder of the experiment (>300 days).
2.4. Assays for microbial growth
The ability of the martian and control brines to support microbial growth was assayed via three independent methods. Firstly, samples of brines (approx. 20 μL) were mounted on microscope slides and examined for evidence of cells under phase contrast microscopy (Leica DM4000B). Secondly, brine samples (200 μL) were stained with 1× SYBR gold nucleic acids stain (Life Technologies) for 15 min in the dark, mounted on black 25 mm diameter polycarbonate filters (Merck Millipore), excited at 450–490 nm, and examined at 1000× magnification using a Leica DM4000B digital microscope and a Leica DFC 450 C microscope-mounted camera. For iron-rich brines, 1× DAPI (4′,6-diamidino-2-phenylindole) (Sigma) was found to be more reliable. For DAPI staining, samples were prepared in an identical way to SYBR-stained samples, and excited at 358 nm. Where applicable, cells were enumerated by counting 20 randomly selected fields of view and averaging over triplicate samples.
To validate microscopic approaches, we enriched communities from our composite inoculum in nutrient broth media (Oxoid), harvested aliquots by centrifugation, suspended them in samples of each brine, and subjected them to identical staining and imaging protocols as those used for the brine enrichments. Imaging of the organisms was possible in all brines (data not shown).
Thirdly, DNA was extracted from 2–10 mL of brine from the final transfer stage using a modified phenol:chloroform:isoamyl alcohol and isopropanol precipitation protocol as detailed by Urakawa et al. (2010). Briefly, samples were passed through 0.22 μm, 25 mm diameter polycarbonate filters. Filters were treated with proteinase K (2 mg mL−1) and TENS buffer (50 mM Tris-HCl; pH 8.0, 20 mM EDTA, 100 mM NaCl, 1% w/v SDS) at 50°C for 1 h. DNA, if present, was then extracted with phenol:chloroform:isoamyl alcohol (25:24:1) and precipitated with isopropanol. DNA extracts were quantified by spectrophotometric absorbance at 280 nm (NanoDrop Lite, BioRad), visualized in 1% agarose gels with a SynGene G-Box UV transilluminator, and further interrogated by polymerase chain reaction (PCR) (see below).
This third approach was validated by adding communities enriched from our composite inoculum in nutrient broth media (Oxoid) to quantities of all brines, at cell densities approximately equivalent to the lowest obtained in our experiments (Type I Stage [b]), and subjecting them to identical extraction and DNA detection procedures. Positive extraction and domain-specific PCR amplification were achieved from all brines. For a brine to be labeled “uninhabitable” in the context of the current study required concurrent negative results from both microscopic methods at all transfer stages as well as negative DNA-based detection.
2.5. Ionic strength, pH, water activity, and chaotropic/kosmotropic activity quantification
Ionic strength was calculated from measured ion concentrations using the following equation:
where ci = the concentration of ion i (in mol L−1), and zi = the charge of ion i. pH was measured in triplicate using an Omega PHH-37 pH meter with Omega PHE 1335 probe setup calibrated to three points (pH 4.0, 7.0, and 10.0) with standard solutions supplied by the manufacturer. Water activity was quantified using 5 mL samples at 30°C in a Rotronic HP23-AW water activity meter, calibrated to five points (a w = 0.325, 0.595, 0.755, 0.845, and 0.935) using saturated calibration standards (MgCl2, NH4NO3, NaCl, KCl, and KH2PO4, respectively) prepared as described by Winston and Bates (1960). Each brine was measured three times, and results were found to be within ±0.002 a w (data not shown). During incubation, water activity was quantified at approximately 2-week (14-day) intervals and found to vary by ≤0.008 a w over the course of 60-day incubation periods (data not shown).
Chaotropic/kosmotropic activities of the eight brines were quantified by measuring the increase or decrease in gelation temperature of a brine/agar solution relative to a pure agar solution as described previously (Hallsworth et al., 2003; Cray et al., 2013). An increase in agar-gelation temperature relative to that of pure agar was indicative of kosmotropicity, whereas a decrease in gelation temperature was indicative of chaotropicity. Where brines caused precipitation of agar, a dilution series was made in order to construct curves that were used to derive extrapolated values (see Cray et al., 2013).
2.6. Ionic composition analysis
Chloride and sulfate ions were analyzed at the University of Edinburgh, UK, via ion chromatography using a Dionex DX-120 system fitted with a conductivity detector, according to manufacturer's instructions. Magnesium, potassium, sodium, and total iron concentrations were quantified via atomic absorption spectroscopy by the University of Sheffield Groundwater Protection and Restoration Group using a Perkin Elmer AAnalyst 200 spectrometer. Radiation was provided at 248.3 nm by an iron hollow cathode lamp (slit 1.8/1.35), and measurements were integrated over 5 s and performed in triplicate.
Changes in ferrous and ferric iron concentrations in Control-4 were monitored colorimetrically throughout incubation periods using the ferrozine assay as previously described (Stookey, 1970). Briefly, samples were digested in 0.5 M HCl for 1 h and added to HEPES-buffered ferrozine solution. Absorbance was measured at 562 nm in a Helios Alpha spectrophotometer (Thermo Fischer Scientific).
Bicarbonate concentrations in Type I martian brines were quantified by titrimetric determination of alkalinity. Samples were titrated with HCl until pH 4.5 was reached, indicating all bicarbonate had been neutralized. HCO3 concentration was then determined using the equation
where A is the total alkalinity (in mg L−1), c(HCl) is the concentration (mol L−1) of the HCl solution used, v 1 is the volume of HCl titrated, and v 2 is the volume of sample used.
2.7. Comparison with physicochemical data from terrestrial brines
For comparisons of martian brines and terrestrial brine environments, physicochemical data was derived from sites summarized in Table 4. When not reported in the source publications, pH and water activity of natural terrestrial brines were calculated from ionic composition using the thermodynamic model FREZCHEM version 16 (Marion and Kargel, 2008). FREZCHEM v. 16 employs Pitzer equations for calculating ion interactions at high ionic strength. Ion compositions were converted from units reported in source publications to moles per kilogram of water, and calculations were performed at 30°C, with pH controlled through equilibrium between H+ and CO2 (gaseous) at approximately terrestrial atmospheric partial pressure (0.04 atm). For more information, see Marion and Kargel (2008).
a w = water activity.
2.8. PCR amplification
Community DNA was interrogated by bacterial, archaeal, and eukaryotic domain-specific primers targeting ribosomal small subunit (SSU) RNA. For oligomer sequences used as primers in the current study, see Table 5. Each individual 25 μL PCR reaction contained 1 μL template, 0.4 μM of the relevant forward and reverse primer, 200 μM dNTPs, 1.5 mM MgCl2, 1× PCR buffer, and 1 unit Taq polymerase (Invitrogen). PCR conditions were as follows: for 28F-519R, reactions were subjected to denaturation at 95°C for 5 min, followed by 30 cycles of 94°C for 30 s, annealing at 60°C for 40 s and extension at 72°C for 60 s, and finished with a final extension step at 72°C for 10 min. For 341F-958R, reactions were subjected to denaturation at 95°C for 5 min, followed by 35 cycles of denaturation at 95°C for 30 s, annealing at 54°C and extension at 72°C, and finished with a final extension step at 72°C for 10 min. For Euk1A-516R, reactions were subjected to denaturation at 94°C for 5 min, followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 56°C for 45 s and extension at 72°C for 60 s, and finished with a final extension step at 72°C for 5 min. Positive PCR amplification was confirmed by electrophoresis in 1% agarose gels made up in TAE buffer (40 mM Tris base, 20 mM acetic acid, 1.5 mM EDTA) and visualized using a SynGene G-Box UV transilluminator.
2.9. 16S rRNA 454 pyrosequencing and bioinformatic analyses
Martian brine enrichments originating from the composite inoculum that yielded positive DNA extractions and either bacteria or archaea domain-specific PCR amplification were pyrosequenced using the Roche 454 platform (Research and Testing Laboratory of the South Plains, Lubbock, Texas, USA). A composite inoculum–derived Control-1 enrichment community was also sequenced for comparison. Initial trimming, denoising, and chimera checking were carried out by Research and Testing (Edgar, 2010, 2011; Edgar et al., 2013). Operational taxonomic unit (OTU) clustering and taxonomic identification were performed in the MOTHUR program using previously described standard operating procedures (Schloss et al., 2009, 2011; Quast et al., 2013). Pyrosequencing data sets were deposited with the Sequence Read Archive (NCBI) under the accession number SRP052574.
3. Results
3.1. Habitability of martian brines
Only three of the eight simulated martian brines supported microbial growth, despite several brines exhibiting permissive water activities and regardless of inoculum source or oxygen availability (Table 6). Among simulated martian brines, there were no differences in colonization when diverse inoculum sources were used: those brines that were colonized were colonized by all environmental inocula tested, and those that remained uninhabited were consistently prohibitive across all inoculum sources (Table 6). Furthermore, initial enrichment stages of uninhabited brines did not yield any evidence of growth after incubation for more than 300 days.
Columns correspond to the different inoculum sources used and to oxygen status (whether aerobic or anaerobic conditions). The + indicates successful colonization, and the − indicates lack of growth. nd = not determined.
Type I brines, similar to the composition of Na-K-Cl-HCO3 hydrothermal brines that likely chemically altered the Nakhla martian meteorite (Bridges and Schwenzer, 2012), were colonized at both stages of concentration (Table 6). Type II brines, relevant to large areas of martian layered sulfate terrains including those in Valles Marineris, Margaritifer Sinus, and Terra Meridiani (Gendrin et al., 2005), were inhabited at the initial dilute Stage [a], but evaporative Stage [b] was hostile to all sources of inoculum under all conditions (Table 6). Type III brines, which resemble an ancient Meridiani Planum and other Fe-Mg-SO4-Cl Hesperian environments (Knoll et al., 2005), were not colonized at either stage of concentration. Consistently, exposure to the atmosphere for 1 month did not result in successful colonization of Type II Stage [b] or Type III brines.
3.2. Microbial communities in martian and control brines
Among those brines that were colonized, biodiversity, cellular morphologies, and growth dynamics varied substantially between brine types and evaporitic stages (Supplementary Figs. S1 and S2; Supplementary Data are available online at
The + and − indicate presence or absence (respectively) of domain. Oxygen status is indicated by an A (aerobic conditions) or An (anaerobic conditions).
Growth demonstrated by direct cell counts only (DNA was not successfully extracted).
In brine enrichments that originated from the composite inoculum, archaeal and bacterial 16S rRNA pyrosequencing revealed distinct prokaryotic communities, which varied depending on the presence or absence of oxygen (Fig. 2). The highest bacterial diversity was recorded in the anaerobic treatment of the most dilute of all simulated martian brines: Type I Stage [a] (Shannon's H′ = 3.500 ± 0.051; Good's coverage = 96.8% at 97% OTU similarity; Figs. 2 and S3). This community was dominated by members of the Firmicutes, notably the genus Anaerobranca and an unclassified genus within Peptostreptococcaceae (Figs. 2 and S3). The aerobic treatment of this brine supported a lower-diversity community in which the genera Brevundimonas and Achromobacter, Alpha- and Betaproteobacteria, respectively, were dominant members (Shannon's H′ = 1.445 ± 0.026; Good's coverage = 99.1% at 97% OTU similarity; Figs. 2 and S3). Type I Stage [b], a later evaporative stage of Type I brines rich in chloride salts, supported a moderately diverse, mixed population of Firmicutes and Gammaproteobacteria, including Oceanobacillus and Halovibrio, both genera known to exhibit halotolerance (Takami et al., 2002; Sorokin et al., 2006) (Shannon's H′ = 1.800 ± 0.081; Good's coverage = 97.8% at 97% OTU similarity; Figs. 2 and S3).
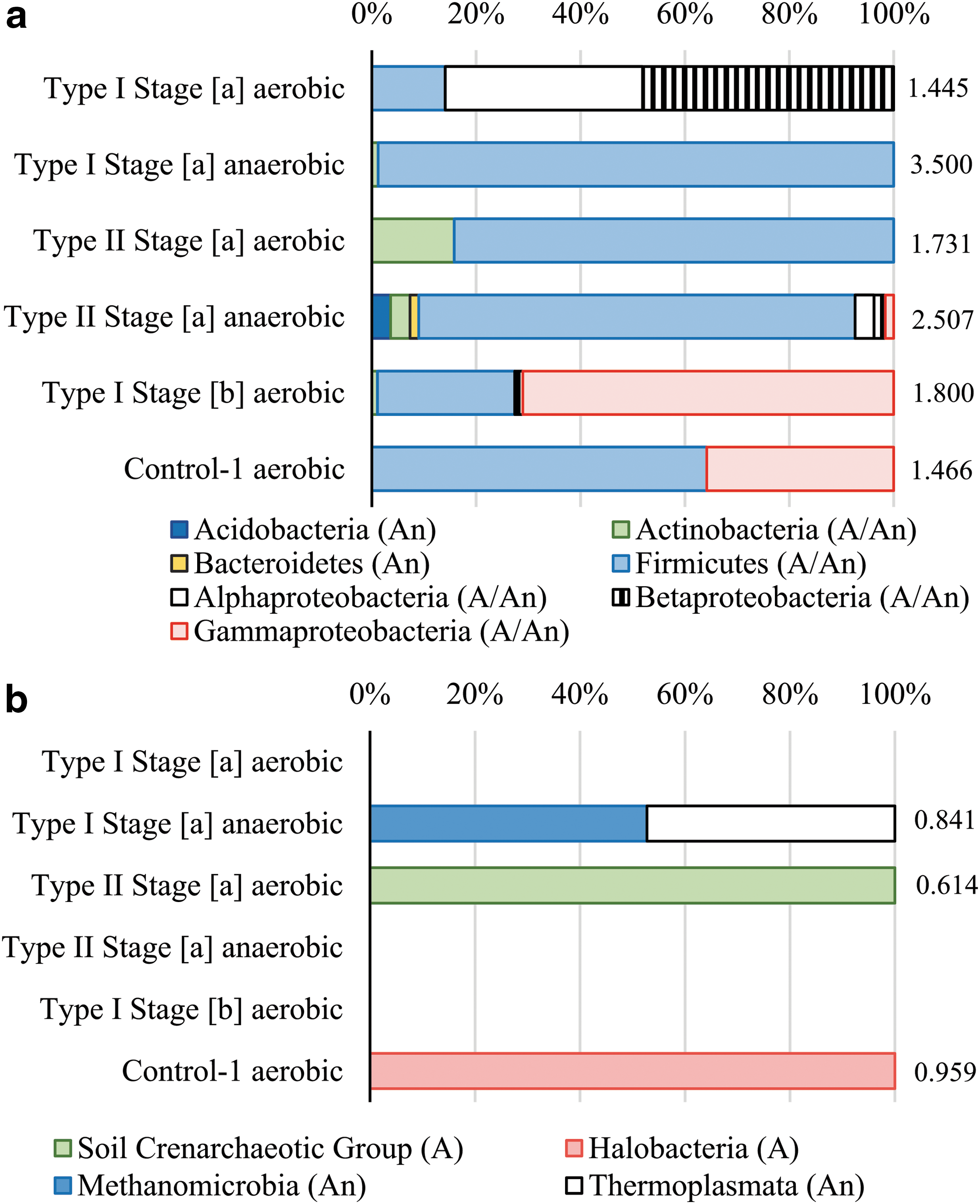
Relative abundances of bacterial phyla (
Type II Stage [a], a magnesium- and sulfate-dominated brine with the highest divalent ion content of any inhabited Mars-relevant brines, supported a moderately diverse community of Firmicutes (including Bacillus) and Actinobacteria (including Arthrobacter) under aerobic conditions (Shannon's H′ = 1.731 ± 0.038; Good's coverage = 98.5% at 97% OTU similarity; Figs. 2 and S3) and a marginally more diverse anaerobic community consisting mainly of facultatively anaerobic Firmicutes such as Virgibacillus (Shannon's H′ = 2.507 ± 0.087; Good's coverage = 95.9% at 97% OTU similarity; Figs. 2 and S3).
Among the sequenced communities found to contain archaea, the archaeal diversity was typically low. The anaerobic Type I Stage [a] (Shannon's H′ = 0.841 ± 0.024; Good's coverage = 99.3% at 97% OTU similarity) was dominated by methanogenic genus Methanoculleus, as well as an unclassified genus within the Thermoplasmata (Figs. 2 and S3). Type II Stage [a], by contrast, was colonized by archaea only under aerobic conditions, and the community was entirely dominated by the Nitrososphaera genus within the Crenarchaeota (Shannon's H′ = 0.614 ± 0.035; Good's coverage = 99.4% at 97% OTU similarity; Figs. 2 and S3).
Control-1 exhibited a similar bacterial community to Type I brine Stage [b], including the Firmicutes Oceanobacillus and the Gammaproteobacteria Halovibrio (Shannon's H′ = 1.466 ± 0.034; Good's coverage = 98.9% at 97% OTU similarity). However, despite the similarities in bacterial community, the archaeal community in Control-1 (Shannon's H′ = 0.959 ± 0.046; Good's coverage = 98.8% at 97% OTU similarity) was markedly different from any simulated martian brine, being dominated by a single class of extremely halophilic archaea: the Halobacteria (Figs. 2 and S3).
3.3 Physicochemical controls on martian brine habitability
3.3.1. Water activity
The currently accepted limit to life in high salt is reached at a w = 0.611, and terrestrial environments that fall below this value are widely considered to be functionally sterile (Fig. 3a) (Stevenson et al., 2015a, 2015b). While the terrestrial brines with the lowest water activities, including the deep-sea Lakes Discovery and Kryos (located in the Mediterranean Sea) and Don Juan Pond in the McMurdo Dry Valleys, Antarctica, exhibit other biologically hostile physicochemical traits, their water activities fall below the minimum required for cellular division (Hallsworth et al., 2007; Samarkin et al., 2010; Yakimov et al., 2015). Apart from in some localized environments, such as the brine/seawater interface in Lakes Kryos and Discovery, where chaotropicity defines microbial habitability (Hallsworth et al., 2007; Yakimov et al., 2015), water activity sufficiently delineates the habitability of terrestrial saline environments (Fig. 3a).

Habitability of simulated martian brines (Type I–III, Stages [a] and [b]), control brines (C-1 to C-6), and terrestrial examples plotted as a function of water activity and pH (
The water activity of Type II Stage [b] (0.633 a w) was close to the biophysical limit for proliferation of extreme halophiles (Stevenson et al., 2015b) and lower than the water activity of any of the brines identified as habitable in the current study (Fig. 3a). By contrast, martian brine Type III Stage [a] exhibited permissive water activity (0.894 and 0.885) but did not allow growth of any microorganisms (Fig. 3a). This was despite the inoculum communities' ability to tolerate lower water activities: Type I Stage [b] (0.789 a w) and Control-1 (0.764 a w) were successfully colonized. Control-3 (0.889 a w), which was designed to directly simulate the water activity of Type III Stage [a], also supported a community of organisms (Fig. 3a).
3.3.2. pH
Low pH can be ruled out as the sole inhibitory factor in Type III Stage [a] due to the colonization by several inoculum sources of Control-2, which exhibited an equivalent pH to Type III Stage [a] (Fig. 3a; Tables 3 and 6). However, combined stresses of low pH and low water activity equivalent to those found in Type III Stage [a] restricted colonization to just one inoculum source, under aerobic conditions only (Control-3; pH 2.5, a w = 0.889) (Fig. 3a; Table 6). The community from Control-3 was not able to grow in Type III Stage [a].
3.3.3. Kosmotropicity
All the simulated martian brines investigated were found to be kosmotropic (macromolecule-rigidifying) (Fig. 3b). Type III Stage [b] exhibited a kosmotropic activity approximately equivalent to a solution of 5.5 M ammonium sulfate (Fig. 3b). This is despite Type III brines possessing high concentrations of ions including Mg2+, Fe2+, and Cl-, the salts of which are strong chaotropes when measured as solutions made up from pure salts (Cray et al., 2013). Although Type III martian brines exhibit extreme kosmotropic activities, the MgSO4-rich Type II Stage [a] was densely colonized by all inoculum sources and under both aerobic and anaerobic conditions, despite imposing a kosmotropic activity higher than the uninhabited Type III Stage [a] brines (Fig. 3b).
3.3.4. Iron toxicity
Despite the presence of high levels of iron in Type III brines, iron-induced oxidative stress can be eliminated as the sole determinant of their habitability. An aerobic community of bacteria from a single inoculum source (the acidic hydrothermal pool inoculum; see Materials and Methods) became established and grew successfully at pH 1.95 in the presence of approximately 600 mM dissolved iron in Control-4 (Fig. S2; Tables 6 and 7). This result is significant; no other inoculum source yielded organisms capable of growing in Control-4. Type III Stage [a] brines, which contained 597 and 628 mM Fe, did not support the growth of these organisms.
3.3.5. Ionic strength
All uninhabited brines, including both Type III Stage [a] brines, were characterized by extremely high ionic strength (>10 mol L−1) (Fig. 3). Control-5 and Control-6 were designed to exhibit high ionic strength but otherwise permissive physicochemical parameters. When all other stresses were minimized, high ionic strength dramatically restricted habitability. Only the MgSO4-rich Basque Lakes, British Columbia, which possess one of the highest divalent:monovalent ratios known in terrestrial brines (see Fig. 1, Materials and Methods), contained organisms capable of growth in Control-5 (ionic strength = 12.141 mol L−1; 0.821 a w; pH 7.0), and these only grew in the presence of oxygen (see Figs. 3c–3d, S1, S2). Domain-specific PCR revealed that the colonizing population consisted solely of bacteria (Table 7). Although they were tolerant of ionic strength higher than that found in Type III Stage [a] brines, the bacteria that colonized Control-5 were not capable of growth in Type III Stage [a].
The level at which ionic strength becomes inhibitory was influenced by water activity. At moderate ionic strength (5 mol L−1) and 0.764 a w in Control-1, rapid and extensive growth was observed (Figs. 3d and S1). However, at a slightly higher water activity (0.801) but greatly increased ionic strength (Control-6; 10.113 mol L−1), growth was inhibited under both oxygenated and anoxic conditions, regardless of inoculum source (Table 6). The Control-6 brine was the only control to remain uninhabited after inoculation across all inoculum sources. This was despite Control-6 exhibiting permissive water activity (0.801), pH (7.1), kosmotropicity (−76.42 kJ mol−1) and iron concentration (approximately 50 μM), levels that were directly demonstrated to be habitable by other control and martian brines (Fig. 3). Initial enrichments of Control-6 were also devoid of growth, after incubation for a period of >300 days.
4. Discussion
4.1. Microbial communities in martian brines
Brines relevant to saline environments on Mars supported distinct, complex, active microbial communities following inoculation by a variety of environmental sources. Variations in microbial community structure revealed by molecular analyses on the domain (Table 7), phylum and class (Fig. 2), and genus levels (Fig. S3), as well as different growth dynamics and cell densities (Figs. S1 and S2), demonstrated that differing ionic compositions can have an important influence in defining community structure. The notable detection of methanogenic Archaea in anaerobic treatments of Type I Stage [a], which was the most dilute Mars-relevant brine and most closely aligned with the Gale Crater paleoenvironment (Léveillé et al., 2014), shows that biological methanogenesis is possible in ancient Mars-relevant fluids. One plausible explanation for methanogenic growth is the production of hydrogen through fermentation driven by the bacterial community in this brine.
One notable finding from the microbial community composition data was that in all cases, martian brine microbial communities were distinct from that of Control-1, which represents the typical composition of NaCl-rich terrestrial environments. The high abundance of one particular archaeal genus (Haloarcula) in Control-1 is typical of NaCl brine lakes, which during blooms can become dominated by relatively few microbial taxa (in comparison to lower-salinity lakes) (Benlloch et al., 2002; Oren and Hallsworth, 2014). Despite some martian brines supporting colonization by known NaCl-tolerant bacteria, they all lacked halophilic archaea and other common inhabitants of NaCl-dominated brines (Figs. 2 and S3). Instead, they supported a diverse community of primarily nonhalophilic organisms. This observation provides a direct demonstration that martian brine environments are distinct from terrestrial brines and that the different geochemical histories of brines have implications for the types of communities that they can potentially support. These data also show that the use of terrestrial brines as analogues for brines found on Mars cannot necessarily reveal the microbial habitability of the latter; instead it is important to augment field studies with the synthesis of martian brines in the laboratory to understand more empirically the factors that define microbial habitability.
4.2. Factors that influence the habitability of martian brines
We systematically investigated the factors that influence habitability in extreme martian brines. This revealed that the habitability of Type I and II brines was consistent with predictions made from water activity. These relatively dilute brines supported growth at water activities above the currently accepted limit for life (0.611), except for Type II Stage [b], which was close to this limit (0.633). There have thus far been only three halophilic bacteria or archaea reported to grow at <0.700 water activity, according to empirical determinations (Stevenson et al., 2015a, 2015b). However, Type III Fe-Mg-SO4 brines were not habitable, even when possessing biologically permissive water activity (Fig. 3; Table 6).
The control solutions that we synthesized allowed us to identify the different physical and chemical extremes associated with the brines and to determine whether they, alone, can explain the hostility of the Type III brines. Low water activity (down to 0.764 a w), low pH (down to 1.95), and high kosmotropic activity (up to −324.35 kJ kg−1) were ruled out as sole inhibitory factors in Type III Stage [a] brines due to the colonization of control solutions possessing these extremes (Fig. 3; Table 6). Colonization of these control brines also rules out osmotic changes experienced by the inoculum communities during transfer from their source environment as the determinant of ability to grow in Type III Stage [a]. Organisms would have experienced equivalent or greater osmotic changes in the control solutions, and growth was not precluded.
High kosmotropicity in martian brines is notable; while chaotropicity can be a life-limiting parameter in diverse types of natural environments (e.g., Hallsworth et al., 2007; Cray et al., 2015; Yakimov et al., 2015), the level of kosmotropicity encountered in Type III martian brines (Fig. 3b) is rarely, if ever, encountered on Earth (Williams and Hallsworth, 2009; Lievens et al., 2015). The biophysical mechanisms that give rise to chaotropic/kosmotropic activities of solutes are extremely complex and not fully understood (Ball and Hallsworth, 2015). Such a high kosmotropic activity as that found in Type III martian brines, despite the presence of chaotropic salts (such as MgCl2 and FeCl2), highlights the need for empirical determinations of these activities in studies of natural environments, as kosmotropicity of complex mixtures cannot be predicted from those of pure salt values (Alves et al., 2015; Yakimov et al., 2015). Nevertheless, the establishment of microbial communities in Type II Stage [a] (−270.69 kJ kg−1) and Control-5 (−324.35 kJ kg−1), brines with higher kosmotropicity than Type III Stage [a], demonstrates that kosmotropicity at these levels alone does not limit microbial growth (Fig. 3b).
If we consider the number of environmental inocula established in each brine to be a crude proxy of its habitability, the data also allow us to extract generalizations regarding the biological hostility of single and combined extremes (Table 6). Combined low pH/low water activity (Control-3), iron toxicity (Control-4), and high ionic strength (Control-5) all only allowed growth from one inoculum source, which differed for each of these controls. This shows that although these extremes in isolation do not prevent growth from all the inocula used, they do restrict colonization to organisms from fewer environments, suggesting that they contribute to the limits of habitability of the most extreme martian brines (Fig. 3; Table 6).
This finding is consistent with previous observations. Coping with co-occurring extremes of low pH and low water activity demands energetically expensive homeostasis strategies, and this combination is known to restrict the growth of terrestrial microorganisms (Harrison et al., 2013, 2015). Iron toxicity is caused primarily by the generation of oxidative hydroxide radicals through Fenton's reaction series (Gutteridge and Halliwell, 1989), and the hostility of this process toward biologically important organic molecules has previously been demonstrated in simulated martian brines (Johnson and Pratt, 2010). Ionic strength, a measure of charge density, is capable of inducing structural deformities and inhibition of biological molecules (Baldwin, 1996; Kohn et al., 1997; Kunz et al., 2004; Cray et al., 2013). At high ionic strength, therefore, the magnitude and extent of ion-biomolecule interactions may function as a stressor on microbial cells.
4.3. Ionic strength is a novel factor that limits the habitability of martian aqueous environments
Ionic strength was found to limit the habitability of control brines. Colonization was restricted to only one inoculum source in Control-5 (ionic strength = 12.141 mol L−1), which possessed a relatively clement water activity (0.821 a w). Furthermore, growth was inhibited entirely in Control-6 (ionic strength = 10.113 mol L−1), which exhibited a lower, but still demonstrably permissive, water activity (0.801 a w) (Table 3). The higher water activity in Control-5 (0.821 compared to 0.801 in Control-6) may explain its capacity to support restricted growth. These data indicate that in martian brines with high divalent ion content, particularly the Type III brines, ionic strength can act as a barrier to habitability.
Ionic strength per se has not previously been considered as an important parameter in restricting microbial growth in natural environments. This is likely due to the dearth of large-scale environments on Earth with sufficient divalent ion content. Terrestrial saline waters, which typically exhibit low divalent:monovalent ratios (Fig. 1) (Eugster and Hardie, 1978), only develop high ionic strength in extremely concentrated brines that also impose hostile water activities (Fig. 3d). Indeed, even Mg2+-rich bittern brines commonly contain chloride as the dominant anion, ensuring that the divalent:monovalent ratio does not exceed 1 (Fig. 1). By contrast, throughout large periods of Mars' surface evolution, high divalent:monovalent ion ratios were common (Catling, 1999; Vaniman et al., 2004; Knoll et al., 2005; Tosca et al., 2011), allowing the formation of brines with high ionic strength, even at moderate, biologically permissive water activities (Figs. 1 and 3d).
It is thought that more than 99% of microorganisms on Earth resist cultivation using current techniques (Amann et al., 1995). Therefore, it cannot be ruled out that organisms currently resistant to cultivation exist that are capable of growth under the conditions found to be uninhabitable in this study. This potential bias was mitigated here by studying a wide range of inocula and using enrichment communities. Cultured communities simulate the complex interdependences of organisms in the natural environment and thus capture a more representative snapshot of natural microbial assemblages (Alain and Querellou, 2009).
The data obtained in the current study demonstrate that a sampling or experimental bias does not explain our results: many organisms were successfully enriched under single or combined conditions found in Type III martian brines and yet were not capable of growth in Type III Stage [a], even after incubation for >300 days. This lack of growth, observed across all inoculum sources and independent of the presence or absence of oxygen, must therefore be attributable to conditions present in the Type III martian brines but that are not present in the habitable martian and control brines. Based on the elimination of other possible explanations, ionic strength must be one of these conditions that limits habitability in martian brines.
4.4. Conclusions and implications
Martian brines are complex, multi-stress environments that present significant challenges to biology. The results presented here support the hypothesis that high ionic strength can restrict habitability in high salt environments, even if water activity is permissive. In combination with other extremes such as high iron concentration and combined low pH/low water activity, high ionic strength explained the lack of colonization in Type III martian brines. Ionic strength can therefore act as a barrier to martian habitability.
We note that our results are conservative, since when combined with other multiple stressors such as low temperature, low energy availability, and high radiation flux, as might be expected on Mars, the brines would likely be even more hostile than under the conditions investigated here. As brines with extremely high divalent ion content have formed on Mars but do not commonly form on Earth, these findings are an example of how differing planetary-scale geochemistries, themselves dictated by geological evolution, can drive fundamental differences in habitability. On Earth, a chloride and monovalent ion-rich aqueous chemistry permits the microbial colonization of brines with exceptionally low water availability—indeed close to the absolute limit for life. By contrast, on Mars a chemistry dominated by divalent ions such as sulfates means that high ionic strength constrains habitability to a smaller window. An enrichment of divalent ions relative to Earth may not be limited to martian aqueous geochemistry. There is evidence that the putative subsurface ocean on Europa may contain significant amounts of Mg2+ and SO4 2- ions (Orlando et al., 2005). Constraints placed on this composition by future missions will allow for a prediction of the habitability of this jovian satellite.
Whereas brines are considered a reservoir of possibly habitable liquid water on present-day Mars, their prohibitively high ionic strength now casts doubt on this assumption. We question whether the current definition of Mars Special Regions based on temperature and water activity alone (Rummel et al., 2014) is sufficiently accurate for the purpose of planetary protection. High ionic strength may render an environment uninhabitable even if temperature and water activity are permissive. Meaningful assessments of biological permissibility for such brines are critical, both in considerations for extant or historical martian biota and in considering regions at risk from contamination with terrestrial microbes. These data also challenge the paradigm of “Follow the Water” in Mars exploration (Hubbard et al., 2002), demonstrating experimentally that aqueous environments need not be habitable. Indeed, martian brines may be some of the least-promising places to search for life.
Footnotes
Acknowledgments
Thanks to Nicholas J. Tosca (University of Oxford), Lorna Dougan (University of Leeds), and Jonathan A. Cray (Queen's University Belfast) for useful discussions. Thanks also to Samuel J. Payler (University of Edinburgh) and to Cleveland Potash Ltd. for their cooperation and for allowing access to the deep subsurface evaporite deposits and brines at Boulby Mine. Claire R. Cousins is supported by a Royal Society of Edinburgh Personal Research Fellowship. We acknowledge Vatnajökull National Park, Iceland, for a research permit to obtain the sample from Kverkfjöll that was used in this study. Funding for this work was provided by the UK Space Agency as part of the Aurora Science program. Support was also provided by Science and Technology Facilities Council (STFC) Grant no. ST/M001261/1.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.