
Abstract
On Earth, microorganisms commonly enhance mineral precipitation and mediate mineralogical and chemical compositions of resulting deposits, particularly at spring systems. However, preservation of any type of microbial fossil or chemical or textural biosignature depends on the degree of alteration during diagenesis and factors such as exposure to diagenetic fluids. Little is known about the transformation of biosignatures during diagenesis over geologic time. Ten Mile Graben, Utah, USA, hosts a cold spring system that is an exceptional site for evaluation of diagenetic alteration of biosignatures because of the presence of modern springs with actively precipitating microbial mats and a series of progressively older tufa terraces (<400 ka) preserved in the area from the same spring system. A previously undescribed Jurassic laminated carbonate unit within the upper part of the Brushy Basin Member of the Morrison Formation is also exposed in Ten Mile Graben. This research characterizes the geology of these modern and Quaternary saline, Fe-undersaturated, circumneutral Ten Mile Graben cold springs and provides the first description in the literature of the Jurassic Brushy Basin Member of the Morrison Formation carbonate deposit. Taphonomy of microbial fossils is characterized by scanning electron microscopy (SEM). The data highlight two distinct methods of biosignature formation: (1) precipitation of minerals from an undersaturated solution owing to metabolic activity of the cells and (2) mineral precipitation on charged cell surfaces that produce distinctive microbial trace fossils. Although diagenesis can destroy or severely degrade biosignatures, particularly microbial fossils, some fossils and trace fossils are preserved because entombment by Ostwald ripening limits diagenetic alteration. Recognizing spring-fed, biogenic tufas is crucial for astrobiological research and the search for life on Mars. Key Words: Biosignatures—Taphonomy—Diagenesis—Carbonates—Hot springs. Astrobiology 17, 216–230.
1. Introduction
O
Biosignatures are fingerprints of life preserved as a result of fluid/rock/biota interaction, owing to microbial enhancement of mineral precipitation during deposition and/or early diagenesis (Farmer and Des Marais, 1999; Banfield et al., 2000, 2001; Cady and Farmer, 1996; Cady, 2001, Cady et al., 2003; Westall, 2008; Cady and Noffke, 2009; Chan et al., 2009, 2010). Three types of biosignatures are described, as follows: (1) morphological microfossils that retain the carbonaceous remains of cells, (2) macroscopic microbially induced sedimentary structures (MISS) such as microbialites (wavy laminated sedimentary structures), (3) chemofossils that include organic compounds such as lipids, biominerals, isotopic signatures, and concentrations of trace elements specific to sequestration by microbes (Cady et al., 2003; Westall, 2008). Biosignatures can be preserved via many different pathways in many lithologies (e.g., carbonates, clay-rich shales, water-laid pyroclastics, and some iron formations; Allison and Briggs, 1991; Farmer and Des Marais, 1999; Cady, 2001; Rainey and Jones, 2010). Most previous research on biosignatures has focused on comparing modern processes with very ancient (>1 Ga) putative microbially induced features (e.g., Cady, 2001; Konhauser et al., 2003; Mojzsis et al., 2003; Schopf et al., 2010; Corkeron et al., 2012), creating inherent uncertainty in their interpretation because diagenetic alteration may be significant (Parenteau and Cady, 2010; Potter-McIntyre et al., 2014a). Moreover, establishing biogenicity of putative morphological microbial fossils can often be problematic, especially because of changes that occur during diagenesis (Farmer and Des Marais, 1999; Cady et al., 2003; Schopf et al., 2010).
Preservation of any type of microbial fossil or chemical or textural biosignature depends on the degree of alteration during diagenesis and factors such as exposure to diagenetic fluids (Cady, 2001; Westall, 2008). Diagenetic alteration includes processes such as destruction of organic matter, compaction, dissolution of minerals, precipitation of cements, recrystallization of metastable minerals to more stable phases, dolomitization, and biogenic alteration of sediments—all of which can alter or destroy biosignatures. Microbial carbonates have been extensively examined in modern systems; however, little is known about the transformation of biosignatures during diagenesis over geologic time (Parenteau et al., 2010; Potter-McIntyre et al., 2014a). This is one of the outstanding challenges to interpreting biosignatures on early Earth and Mars. We hypothesize that biosignatures are preserved, even in very ancient deposits, because iron (oxyhydr)oxide and carbonates have outstanding capacities to preserve biogenetic fingerprints—both by permineralization of cell structures (Cady and Farmer, 1996; Konhauser et al., 2003; Rancourt et al., 2005; Westall, 2008; Parenteau and Cady, 2010) and by preserving diagnostic trace element configurations and/or kinetically unusual mineral phases (Westall, 2008; Chan et al., 2009; Potter-McIntyre et al., 2014a).
Ten Mile Graben, Utah, USA, hosts a cold spring system that is an exceptional site in which to evaluate diagenetic alteration of biosignatures because of the presence of modern springs with actively precipitating microbial mats and a nearby series of progressively older tufa terraces (<400 ka) preserved in the area from the same spring system (Burnside, 2010). A previously undescribed Jurassic carbonate unit within the upper part of the Brushy Basin Member of the Morrison Formation is also exposed in Ten Mile Graben.
The Ten Mile Graben spring system has received attention with application to carbon capture and sequestration research because it emanates from a natural CO2 reservoir in the Jurassic Navajo Sandstone and both methane and CO2 escape along the fault system (Shipton et al., 2004; Heath et al., 2009; Burnside, 2010; Dubacq et al., 2011; Han et al., 2013; Potter-McIntyre et al., 2013; Jung et al., 2014). However, previous research has not focused specifically on the diagenetic alteration and preservation of biosignatures in carbonates and iron (oxyhydr)oxides.
2. Purpose of the Study
This research characterizes the geology, mineralogy, taphonomy of these modern and Quaternary saline, Fe-undersaturated, circumneutral Ten Mile Graben cold springs, and provides the first description in the literature of the Jurassic Brushy Basin Member of the Morrison Formation carbonate deposit. Chemical, mineralogical, and morphological biosignatures in calcium carbonates and iron (oxyhydr)oxides precipitated in the modern springs are compared with the Quaternary and Jurassic examples to show how these biosignatures are modified over geologic time. These successively older carbonate microbialites provide a novel opportunity to investigate how biosignatures are progressively altered and preserved during diagenesis on geologic timescales.
Understanding the alteration and preservation of biosignatures is essential for recognizing these signatures in the rock record of both early Earth and Mars. Noffke (2015) hypothesized that the Gillespie Lake Member at Gale Crater contains putative MISS, and the Sample Analysis at Mars (SAM) instrument on Curiosity has detected carbonates at Gale Crater (Archer et al., 2014). Carbonates have been detected in multiple locations on the martian surface such as at Gusev Crater, which is hypothesized to represent a spring environment (Squyres et al., 2004; Ehlmann et al., 2008; Boynton et al., 2009; Morris et al., 2010; Sutter et al., 2012), and in meteorites, such as ALH84001 (Mittlefehldt, 1994). Compact Reconnaissance Imaging Spectrometer for Mars (CRISM) spectral mapping of Mars' surface indicates the presence of carbonates in the Nilli Fossae region, which is also rich in clays (Ehlmann et al., 2008, 2009).
3. Geologic Setting
Ten Mile Graben hosts a series of modern CO2-driven geysers and springs, active and Quaternary fossil microbial carbonate mats, and carbonate veins that imply fault-controlled CO2 leakage from an underlying reservoir (Jung et al., 2014; Figs. 1 –3). The waters are saline (13,500–22,000 mg/L) and circumneutral (pH = 6.2–7; Shipton et al., 2004). U-Th dating of Quaternary carbonates and embedded veins reveals that CO2 has constantly leaked to the surface for >400 ka (Burnside, 2010; Burnside et al., 2013). Yet, spatially dispersed tufas with different volumes indicate that the location of CO2 leakage has varied depending on permeability variation by a combination of mineral precipitation, seismic activity, climate changes, and regional unloading (Burnside, 2010; Burnside et al., 2013). Mineral precipitation has likely also been influenced to a large extent by microbial participation (Potter-McIntyre et al., 2014a).

Google Earth image of the study area. Inset shows Ten Mile Graben on a map of Utah. Red squares show study sites (200 and 250 ka terraces are not shown because they are right next to the 300 ka terrace). Black squares show springs that were used by Shipton et al. (2004) to collect water data but were not evaluated in this study. Jmb = Jurassic Brushy Basin Member of the Morrison Formation.

Big Bubbling Creek and Crystal Geyser. (
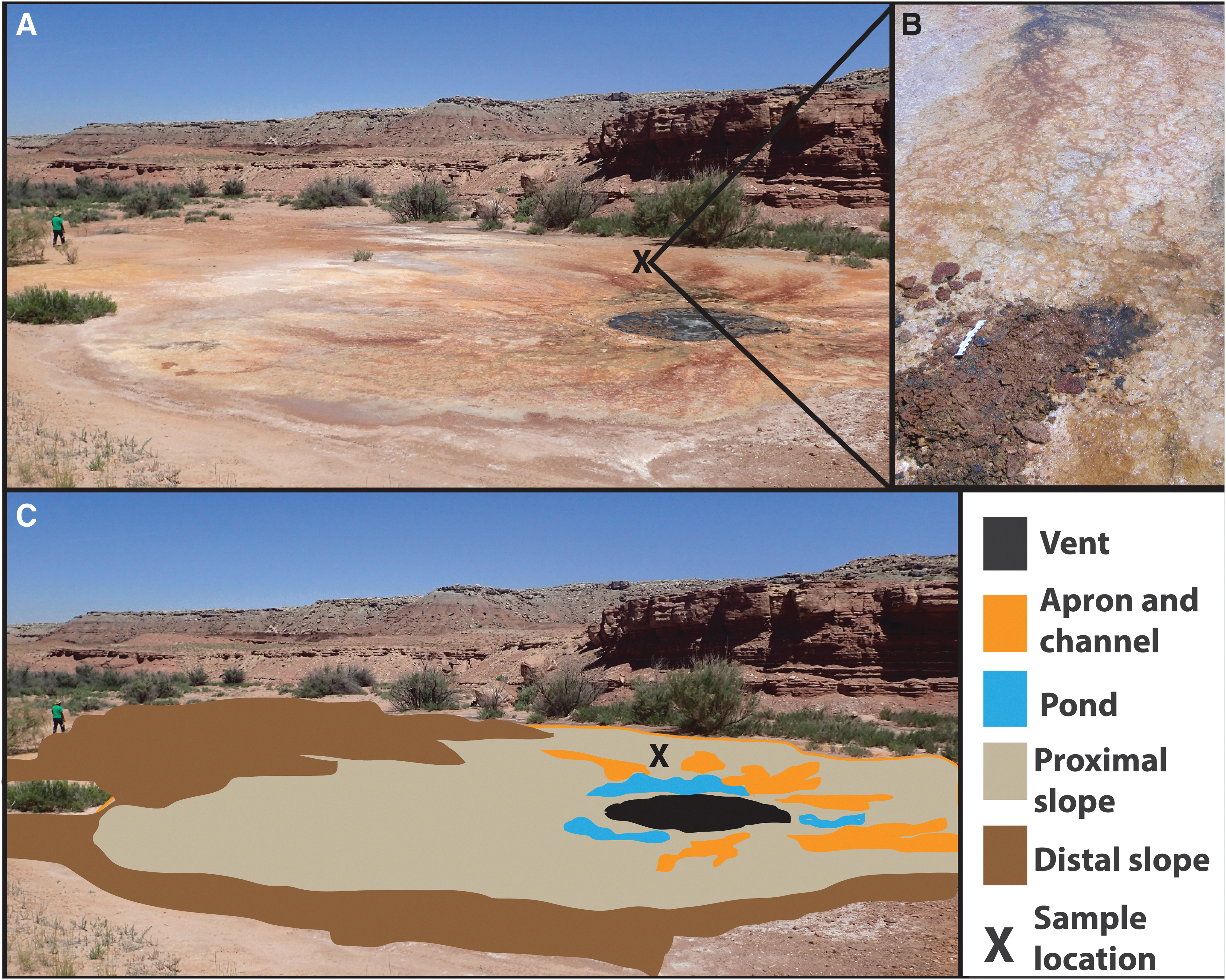
Big Bubbling Spring and spring facies. (
The 148 Ma Brushy Basin Member of the Morrison Formation is primarily a volcaniclastic shale deposited in a fluvio-lacustrine setting that extends from New Mexico to Montana in the western United States (Turner and Fishman, 1991; Kowallis et al., 1998; Turner and Peterson, 2004; Potter-McIntyre et al., 2014a, 2014b). The upper part of the unit in the Four Corners region of the United States is interpreted to have been deposited in an ephemeral, groundwater-fed, alkaline saline lake system named Lake T'oo'dichi' (Turner and Fishman, 1991). Abundant volcanic ash sourced from a back arc basin to the west created a pore-water chemistry that resulted in a bull's-eye pattern of increasingly altered minerals toward the basin center (Turner and Fishman, 1991; Turner and Peterson, 2004). Thick carbonate deposits (> a few centimeters) are undocumented in this member; however, a laterally restricted (∼10 km2), 3–10 m thick carbonate is present in the Ten Mile Graben area (to the northeast of the extent of Lake T'oo'dichi' deposits) and described for the first time herein.
4. Methods
Field research was performed by using traditional field characterization methods in the modern, Quaternary, and Jurassic deposits. Lithofacies were documented, and relations to adjacent lithofacies were recorded. Representative samples were collected for scanning electron microscopy (SEM) to identify morphological modern microbial features and associated mineral compositions and habits. The modern sample from Big Bubbling Spring was collected from a partially lithified section on the proximal slope of the spring, which is more likely to show biotic mediation (spring facies are identified after Fouke et al., 2000; Fig. 3). However, the sample was collected from ∼0–5 cm depth, and it has been shown that the precipitation rate for hot springs is ∼30 cm/yr in the proximal facies and <5 cm/yr in the distal slope facies. So it is possible (but not likely) that some of the sample was precipitated in a different facies from which it was collected. The sample was air-dried, as this was a lithified sample rather than a sample of the actively precipitating mat and was not in standing water. It should be noted that air-drying can collapse cells and alter morphology; however, this sample was partially lithified and was not collected to identify extant morphology.
X-ray diffraction (XRD) was used to identify bulk mineralogy. SEM was performed at the Southern Illinois University IMAGE facility on an FEI Quanta FEG 450 SEM with EDS detectors. Samples were gold-coated and put under high vacuum. Energy-dispersive X-ray technology (EDS) associated with the SEM was used to identify chemical composition and infer mineralogy. XRD was performed at Southern Illinois University in the Organic Petrology Lab on a Rigaku Ultima IV X-ray diffractometer to determine mineralogy. Samples were disaggregated and analyzed by using randomly oriented powder mounts. Geochemist's Workbench software was used to calculate mineral saturation using published water chemistry data from some of the springs in the Ten Mile Graben system (Shipton et al., 2004). These predicted saturations were compared with measured mineralogy from this study.
5. Results
5.1. Modern springs
Three modern spring sites (Big Bubbling Spring, Big Bubbling Creek, and Crystal Geyser) were characterized in the field. Spring facies are categorized after the work of Fouke et al. (2000) and exhibit the similar facies relationships to hot spring deposits (Fig. 3). It should be noted, however, that the cold spring deposits are on a much smaller lateral scale and do not have the vertical mound height that Yellowstone hot springs examples typically have.
Water chemistry from Crystal Geyser, Torrey's Spring, and Ten Mile Geyser was modeled with Geochemist's Workbench to determine mineral saturation (log Q/K; Fig. 1; Table 1: water data from Shipton et al., 2004). Waters from all springs are saturated with respect to halite and gypsum; calcite, aragonite, hematite, goethite, and Fe(OH) are undersaturated (Table 1). The minerals present in the Big Bubbling Spring mat are calcite, aragonite, halite, quartz, smectite, goethite, and ferrihydrite (from XRD analysis). Interestingly, gypsum was not detected with XRD in any of the samples analyzed, and although iron minerals are present in all samples, Fe2+ concentrations are very low or undetectable in all water samples.
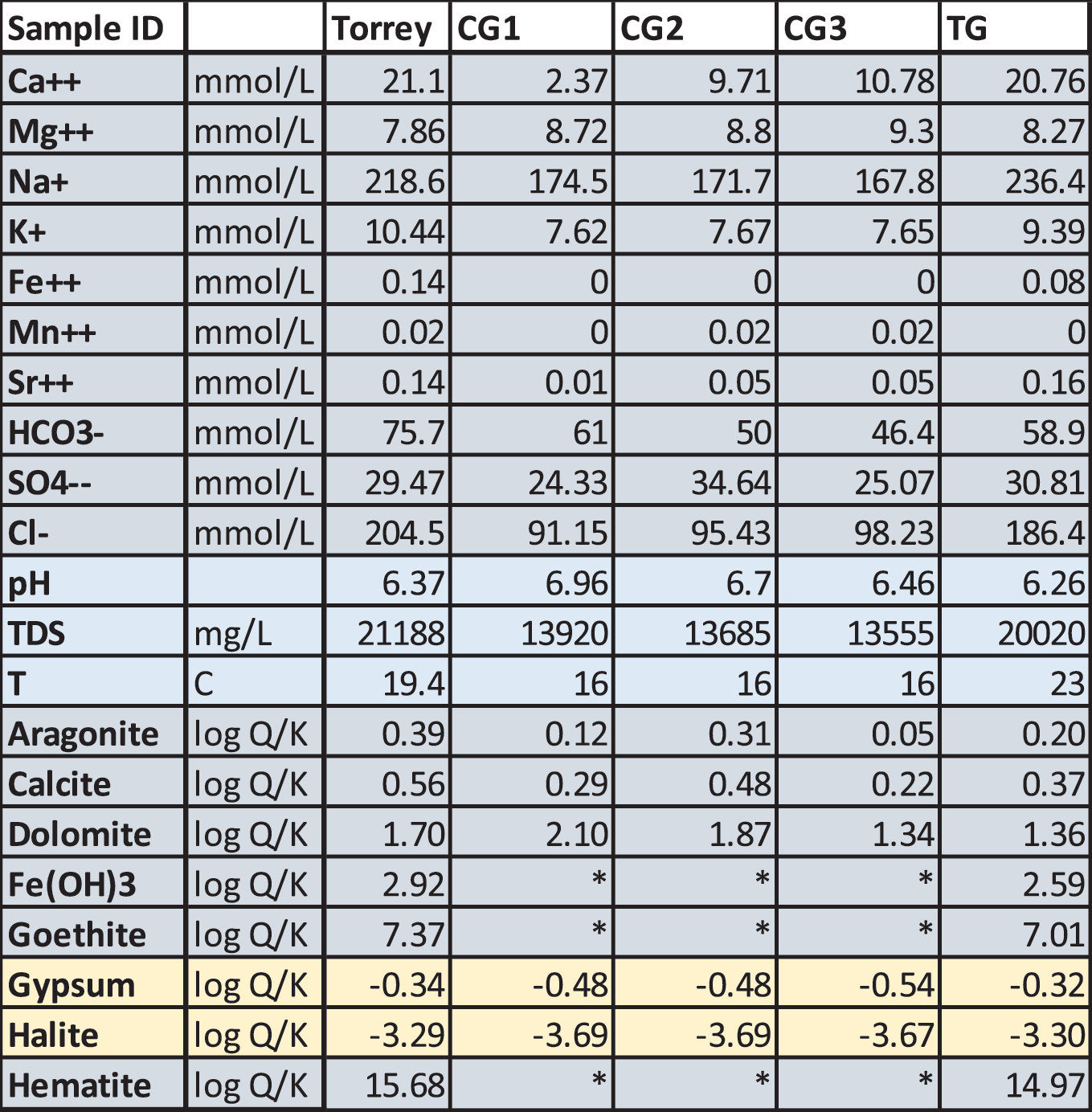
Water chemistry data from Shipton et al. (2004), and samples are from Torrey's Spring (Torrey), Crystal Geyser (CG1, CG2, CG3), and Ten Mile Geyser (TG). *, below detection limits.
Big Bubbling Spring is located in Salt Wash and consists of an active spring surrounded by a microbial mat with laminated tufas and iron minerals, which is approximately 12 m in diameter (Figs. 1 and 3A). Older tufa deposits extend approximately 30 m into a small drainage (informally named Big Bubbling Creek) to the west. Big Bubbling Creek is a west-flowing, ephemeral drainage that winds to the south and west of Big Bubbling Spring. Vents emanate along the sides of the wash, and the water precipitates red tufa crusts that eventually turn gray as the fluid flows downstream and presumably iron is depleted (Fig. 2A). (The only water in the drainage during the summer 2015 field season was water from the springs.) Small vents are present in clusters along the stream banks (Fig. 2B), and terracettes precipitate along the stream that are similar to those at the other springs (Fig. 2C).
Crystal Geyser is a modern spring set on the east bank of the Green River (Fig. 1). An oil well was drilled into it in the 1930s, which unintentionally created the geyser, and it is now the largest CO2-charged cold water geyser in the world (Gouveia and Friedmann, 2006). The area consists of tufa terraces that surround the drill pipe and cascade into the river (Fig. 2D). The terrace is approximately 30 m diameter and 10 m high. Pisoids (∼2–5 mm diameter) are collected in the pools that surround the pipe. These pisoids are not present in any of the pristine springs and likely formed via the agitation of the water from eruption of the geysers. Photosynthetic microbial communities turn some pools green (Fig. 2E).
5.2. Quaternary terraces
The 100 ka terrace is located just southwest of Crystal Geyser (Fig. 1) and consists of a 10 m terrace deposited on the paleo-landscape (Fig. 4A; Pederson et al., 2013). Present in this terrace are delicate structures that consist of clusters of bushy carbonate spheres and terracettes (Fig. 4B, 4C) as well as laminated iron-rich mat structures (Fig. 4D). Secondary carbonate veins also intrude the mat (Fig. 4E). Major minerals present in the tufa are aragonite, ankerite, goethite, and illite/smectite interstratified clay based on XRD analysis.

Tufa terrace, 100 ka. (
A series of older tufa terraces up to 400 ka are also perched on paleo-surfaces along Salt Wash southwest of Big Bubbling Spring (Fig. 1). This study focuses on the 200, 250, 300, and 400 ka terraces (Figs. 1, 4, 5; 200 and 250 ka terraces are not shown on the map because they are right next to the 300 ka terrace). These remnant terraces are less than half a meter thick and approximately 3–4 m in diameter. The 400 ka terrace is heavily eroded and consists of broken pieces of carbonate (Fig. 5C); however, layering and delicate structures (terracettes) are still preserved in all three examples.

Older Quaternary tufa terraces. (
5.3. Jurassic tufa
The Jurassic carbonate deposits in upper Brushy Basin Member are exposed along approximately 5 km of eroded cliff. The main lithofacies consist of a lower massive carbonate (Fig. 6A, 6D) and an upper layered unit (Fig. 6A, 6B) that marks the top of the Brushy Basin Member in the region. Below the lower massive carbonate are lenses of silcretes that are typically less than 1 m thick and 3 m long (Fig. 6A, 6C) and are interspersed in the variegated mudrock that makes up the majority of the Brushy Basin Member throughout the Morrison Formation (see Turner and Fishman, 1991, Potter-McIntyre et al., 2014b, for in-depth descriptions of the Brushy Basin Member).

Carbonate deposit in the Jurassic Brushy Basin Member of the Morrison Formation. (
The lower massive carbonate is ∼15 m thick, although the thickness varies throughout the exposure, and the carbonate grades into a sandstone toward the selvages of the deposit. The bedding of the carbonate is mostly massive and heavily weathered, although 0.5 m beds are present. Brecciated carbonate clasts are mixed with mudrock in sections that are approximately 2 m tall by <0.5 m wide. Mineralogy is composed of calcite, quartz, dolomite, and ferrihydrite.
The uppermost unit is ∼2 m thick and exhibits wavy laminations as well as some wavy bedding (>1 cm thick). These wavy laminations are a primary sedimentary structure and not a result of compaction nor structural deformation, as other strata above and below do not exhibit this structure. The wavy strata are commonly interbedded with chert (Fig. 8B). This unit is only present along ∼1 km of the outcrop where the massive carbonate is thickest. Mineralogy was not analyzed in this sample.
The silcrete lithofacies is present throughout the area where the massive carbonate is exposed, although the concentration of lenses is densest where the massive carbonate is thickest. The silcretes are typically black; however, some silcretes are white. They exhibit horizontal planar bedding (∼1–5 cm; Fig. 6C), and some examples have wavy lamination within the thicker bedding (Fig. 8C, 8D). Mineralogy consists of chert and calcite (from XRD) and clay and barite (observed with SEM).
5.4. Macroscopic features
All the outcrops exhibit some features in common that are retained throughout geologic time. Terracette structures are primary features formed during precipitation and are present in all modern spring sites (Fig. 2C–2E; Fig. 7A, 7B). These features show some degradation but are observed in ancient outcrops including the Jurassic deposit (Fig. 7C, 7D). Laminated mat structures are also primary features that formed during mat precipitation and are present in the modern outcrops and preserved throughout geologic time (Fig. 8). These features are typically iron-rich except in the silcretes. Secondary veins are present in all the older units where aragonite has precipitated from fluid moving through the terraces (in the Quaternary examples), and chert is present in these veins in the Jurassic example (Fig. 8).

Comparison of terracette structures in modern and ancient tufas. (

Comparison of modern and ancient laminated structures. (
5.5. SEM
Evaluation with SEM revealed the presence of distinct microbial forms and associated minerals with unusual habits and/or unexpected composition. In the modern Big Bubbling Spring sample, a cluster of ∼2 μm diameter smooth hollow sheaths was observed and is associated with an anomalous fibrous mineral (Fig. 9A, 10C). EDS analysis showed that the sheaths contain 65 wt % iron (although results are semiquantitative). The fibrous mineral phase is composed of copper and sulfur. A similar ∼2 μm diameter iron-rich sheath is preserved in the 200 ka samples that also exhibits a high iron content (67 wt %; Fig. 9B). The 200 ka sheath is encased in an Fe-rich honeycomb structure (Fig. 9B). Honeycomb structures and possible preserved sheaths (2 μm diameter and 60 wt % iron) were observed in the 400 ka sample (Fig. 9C, 9D). Large crystals (compared to the smaller crystal size in the younger examples) were observed in the 250 ka example in an area where interlocking calcite crystals up to 40 μm (long axis) are present (Fig. 10D).
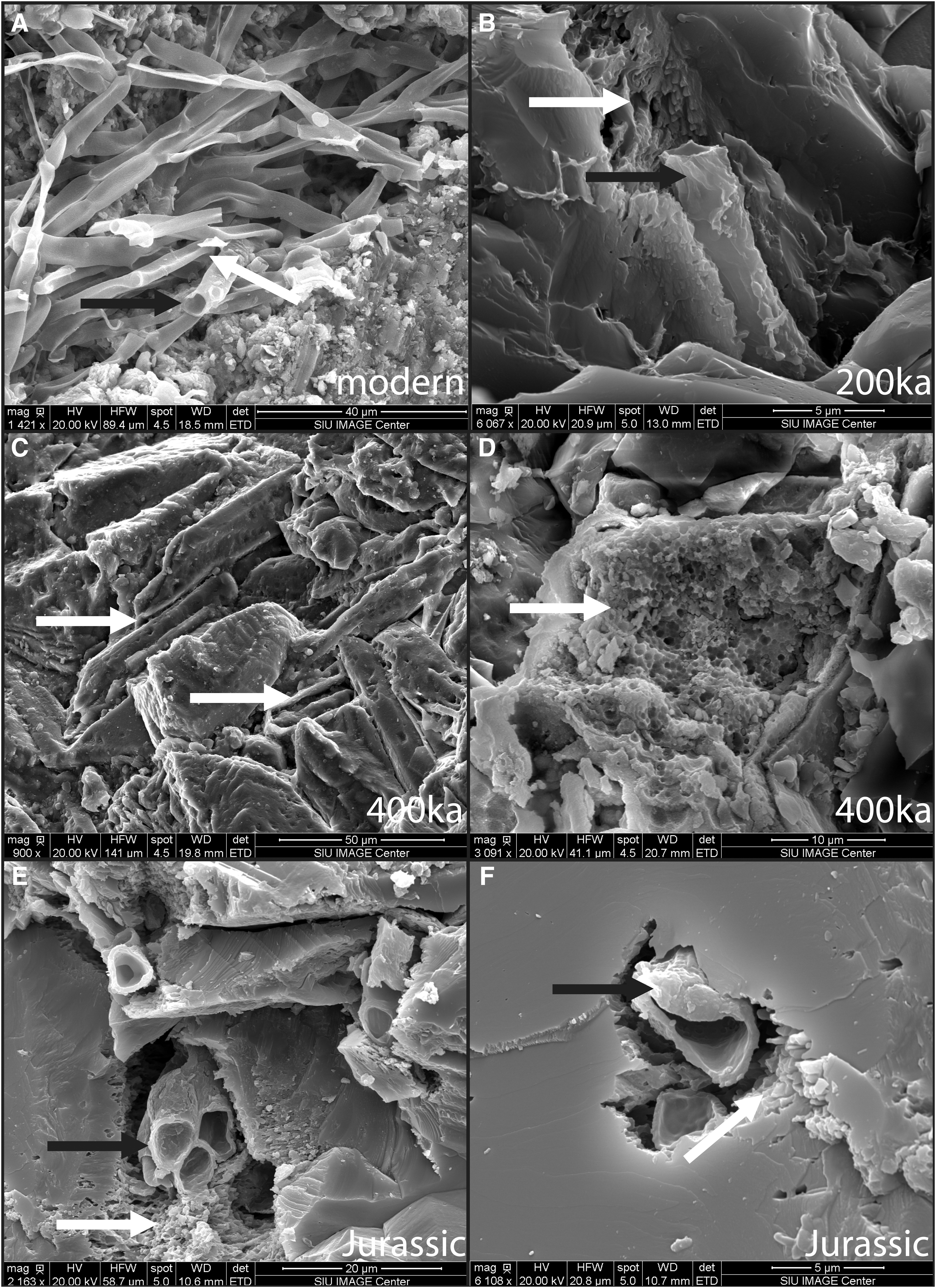
Comparison of modern and ancient sheath structures. (
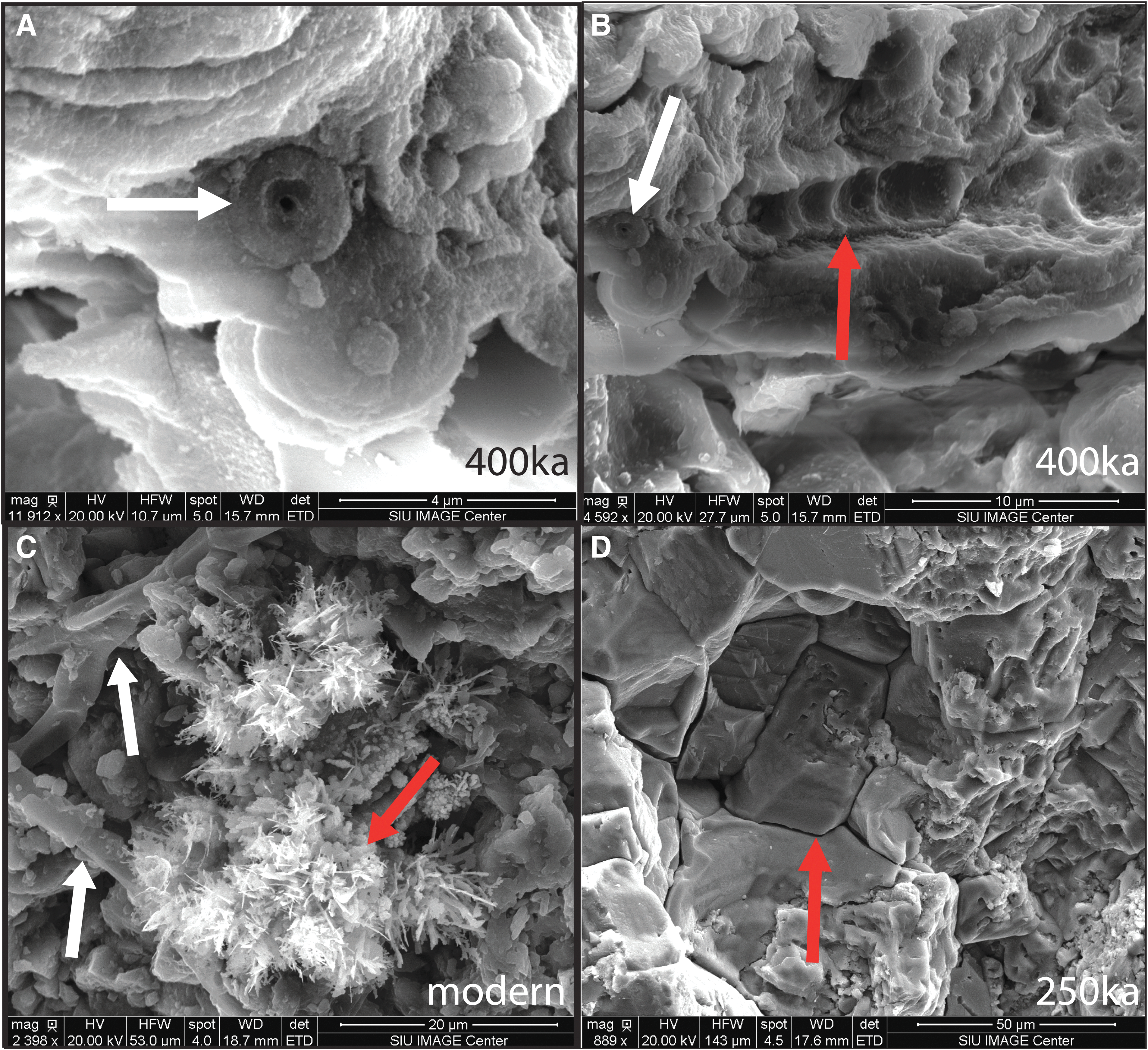
400 ka fossils and trace fossils. (
Other microbial fossils are present in the 400 ka sample: a spherule with a hollow center (Fig. 10A) and a mold of a segmented stalk (Fig. 10B). The spherule and the segmented stalk mold are present within the honeycomb structure. The hollow spherule is ∼2 μm in diameter with a smooth-walled hollow center that is ∼0.5 μm in diameter. The spherule is iron-rich (66 wt %), and arsenic (<1 wt %) and manganese (<1 wt %) are also present in trace amounts. The segmented stalk is a mold of a linear series of ∼2 μm diameter spherical impressions within the honeycomb structure. It is also iron-rich (72 wt %) and contains arsenic (<1 wt %) and manganese (<1 wt %).
The Jurassic tufa example preserves tubes that are 3–4 μm in diameter and sometimes clustered (Fig. 9E, 9F). EDS analysis results suggest that these features are dominated by calcium with only trace amounts of iron. However, EDS is semiquantitative, and the results from this particular analysis are questionable especially for the calcium (Ca = 366–540 wt %, Fe = 2 wt %, Si = 2 wt %, K = 9 wt %). The tubes in the Jurassic example are enclosed by honeycomb features (Fig. 9E, 9F) and surrounded by massive (anhedral) calcium carbonate. The silcrete example exhibits layers of clays, barite, and chert; however, no obvious microbial fossils or trace fossils were observed.
6. Discussion
The Big Bubbling Spring sample contains sheaths that are encased in iron (oxyhydr)oxides. This early mineralization helps preserve the sheath through geologic time, and although some deterioration was observed in particular in the 400 ka example, the microbial fossils are still recognizable.
Similar sheaths with surrounding honeycomb features are preserved in the older samples (200 ka, 400 ka, and Jurassic). Honeycomb features surround all these sheaths. Other microbial fossils are also preserved in the 400 ka samples such as a spherical cell with a hollow center and the segmented stalk mold. These features are preserved within the honeycomb structure, which suggests that the honeycomb features are likely molds of microbial cells (possibly cyanobacteria based on the morphology). This association suggests that both body fossils and trace fossils (molds of microbes) can be co-preserved.
In the 250 ka and Jurassic samples, large crystals have replaced the smaller crystals, and we interpret this as larger crystals forming at the expense of smaller ones because larger crystals are energetically favored (Ratke and Voorhees, 2002). This process is called Ostwald ripening and is observed in geologic environments (Steefel and Van Cappellen, 1990; Potter et al., 2011). In the Jurassic sample, Ostwald ripening has produced anhedral carbonate minerals that surround the sheaths and associated honeycomb features. This entombment of permineralized body fossils by large, massive crystals has likely preserved these features on mega-annum time scales, whereas long-term weathering in other examples, such as in the 400 ka example, has allowed the sheaths to deteriorate.
The presence of unexpected and trace minerals associated with microbial features is also documented. In the modern sample, a delicate fibrous mineral associated with the sheaths is most likely the hydrated salt, calcanthite (CuSO4•5H2O). Interestingly, no copper and low SO4 (relative to CO3 and Cl) are reported in the spring waters (Shipton et al., 2004). The presence of barite within the silcretes suggests that the Jurassic deposit was a hot spring–fed system.
6.1. Macroscopic spring features
Delicate features associated with specific spring facies were observed in older terraces such as terracettes (proximal slope) and bushy spheres (pond facies), and these facies are therefore identifiable throughout geologic time. These features (terracettes and bushy spheres) have been proposed to be biogenic in modern spring systems (Fouke et al., 2000); therefore, it is important to note that these are preserved during diagenesis on geologic time scales. Secondary processes such as vein precipitation during early diagenesis and Ostwald ripening likely do affect and degrade some of these macroscopic features; however, these processes do not completely destroy macroscopic identifying features. Moreover, secondary veins can be added to criteria used to identify spring-fed microbially mediated tufas in the rock record.
Distinguishing features observed in modern and Quaternary tufas such as terracettes, layering, and secondary veins are also present in the Brushy Basin Member of the Morrison Formation, suggesting that this carbonate is a spring-fed system. Brecciated carbonates mixed with mudstone are possibly paleokarst features, or the result of collapsed tufa towers filled with siliciclastic sediment, such as those observed in spring-fed lacustrine settings like Mono Lake. Additionally, the gradation of the massive carbonate into a sandstone with features consistent with a shoreline (parallel, slightly dipping laminae) suggests that standing water was present to the northwest of Lake T'oo'dichi' and this northwestern lake had a very different water chemistry from Lake T'oo'dichi, which is exclusively siliciclastic. The uppermost unit contains primary wavy stratification interbedded with chert veins and terracettes; therefore, this unit is interpreted as a microbialite. The presence of silcretes that increase in abundance near the thickest parts of the massive carbonate provides additional evidence that this carbonate was spring-fed, and the presence of barite suggests that the spring was hot.
6.2. Two types of biomineralization
Biomineralization caused by metabolism in the modern samples can explain why iron minerals are precipitating despite the nearly undetectable concentrations of iron in the water. Calculations of log Q/K using published springwater compositions suggest that the precipitates should be gypsum and halite. The presence of hematite (and even ferrihydrite) suggests that microbial metabolisms are participating in the precipitation process, because these minerals would not precipitate abiotically from the concentrations of iron present in the water. Therefore, even abundant oxidized iron minerals may possibly function as a biosignature, especially in undersaturated environments such as Archean marine environments that were oxygen limited.
Biomineralization on the charged surfaces of the microbes is another mechanism that is occurring at the springs. This is evidenced by the MISS such as laminae and bushy spherules. These features form by Ca and/or Mg adsorbing to cell surfaces, which are negatively charged at circumneutral pH; then CO3 binds to the cations, and carbonate minerals precipitate (Dupraz et al., 2009; Corkeron et al., 2012). The microscopic honeycomb textures are likely formed in this manner as minerals precipitate around cells, and then the cells degrade leaving molds (trace fossils). This process is evidenced by a segmented stalk mold and the remains of a hollow cell preserved within the honeycomb feature.
Although some microbes, such as cyanobacteria, may decompose in oxidizing surface conditions, this study shows that some microbial fossils and trace fossils such as honeycomb textures can persist owing to encasement by iron (oxyhydr)oxides and/or by entombment by Ostwald ripening. For example, early permineralization of sheaths renders this microbial population particularly resistant to destruction and degradation during early diagenesis. Other features such as sheaths and honeycomb trace fossils were preserved in the Jurassic example because Ostwald ripening entombed these features, which shielded them from diagenetic alteration and destruction. Additionally, cyanobacteria may actually encourage this entombment by increasing pH via photosynthesis and creating an environment conducive to calcium carbonate precipitation (e.g., McConnaughey and Whelan, 1997; Golubic et al., 2000; Dittrich and Sibler, 2010).
7. Conclusions
This study furthers our understanding of biosignature preservation in the rock record by demonstrating that Fe-oxide and carbonate biosignatures are well preserved on both millennial and geologic time scales despite diagenetic changes. The data highlight two distinct methods of biosignature formation: (1) minerals are precipitating from undersaturated solutions owing to metabolic activity of cells, and (2) mineral precipitation on charged cell surfaces. Microbes produce trace fossils by creating an environment conducive to mineral precipitation, and in turn, the presence of these minerals helps preserve these features. Although diagenesis can destroy or severely degrade biosignatures and especially microbial fossils, some fossils are preserved because Ostwald ripening encases fossils and limits diagenetic alteration. This field site preserves an excellent record with which to understand the taphonomy of microbial fossils preserved in discrete time slices in the geologic record. Recognizing spring-fed, biogenic tufas is crucial for astrobiological research and the search for life on Mars.
Footnotes
Acknowledgments
This research was funded by a grant to Potter-McIntyre by the American Chemical Society Petroleum Research Fund and by start-up funds from Southern Illinois University.