
Abstract
Nitrogen is an essential nutrient for all life on Earth and possibly elsewhere. Burial of nitrogen bound to organic matter constitutes the major flux of nitrogen into sediments today, which has led to the inference that nitrogen enrichments in sedimentary rocks may be a biosignature. However, abiotic processes such as lightning or volcanism can fix atmospheric N2 and contribute to sedimentary nitrogen burial in the absence of life. It is therefore uncertain whether observed nitrogen enrichments of up to 430 ppm in Paleoarchean metasedimentary biotite grains are indeed biogenic. This study seeks to address that problem with a numerical model. The NH4 + concentration of an abiotic ocean is modeled as a function of source fluxes, pH-dependent NH3 volatilization, and equilibrated adsorption of NH4 + onto clay particles. The results suggest that the observed nitrogen concentrations in Paleoarchean biotite can only be reconciled with purely abiotic processes if the ocean was more acidic (pH <6) and/or if the source fluxes from lightning and volcanism were at least an order of magnitude higher (≥1012 mol/yr) than previously thought. The bulk of the nitrogen is thus most likely of biological origin. While this does not necessitate a particular metabolism such as biological N2 fixation, the data provide evidence of nitrogen utilization back to 3.8 Gyr. Nitrogen abundances could thus provide useful information in extraterrestrial missions. Key Words: Early Earth—Biosignatures—Nitrogen fixation. Astrobiology 16, 730–735.
1. Introduction
T
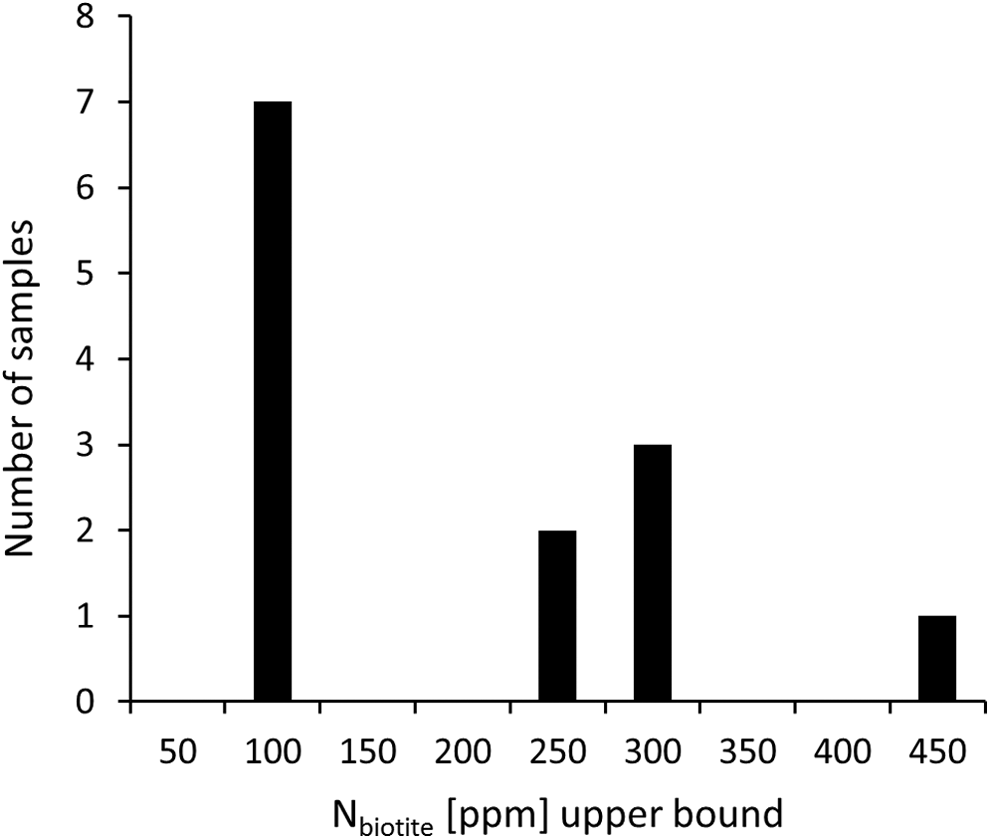
N concentrations in Paleoarchean biotite from the Isua Supracrustal Belt. Biotite grains were separated from metasedimentary rocks. Data are compiled from the works of Honma (1996), Pinti et al. (2001), and Papineau et al. (2005). Concentrations tend to be lower in whole-rock samples due to dilution with N-poor minerals such as quartz or volcaniclastic debris.
NH4 + uptake into the crystal lattice of clay minerals is preceded by adsorption onto the mineral surface (Schroeder and McLain, 1998). On modern Earth, this occurs primarily in pore waters of anoxic sediments, where dissolved NH4 + can be present in millimolar concentrations due to the degradation of organic matter (e.g., Rosenfeld, 1979; Boudreau and Canfield, 1988). It is also relevant in anoxic water columns such as the Saanich Inlet or the Black Sea where NH4 + levels are on the order of 100 μM (Brewer and Murray, 1973). Boatman and Murray (1982) derived a relationship between dissolved and adsorbed NH4 + as a function of clay mineralogy (Fig. 2), following standard formulations of adsorption isotherms (Stumm and Morgan, 1996). This relationship applies over a wide range of NH4 + concentrations, and it reproduces measured values in other settings (e.g., Rosenfeld, 1979; van Raaphorst and Malschaert, 1996). [It is preferred here over alternative formulations by Abdulgawad et al. (2009) because the experiments by Boatman and Murray (1982) were carried out under more controlled conditions.] Comparison to the Paleoarchean biotite data suggests that the water column or pore fluids from which the preceding clay minerals obtained their NH4 + must have had dissolved NH4 + concentrations between 0.2 and 10 mM (Fig. 2). Importantly, this is a minimum estimate, because some nitrogen may have been lost from the clay minerals during metamorphism. The calculated concentrations are well within the range of modern biologically controlled pore waters (Fig. 2). However, it is unknown whether similar concentrations could have been achieved on an abiotic planet or whether they are indeed a biosignature. This study seeks to address this question with a numerical model of nitrogen sources and sinks.
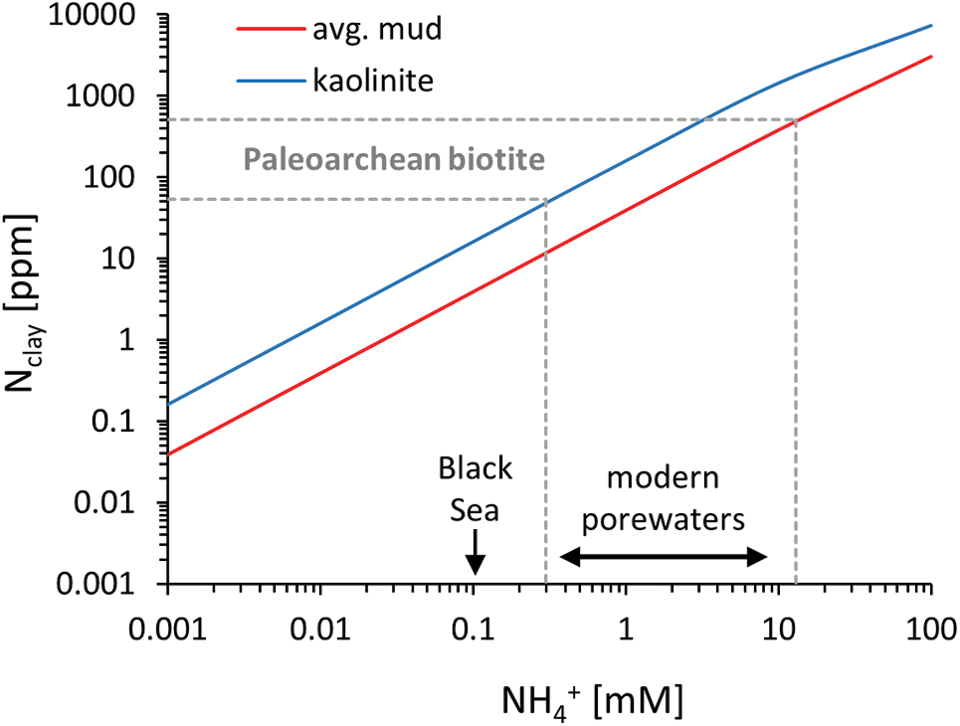
Adsorbed N content of clay as a function of dissolved NH4
+. The relationship is modeled after the work of Boatman and Murray (1982). Red = calibrated with average mud from the Saanich Inlet, after removal of organic matter, sulfides, and oxide minerals; blue = calibrated with pure kaolinite, which has the highest adsorption affinity from NH4
+ among clays. Horizontal gray lines = range of N concentrations in Paleoarchean biotite (Fig. 1). Maximum dissolved NH4
+ concentrations of modern pore waters are taken from the works of Rosenfeld (1979) and Boudreau and Canfield (1988). The Black Sea (Brewer and Murray, 1973) exemplifies a modern anoxic marine basin with largely biogenic NH4
+ sources. (Color graphics available at
2. Model Description
A simple one-box model was built with the isee Stella modeling software (Fig. 3). The initial NH4 + concentration of the ocean was set to zero; the model was run until the concentration had reached steady state. Source fluxes included hydrothermal input, impact shock heating, lightning, and volcanism. Hydrothermal N2 reduction to NH4 + was varied from 109 mol/yr to 1012 mol/yr (Smirnov et al., 2008), impact shock heating from 1010 mol/yr to 1011 mol/yr (Kasting, 1990), volcanism from 109 mol/yr to 1011 mol/yr (Mather et al., 2004), and lightning was set to 1010 mol/yr (Kasting and Walker, 1981; Navarro-González et al., 1998, 2001). Impacts, volcanism, and lightning produce NO x instead of NH4 +. It was assumed here that all NO x is reduced to NH4 + during hydrothermal circulation or by reaction with ferrous iron (Summers and Chang, 1993; Brandes et al., 1998). It is possible that some NO x was instead reduced to N2, but the relative proportions of NH4 + and N2 production are unknown. The assumption of complete conversion to NH4 + is thus conservative and provides an upper bound of the NH4 + source flux. The model was run with all fluxes set to either their maximum or their minimum value to explore the full range of total NH4 + source fluxes. Further discussion of the source fluxes is given below.
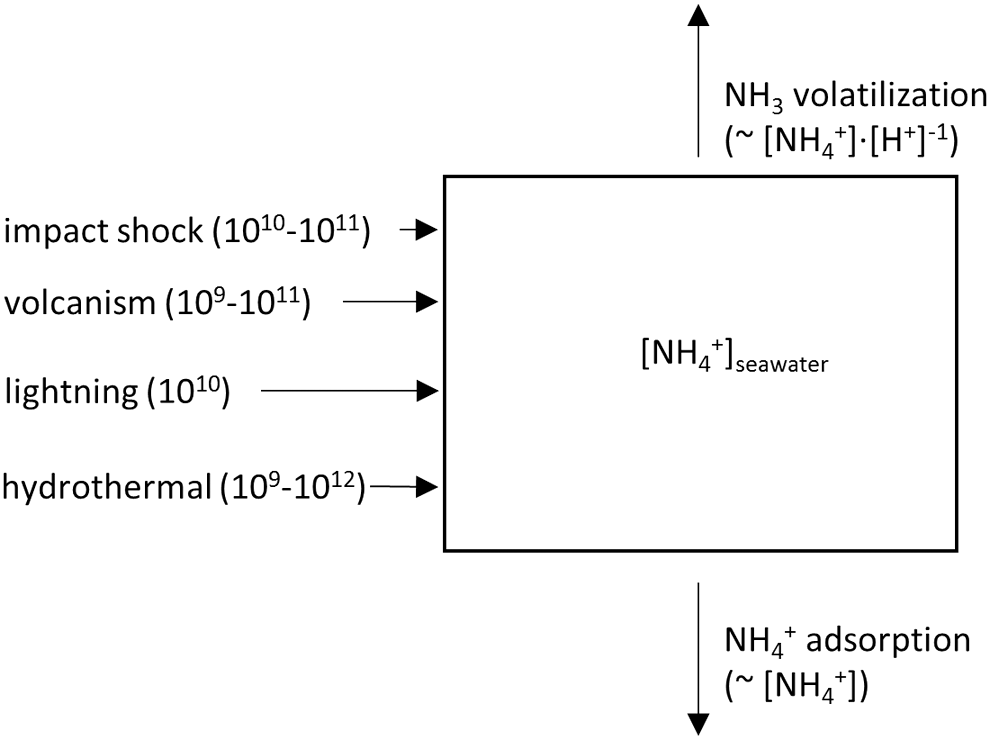
Schematic of the abiotic box model. Sources (in mol/yr) and sinks are described in the text. Minor source fluxes (≤109 mol/yr) such as extraterrestrial delivery or photochemical reactions are left out.
The two abiotic sinks in the model were adsorption of NH4 + onto settling clay particles and dissociation of NH4 + to NH3. NH3 is volatile and can escape into the atmosphere, where it is rapidly photolyzed back to N2 (Kuhn and Atreya, 1979). For clay adsorption, the settling flux was set to 5·1015 g/yr (Li, 2000, p 326, assuming seafloor area of 3.6 km2), which is in good agreement with average sedimentation rates of 4·1015 g/yr on continental shelves (Gregor, 1985). The model is fairly insensitive to this parameter above pH 6, where NH3 volatilization is the dominant N sink from the ocean. Here, an order-of-magnitude change in the clay flux changes the clay-bound N concentration by only a few percent. Below pH 6, a decrease in the clay flux has similarly small effects, whereas an increase leads to a significant drop in clay-bound N. If continental land masses were smaller in the early Archean, then the clay flux is more likely to have been smaller than today, meaning that the N concentration is overall not strongly affected by this parameter. The adsorption process itself was modeled after the work of Boatman and Murray (1982) as N clay = a NH4+/([1/K ads] + a NH4+) · Γ · M N (Fig. 2). Here, a NH4+ is the activity of NH4 +, that is, the molar concentration multiplied by an activity coefficient of 0.57 for seawater. M N is the molar mass of 14.01 g/mol for nitrogen. K ads is the adsorption constant, which is 5.1 L/mol for average mud from the Saanich Inlet and 21.0 L/mol for kaolinite (Boatman and Murray, 1982). Other clay minerals fall close to, or within, this range (montmorillonite 11.7 L/mol, illite 14.8 L/mol, chlorite 3.8 L/mol). The constant Γ is the cation exchange capacity of clay, which is taken to be 0.96 mmol/g for average surface mud (Boatman and Murray, 1982). Deeper mud samples and some individual clay minerals tend to have lower values for Γ (down to 0.65 mmol/g), making this a conservative estimate. The adsorption isotherm of Boatman and Murray (1982) has not been calibrated for low pH, but other studies have observed a decrease in NH4 + adsorption with decreasing pH (Kithome et al., 1999). This is expected, because mineral surfaces become progressively more positively charged with an increase of dissolved H+ ions (Stumm and Morgan, 1996), leading to increasing repulsion of the positively charged NH4 + ion under more acidic conditions. Keeping adsorption constant across pH is thus again a conservative approach. NH4 + adsorption has been shown to decrease with increasing salinity (Hou et al., 2003). The salinity of the Paleoarchean ocean is not well constrained but may have been 1.5–2 times higher than it is today (Knauth, 2005). Keeping salinity constant is therefore a conservative choice that may overestimate the abiotic NH4 + burial flux.
NH3 volatilization is a function of pH: (NH3) = [(NH4 +) · K NH3]/(H+), where K NH3 = 10−9.25 mol/L at standard conditions (Li et al., 2012). It is assumed that NH3 escape is limited by ocean circulation with an overturn rate of 1000 years as today. This is a conservative estimate, because NH3 may have escaped more rapidly to the atmosphere if it was produced in shallow water.
3. Results
The major results of the model are depicted in Fig. 4 and summarized in Table 2. The concentration of ammonium in clay minerals shows a strong dependence on pH, because under abiotic conditions, pH-dependent NH3 volatilization is a major sink of fixed nitrogen from the ocean. (As noted above, this is under the conservative assumption that NO x is mostly reduced to NH4 + instead of N2.) The higher the pH, the lower the concentration of dissolved NH4 + in seawater and the lower the abundance of adsorbed NH4 + in clay minerals. The results are also strongly dependent on the total source flux and the mineralogy of the clay mineral. Kaolinite with its higher adsorption constant (Boatman and Murray, 1982) tends to accumulate more nitrogen than other clay minerals. The original mineralogy of the clay minerals that preceded the Paleoarchean metasedimentary biotite grains is unknown. Overall, the dissolved NH4 + concentrations that are necessary to explain the nitrogen content of the biotite grains by purely abiotic mechanisms can only be achieved if the total source flux of fixed nitrogen was on the order of 1012 mol/yr or higher, or if the pH of the ocean was less than 6.
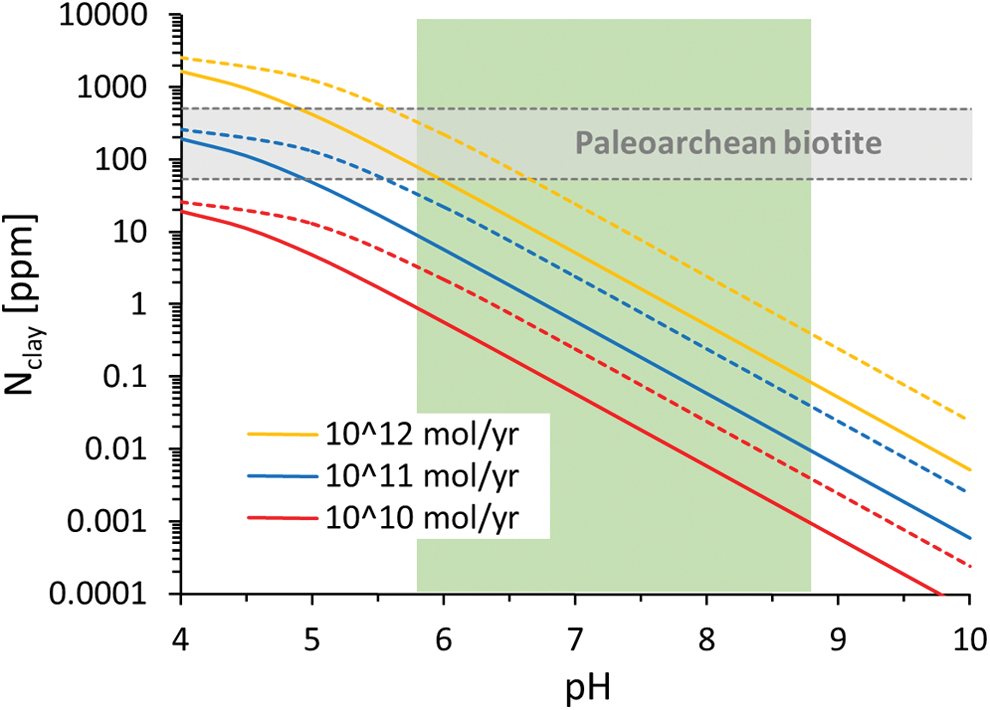
Modeled N contents of clay as a function of pH and total N source flux. The most realistic source flux is between 1010 mol/yr and 1011 mol/yr, based on a combination of volcanism, impact shock heating, and lightning (see text). Colored solid lines = average mud; colored dashed lines = kaolinite. Gray horizontal bar = N contents of Paleoarchean biotite (Fig. 1); green vertical bar = modeled pH range for the Archean ocean (Grotzinger and Kasting, 1993). (Color graphics available at
Data are for average mud from the Saanich Inlet (Boatman and Murray, 1982).
4. Discussion
The results suggest that the observed N concentrations are most probably of biogenic origin for a number of reasons. First, an abiotic source flux of >1011 mol/yr seems unlikely. High-temperature volcanism and impact shock heating of N2 and CO2 could potentially have maintained a fixed N flux of 1011 mol/yr (Kasting, 1990; Mather et al., 2004); however, estimates of up to 1012 mol/yr for hydrothermal reduction of N2 to NH4 + (Smirnov et al., 2008) seem implausible. Even modern hydrothermal fluids from unsedimented vents that do not recycle sedimentary nitrogen are noticeably NH4 + poor (<10 μM, von Damm, 1990; Lilley et al., 1993), despite the fact that the modern ocean is rich in NO3 −, which can also be reduced to NH4 + (Brandes et al., 1998). Given hydrothermal fluid flux rates of 0.5–6·1013 L/yr (Elderfield and Schultz, 1996; Emerson and Hedges, 2008), a hydrothermal NH4 + flux of more than 109 mol/yr from N2 reduction is unrealistic. Other studies of the hydrothermal NH4 + flux, based on iron-sulfide minerals instead of iron-nickel alloys as a catalyst, derived much lower values (108 mol/yr) (Schoonen and Xu, 2001). The low NH4 + concentration of <10 μM in hydrothermal fluids further rules out post-depositional alteration of the biotite grains (cf. van Zuilen et al., 2005). Nitrogen delivery by extraterrestrial material is thought to have been minor (103 to 109 mol/yr) (Chyba and Sagan, 1992; Smirnov et al., 2008). The same is true for photochemical HCN production in the absence of biogenic CH4 (109 mol/yr) (Tian et al., 2011). The total source flux, therefore, probably did not exceed 1011 mol/yr. Importantly, this limit would be even lower if a significant portion of NO x was reduced to N2 instead of NH4 +.
Second, assuming a total source flux of 1011 mol/yr or less, the model indicates that a pH of less than 5.5 would be necessary to explain the observed N enrichments in Paleoarchean biotite (Fig. 4). If the adsorption properties of the initial clay mineral were closer to that of average mud than to kaolinite, then a pH of less than 5 would be required. The pH of the Archean ocean is not well known, but model estimates by Grotzinger and Kasting (1993) suggest a range from roughly 5.8 to 8.8, depending on atmospheric pCO2. If the nitrogen were completely abiotic, then it would imply a more CO2-rich atmosphere than previously thought.
Lastly, the required concentrations of dissolved NH4 + are well within the range of modern anoxic pore waters (Fig. 2). They are thus plausibly explained by degradation of organic matter.
Isotopic data collected from Paleoarchean metasediments cannot unambiguously resolve this question, but they are consistent with a biological interpretation. Reported values range from −3‰ to +12‰ (Pinti et al., 2001; Nishizawa et al., 2005; Papineau et al., 2005; van Zuilen et al., 2005). Metamorphic overprint up to amphibolite facies has likely increased initial 15N/14N ratios through preferential 14N loss (Haendel et al., 1986). It is not possible to correct for the effect, but original δ15N values were likely lower than measured. The isotopic composition of abiotically fixed N is not well known, but measurements suggest that NO x produced by lightning may be isotopically light (−5‰ to −15‰, Ingerson, 1953; Moore, 1977), similar to photochemical reaction products (Kuga et al., 2014). The isotopic effects of volcanic and impact-induced nitrogen fixation have not been determined, but the reaction pathway and energy input are perhaps comparable to lightning; hence one may speculate that the product is isotopically light as well. Hence, if nitrogen was fixed abiotically, then dissolved NH4 + may have started out with a light δ15N value of <−5‰. Further fractionation may have occurred during NO x reduction to NH4 + (McCready et al., 1983), if the reduction did not go to completion. In the presence of life, a large proportion of this light NH4 + would likely have been assimilated into biomass. In a purely abiotic ocean, however, NH3 volatilization would have been a major sink of fixed nitrogen from the ocean, especially above pH 6, and this process would have imparted a large isotopic fractionation, rendering residual NH4 + heavy (ɛ = 45‰ at standard conditions, Li et al., 2012). It is conceivable that values from the heavier end of the observed spectrum in Paleoarchean metasediments reflect this process. However, none of the reported data points suggest fractionations up to 45‰. Instead, biological NH4 + uptake and burial, followed by metamorphic alteration to amphibolite facies, appears to be the more plausible mechanism. Biological N2 fixation cannot be ruled out from these data.
5. Conclusion
Given our currently best constraints for abiotic source fluxes of fixed nitrogen through volcanism, lightning, impacts, and hydrothermal activity, and current model constraints on the pH of the Archean ocean, the nitrogen contents of the Paleoarchean biotite cannot plausibly be explained by abiotic adsorption of NH4 + from seawater. Instead, it is more likely that NH4 + was enriched in sedimentary pore waters due to degradation of organic matter, as it is today. The nitrogen in these biotite separates is thus a biosignature and evidence of biological nitrogen utilization at 3.8 Gyr. Although the concentration and isotopic data cannot resolve by which metabolism nitrogen was assimilated into biomass, that is, by N2 fixation or by uptake of abiotic or biotic NH4 + or NO3 −, the inference of biological involvement supports previous advocates of a relatively early origin of life on Earth dating back to at least the Paleoarchean (Mojzsis et al., 1996; Rosing, 1999). This study further shows that nitrogen abundances can potentially serve as a useful biosignature on extraterrestrial missions, if source fluxes and solution pH can be estimated independently. For example, this proxy could prove useful in upcoming investigations of sediments on Mars.
Footnotes
Acknowledgments
Financial support was provided by the NASA postdoctoral fellowship program. I further thank Sherry Cady, Jim Kasting, and Christophe Thomazo for constructive comments.