
Abstract
We tested the potential of a miniaturized Raman spectrometer for use in field detection of snow algae pigments. A miniature Raman spectrometer, equipped with an excitation laser at 532 nm, allowed for the detection of carotenoids in cells of Chloromonas nivalis and Chlamydomonas nivalis at different stages of their life cycle. Astaxanthin, the major photoprotective pigment, was detected in algal blooms originating in snows at two alpine European sites that differed in altitude (Krkonoše Mts., Czech Republic, 1502 m a.s.l., and Ötztal Alps, Austria, 2790 m a.s.l.). Comparison is made with a common microalga exclusively producing astaxanthin (Haematococcus pluvialis). The handheld Raman spectrometer is a useful tool for fast and direct field estimations of the presence of carotenoids (mainly astaxanthin) within blooms of snow algae. Application of miniature Raman instruments as well as flight prototypes in areas where microbes are surviving under extreme conditions is an important stage in preparation for successful deployment of this kind of instrumentation in the framework of forthcoming astrobiological missions to Mars. Key Words: Snow algae—Chloromonas nivalis—Chlamydomonas nivalis—On-site field detection—Raman spectroscopy—Astaxanthin. Astrobiology 16, 913–924.
1. Introduction
E
Mars and some icy moons, especially Europa, are well known as bodies in the Solar System that have attracted the attention of astrobiologists over the decades. The presence of water on Mars in solid form today, and even at the surface (Martinez and Renno, 2013), as well as in liquid form in the past (Westall et al., 2013; Dohm et al., 2015), is an important general aspect and major reason for this extremely high level of interest of astrobiologists. Osterloo et al. (2008) demonstrated that chlorides are globally present on Mars and pointed to their importance as signs of the previous presence of water bodies. Additionally, it has been suggested that the chaotropic salts detected on Mars may also favor the survival of psychrophilic microorganisms at very low temperatures (Chin et al., 2010). Fischer et al. (2014) confirmed regions where salts and ice coexist at the martian surface or subsurface. Formation of liquid brines might occur and could allow microbial growth to thrive, even on the surface of the planet. Hynek et al. (2015) proposed time sequences for the formation of superficial ponds in the Meridiani Planum region. Dohm et al. (2015) demonstrated the major stages of past water movements in the Argyre province and its surroundings. On the basis of thorough analyses of both the surface features and local sedimentology, they documented complex water migration processes such as groundwater discharges into an ice-covered lake, spring-fed activity, catastrophic outburst floods, debris flows, and alluvial fan development, as well as glacier accumulation and inflow into the basin. Surface martian physical and physicochemical conditions, as well as up-to-date knowledge of martian geology, have been described to such a detailed level as to allow the suggestion of potential local developments of habitable zones (Cockell and Raven, 2004). Those zones are not located at the surface, where harsh radiation is expected to prohibit the evolution of life (Thomas and Schimel, 1991; Clark, 1998, Ronto et al., 2003) or even conservation of its vestiges. Deeper zones are of interest to astrobiologists, and the existence of ice or liquid water bodies within the martian subsurface in the past (or even currently) has been suggested (Cabrol and Grin, 2001; Baker et al., 2005). By inhabiting a subsurface environment (i.e., soil, ice, or snow), the substrate would attenuate harmful UV radiation, and DNA-based photosynthetic organisms may be able to find a habitable niche within it (Cockell and Raven, 2004). France et al. (2010) showed that dusty water-ice snowpacks on Mars may also provide a habitable zone even for DNA-based photosynthetic life. Habitability was recently reviewed by Cockell et al. (2016).
On Europa, the presence of a liquid ocean beneath the icy crust is currently being examined in greater detail and represents an important area for potential future astrobiological research (Gaidos et al., 1999; Lipps and Rieboldt, 2005; Greenberg, 2011). Hypotheses and research focused on the possibility of a sustaining biosphere in the past, or even today, have been reported (Reynolds et al., 1983; Jakosky and Shock, 1998; Priscu et al., 1998; Spencer et al., 1999; Price 2007; Wilhelm et al., 2012; Noell et al., 2015). General strategies for searching for life on Europa have been suggested and have included searches for biomarkers and biochemical testing of materials (i.e., antibody microarray immunoassays) (Prieto-Ballesteros et al., 2011).
On Earth, studies in the field of the development of psychrophiles' ecosystems, as well as other snow- and ice-tolerant environments, are ideal for understanding the survival and protection strategies of such organisms (Gleeson et al., 2012). Detecting them and describing them under complex situations on-site can also be both useful tasks and exercises for training in the search for life beyond Earth (i.e., Preston and Dartnell, 2014).
1.1. Snow algae and life under low temperatures
Colored snow is a striking natural phenomenon caused by the mass accumulation of snow algae. They represent a specific and well-adapted group of microorganisms, which have developed physiologically optimized to such an extreme environment. The snow environment is characterized by low temperature, high and variable irradiance (photosynthetically active radiation and UV radiation), freeze-thaw cycles, nutrient depletion, and so on. The physiological and biochemical processes of snow algae are optimized to their extreme environment (Hoham and Duval, 2001). The major protection against high irradiance levels is accomplished by the accumulation of secondary carotenoids, specifically astaxanthin. Large amounts of free and esterified astaxanthin are located in cytoplasmic lipid globules, which often occupy most of the cell volume. Secondary carotenoids shield the chloroplast from high irradiation (reducing the risk of photoinhibition) and may also be a potential carbon source during unfavorable environmental conditions (Remias et al., 2005). Optimum temperatures for growth of snow algae are, for the most part, shifted to 1–10°C, and the great extent of fatty acid unsaturation of membrane lipids plays a major role in avoiding membrane rigidity at low temperatures (Morgan-Kiss et al., 2006). The esterification of astaxanthin with unsaturated fatty acids can also provide cryoprotection (Řezanka et al., 2008, 2013). Most snow algae belong to the green algal order Chlamydomonadales (Chlorophyta), and the genera Chloromonas and Chlamydomonas are widely distributed. The life history, involving flagellated vegetative cells and immotile resting stages, is a successful adaptation to the extreme environment of mountain or polar snowfields. The species Chlamydomonas nivalis forming the “red snow phenomenon” is well known from alpine localities on all continents, as well as from the polar regions, and is regarded as a cosmopolitan cryophilic species (Remias et al., 2005). Chloromonas nivalis has been reported as a species causing green, orange, or pink blooms in alpine and polar regions worldwide. The ecology of snow algae in the Giant (Krkonoše) Mountains, Czech Republic, has been described in detail (Nedbalová et al., 2008). In contrast to Chlamydomonas nivalis, this taxon is less frequently found, and the flagellates are always green (Remias et al., 2010).
1.2. Raman spectroscopy for the search for life signatures in extraterrestrial scenarios
Several different strategies for the search for life on Mars have been suggested and developed by NASA and ESA in the framework of the coming astrobiology-dedicated missions. The recommended protocols and methods have been detailed (e.g., Des Marais et al., 2008; Shapiro and Schulze-Makuch, 2009; Prieto-Ballesteros et al., 2011) and are being tested within the framework of terrestrial scenarios and analogues (i.e., as reviewed by Preston and Dartnell, 2014). Strategies proposed are based on the deployment of miniature Raman spectrometers in the frame of Mars rovers under martian conditions as suggested first by Dickensheets et al. (2000) and Ellery and Wynn-Williams (2003). This proposal was developed since and tested due to the technical and analytical progress that allows reliable recording of Raman spectra with miniature instruments (or even Mars Raman spectrometer prototypes) under outdoor conditions (see below). During the last decade, in fact, transportable and portable Raman spectrometers have been developed and are at present used commonly in very different areas, for example, in forensics, the geosciences, and cultural heritage domains (as reviewed for example by Colomban, 2012; Vandenabeele et al., 2014).
The European Space Agency (ESA) has proposed that a miniaturized Raman spectrometer be part of the Pasteur analytical life-detection protocol in the ExoMars mission, which is scheduled to launch in 2018 with the intent to search for signs of life on Mars in the Aurora program. Raman spectroscopy will perform a key role in the molecular analytical protocols aboard the ExoMars rover. Several different prototypes of miniature Raman spectrometers have been designed; however, a final decision was made for a 532 nm excitation laser line (Rull et al., 2011). It has been suggested that preliminary testing of the instrumentation should be performed within the framework of laboratory conditions. Additionally, field testing under common or even complex outdoor conditions has been recommended (Edwards et al., 2014). A laboratory model of the ExoMars Raman Laser Spectrometer (RLS) device was developed at the Associated Unit University of Valladolid–CSIC Center of Astrobiology (UVa-CAB) (Rull et al., 2011). The Raman prototype (532 nm) has an X, Y, Z coordinate positioning system, with a precision of 2.5 μm, as well as imaging, autofocus, and spectral acquisition capabilities. The analytical strategy of the RLS instrument is the acquisition of spectra along a linear series of points. At each point, the optical head is focused on the sample, the acquisition parameters are automatically calculated, and the spectrum is acquired. The ExoMars RLS prototype developed at the University of Leicester is another miniaturized prototype (Edwards et al., 2012). This system consists of a continuous-wave diode laser (100 mW at 532 nm), a detector system that incorporates thermoelectrically cooled CCD and drive/acquisition electronics, and a transmission grating with two spectral orders.
The advantage of the Raman spectroscopic investigations in situ, or remotely under martian conditions, is the possibility for acquisition of information on minerals or organics (biomarkers) (Edwards et al., 2014; Harris et al., 2015). Obtaining Raman spectra by using miniature devices currently planned to be installed (i.e., aboard the ESA rover for ExoMars) was identified as a key step in the search for biomarkers and minerals in the martian subsurface. Identifying biomarkers allows for gathering information about extinct or extant life; knowledge about mineral distributions can be seen as a key to estimating the environmental conditions in the past.
Raman spectroscopy has been evaluated many times within the fields of geobiology and astrobiology, with the goal of detecting key biomarkers of different organisms: pigments of lichens (Edwards et al., 2003), carotenoids of corals (de Oliveira et al., 2010), carotenoids of halophiles (Jehlička et al., 2013a), or osmotic solutes of halophiles (Jehlička et al., 2012). Carotenoids have been detected with Raman spectroscopy as a group of organic traces under different scenarios and matrices, and proposed as biomarkers (Edwards et al., 2014). Blue and green lasers (485, 514, 532 nm) are well adapted for obtaining sharp resonance Raman signals of polyenes. Due to the resonance Raman effects and resonance enhancement of Raman signals of carotenoids, the detection limits can be very low. The search for these molecules in extraterrestrial environments, including the martian subsurface, is seen as a major task for astrobiological investigations.
The limits, potential pitfalls, as well as advantages of such techniques for the detection of pigments of microorganisms have also been highlighted (Jehlička et al., 2014a; Harris et al., 2015). Pigments from a pure biomass, originating from cultivations of archaea, cyanobacteria, or microalgae, have been detected (Rosch et al., 2005; Marshall et al., 2006, 2007; Harz et al., 2009; Jehlička et al., 2013b, 2014b), as well as pigments from inclusions within crystalline halite (Winters et al., 2013). Details of the distribution of microbes in the pattern of endolithic communities from the Atacama were successfully obtained by using conventional Raman spectroscopy and a Raman imaging approach (Vítek et al., 2013, 2014). Culka et al. (2012, 2014a, 2014b), Jehlička et al., (2009, 2010, 2011), and Jehlička and Oren (2013) additionally described how miniature Raman spectrometers and Mars prototypes perform to detect minerals, biomarkers of halophiles, endolithic colonizations, and carbon compounds having possible relevance in Mars studies. Additionally, small, light, and portable spectrometers can be applied directly under field conditions, which is not feasible with other analytical techniques. Raman spectroscopy has been evaluated for the detection of compounds at low temperatures, and miniaturized Raman spectrometers with 785 nm lasers have been used to investigate mixtures of amino acids in mineral matrices at about −10°C (Culka et al., 2011) and complex mixtures of amino acids in water ice even in small quantities (Culka et al., 2014c).
Carotenoids have been detected by Raman spectroscopy as a group of organic traces under different scenarios and matrices, and have been proposed as biomarkers (Edwards et al., 2014). The search for these molecules in extraterrestrial environments, including the martian subsurface, is seen as a major task for astrobiological investigations. Importantly, mainly due to the above-mentioned resonance enhancement of the Raman signals of the polyenic chain, portable instrumentation (green excitation) has performed well in carotenoid detection outdoors (Prinsloo et al., 2008; Tournié et al., 2011; Jehlička and Oren, 2013). Due to the resonance Raman effects and resonance enhancement of Raman signals of carotenoids, the detection limits can be very low.
In the present study, icy environments were investigated with the intent to estimate the performance of lightweight miniature portable Raman spectrometers in the detection of pigments of snow algae. Specific carotenoids are expected to be produced by these microorganisms in conjunction with environmental conditions. Is Raman spectroscopy the ideal tool for discriminating these pigments? Is it possible for miniature instruments to be applied under field conditions for this kind of task?
2. Materials and Methods
2.1. Description of sites
Measurements were performed at two sites above the timberline on two different European mountains. The snowfields available at the time were intensely colored due to massive occurrences of snow algae from the order Chlamydomonadales (Chlorophyta) (Table 1). In the Krkonoše Mountains, the algal communities were most densely developed at the edges and lower parts of the snowfields. This distribution is probably due to the flow of the thawing snow, but the availability of nutrients from animal excrement surrounding the snowfields may also be important (Kvíderová and Kociánová, 2011). The snow algae formed patches of a distinct dark orange-pink color on the surface of the snow (Fig. 1A, 1C); additionally, yellow-green colored colonies were found in the soft snow below the surface (Fig. 1E).

(
In the Ötztal Alps, dark red-violet colored patches in depressions on the snow surface were observed on the remaining and quickly thawing snowfields (Fig. 1B). Algal biomasses accumulated at the edges of the snow and rocks due to thawing and rain (Fig. 1D, 1F).
In addition, two other members of the common green algae were analyzed, with the values of these Raman bands reported in Table 2 for comparison purposes. While Haematoccoccus pluvialis is known to produce very large amounts of astaxanthin, Trentepohlia produces mainly β-carotene. Haematoccoccus pluvialis colonies were investigated directly in a shallow cistern on the grounds of the Třeboň Institute of Botany of the Academy of Science of the Czech Republic on August 10, 2015. The Raman spectra of Trentepohlia were obtained from the surface of a gneissic boulder in the Labe streambed, about 1 km downstream from the spring on May 20, 2015. Both samples were analyzed in situ with a Rigaku handheld instrument.
Expected major carotenoid reported.
Handheld instrument (H), laboratory instrument (L); both using 532 nm excitation.
2.2. Sampling
At both sites, the snow algae were collected in sterilized plastic vials from the colored layer and then transported alive in a thermos bottle to the laboratory. A fraction of the samples were frozen for further study. Live samples were examined and photographed in the laboratory as soon as possible using a Nikon Eclipse E400 microscope (magnification 1000×) with a Canon EOS 650DX camera system.
2.3. Raman spectrometric analyses
The primary Raman spectrometer used for analyses of snow algae in situ was a handheld Rigaku First Guard Raman spectrometer (Rigaku Raman Technologies, Burlington, MA, USA). This instrument (Fig. 2) weighs 2.7 kg and is equipped with a 532 nm diode laser and thermoelectrically cooled CCD detector. For this instrument, the laser power at the target for all the analyses reported was around 20 mW (as measured with the LaserCheck handheld laser power meter from Coherent Inc.), and the spectral range was 200–3000 cm−1, with a spectral resolution of 10–15 cm−1. The instrument uses a resistive touch display, which allows for a range of experimental settings such as the number and duration of scans to be collected.

Recording Raman spectra of the reddish zones on the surface of thawing snow at the Rettenbach Glacier.
In the present study, a short measurement was typically tested. When the positioning was optimal, a longer measurement for the spectrum with a better signal-to-noise ratio was then executed. When performing the analysis, the head of the instrument was in direct contact with the sample (algae dispersed on/in snow, or accumulated algal biomass). During acquisition of spectra, the instrument head and the surroundings of the point being analyzed were covered with a black cloth to shield from sunlight. In the case of field measurements, 30 accumulations of a 1 s shot were acquired.
For comparative purposes, all the samples were also analyzed with a laboratory Raman microspectrometer. A Thermo Scientific DXR Raman microscope interfaced to an Olympus microscope (with a 50× long focus objective) was used. A laser with the same wavelength (532 nm) as the handheld instrument was selected. The laser power at the target for the laboratory Raman microspectrometric measurements was roughly 0.1 mW, and the spectra were recorded using a high-resolution grating from 100 to 2000 cm−1. A total of 64 accumulations of a 1 s shot were optimal in the case of Raman microspectrometry to achieve a very good signal-to-noise ratio. Laser spot size at the sample for the handheld instrument was roughly 1 mm in diameter when the optical head was at the contact with the target. For the laboratory instrument, the spot size was roughly 2 μm in diameter when focusing at the surface.
Under the microscope, it was possible to analyze single algal cells, in either the vegetative or immotile resting stages.
The spectra from both instruments were exported in .spc format and corrected for baseline with GRAMS/AI 9.1 spectroscopy software. For uniform results, the position of the Raman bands was determined by the peak fitting procedure.
3. Results
3.1. Snow algae
At the Luční hora site (see Table 1, Fig. 1A), several colored patches of the snow algae Chloromonas nivalis (Fig. 1C) were located on two snowfields; the density of individual patches varied significantly. In a suitable green-yellowish spot of snow cover, a profile (ca. 15 cm thick) was cut in the slowly melting snowfield (Fig. 1E). At the bottom of the profile, the snow was compacted to form ice covered with melting water. Both the orange-pink surface layer of snow and the yellow-green layer (ca. 10 cm thick) of snow were analyzed with the handheld instrument. The laser beam from the probe head was focused directly onto the colored zones, in the contact mode.
The Raman signal strength arising from the presence of the carotenoid pigments varied significantly between the two layers. Spectra acquired on the top orange-pink layer (see Fig. 3A; the reference spectrum of the same sample taken by the laboratory Raman is in Fig. 3B) were of a very good quality and with a favorable signal-to-noise ratio. The positions of the three main carotenoid bands are 1518, 1157, and 1006 cm−1, which can be attributed to the ν1(C═C), ν1(C–C), and ρ(C–CH3), respectively. Generally, the positions of the Raman bands in the spectra from the laboratory instrument were almost identical: 1518, 1157, and 1006 cm−1. At least 10 measurements were performed on different spots to evaluate potential nonsystematic shifts in the wavenumber position of the bands (which can sometimes occur when using handheld instruments). The values reported here are median values; standard deviation ranged from 5 to 3 cm−1 when using the handheld instrument, with respect to dispersive laboratory instruments.
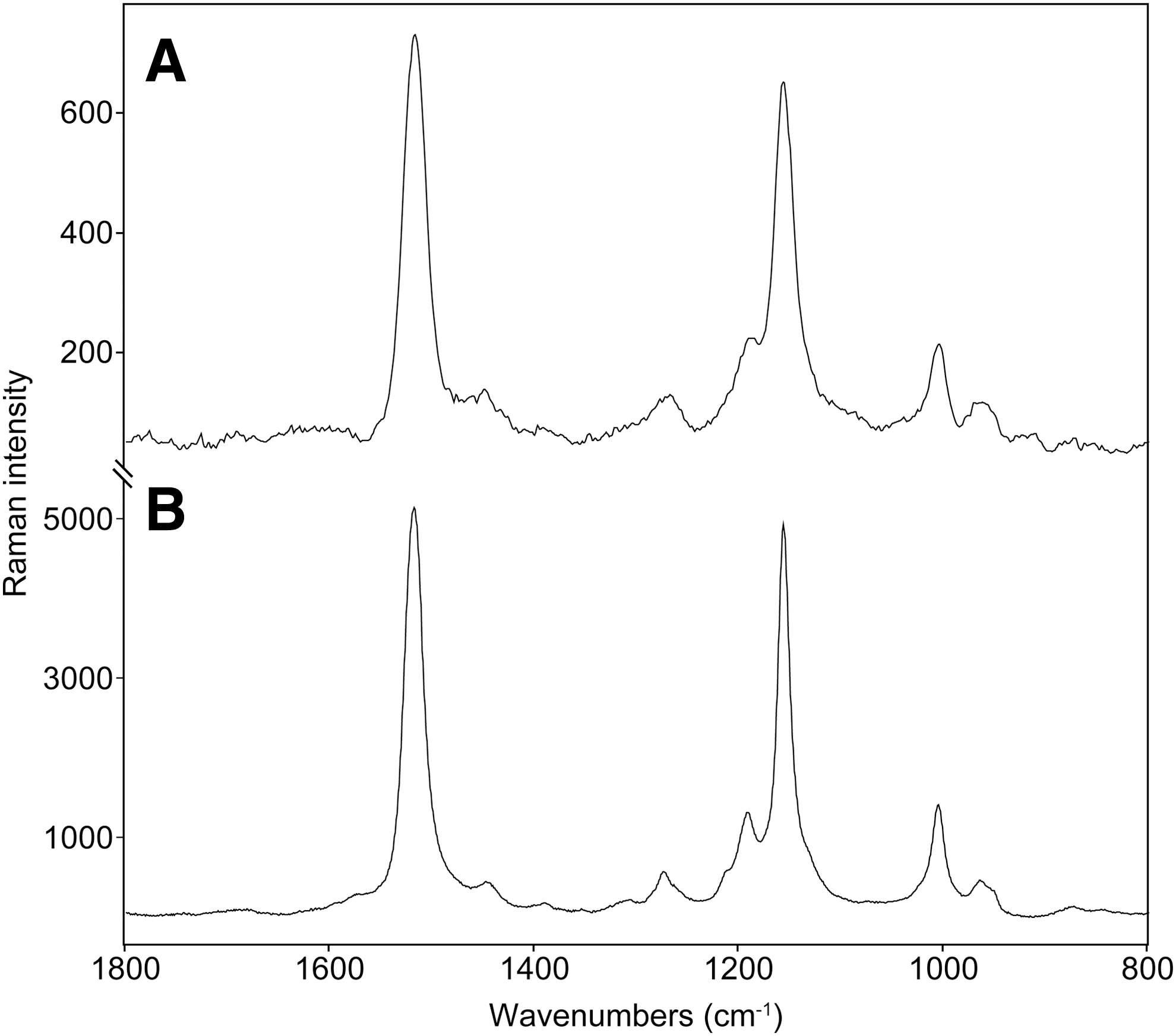
Representative Raman spectra taken (
The spectra that were taken on the green layer by the handheld instrument were characterized by a high level of noise, and quite a low-intensity Raman signal, due to the significantly lower presence of carotenoid pigments (see Fig. 4A). Additionally, a small shift toward higher wavenumbers was detected in the position of the ν1(C═C) Raman band. Raman bands were observed at 1521, 1156, and 1005 cm−1. The Raman spectra also manifest a relatively high level of noise, and the carotenoid Raman band occurs at 1522, 1156, and 1004 cm−1. The shift (between orange-pink and yellow-green Chloromonas nivalis) in the position of the ν1(C═C) Raman band is consistent with the expected change in the ratios of the different occurring carotenoid pigments, such as astaxanthin and β-carotene. This will be discussed later.
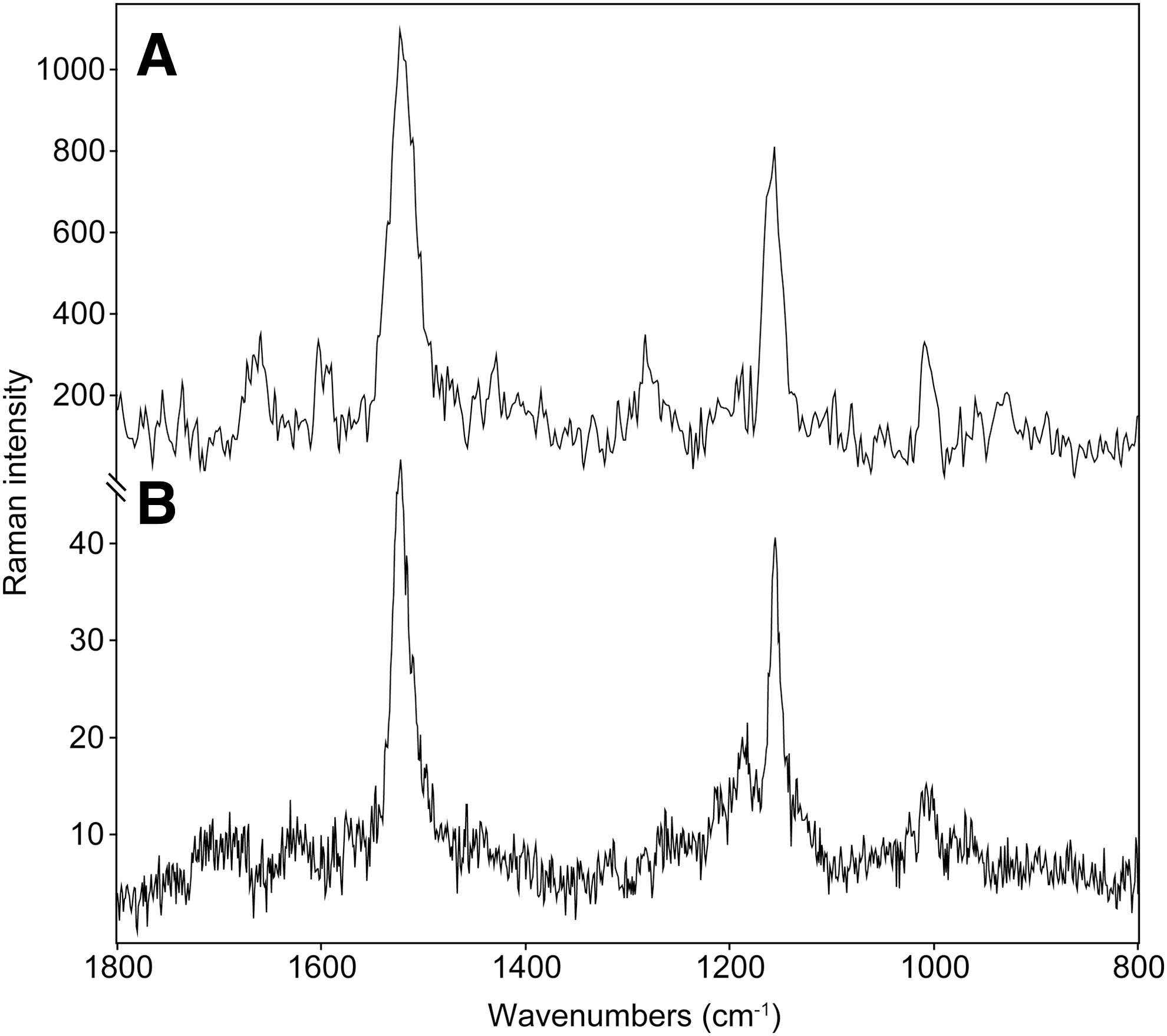
Raman spectra taken (
At the Rettenbach Glacier site (see Figs. 1B and 2), patches of “red snow” were distributed to a greater extent as well as with a greater density of the algal colonizations (based on the color of the snow). The overall amount of accumulated algal biomass on the surface of the snowfields was much greater than at the Luční hora site. Prominent red-violet colored patches, with algal biomass accumulations within depressions on the surface of snow, suggested massive thawing and rain activity. Allochtonous algal accumulations occurred at the rock/snow boundaries as well (see Fig. 1F). The algae were later microscopically identified as Chlamydomonas cf. nivalis in a resting stage (see Fig. 1I). Thus, the best-quality spectra taken in this study of snow algae by a handheld instrument (see Fig. 5A) were recorded at this site. The three main carotenoid Raman bands were located at 1516, 1154, and 1005 cm−1 in the spectra that were taken by the handheld instrument in situ, compared to the laboratory reference values of 1517, 1156, and 1006 cm−1. These values can be seen as indicative of the presence of astaxanthin. Snow algae transported from the snowpack and thawing ice accumulated on stones and at the stone/snow interface. In this case, it was even possible to analyze the accumulated algal biomass directly on the surface of stones, and the positions of the Raman bands were the same when compared to the values obtained directly on the snow surface.
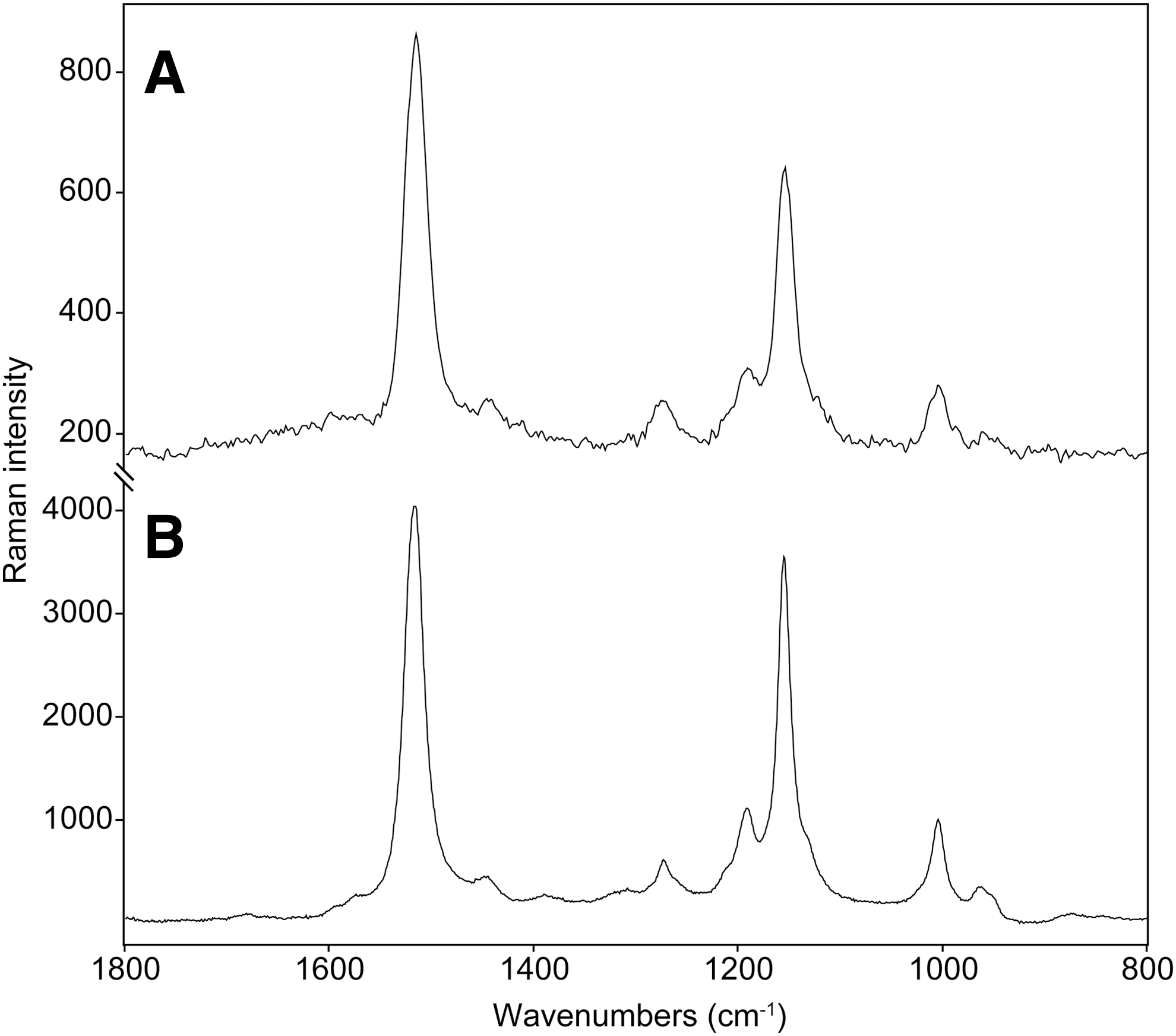
Raman spectra taken (
All the Raman band positions in the text and tables were determined by the peak fitting procedure, using one band for fitting each of the three main carotenoid bands. However, for some of the spectra (from yellowish/orange colored zones with a low concentration of astaxanthin), a better fit for the diagnostic Raman band, located at around 1522 cm−1, was achieved by using two bands. The band positions for the two resulting Raman bands were typically very close to 1524 and 1516 cm−1, which is in very good agreement with the values in our samples that we believe are the values indicative for β-carotene and astaxanthin, respectively. This illustrates the fact that this band is actually a compound band, which is due to the signal of all carotenoid pigments present in the sample. In some situations, when using high-quality spectra, a deconvolution (peak fitting) along with the band position can be used for a tentative characterization of the mixture of carotenoids.
4. Discussion
The detection of snow algae pigments in situ on snowfields while using a handheld Raman spectrometer was expected to be a challenge. From our previous experience with handheld instruments, difficulties such as partial melting of samples and/or low concentrations of target molecules were anticipated. A unique aspect of this type of snow coloration is that the studied system contained microorganisms that were highly mobile in the vegetative state. For example, in the vertical profile of the snowfield at Luční hora, the yellow-green Chloromonas nivalis cells were highly mobile and escaped into the interior of snow mass on the order of minutes. This was definitely active movement, caused by the flagellated algae cells, as opposed to the moderate flow of water caused by the melting of ice by the laser. The flagellated stages occur in all snow algae from the order Chlamydomonadales and apparently represent an important adaptation to life in the snow, enabling them to find optimal conditions (Hoham and Duval, 2001). Nevertheless, it was possible to acquire a good-quality Raman spectra of the carotenoid pigments even when the concentration of algae in the snow was low and when only the yellow-green Chloromonas nivalis cells (i.e., flagellated cells, which contain minimal amount of the secondary, photoprotective pigments) were present. This was primarily feasible because a 532 nm laser excitation was used, which allowed the enhancement of the Raman signal of the carotenoids due to the resonance effect. On the other hand, it was easy to acquire very-good-quality Raman spectra of the carotenoids in the samples of orange-pink Chloromonas nivalis immotile resting stages on the surface of snowfields, which contained high amounts of secondary carotenoid pigments, especially astaxanthin. The physiological changes of Chloromonas nivalis cells during cyst formation were described in detail by Remias et al. (2010). The content of astaxanthin, indicated by the ratio to chlorophyll, significantly increased in mature cysts in comparison with younger stages. Other secondary carotenoids like canthaxanthin or echinenone were not detected in this species.
A detailed study involving determination of the ratios of carotenoid pigments in the different life cycle stages of Chloromonas nivalis, and how these changes are reflected in the Raman spectra, is a work in progress; however, this exceeds the scope of the present study.
The Raman spectra taken on Chlamydomonas nivalis generally had the highest signal-to-noise ratio, as well as the highest absolute Raman intensity of the three major carotenoid bands. The Raman spectra taken on the orange-pink Chloromonas nivalis generally showed a lower Raman intensity of the carotenoid bands, but the signal-to-noise was quite high. In the case of the Raman spectra taken on the yellow-green Chloromonas nivalis, the Raman signal from carotenoid pigments was almost 2 orders of magnitude lower, with the level of noise substantial, indicating an almost complete absence of the secondary carotenoid pigment astaxanthin. This reflects the fact that the content of astaxanthin in the resting stages of Chlamydomonas nivalis is generally much higher in comparison with previous species. In a study from the Austrian Alps, Remias et al. (2005) reported values up to 25 for the astaxanthin to chlorophyll a ratio in field samples. To our knowledge, these values are among the highest reported for snow algae.
The results of this study suggest that some changes in the presence of individual carotenoid pigments in snow algae can be documented by Raman spectra acquired by handheld instruments.
Table 2 summarizes the positions of the major carotenoid bands from the Raman spectra taken on snow algae. For comparison, the carotenoid Raman bands from spectra taken on other members of the green algae group (not shown here) such as Haematoccoccus pluvialis (with dominant astaxanthin) and Trentepohlia (dominant β-carotene) are listed.
The position of the Raman band due to the ν1(C═C) is diagnostic of the length of the polyene chain, and this position shifts from 1522–1521 cm−1 (Chloromonas nivalis vegetative cells containing β-carotene and minimal amounts of astaxanthin) to about 1518 cm−1 (Chloromonas nivalis cysts with accumulated astaxanthin). Even lower values (1516–1517 cm−1) were detected in the spectra of Chlamydomonas nivalis cysts. Similar low wavenumber values were recorded in the spectra of Haematoccoccus pluvialis cysts, which accumulate significant amounts of astaxanthin in the form of astaxanthin esters that can amount to up to 4% of the dry weight (Boussiba et al., 1999). An increase in the concentration of the carotenoid pigments (vegetative cells versus cysts) can only be roughly estimated based on the relative intensities of the Raman bands of the carotenoids. This was attempted using the laboratory Raman instrument in the analysis of a single cell; the carotenoid Raman signals from the cysts were typically 50–150× stronger due to the presence of astaxanthin.
Kaczor et al. (2011) used Raman microimaging of astaxanthin in H. pluvialis cells to obtain information about the distribution, concentration, and molecular structure of the studied object without simultaneous degradation of the sample. They also showed that in situ and in vivo analysis of a single cell can be conducted when the compound (or compounds) of interest has a large Raman cross-section. They found that the distribution and concentration of astaxanthin in the cysts of H. pluvialis differ significantly, depending on the morphological form of the cell; however, the molecular structure of astaxanthin seems to be uniform. Several studies that focused on carotenoids and their analyses using Raman spectroscopy have shown differences in the spectra of a given compound in different organisms and in different chemical environments. Modifications in band positions can reflect not only the length of a polyenic chain but also the mode of binding of such a molecule in the context of the biological tissue (de Oliveira et al., 2010; Maia et al., 2013). Differences in the band positions of the Raman spectra of carotenoids, between different organisms under optimal conditions, can allow for an estimation of the length of the polyenic chain and a precise discrimination of the given carotenoid (Withnall et al., 2003; Jehlička and Oren, 2013). However, in many cases, the length of the polyenic chain of carotenoids is the same (or very similar), and the unambiguous discrimination of unknown carotenoids can be tricky or even impossible (Jehlička et al., 2014a; Harris et al., 2015). Differences observed between spectra obtained when investigating reference crystalline solid astaxanthin and Raman spectra obtained on biomasses are related to the precise occurrence of the pigment in a molecule or chemical environment. The change of the astaxanthin molecular structure in microalgae compared to the standard is probably related to the change of the conformation between the ionone rings and the chain, as the Raman shifts of the key bands depend on the dihedral angles defining this conformation (Kaczor et al., 2011). Additionally, effects of the potential presence of the astaxanthin monoester can also be expected (in the same manner as in the case of H. pluvialis; Renstrøm et al., 1981); however, the spectral resolution and stability of these miniature systems do not allow one to ascertain this possibility.
The present study shows Raman spectroscopic results obtained on microbial communities in the frame of snow. Direct mode of illuminating targets and quasi contact collecting of scattered radiation without any preliminary treatment was selected. This mode of analysis differs from the protocol proposed for the next ExoMars mission (2020, ESA) (Rull et al., 2011, 2013; Foucher et al., 2013). In this case, rocky material will be obtained from depths from 0.5 to 2 m by drilling and will be crushed for subsequent analysis by a suite of instruments on board the rover, including the Raman spectrometer. It is also different from the mode suggested and tested that will be applied in the framework of forthcoming planetary missions by American teams, which will involve the deployment of a stand-off Raman spectrometer that allows for remote acquisition of the spectra of minerals and organics (Sharma et al., 2006, 2010). In this case, a pulse-gated spectrometer can use a 532 nm excitation (Clegg et al., 2014). An alternative, which has been investigated at the Jet Propulsion Laboratory (NASA), is an in situ Raman spectrometer that employs a continuous-wave excitation laser emitting at 248.6 nm (SHERLOC) (Beegle et al., 2014).
Advantages and critical issues of each of the protocols are thoroughly evaluated. For the successful realization of the planned missions, with respect to technical limitations, it is clear that it will be necessary to accept some compromises. This concern, for example, limits acquisition of acceptable Raman spectra of some biomolecules when using green lasers (conversely to the excellent possibility to detect carotenoids or elemental carbon in this case). A drawback can be seen by studying powdered samples prepared by grinding (ExoMars). Here, part of the spectroscopic and geochemical information of relevance (spatial context, relationship between a potential biomolecule and mineral matrix) will probably be lost. Critical issues of Raman spectroscopic investigations also include presumed general instability of biomolecules under laser illumination and potential degradation of the targets when using higher laser energy. Those aspects can well be addressed in the framework of studies of terrestrial analogues (including this study). Testing on Earth can clearly be seen as an important step for further preparing planetary missions.
5. Conclusions
We evaluated the potential of a miniaturized Raman spectrometer with a 532 nm laser for field detection of pigments of snow algae. The instrument allowed for the unambiguous detection of carotenoids in blooms of Chloromonas nivalis and Chlamydomonas nivalis (Chlamydomonadales, Chlorophyta) at different life-cycle stages. Astaxanthin, the major pigment, was detected in colored patches that had developed in snow at different depths and at two alpine sites (Krkonoše Mts., Czech Republic, 1200 m, and Rettenbach Glacier, Austria, 2790 m). A rough estimation of the ratio between astaxanthin and β-carotene can be carried out when using conventional Raman microspectrometers under laboratory conditions. The application of miniature Raman instruments as well as flight prototypes in areas of microbial survival under extreme conditions is an important stage to prepare for the successful deployment of this kind of instrumentation in the context of forthcoming astrobiological missions to Mars.
Footnotes
Acknowledgments
This work was partly funded by a grant of the Grant Agency of the Czech Republic (P210/10/0467) and by institutional support. The authors wish to thank Milena Kociánová for her assistance and guidance in the Krkonoše Mountains National Park and location of suitable snowfields. We also wish to thank Irena Matulková and Ivan Němec for the use of a DXR Raman microscope.
Disclosure Statement
No competing financial interests exist.