
Abstract
As part of its biosignature detection package, the Mars 2020 rover will carry PIXL, the Planetary Instrument for X-ray Lithochemistry, a spatially resolved X-ray fluorescence (μXRF) spectrometer. Understanding the types of biosignatures detectable by μXRF and the rock types μXRF is most effective at analyzing is therefore an important goal in preparation for in situ Mars 2020 science and sample selection. We tested mesoscale chemical mapping for biosignature interpretation in microbialites. In particular, we used μXRF to identify spatial distributions and associations between various elements (“fluorescence microfacies”) to infer the physical, biological, and chemical processes that produced the observed compositional distributions.
As a test case, elemental distributions from μXRF scans of stromatolites from the Mesoarchean Nsuze Group (2.98 Ga) were analyzed. We included five fluorescence microfacies: laminated dolostone, laminated chert, clotted dolostone and chert, stromatolite clast breccia, and cavity fill. Laminated dolostone was formed primarily by microbial mats that trapped and bound loose sediment and likely precipitated carbonate mud at a shallow depth below the mat surface. Laminated chert was produced by the secondary silicification of microbial mats. Clotted dolostone and chert grew as cauliform, cryptically laminated mounds similar to younger thrombolites and was likely formed by a combination of mat growth and patchy precipitation of early-formed carbonate. Stromatolite clast breccias formed as lag deposits filling erosional scours and interstromatolite spaces. Cavities were filled by microquartz, Mn-rich dolomite, and partially dolomitized calcite. Overall, we concluded that μXRF is effective for inferring genetic processes and identifying biosignatures in compositionally heterogeneous rocks. Key Words: Stromatolites—Biosignatures—Spectroscopy—Archean. Astrobiology 17, 1161–1172.
1. Introduction
T
Because one of PIXL's key missions will be to analyze microbial biosignatures that may occur in martian rocks and to screen for rocks that may contain biosignatures detectable after sample return, it is important to understand how μXRF mapping over centimeter-scale features can contribute to in situ biosignature analysis. Microbial biosignatures are classified (Cady et al., 2003) as (1) microbial body fossils, (2) organic biomarkers and isotopically fractionated compounds, and (3) sedimentary traces of benthic communities (microbial mats). Microbial body fossils are generally impossible to detect in situ on Earth and will likely be so on Mars, meaning that such evidence, if any, will need to be in returned samples. The rover will be equipped with a sensitive UV/visible/Raman spectrometer for in situ biomarker detection (SHERLOC). PIXL will be tasked primarily with detecting and characterizing inorganic trace evidence for past life. Thus, understanding the relationships between microbial textures, associated compositional variations, and the processes that produced them will be critical to interpreting similar PIXL data from Mars.
To test μXRF as a technique for microbial biosignature detection and interpretation, we conducted systematic scans of Mesoarchean stromatolite samples from the 2.98 Ga Nsuze Group, South Africa, at spatial resolution similar to that which will be achievable by PIXL. Results were compared with higher-resolution analyses and analyses from optical microscopy in order to explore potential limits to interpretations possible with in situ data generated on Mars.
Although stromatolites are obvious targets for astrobiological investigation, their value as biosignatures depends in part on the ability to infer biological modes of growth from constituent fabrics and textures, and this can be complicated by postdepositional carbonate mineral recrystallization (e.g., Grotzinger and Knoll, 1999; Allwood et al., 2009). For instance, overprinting of primary grain sizes and orientations can make it difficult to identify layers formed by different physical and biological mechanisms. Carbonate recrystallization is likely in any rocks subjected to subsurface fluid flow on Earth or Mars, complicating the utility of regions exposed to such flow for astrobiological field operations. However, because subsurface aquifers are also habitable environments, they are priority targets for astrobiological exploration on Mars (Lin et al., 2006; McLennan et al., 2012). It is therefore necessary to develop robust in situ techniques for analyzing potential biosignatures of surficial microbial communities in recrystallized carbonate rocks. Chemical mapping by centimeter-scale μXRF techniques such as PIXL has the potential to do this. In particular, isolated noncarbonate minerals (quartz, feldspar, clay, zircon, rutile, etc.) are all potentially identifiable in layers that accumulated detrital grains, and they can be used to identify sedimentary structures or to estimate primary grain sizes. These grains are all particularly accessible to chemical mapping by μXRF based on their distinctive chemical compositions. Moreover, if distinct phases of carbonate mineralization or recrystallization produce varying cation compositions, this could also be detected by chemical mapping.
Despite the fact that some abiotic stromatolites are known, stromatolites are important terrestrial records of microbial processes from the Paleoarchean to the modern (Hofmann, 1973; Riding, 1991, 2011; Grotzinger and Knoll, 1999; Allwood et al., 2009; Tice et al., 2011; Bosak et al., 2013a, 2013b). Nsuze stromatolites, in particular, are significant because their morphologies and textures are well preserved compared to other Archean stromatolites (Beukes and Lowe, 1989; Altermann, 2008; Hicks et al., 2011). They are also some of the oldest stromatolites with morphological and textural diversity similar to stromatolites of the Proterozoic, Phanerozoic, and the modern (Lowe and Tice, 2007). Stromatolites formed before 3.0 Ga were usually small and poorly developed when compared to the large, morphologically and texturally varied stromatolites that formed < 3.0 Ga. It is unclear whether this change was due to changing environmental conditions, the evolution of new stromatolite-constructing microorganisms, or qualitative changes in the community processes leading to stromatolite growth (Tice et al., 2011). Determining the relative roles of different biological and abiotic processes in the morphogenesis of these stromatolites thus has the potential to contribute to a broader understanding of the changing roles of microbial mats shaping the historical stromatolite record.
For the purposes of this study, a stromatolite is an attached, laminated, lithified sedimentary growth structure, accretionary away from a point or limited surface of initiation (Semikhatov, 1979; Grotzinger and Knoll, 1999; Tice et al., 2011). Stromatolites are formed by the interaction of various biological, physical, and chemical processes (Hofmann, 1973; Riding, 1991, 2011; Grotzinger and Knoll, 1999; Allwood et al., 2009; Tice et al., 2011; Bosak et al., 2013a, 2013b). Biological processes include trapping and binding, microbial growth and metabolism, taxis, and extracellular polymeric substance production. Physical processes include erosion of microbial mats and stromatolites as well as deposition, sorting, and current reworking of detrital sediment. Chemical processes include precipitation, dissolution, and diagenetic modification of cements. These processes can produce distinctive textures or variations in lamination thickness.
Repeated trapping and binding of sediment commonly produces irregular laminated textures in stromatolites (Grotzinger and Knoll, 1999). Mat roughness and the stickiness of extracellular polymeric substances trap sediment grains (Kawaguchi and Decho, 2000; Tice et al., 2011). Binding occurs when microorganisms move or grow over a layer of trapped sediment.
The distribution of net metabolic processes within the mat is typically not preserved directly in stromatolitic textures, but it can affect other processes such as precipitation (Pentecost and Riding, 1986; Meister, 2013; Zeng and Tice, 2014). Mats commonly are self-organized into zones by depth in which metabolisms dependent on distinct electron acceptors dominate. In modern mats, the layer closest to the surface commonly comprises organisms that perform oxygenic photosynthesis, typically cyanobacteria, diatoms, and algae, followed by a layer of anoxygenic photosynthetic bacteria, and then a deep layer of anaerobic heterotrophic organisms dominated by sulfate reducers (Riding, 2000; Dupraz et al., 2013). This modern structure results from growth in an aerobic, sulfate-rich environment; however, microbial mat structures were most likely different in the Archean, when ambient oxygen and sulfate levels were significantly lower (Olsen, 2006).
Precipitation of early diagenetic cements, particularly carbonate minerals, is critical to the formation and preservation of stromatolites (Ginsburg, 1991; Grotzinger and Knoll, 1999; Riding, 2000; Bosak et al., 2013a). Precipitation and dissolution, although chemical processes, also commonly are controlled or induced by biological processes of the mat community. Microbial processes such as photosynthesis, sulfate reduction, and metal reduction commonly alter the concentrations of HCO3 −, OH−, or both, potentially increasing carbonate saturation (Pentecost and Riding, 1986; Meister, 2013; Zeng and Tice, 2014). Mat preservation can therefore be controlled by the microbial mat layer that promotes carbonate precipitation, unless precipitation is driven by environmental forcing. If carbonate precipitation is driven by photosynthesis, then the upper layer of mat will be preserved and lower layers may be only poorly developed. If the anaerobic oxidation of organic matter promotes carbonate precipitation, then the lower layer of the microbial mat will be preserved. Precipitation induced by externally driven saturation such as episodic influx of evaporitic fluids has the potential to preserve the entire mat profile.
To identify the relative roles of physical, biological, and chemical/diagenetic processes in Nsuze stromatolite morphogenesis, elemental compositions and spatial distributions were related to early sedimentary features of the rock in “fluorescence microfacies” and used to infer growth processes. We define fluorescence microfacies as packages of rock identifiable by XRF and mappable at the centimeter scale. A growth model for an Nsuze stromatolite was ultimately derived from the distribution of different fluorescence microfacies.
2. Geological Background
The 2.98 Ga Nsuze Group was deposited nonconformably on basement granitoid rocks and is the basal unit of the 2.98–2.87 Ga Pongola Supergroup, located in South Africa and Swaziland (Hegner et al., 1994). Major lithologies are volcanic and siliciclastic rocks that were deposited in an intracratonic basin and experienced burial to greenschist facies metamorphism. The Nsuze Group is subdivided into three units: the lower fluvial feldspathic sandstone of the Montonga Formation, the andesite deposits of the Nhlebela Formation, and the tidal deposits of the White Mfolozi Formation (Hicks et al., 2011).
The White Mfolozi Formation is a volcano-sedimentary succession comprised of altering layers of volcaniclastic sandstone, shale, quartz arenite, and stromatolitic dolostone. The quartz arenite in this unit is medium- to fine-grained and well rounded, and contains cross beds, herringbone cross beds, ripple marks, and flaser and lenticular bedding. These features are consistent with formation by deposition and reworking by tidal currents (Balazs and Klein, 1972; von Brunn and Mason, 1977; Hicks et al., 2011). Structures related to the cohesion and metabolism of shallow-water microbial mats (MISS, or microbially induced sedimentary structures) have been described in well-preserved Pongola sandstones from nearby localities (Noffke et al., 2003, 2008). The stromatolitic carbonate unit is in the upper part of the formation and likely formed in intertidal and shallow subtidal settings (Beukes and Lowe, 1989). Stratiform, domal, columnar, and conical stromatolites composed of interlaminated gray dolostone and chert are common in this unit (Beukes and Lowe, 1989). Dolostone and chert are secondary lithologies formed after the growth of the stromatolite, with dolostone formed by the replacement of calcium carbonate by dolomite, and chert formed by replacement of some carbonate and other primary materials during silicification (Beukes and Lowe, 1989). Stromatolitic laminations commonly contain admixed siliclastic grains interpreted to have been trapped and bound by microbial mats (von Brunn and Mason, 1977; Beukes and Lowe, 1989). Nsuze stromatolites thus form an ideal test case to determine whether centimeter-scale μXRF mapping could be useful for inferring biological, physical, and chemical growth processes in partially recrystallized microbialites.
3. Materials and Methods
To analyze elemental distributions in Nsuze Group stromatolites, a large block was collected from float along the White Mfolozi River, South Africa, at the location described by Beukes and Lowe (1989). The block was cut into three slabs approximately 1 cm thick in order to describe the lateral continuity of observed microfacies. Each slab face was scanned by μXRF (Horiba XGT-7000). Scans were performed at a spatial resolution of 100 μm over 5.12 × 5.12 cm2 to approximate the resolution of PIXL. The X-ray generator uses a rhodium block as a target material for generating X-rays, an accelerating tube voltage of 50 kV, and a filament current of 1 mA. Full spectra from 0 to 40 keV were collected at each point with a duration time of 0.02–0.05 s over 8–16 accumulations. Resulting fluorescence maps were analyzed with ImageJ (Rasband, 2014). Fluorescence microfacies were defined by elemental associations with each other and with physical structures.
To evaluate conclusions drawn from PIXL-like data relative to more detailed analyses that could only be accomplished upon sample return to Earth, thin sections were cut and analyzed by optical petrography to confirm or refine mineral grain identifications from μXRF scans.
The μXRF instrument used for this study and PIXL are similar in that both use Rh as the target material and produce fluorescence elemental scans at approximately 100 μm resolution. However, PIXL will use a tube voltage of 28 kV and a filament current of 200 μA. The higher voltage and current used in this study produce a more intense fluorescence and thus allow for data collection over a shorter period of time. PIXL also scans smaller areas by directing an X-ray beam from a stationary position across a stationary target, causing the angle of incidence to change across the scanning surface and changing the measurement resolution slightly away from the center of the target. In contrast, the Horiba μXRF instrument rasters the target below a stationary X-ray beam, producing constant spatial resolution for flat samples such as those in this study.
Although PIXL will be capable of collecting measurements on points and sparse arrays of points on rough surfaces without significant degradation of data quality (Allwood et al., 2015), current operational plans call for more intensive mapping such as the analyses conducted here to be performed on abraded or naturally flat surfaces. Moreover, the angle of the X-ray detector relative to the incident X-ray beam differs significantly between PIXL and the Horiba XGT-7000, limiting comparisons between their data-collecting capabilities over uneven samples. For these reasons, analyses were not attempted on uncut samples.
4. Results
4.1. Fluorescence microfacies
Ca, Si, K, Fe, S, Mn, Mg, Ti, and Zr were all detected in the Nsuze Group samples (Fig. 1) and used to subdivide it into five fluorescence microfacies: laminated dolostone, laminated chert, clotted dolostone and chert, stromatolite clast breccia, and cavity fill (Table 1; Figs. 2 –4).
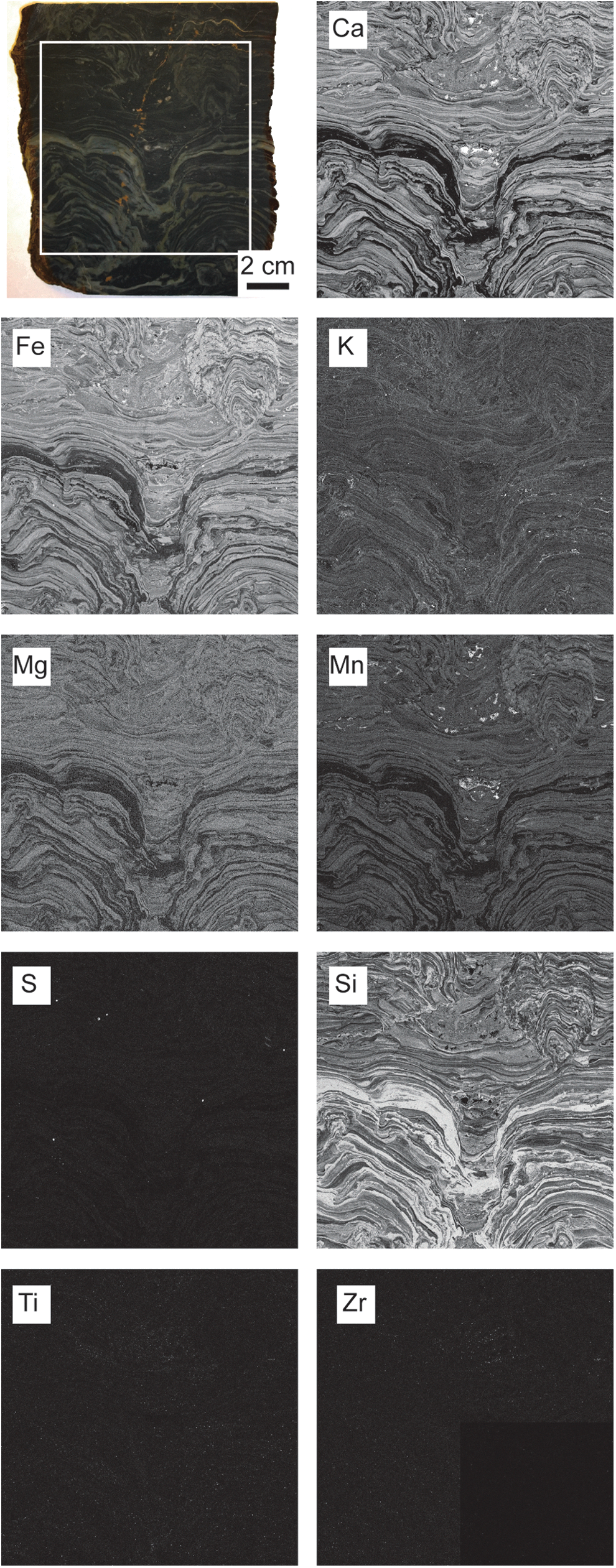
Slab cut perpendicular to bedding through Nsuze Group stromatolite sample with corresponding elemental fluorescence maps of boxed area. Ca, Si, K, Fe, S, Mn, Mg, Ti, and Zr were all detected at mappable intensities. All mapping performed at 100 μm resolution.

False-color elemental fluorescence images of six slab faces from the Nsuze sample with red = Fe Kα1, green = Ca Kα1, and blue = Mn Kα1 intensity. In this color mapping, green layers are dolostone, black layers are chert, and light blue is Mn-rich cavity fill. All mapping performed at 100 μm resolution.
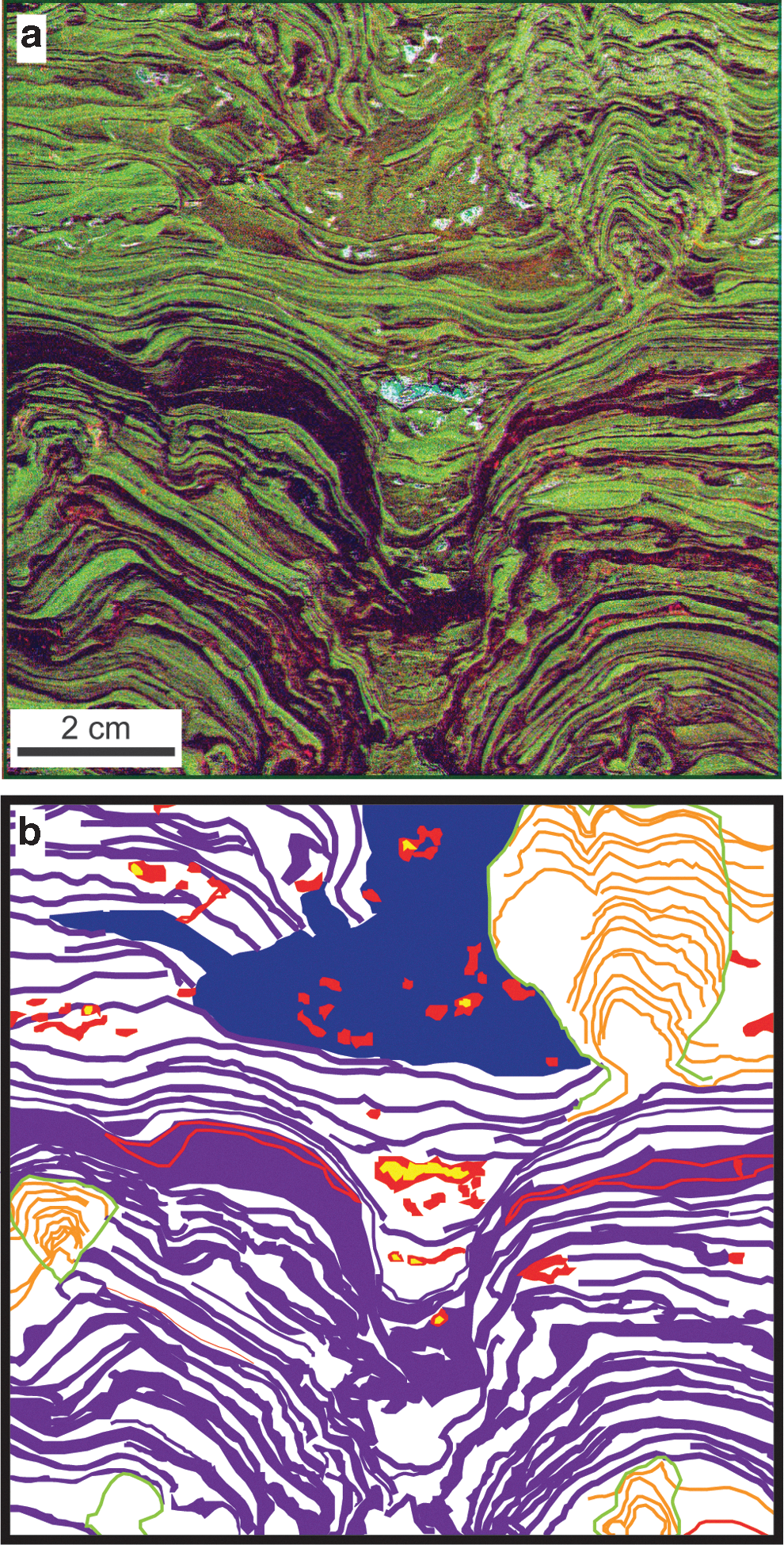
Fluorescence microfacies distributions. (

Fluorescence microfacies. The sample was divided into five microfacies. (
Fluorescence intensities integrated over 200 eV windows centered on Kα1 lines with background subtracted and reported as mean ± standard deviation in counts mA−1 s−1.
4.1.1. Laminated dolostone
Description: Common elements in the laminated dolostone fluorescence microfacies are Ca, Fe, Si, K, Ti, and Zr (Table 1). Ca, Mg, Fe, and Mn have highly similar physical distributions and are correlated in this microfacies (R 2 ≥ 0.953 for pairs of each element averaged over 10 distinct layers), implying that the Fe and Mn are a part of the dolomite (Fig. 4a). Coarse (500–700 μm) Fe- and S-rich grains (pyrite) are randomly dispersed throughout the facies (Fig. 1 Fe, S). Some Fe also occurs in unidentified isolated grains without S. Si-rich grains occur in the dolostone laminations, most likely in quartz, and commonly define microlaminations within larger dolomitic laminations (Fig. 4a). The occurrence of contrasting sediment types allows visualization of sedimentary structures in the dolomitic laminations that would otherwise be impossible to detect, such as grading, laminations, and cross laminations (Fig. 4a). Syndepositional slumps and faults (Fig. 4c) occur on the tops of both small domes, whereas dolostone chip breccia occurs on dome flanks (Fig. 4d). Ti and Zr are dispersed in isolated grains having sizes similar to the resolution of the μXRF probe head (100 μm) throughout the laminations and are likely present as rutile or anatase and zircon grains, respectively (Fig. 5). Very fine K-rich minerals line lamination boundaries and are inferred to represent drapes of K-rich phyllosilicate minerals (Fig. 1 K). Individual dolomitic laminations frequently form slopes greater than the angle of repose for unconsolidated sediment (Fig. 3a).

Rutile and zircon distributions. False-color elemental fluorescence image showing distributions of Ti Kα1 (cyan) and Zr Kα1 (yellow) intensity with respect to dolostone (magenta) and chert (black) layers. Color mapping emphasizes distribution of fine, isolated grains. Field of view as in Fig. 3. All mapping performed at 100 μm resolution.
Interpretation: This fluorescence microfacies was likely formed primarily by the growth of microbial mats that trapped and bound carbonate and siliciclastic sediment and precipitated micritic carbonate cement. Mats produced cohesive layers and highly inclined surfaces. A single layer containing truncated cross-laminated dolomite (Fig. 4a) indicates episodes of bed-load transport and deposition of up to medium sand-sized carbonate grains.
Syndepositional slumps, faults, and dolostone chip breccias are contained in one layer of the stromatolite, marking disturbance by a single event that induced both plastic and brittle deformation. The variation in deformation style suggests a steep gradient in material properties near the sediment/water interface, with a shallow cohesive layer capable of flow and a lithified layer <2 cm below the surface. The strength gradient with respect to distance from the stromatolite surface was steeper on the flanks, producing more brittle failure along dome margins than on dome tops, where slumping dominated.
Pyrite grains were unlikely to have been transported by currents based on comparison to co-occurring grain types. Carbonate grains had a maximum diameter of 300 μm based on the maximum thickness of laminations in the cross-laminated dolostone layer. The ratio of the Stokes settling velocities of the pyrite grains (ρ
pyr = 5.0 g cm−3) to carbonate grains (ρ
carb = 2.7–2.9 g cm−3), using the maximum inferred diameters, is ∼8.9 (Eq. 1).
Here, ρ pyr, ρ carb, and ρ water are the densities of pyrite, carbonate, and water, respectively; d pyr and d carb are the diameters of pyrite and carbonate grains; and ω pyr and ω carb are the Stokes settling velocities of pyrite and carbonate grains.
The Rouse number (Eq. 2) of a grain relates the hydraulic properties of a flow and grain properties to transport mechanisms. Here, ω
s is the setting velocity of the grain; κ is the von Kármán constant; u
* is the shear velocity of the fluid, and R
pyr and R
carb are the Rouse numbers for pyrite and carbonate grains. R
o greater than 6.25 indicates bed-load transport, R
o between 6.25 and 1 indicates mixed load transport, and R
o less than 1 indicates suspended load transport. By comparing the ratios of the Stokes settling velocity and the Rouse number of the pyrite grains to the carbonate grains and allowing R
opyr to equal the minimum value for bed-load transport, the Rouse number of the carbonate grains is 0.7 (Eq. 3). The carbonate grain would therefore be transported by suspended load if the pyrite grains were transported by bed load. Therefore, these grains are not hydraulically equivalent and would not have been deposited by the same current.
Alternatively, if the pyrite had recrystallized from primary grains 100 μm in diameter, they would have had similar settling velocities and facies distributions to zircon (ρ zircon = 4.7 g cm−3) and rutile (ρ rutile = 4.2 g cm−3) grains. Instead, pyrite is distributed randomly and sparsely in the laminated dolostone fluorescence microfacies, whereas zircon and rutile are distributed along microlaminations, in interstromatolite spaces, and in stromatolite clast breccias (Fig. 5). Pyrite is therefore interpreted to have formed by in situ precipitation and does not reflect the oxidation state of the overlying water column during stromatolite growth.
4.1.2. Laminated chert
Description: The primary element detected in the laminated chert fluorescence microfacies is Si (Table 1), although other elements associated with siliclastic sedimentary grains (K, Ti, Zr) occur in laminations and lenses of fine-grained phyllosilicates (Fig. 1 K), randomly disseminated heavy minerals (Fig. 5), and rare sand-sized K-rich grains (Figs. 1 K and 6). Isolated carbonate grains and unidentified Fe-rich grains also occur in the chert laminations (Fig. 4a). Like some dolostone laminations, some chert laminations formed slopes above the angle of repose for unconsolidated sediment (Fig. 3a). Some chert laminations include 2–4 mm tall peaked structures that are vertically oriented on both the tops and sloping flanks of domes (Fig. 6). Dolostone laminations sometimes terminate or lap onto these peaks (Fig. 6). Some peaks contain sand-sized K-rich grains organized into small conical piles 2–3 grains deep (Fig. 6). Partially rolled-up chert chips occur in a single dolostone lamination (Fig. 6).
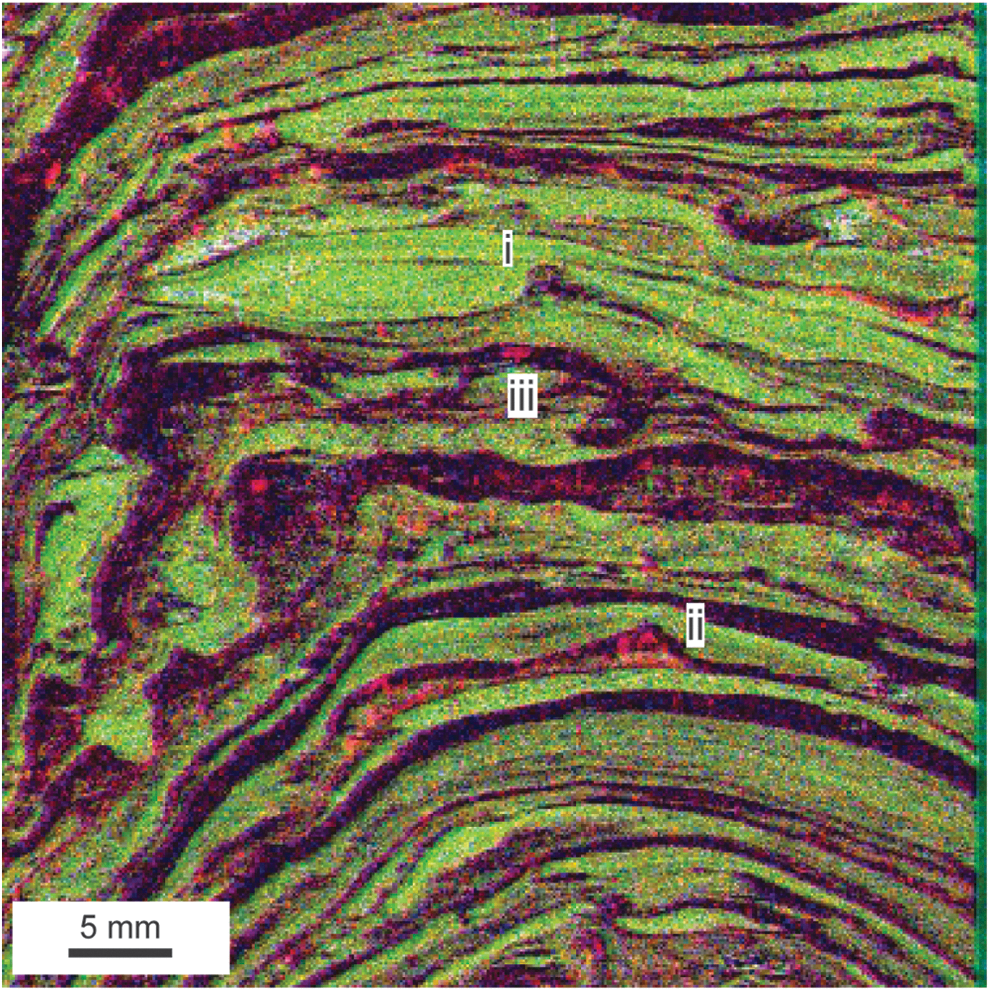
Matlike structures in the laminated chert facies. Peaks with onlapping truncated dolostone layers (i, ii) and rolled chert chip (iii). Color mapping as in Fig. 2. Peak grew on top of a small pile of K- and Fe-rich sand-sized grains. All mapping performed at 100 μm resolution.
Interpretation: The chert in the laminated chert fluorescence microfacies formed during silicification of a preexisting material that was cohesive as evidenced by oversteepened layers. It also flowed plastically as indicated by the presence of floating dolostone chips in disrupted chert laminations (Figs. 3a, 4a). Rare rolled-up chert bodies are consistent with this interpretation and likely represent microbial roll-up structures (Sumner, 1997; Simonson and Carney, 1999; Tice and Lowe, 2004; Bosak et al., 2013b). Uniform orientation of chert peaks and their effect on the distribution of detrital sediment cannot be explained by formation by soft sediment deformation or diagenesis; instead, they likely formed as microbial cones or pinnacles. These inferences suggest that the laminated chert fluorescence microfacies represents silicified microbial mat that incorporated little to no sand-sized detrital material, although it is possible that some trapped or precipitated micrite was present and later replaced during silicification. Chert laminae in domical stromatolites of the 3.45 Ga Strelley Pool Formation likely formed by a similar replacement mechanism (Allwood et al., 2009).
4.1.3. Clotted dolostone and chert
Description: The clotted dolostone and chert fluorescence microfacies is composed of high-relief cauliform mounds composed of intergrown dolomite and chert forming clots up to 2 mm in diameter and cryptically laminated patches (Figs. 3 and 4c). Cryptic laminations can be vertical or even overhanging. This fluorescence microfacies contains abundant cavities filled with Mn-rich dolomite of the cavity fill fluorescence microfacies (Fig. 4c). Clotted dolostone and chert fluorescence microfacies form the core of each stromatolite as well as the branches formed on top of other stromatolites. From thresholded maps of Mn and Si, the primary porosity of the clotted microfacies defined as the fraction of the facies filled with the cavity fill microfacies was estimated to range from 37% to 50%.
Interpretation: Mounds of clotted dolostone and chert formed at the sediment/water interface; they seeded new stromatolites and stromatolite branches, and dolostone laminations onlapped and truncated against them (Fig. 4c). Therefore, they were not formed through the diagenetic alteration of older stromatolites. Instead, their steep relief and high initial porosity likely resulted from patchy mineralizing microbial colonies that locally graded into thin flat mats or carbonate encrustations. They are here interpreted as thrombolites that transitioned upward into stromatolites as in many younger microbialites (Aitken, 1967; Kennard and James, 1986). This also makes them the oldest known thrombolites by ∼1 Ga (Kah and Grotzinger, 1992) and highlights a potentially important similarity between Nsuze Group microbialites and younger assemblages.
4.1.4. Stromatolite clast breccia
Description: The stromatolite clast breccia fluorescence facies is a breccia composed of stromatolite clasts supported by a massive matrix composed of mixed chert and dolomite and containing an abundance of heavy mineral grains and unidentified Fe-rich grains (Figs. 3 and 4d). This microfacies fills erosional scours and space between stromatolites. The stromatolite clast breccia hosts the majority of the large cavities filled by the cavity fill fluorescence microfacies (Fig. 4d).
Interpretation: This microfacies was deposited when energetic currents eroded stromatolites and other sediment and rapidly redeposited them as lags in topographic lows with little to no sorting. Cavities likely formed by decomposition of eroded mat fragments.
4.1.5. Cavity fill
Description: Cavities in both the clotted dolostone and chert and stromatolite clast breccia fluorescence microfacies are filled with intergrown Mn-rich dolomite and microquartz, and where cavities are larger than ∼0.5 mm, by an inner core of partially dolomitized calcite (Fig. 4b). This microfacies is therefore relatively rich in both Mn and Ca (Table 1).
Interpretation: The Mn-rich cavity fill fluorescence microfacies reflects the composition of near-surface pore water that filled pore spaces during early diagenesis. Calcite cavity fill reflects changing pore water compositions during early burial.
4.2. Fluorescence microfacies petrographic analysis
Laminated dolomite is composed of alternating laminations of ∼30–200 μm sized dolomite grains, including euhedral to subhedral rhombs, and microquartz laminations (Fig. 7a–7c). Very fine quartz grains (∼50–100 μm) also occur in some laminations (Fig. 7a, 7d). Fine K-rich grains were identified as sericite micromosaics (Fig. 7e), which are common in the Nsuze Group and a likely metasomatic product of vitric volcaniclastic sand (von Brunn and Mason, 1977; Beukes and Lowe, 1989).

Laminated dolostone and laminated chert fluorescence microfacies in thin section (cross-polarized light). (
The clotted dolostone and chert microfacies as seen in thin section has similar geometries to those observed by μXRF. Clots are comprised of clusters of dolomicrite and dolomite rhombs with intergrown quartz. Dolomite appears similar in grain size to dolomite from other microfacies (Fig. 8).

Clotted dolostone and chert fluorescence microfacies in thin section (cross-polarized light). (
5. Discussion
Nsuze Group stromatolites observed in this study are composed of basal clotted zones that transition upward into laminated zones, making them zoned microbialites sensu Kennard and James (1986). Microbialites were seeded by the growth of high-relief cauliform thrombolites, which formed shrublike projections on the seafloor and acted as templates for subsequent accumulation of laminated stromatolites. Capping domal stromatolites formed the bulk of these structures and were built by the mat-aided accumulation and cementation of micrite and granular sediment, now preserved as laminated dolomite layers, and clastic-poor microbial mats, now preserved as secondary laminated chert layers. Alternating clastic-rich dolostone layers and clastic-poor mat layers likely reflect changing current intensities and sediment supply relative to mat growth rate.
The combination of plastic failure at shallow depths in the stromatolites and brittle failure at slightly greater depths was likely related to different sources of material strength in each zone. In the shallow zone, strength likely resulted from the cohesion of the microbial mat itself, that is, from the presence of extracellular polymeric substances produced by microorganisms and potentially from intergrown filamentous organisms (Stal, 1995; de Winder et al. 1999; de Brouwer et al., 2005; Gerdes, 2007; Gerbersdorf and Wieprecht, 2014). In the deep zone, strength likely resulted from precipitation of early diagenetic carbonate cements that lithified the mat. In modern mats, cementation at depth frequently occurs as mats decompose, often during sulfate reduction (Riding, 2000; Dupraz et al., 2009). In the low-sulfate oceans of the Mesoarchean (Habicht et al., 2002; Kah et al., 2004), cementation may have been associated with iron reduction (Olsen, 2006; Meister, 2013; Zeng and Tice, 2014). In either case, the transition from plastic to brittle deformation from dome tops to flanks in Nsuze Group stromatolites likely occurred at, or near, the boundary between net autotrophic production of organic matter in the shallow mat and net heterotrophic oxidation of organic matter in the deep mat, and thus reflected metabolic zonation within the original mat communities (Fig. 9). If this boundary occurred at a constant vertical depth within the mat (e.g., at a depth of constant light attenuation), then it would occur at a shallower depth measured perpendicular to the mat surface on dome flanks than on dome tops. For instance, on a flank dipping at 45° with respect to horizontal, such an autotrophic/plastic to heterotrophic/brittle transition would be 30% closer to the surface than the same transition below a horizontal surface.
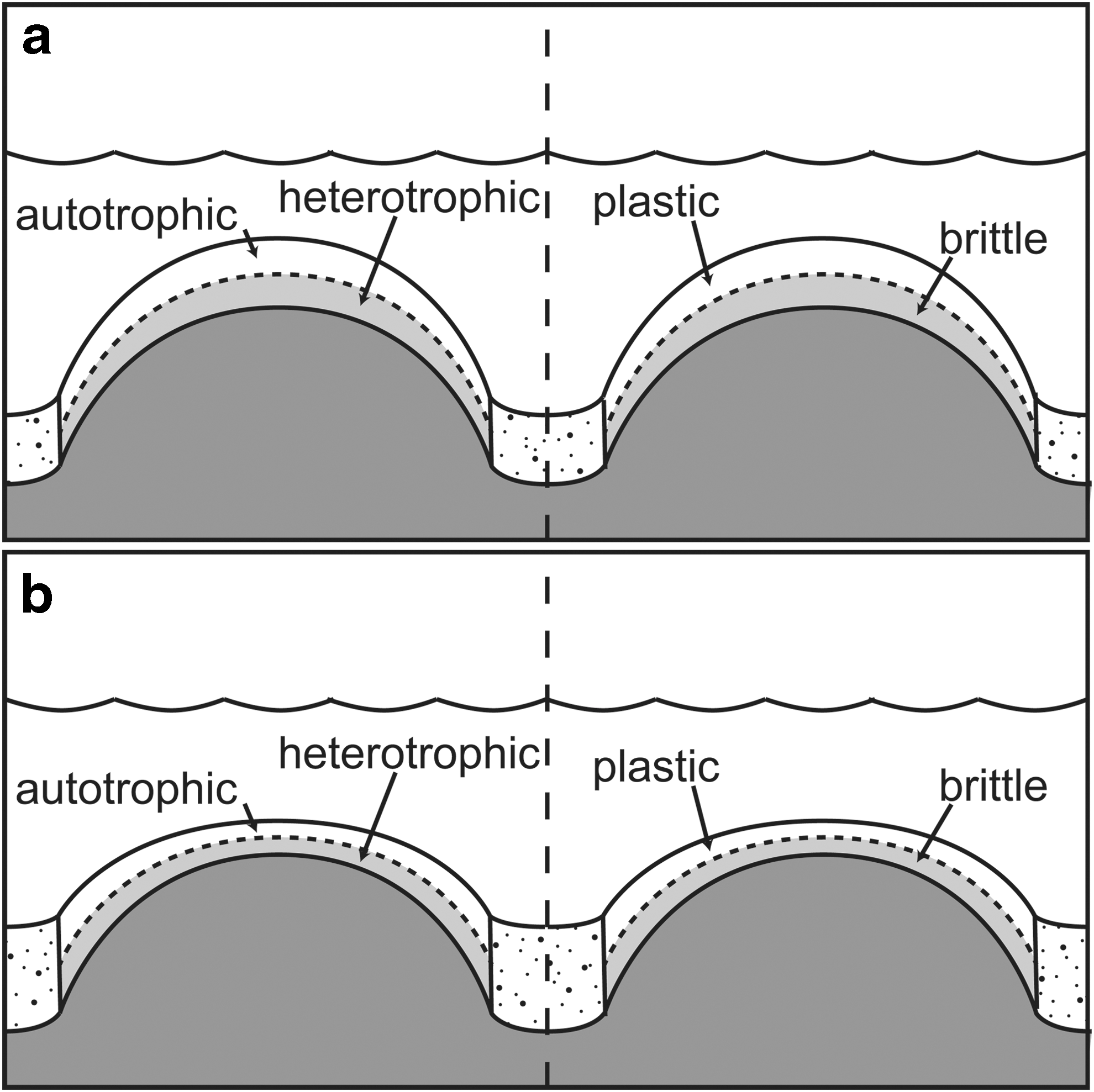
Mat profiles across domal stromatolites during size and shape maintenance and during drowning. Hypothesized metabolic and deformational zonation are illustrated. Constant vertical thickness of net autotrophic and heterotrophic zones (left side of figure) resulted in corresponding plastic and brittle zones (right side of figure) that thinned perpendicular to the mat surface along dome flanks. (
Domal stromatolite growth maintained constant synoptic relief above the surrounding sediment as long as mat growth, precipitation, and sediment trapping and binding kept pace with accumulation of detrital material and slumped mat between domes. Constant synoptic relief is shown by the continued upward growth of dolostone and chert laminations and the maintained domal structure of the stromatolite, as detrital grains were deposited in the space between the domes. In the sample examined, domes began to drown when dolostone layers began to thicken between domes and thin onto their tops. Laminated chert layers are less common in sediments deposited during this time, suggesting that the supply of carbonate sediment had increased, likely due to increased ambient current activity. Relatively flat dolostone layers dominate at this point, consistent with predictions of mat flattening under high-shear currents (Tice et al., 2011).
Traditional petrographic analysis supported many of the observations and interpretations made based solely by μXRF analysis; however, some observations were possible only with one technique. Because they did not contrast in composition, quartz sand grains in chert laminations were not detected by μXRF, and siliciclastic content was instead inferred from Ti-, Zr-, and K-rich grains. Sand-sized K-rich grains were not identifiable from μXRF alone; they were identified as sericitized volcaniclastic grains optically. On the other hand, the spatial distributions of these grains within laminations and across microfacies were more easily detected by μXRF, and the identification of early diagenetic Mn-rich dolomite in both large cavities and thrombolitic pores was only possible by this technique.
6. Conclusions
In this test case, μXRF highlighted sedimentary features and textures that were not apparent upon visual inspection of the cut sample or even by petrographic examination of thin sections. The technique was particularly effective for (i) mapping contacts between lithologies that were visible but not as clear in hand sample, (ii) identifying laminated and cross-laminated textures marked by compositionally contrasting grain types, and (iii) distinguishing an important early diagenetic generation of carbonate void fill. Detection of those features and relationships enabled process interpretation despite pervasive dolomitization of the sample, including identification of previously unrecognized thrombolitic mounds as critical components of early morphogenesis in the observed stromatolites. Because PIXL will generate geochemical maps at similar resolution and detection limits on the surface of martian rocks, we conclude that it could be an effective tool for analyzing putative stromatolitic or mat-related biosignatures in partially recrystallized carbonate rocks.
μXRF scanning leveraged compositional variation for observation of sedimentary features related to microbial processes. In a lithologically homogeneous microbialite (for instance, one composed entirely of calcite or gypsum with no admixed siliclastic sediment), the ability to identify such features would have been limited. This suggests that, from a PIXL perspective, operating sites should be chosen not primarily for the availability of large bodies of relatively pure lithologies visible from orbit, but based on the likelihood that lithologies will be intimately intermixed. Nonetheless, μXRF analysis also identified distinct generations of dolomite despite their optical similarity and distribution across different microfacies. The ability to make these types of interpretations will be useful for in situ astrobiological investigations on Mars regardless of lithology.
Footnotes
Acknowledgments
The authors acknowledge partial support from the PIXL Instrument Project.
Author Disclosure Statement
No competing financial interests exist.