
Abstract
Life on Earth is found in a wide range of environments as long as the basic requirements of a liquid solvent, a nutrient source, and free energy are met. Previous hypotheses have speculated how extraterrestrial microbial life may function, among them that particle radiation might power living cells indirectly through radiolytic products. On Earth, so-called electrophilic organisms can harness electron flow from an extracellular cathode to build biomolecules. Here, we describe two hypothetical mechanisms, termed “direct electrophy” and “indirect electrophy” or “fluorosynthesis,” by which organisms could harness extracellular free electrons to synthesize organic matter, thus expanding the ensemble of potential habitats in which extraterrestrial organisms might be found in the Solar System and beyond. The first mechanism involves the direct flow of secondary electrons from particle radiation to a microbial cell to power the organism. The second involves the indirect utilization of impinging secondary electrons and a fluorescing molecule, either biotic or abiotic in origin, to drive photosynthesis. Both mechanisms involve the attenuation of an incoming particle's energy to create low-energy secondary electrons. The validity of the hypotheses is assessed through simple calculations showing the biomass density attainable from the energy supplied. Also discussed are potential survival strategies that could be used by organisms living in possible habitats with a plentiful supply of secondary electrons, such as near the surface of an icy moon. While we acknowledge that the only definitive test for the hypothesis is to collect specimens, we also describe experiments or terrestrial observations that could support or nullify the hypotheses. Key Words: Radiation—Electrophiles—Subsurface life. Astrobiology 18, 73–85.
1. Introduction
B
Of interest to astrobiology are hypothesized deviations from the oxygenic photosynthesis described in Eq. 1. One hypothesized deviation includes the photolytic oxidation of aqueous chloride, yielding halogen oxides or halogens, rather than molecular oxygen, in what has been termed chlorinic photosynthesis (Haas, 2010):
More common hypothetical deviations from terrestrial photosynthesis include variations in captured wavelengths of light. For example, capture of photons with near-IR wavelengths could enable photosynthesis in deep-sea hydrothermal environments otherwise dark at visible wavelengths (Perez et al., 2013). The wavelength of light captured by extraterrestrial photosynthetic organisms has also been hypothesized to depend on stellar spectral type, leading to different absorptions in the incident light spectra of extrasolar planets (Kiang et al., 2007).
Although photosynthesis dominates the net primary production of biological carbon, it is not the only energy-gathering route to build and sustain biomass: chemosynthetic organisms, which account for less than 0.1% of global oceanic primary production (Middelburg, 2011), mediate a large diversity of chemical processes. Such processes include the oxidation of molecular hydrogen (H2) to water (Drobner et al., 1990); hydrogen sulfide (H2S), elemental sulfur (S), or thiosulfate (S2O3 2-) to sulfate (SO4 2-) (Suzuki, 1999); ammonia (NH3) to nitrate (NO3 2-) (Nicol et al., 2008); ferrous iron (Fe2+) compounds to ferric iron (Fe3+) compounds (Emerson and Weiss, 2004); and others.
In addition to photosynthetic and chemosynthetic organisms, some organisms can also directly exchange electrons with extracellular metal ions in a process termed extracellular electron transfer. Certain organisms, like Methanobacterium palustre and Desulfovibrio vulgaris, can accept or donate electrons; they are sometimes labeled electricigens, anode-respiring bacteria, or exoelectrogenic bacteria if found at the anodes, and electrotrophs if found at the cathodes (Schroder et al., 2015). Metal-reducing bacteria in the genera Shewanella and Geobacter are hypothesized to use a chain of c-type cytochromes, electron transfer proteins found in the membranes of mitochondria and all bacteria, to donate or transfer metabolic electrons to areas outside the organism (reviewed in Richardson et al., 2012; Shi et al., 2009; Rosenbaum et al., 2011; Semenec and Franks, 2015).
This ability to use a current for growth has likely evolved multiple times. Semenec and Franks (2015) pointed out that hydrogenases are also hypothesized to help with extracellular electron transfer in electrotrophs. In addition, the capacity to exchange electrons is found across phyla (Shewanella is a Gammaproteobacterium, whereas Geobacter is a Deltaproteobacterium); Shewanella genes coding for cytochrome c proteins capable of utilizing electrical currents have been found to be paralogs (Brutinel and Gralnick, 2012), that is, resulting from convergent evolution; and electrical currents can also be utilized by mitochondria (Gupta, 1998; Butler et al., 2010).
Direct electron exchange is found in both natural and artificial systems. Within marine sediment communities, there is evidence to suggest that filamentous bacteria transport electrons over centimeter distances (Pfeffer et al., 2012). Direct exchange of electrons is also exploited in artificial systems such as microbial fuel cells to produce electricity from the degradation of organic matter (Semenec and Franks, 2015). This process, which mimics respiration, can be described by the reverse of Eq. 1. Geobacter are known to accept electrons from a cathode (Gregory et al., 2004). In the past decade, biocathodes have been used in research involving biofuel/commodity production and wastewater remediation (Semenec and Franks, 2015). Even mitochondria have been found to transfer electrons and can be used in designing electrochemical cells (Arechederra and Minteer, 2008; Hubenova and Mitov, 2015).
But what are the potential sources of electrical currents on extraterrestrial bodies, and could they power extraterrestrial life? To examine that question in more detail, we briefly review some relevant hypotheses and observations. Recently, the Workman-Reynolds effect, wherein a voltage is produced through the separation of cations and anions in quickly freezing saltwater, has been hypothesized to support microorganisms on icy moons through the production of H2 and molecular oxygen (O2) from the electrolysis of water (Johnson et al., 2014). Radiation-driven ecosystems have been considered too, with the role of radiation often being to provide substrates (radiolytic products) useful for biosynthesis. For example, Chyba and Phillips (2001) hypothesized the production of formaldehyde (H2CO) from irradiated ice on Europa, which could then be used in heterotrophic metabolism:
which is known to support at least one type of soil bacterium on Earth (Chyba and Phillips, 2001). Atri (2015) recently suggested that cosmic rays could power life on planets like Mars through either formation of secondary particles (muons) which could power organisms through radiolysis, formation of secondary particles that could lead to the formation of organics, or direct capture of high-energy photons with pigments such as melanin. On Earth, ionizing radiation has been observed to enhance the growth of some fungi living in Chernobyl, Ukraine, by causing changes in the electronic structure of melanin, a pigment produced by the fungal cells (Dadachova et al., 2007).
Here, we hypothesize two mechanisms involving radiation by which organisms could use secondary electrons from ionizing radiation or beta particles for growth: (1) The use of electrical currents resulting from secondary electrons flowing through a putative electron capture compound (playing a role analogous to the cytochrome c electron carrier or similar proteins), much like in terrestrial electrotrophic organisms. We term this mechanism “direct electrophy.” (2) A process in which secondary electrons excite a fluorescing molecule, either generated by an organism or naturally present in the environment, to drive photosynthesis. We term this putative process “indirect electrophy” or “fluorosynthesis.”
With these two mechanisms, we aim to show that organisms harnessing free electrons such as secondary electrons may be of astrobiological interest.
Such hypothetical organisms would require an environment with both energy and particle fluxes sufficient to produce enough low-energy secondary electrons to support their metabolisms. If the distance between incoming particles is much larger than the size of the organisms, then a direct interaction between a particle, either primary or secondary, and an organism might be too rare to supply energy even if the energy flux itself is sufficient. Furthermore, particles of energy above a few electronvolts would likely mostly just pass through the organisms and ionize molecules within the path of the particle (Mozumder, 1999). A variety of environments could fulfill these requirements. Galactic cosmic radiation provides high-energy particles of at least 10 MeV, but a relatively low particle flux of about 1 m−2 s−1 sr−1 MeV−1 (Fanselow et al., 1969) delivers this energy in erratic bursts rather than in a continuous supply. Each primary particle impact results in a cascade of secondary electrons of energy typically between 1 and 10 eV. If this cascade provides a sufficient flux of secondary electrons, then the discussed organisms might be able to use galactic cosmic radiation as a source of energy. Planetary magnetospheres could also provide a sufficient energy and flux. For example, the surface of Jupiter's moon Europa receives an electron energy flux of 6.2 × 1010 keV cm−2 s−1 and a particle flux of 1.8 × 108 cm−2 s−1 (Cooper et al., 2001). To demonstrate the adequacy of such energy and particle fluxes, we present calculations of the biomass that could be supported by direct electrophy and the amount of photons that could be created by fluorosynthesis in Sections 2 and 3.
2. Direct Electrophy
While the direct use of particle radiation through secondary electrons has not been discussed before, the indirect use of particle radiation as an energy source for life has been previously hypothesized. For example, it has been hypothesized that organisms inhabiting the ocean of an icy moon may derive energy from the electrolysis of water driven by a global electrical current created by incoming particle radiation into a conductive ocean (Reynolds et al., 1983). Here, we hypothesize that organisms might derive energy from electrical currents using an electron capture compound powered by secondary electrons generated from particle radiation. An electron capture compound is defined as any biomolecule involved in electron-transfer reactions similar to the ones undertaken by terrestrial cytochrome c proteins. We call this putative mechanism “direct electrophy.”
Direct electrophy requires attenuation of high particle energies before particles can be used by organisms to generate the energy required to fix carbon, since particles of too high energy can damage the cell (Fig. 1). On an icy body, impacting high-energy particles would be stopped within 0.6 mm of the surface (Cooper et al., 2001). However, the resulting cascade of secondary electrons (energy 1–10 eV) could be exploited as a biological energy source if the organism is able to survive the harsh conditions of an icy moon's near subsurface (Section 4). The number of secondary electrons n(x) generated at a distance x from the entry surface per unit path length dx is (Simon and Williams, 1968; Kimura and Mann, 1999)
Here, ɛ is the energy deposited by the primary electron into the solid target to produce one secondary electron. In ice, ɛ ≈ 25 eV (Suszcynsky et al., 1992). dE/dx is the energy loss of the primary electron per unit path length. We have made the same simplifying assumptions as Scholtz et al. (1996), such as neglecting angular scattering. Simon and Williams (1968) pointed out that energy loss is relatively constant throughout the primary electron penetration depth R:
Thus, the number of secondary electrons generated upon impact is roughly proportional to the primary electron energy E
p. Combining Eqs. 4, 5, and the penetration depth for an electron, R ≈ (1 mm) (1000 kg m−3/ρ) (E
p/1 MeV)1.5 (Simon and Williams, 1968; Kimura and Mann, 1999), we obtain an expression for the number n(x) of electrons of energy 1–10 eV produced in ice (ρ ≈ 1000 kg m−3) per unit length by secondary emission of a primary electron of energy E
p:
Increasing E p has competing effects on n: a primary electron of higher energy can produce more ∼1 eV electrons but will also penetrate deeper and dissipate more energy in the process due to exponential attenuation; the latter effect dominates. This electron quantity value can then be converted to an electrical current value by using the definition of the ampere as a conversion factor. An example calculation is shown in Table 1.
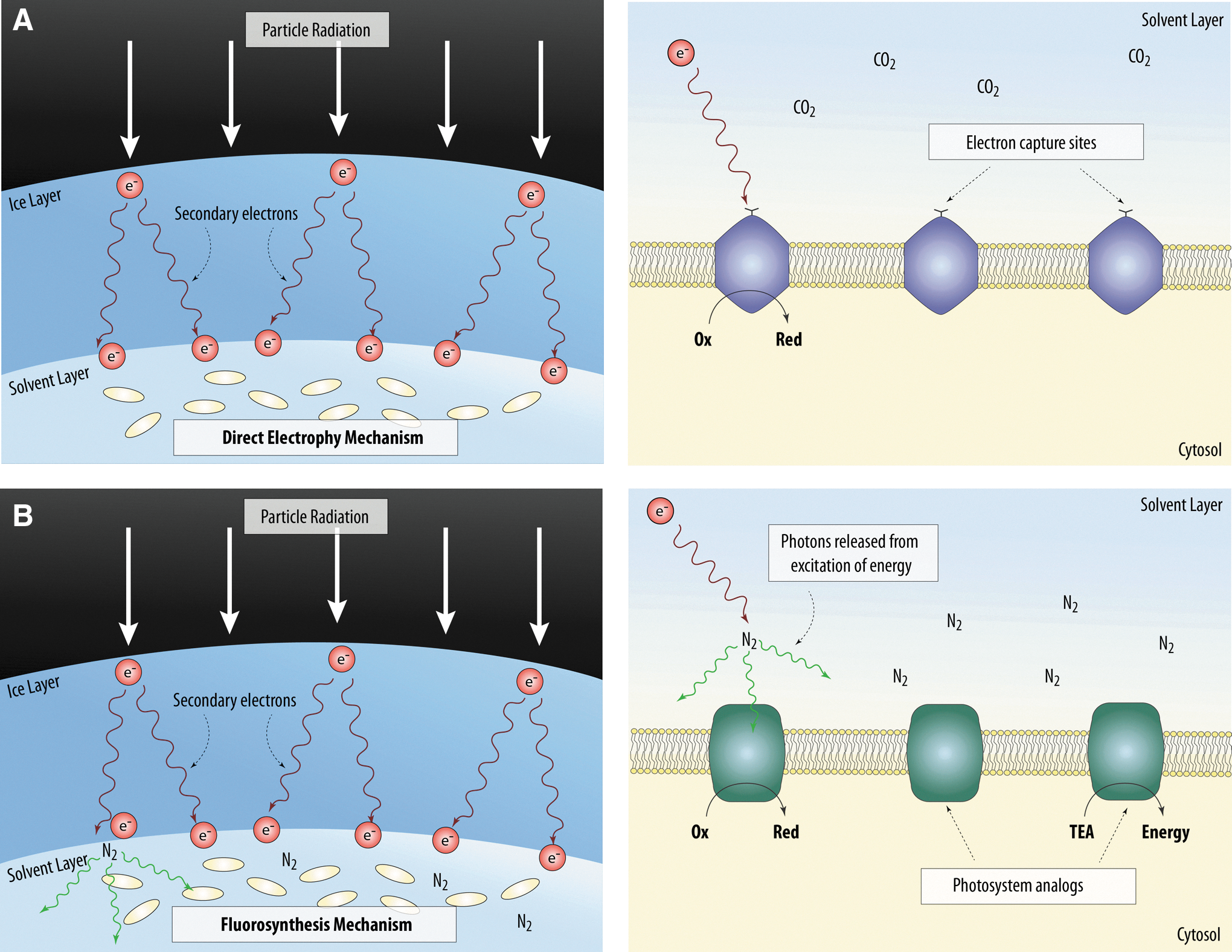
Schematics of how the putative direct electrophy and fluorosynthesis processes might provide metabolic energy to microbial cells. (
The penetration depth and stopping powers were calculated using NIST's EStar program (Berger et al., 2005). Higher-energy particles penetrate deeper into the ice and are responsible for the creation of a larger number of secondary electrons. These simple calculations neglect the energy of secondary electrons that would produce their own cascade of secondary electrons. The supply of electrons in an icy moon's near surface seems to provide sufficient energy to build biomolecules, assuming that nutrients do not limit organismal growth. For comparison, the jovian moons experience a particle flux from 106 to 108 particles cm−2 s−1 (Cooper et al., 2001), and the average biomass density at the Earth's surface is 0.4 g cm−2 (Whitman et al., 1998; Landenmark et al., 2015).
The water-ice calculations shown in Table 1 are relevant to icy bodies such as Europa and Enceladus (Cooper et al., 2001; Brown et al., 2006), and those for nitrogen-dominated ice are similar to the water-ice calculations and are relevant to objects such as Triton and Pluto (Cruikshank et al., 1984; Stern et al., 2015). From the electrical currents alone, one could predict that life could exist in such environments since electrical currents of about ∼10−5 mA are used to sustain terrestrial electrophiles (Gregory et al., 2004); however, there are other considerations to be made.
First, if currents or voltages are too strong, they would probably be deleterious to an organism. A high current through an organism could overwhelm the cell's ability to counteract ion fluxes or redox reactions. A high voltage drop across an organism could exceed the potential difference needed to drive chemical change, such as membrane disruption, protein refolding, or DNA strand separation. Life on Earth has evolved means of channeling photoelectrons of a few electronvolts, that is, comparable to the kinetic energy of secondary electrons (Suszcynsky et al., 1992) but nearly ionizing (Mozumder, 1999). Electrons can be trapped and transferred through the aromaticity of molecules such as the porphyrin rings of chlorophyll and cytochrome compounds, which are also used in molecular electronics (Urbani et al., 2014), and one could imagine then that low-energy secondary electrons could be trapped in a similar way. Electrons captured at active sites of metalloproteins or other conductive biomolecules could then be transferred even further using known biochemistry or at least chemicals analogous to terrestrial chemistry. This can include organic nanowires as described in Leung et al. (2013), metalloproteins (Levar et al., 2012), or even synthetic biological systems like metalloprotein-prion systems (Altamura et al., 2016).
Second, pressures and temperatures just below the surface of any world with a tenuous atmosphere would not be welcoming for any water-based life. Direct electron capture strategies assume the microorganism would be close (<1 cm) to a planetary body's surface with our simple calculations. Could the initial current be sustained deeper to reach more welcoming conditions? This would depend largely on the conductivity of the medium utilized. Water ice, for example, is an insulator that would make it impossible for a current produced on the surface to reach a subsurface ocean (Reynolds et al., 1983), unless preferential trapping of ions (Johnson et al. 2014) leads to a buildup of positive charges in the ice, which are able to interact with near-surface secondary electrons to sustain a minor electrical current. The planetary body could thus be modeled as two large plates: one negative from the creation of secondary electrons and one positive plate if one assumes the trapping of cations. Ultimately, in the case of planetary magnetospheres and as noted by Reynolds et al. (1983), the initial electrical current in the impact zone of the incoming particles is sustained by the sputtering of atoms and molecules that then can get ionized and trapped in a planetary magnetosphere and then come back to the surface and create secondary electrons.
The relevance of these calculations to Solar System objects, such as Pluto and the icy Galilean moons, is shown in Fig. 2. Europa has been observed to receive a flux of about ∼1.8 × 108 particles cm−2 s−1 (Cooper et al., 2001). Such a flux would result in large initial currents that would probably sterilize the surface, but further down, the current could drop to a point suitable for direct electrophy. Other icy moons, such as Enceladus and Triton, receive fewer electrons due to Saturn's lower magnetospheric particle fluxes (Jones et al., 2006) and a shielding ionosphere (Bagenal, 1997), respectively. On these worlds, direct electrophy could still be driven by secondary electrons generated from high-energy photon irradiation, although photons do not create as many secondary electrons in a given substrate as particle radiation does (Baumstark-Khan and Facius, 2006).
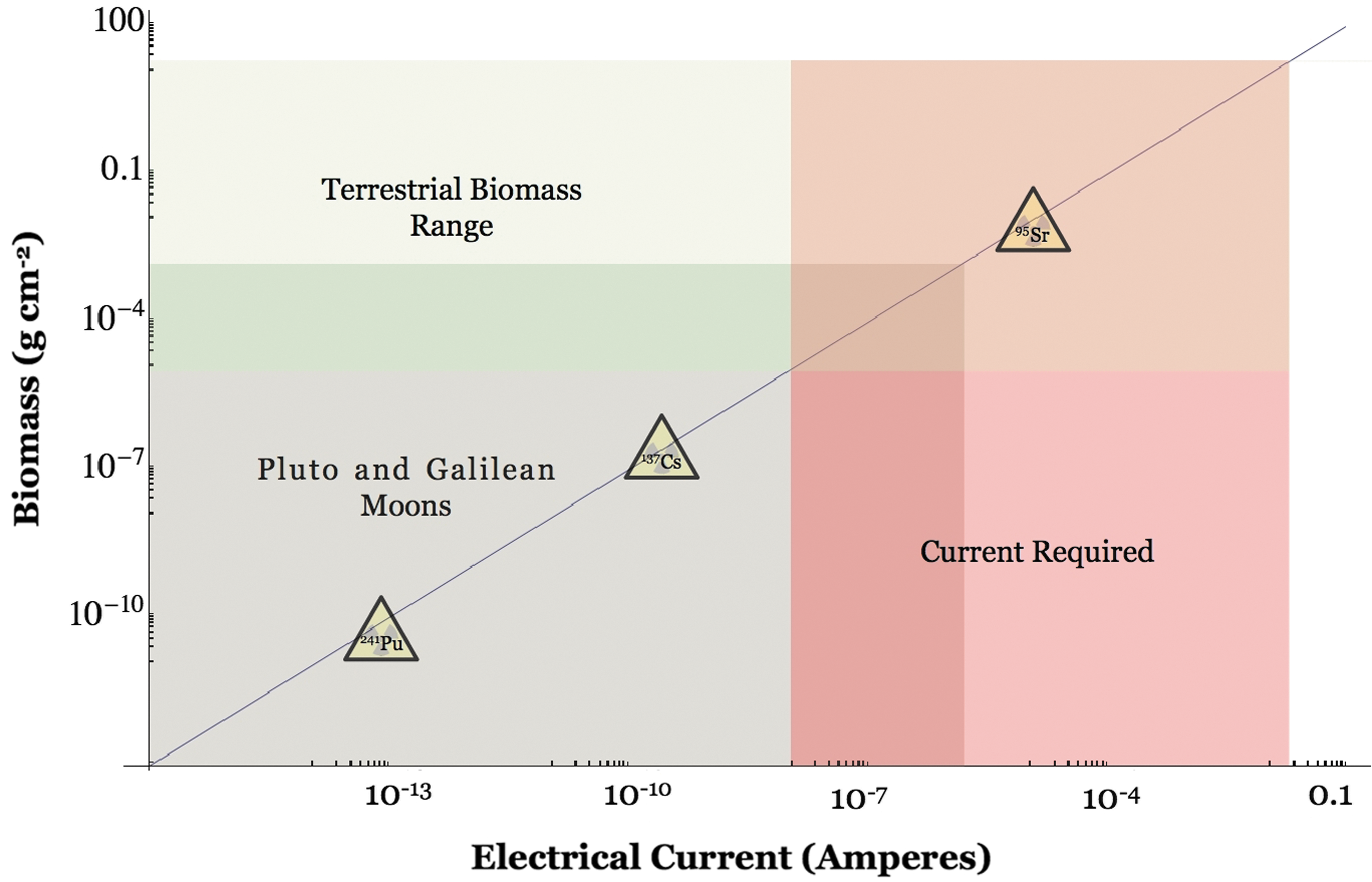
Power-law relationship between maximum biomass sustained by a given current and current intensity (Eq. 7). The estimated terrestrial biomass range and corresponding range of minimal currents that would be needed to support this biomass are shown in green and red, respectively. Currents resulting from secondary electron fluxes estimated for Galilean moons and the Pluto system are plotted in gray along the maximum biomass density line; at the high end, there is a significant overlap with terrestrial biomass ranges. Along this line are also plotted radionuclides found at nuclear plants or natural reactors (yellow triangles).
Our simple calculations assume an equal energy for all incoming particles. In reality, there is a spectrum of exogenic and planetary magnetospheric particle energies, and a cascade of secondary electrons is created. In magnetospheres, for instance, particle energies follow a Maxwellian thermal core at lower energies, with a power-law distribution in the higher energy tail (e.g., Cooper et al., 2001). Our simplified calculations could pertain to a subset of particles having a narrow energy range.
How much biomass could be maintained with this current supply? We make estimates by assuming that this current drives chemical reactions away from equilibrium, yielding products that can then be used as metabolic substrates. Following Seager et al. (2013), we first estimate the energy rate needed for a microorganism to build biomass as the product ΔG*r. Here, ΔG is the Gibbs free energy released or absorbed by the reaction that produces reducing or oxidizing equivalents to fix carbon and hence build biomass, in J mol−1; and r is the rate at which species are consumed or generated through this reaction, in mol g−1 s−1. We express ΔG as -n*F*E, where n is the number of electrons transferred in the reaction, F is the charge of one mole of electrons (Faraday's constant), and E is the reaction's electrochemical potential. Seager et al. (2013) expressed r as the ratio of the source flux F
source of a microbial metabolic product, in units of mol m−2 s−1, to a surface biomass density Σ
B in g m−2. In our case, F
source can be understood as the source flux of electrons whose flow generates a current density J = n*F*F
source with units A m−2, providing the energy needed to produce reducing equivalents to fix carbon and build biomass. The maximum surface biomass density that can be supported can then be derived by comparing this energy rate for growth, ΔG*r = −J*E/Σ
B, to the minimal energy rate needed to maintain active biomass, P
me (in J g−1 s−1):
The quantity P me is independent of the chemistry used to generate that maintenance energy, but it still depends on temperature. Seager et al. (2013), and references therein, provide an empirical, Arrhenius-type relationship between P me and temperature. Using their parameter values for aerobic organisms at 270 K yields P me = 1.46 × 10−3 J (g biomass)−1 s−1. Values for E depend both on the standard electrochemical potential of a given reaction, E°, and on the relative abundances of products and reactants (reaction quotient Q) as E = E° − R G T/nF * ln Q, where R G is the gas constant. Values for E° are typically of order 1 V and negative for reactions that do not spontaneously take place under reference conditions but that can be driven by any energy source. (For example, E° = −1.23, −0.731, and −0.174 V for the reactions H2O = H2 + 0.5O2, C2H6 = C2H4 + H2, and H2S = S + H2, respectively.) Representative current densities J are provided in Table 1 for the near-surface of icy moons irradiated in planetary magnetospheres; they are of order 10−11 to 10−8 A cm−2. Primary electron fluxes at Europa from Fig. 1 of Cooper et al. (2001), combined with information from our Fig. 2, yield a wider range of current densities: 10−12 to 10−6 A cm−2. Together, these values for P me, E, and J yield estimates of maximum sustainable biomass Σ B on the order of 10−8 to 10−5 g cm−2 (Fig. 2). This is many orders of magnitude lower than biomass densities in tropical forests (≤1 g cm−2 of active biomass; e.g., Luckman et al., 1998) and comparable to biomass densities in material from subglacial Lake Vostok in Antarctica entrained in overlying ice (∼100 cells mL−1; Christner et al., 2006; i.e., 107 cells cm−2 or 10−5 g cm−2 assuming integration over a ∼1 km water column).
Planetary radioactive materials also provide endogenic sources of free electrons in the form of β- decay. The long-lived radionuclides 40K, 232Th, 235U, and 238U are dominant sources on Earth (Clark et al., 1966; Armstrong and Harmon, 1981; Van Schmus, 1995) and likely on other planets as well. These radionuclides are enriched in the mantle and crust compared to the bulk Earth. K is largely present in granites and shales, and Th and U are present in trace minerals (Van Schmus, 1995). In addition, short-lived radionuclides such as 26Al, 60Fe, 10Be, or 107Pd may have been substantial free electron sources early in Solar System history (Walter and Trønnes, 2004; Arnould et al., 2006). Nuclear waste is also a β- decay-rich environment. The uranium content in nuclear waste is typically on the order of 100 ppm in contaminated sediment and 1000 ppm in sludge from uranium processing plants (Francis, 1994). Electron fluxes from radioactive decay and corresponding currents that could be harnessed by direct electrophy are shown in Table 2. Long-lived radioisotopes do not produce sufficient electron fluxes to sustain any significant electrical currents, and the short-lived radionuclides preclude their involvement in bioenergetic pathways. This may explain why direct electrophy from radiation sources did not evolve on Earth. Only free electron fluxes from the decay of nuclides found in radioactive waste and natural nuclear reactors could sustain appreciable biomass densities, up to 10−3 g cm−2 (Eq. 7 and Table 2), but only on decadal timescales. Note that such environments seem to enhance the growth of some fungi as has been shown in Chernobyl, Ukraine (Dadachova et al., 2007; see Section 4). To our knowledge, biomass densities sustained solely by radioactivity have not been reported.
Most of the radionuclides would not contribute significantly to the production of biomass. Interestingly, 137Cs could; it is present in Chernobyl, Ukraine, where radiotrophic fungi have also been found. The low biomass values for other radionuclides help explain why radiotrophic electrophilic organisms are not common on Earth: there were not many chances for them to evolve.
IAEA Nuclear Data Services (2014). bAssuming chondritic material (Kita et al., 2005). cWolfram (2014). dArnould et al. (2006). eWadhwa et al. (2006). fGounelle and Meibom (2008). gHuss et al. (2009). hWalter and Trønnes (2004). iMcKeegan et al. (2000). jAssuming 750 kg waste yr−1 for a 1000 MWe reactor (World Nuclear Association, 2012). kGauthier-Lafaye et al. (1996).
A final consideration is the limitation of microbial activity due to nutrient supply. This point lies outside the scope of this paper; the calculations presented here were performed assuming that the limiting factor is energy supply.
3. Indirect Electrophy (Fluorosynthesis)
In the second proposed mechanism, high-energy particles impacting an icy surface produce a cascade of secondary electrons, whose kinetic energy (<30 eV) could excite a substrate within or outside the putative organism; the excess energy would then be released by fluorescence as a photon (Prince et al., 1976). These photons can then be used by a photosystem analogue within the putative organism to drive photosynthesis (Fig. 1). This process, which we term “indirect electrophy” or “fluorosynthesis,” is similar to a hypothesis that postulated that extraterrestrial organisms might be able to use photons created by ionizing radiation in the atmosphere as an energy source (Smith et al., 2004).
A model that places the fluorescing molecule within the organism is a closer analogy to terrestrial photosynthesis, as chlorophyll molecules are located inside cells. This requires prior electron capture as in direct electrophy, but the added steps of fluorescent molecule excitation and subsequent capture of the photon emitted make this intracellular fluorescence model less efficient than direct electrophy. However, fluorosynthesis need not require cellular electron capture, as it could exploit fluorescence of the substrate surrounding the organism. Such a substrate could be water ice, which under irradiation by >10 eV electrons emits near-UV/visible photons primarily at 306 and 400 nm (Prince et al., 1976). Irradiated solid nitrogen also fluoresces (McLennan and Shrum, 1924); to our knowledge a conversion factor relating photon fluxes to energy input has not been reported for solid N2, but simple laboratory experiments can show that the fluorescence of N2 ice is much stronger than that of water ice. In the calculations outlined below, we assume that the fluorescent substrate is water ice, bearing in mind that this assumption provides a lower limit for the photons that can be produced.
From the data of Prince et al. (1976), we first estimate how many 306 and 400 nm photons are emitted in water ice per incoming electron. Below a few electronvolts, the electron energy threshold for fluorescence in ice, no photons are emitted. With electrons of energy 10 eV and an electron source intensity of 0.1 mA * (10 eV/100 eV)3/2 = 3.2 μA, which corresponds to a flux of 2 × 1013 electrons s−1, the fluorescence emission is ≈3 photons s−1. Thus, the yield is 1.5 × 10−13 emitted photons per incoming 10 eV electron.
Europa's surface receives a primary electron energy flux of 6.2 × 1010 keV cm−2 s−1, for a corresponding primary electron number flux of 1.8 × 108 cm−2 s−1 (Cooper et al., 2001). Although the number of electrons in a given energy range decreases exponentially with energy (Cooper et al., 2001), we divide the energy flux by the number flux to derive a “typical” primary electron energy of 340 keV = 0.34 MeV. Upon impact of one such primary electron, the number of secondary electrons (of energy 1–10 eV) generated is about 7 × 104 (Eq. 6). Multiplied by the primary electron number flux and by the 306/400 nm fluorescent photon yield for 10 eV electrons derived above, we obtain a fluorescent photon production rate of order 1 cm−2 s−1. This is an upper limit, because we have assumed that all secondary electrons (energy 1–10 eV) can induce fluorescence, whereas Prince et al. (1976) determined that electrons with energy below ≈10 eV do not induce fluorescence. Europa is the most irradiated icy moon in the Solar System (Cooper et al., 2001; Moore and Hudson, 2003), so we expect ice fluorescence to be lower everywhere else.
In comparison, fluxes of near-UV/visible photons on planetary surfaces across the Solar System are many orders of magnitude higher (Table 3). This does not necessarily mean that life would not utilize fluorosynthesis, but it does make it unlikely just below the surface of any planetary object. This mechanism, however, may still be important at greater depths opaque to visible sunlight but amenable to the presence of high-energy photons or particles or of substantial amounts of beta particles. Lastly, water ice is less fluorescent than N2 ice, which is relatively common in the outer Solar System.
The photosynthetic photon flux for various planetary bodies was estimated using the inverse square law. The average distance from the Sun of the host planet was used for the satellites. The equation I = P/4πr 2 was used to calculate the photosynthetic solar constant for the various planetary bodies listed. The solar output was then translated into a photon flux using the conversion coefficient for global photon output on Earth (1.814 μmol J−1). The flux from the Sun is still much higher on each body than can be produced by fluorosynthesis, thus precluding a near-surface presence.
Processes other than particle radiation could help drive fluorosynthesis. Ionizing photon radiation in the form of X-rays or gamma rays could dissipate their energy by causing molecules to fluoresce as particle radiation does (Mozumder, 1999). Irrespective of the ionization source, flux values of incoming radiation might matter more than total energy supply: icy moons with high irradiation fluxes in planetary magnetospheres are better candidates for searching for fluorosynthetic organisms than environments that have high amounts of particle energy impacting the surface but extremely low particle fluxes. Low particle fluxes here can be defined as those where the space between particles is larger than the organism itself. As with direct electrophy, attenuation of particle energy is important not only to ensure biomolecule resistance to ionizing radiation but also to help spread the energy more evenly throughout the near-subsurface of any icy moon through the creation of secondary electrons.
4. Discussion
4.1. Coping with radiation and cold
The necessity for irradiating particles to reach planetary surfaces in the above mechanisms precludes their viability on any world bearing more than a tenuous atmosphere; although as long as the surface radiation is high enough, direct electrophy could be used. As a consequence, organisms using these putative metabolic pathways would need to cope with additional sources of stress, including high radiation and, in the case of icy worlds, low temperatures. The low pressures and potential drastic pressure changes of near-surface environments could be resisted by terrestrial microorganisms, whose tolerance spans the minute pressures outside the International Space Station (Horneck, 1999; Cox and Battista, 2005; de la Torre et al., 2010; Onofri et al., 2012; Vaishampayan et al., 2012) to 100 MPa at the Challenger Deep (Kato et al., 1998).
Optimal near-surface niches would need to provide sufficient particle and energy fluxes yet sufficiently low levels of ionizing radiation (electrons with energy <10 eV) to prevent excessive damage to biomolecules (Mozumder, 1999; Sanche, 2005; Dartnell, 2011). Despite low radiation levels at Earth's surface, many species have developed strategies to mitigate radiation damage, and even common microorganisms such as E. coli can build radioresistance after multiple exposures to radiation (Pavlov et al., 2006). With special proteins for DNA repair, and up to 10 copies of its genome per cell (Cox and Battista, 2005), Deinococcus radiodurans shows no loss in viability up to a dose of 5 kGy (i.e., an irradiating energy of order 5 × 104 keV absorbed by 1 μm3 of cellular material). Radiation damage can also be mitigated by the electron transfer properties of melanin (Gan et al., 1976; Dadachova et al., 2008), which allow the scavenging of free radicals produced during oxidation-reduction reactions in the cell, thereby reducing DNA damage (Gan et al., 1976; Menter and Willis, 1997). Incidentally, these properties might facilitate secondary electron capture. More indirectly, irradiation of water ice can generate hydrogen peroxide, a source of oxidative stress that microbes on Earth manage with stress-response proteins such as superoxide dismutase, catalase, and peroxidase (Wang et al., 2004; Whitehead et al., 2006; Robinson et al., 2011; Terato et al., 2011).
Low temperatures would be an additional source of stress on icy moons orbiting in the magnetosphere of giant planets. While environments suitable for secondary electron metabolism need not be cold (e.g., the near-surface of a rocky exoplanet in a strong radiation field), those in the Solar System are; this warrants a brief discussion of cold-coping strategies. On Earth, microorganisms can metabolize below 0°C using ice nucleation proteins to induce freezing outside microbial cells (Kawahara, 2002). This induces the osmotic flux of water from the cell to the surrounding ice, increasing solute concentrations in the cell and halting freezing of intracellular fluid (Koop and Zobrist, 2009; Barletta et al., 2012). Such adaptations to freezing can allow microbial activity in solute- and nutrient-rich fluid pockets or veins between ice crystals (Priscu et al., 1998; Mader et al., 2006; Ewert and Deming, 2011). Antifreeze proteins have evolved multiple times (Logsdon and Doolittle, 1997; Graham et al., 2013), suggesting that cold-coping strategies readily result from convergent evolution and adaptation. However, these strategies probably face both chemical and physical limits and may not necessarily be applied at near-subsurface conditions for icy worlds or planetary bodies with tenuous atmospheres. In the Solar System, the warmest environment suitable for secondary electron metabolism is arguably Europa's surface, no warmer than a frigid −140°C (Spencer et al., 1999). This is too cold for any liquid pockets to persist within centimeters of the surface, barring substantial endogenic energy sources (as on Io), greater insolation, or solvents other than water (Bains, 2004; Baross et al., 2007; Stevenson et al., 2015). We note that these stress mitigation strategies involve the expenditure of energy. It remains to be shown whether secondary electrons can supply enough energy for both microbial growth and stress mitigation.
4.2. Detecting electrophic life
To date, evidence for either direct or indirect electrophy has yet to be detected. The absence of electrophilic or fluorosynthetic life in the subsurface icy worlds could be attributed to one of the following: (i) physical-chemical stresses such as low temperatures, and nutrient or solvent limitations; (ii) the absence of material exchange between potential subsurface liquid reservoirs, where presumably such life would emerge, and the solid near-surface where it would adapt; (iii) exchanges occurred too recently for life to have adapted to near-surface conditions; or (iv) life did not evolve on the icy body altogether. Of these possibilities, the postulated age of icy worlds makes possibility (iii) unlikely. Exchange processes, such as ice shell convection, are presumed to occur on icy worlds (e.g. Showman and Han, 2007), precluding (ii). Thus, perhaps only possibilities (i) and (iv) could limit the occurrence of life in the near surface of ocean-bearing icy bodies if direct electrophy and fluorosynthesis are plausible processes.
While terrestrial photosynthesis can be detected remotely, detecting fluorosynthetic organisms may be much more difficult for two reasons. First, the source of the photons is located below the ice surface in the indirect electrophy model, making remote-sensing observations difficult since spectroscopic measurements would only be able to sense the upper portion of the ice crust. However, even if the organisms were located within the top few millimeters of the ice, a positive confirmation of life would still require in situ investigations.
To improve on our order-of-magnitude calculations, the electrical conduction of planetary surfaces could be investigated to see if the initial current created by an incoming plasma can be sustained deeper in the subsurface. Furthermore, our calculations assumed incoming particles of equal energy; refining them by integrating over a range of energies would provide a better idea of the initial electrical current supplied by the incoming plasma.
Studies on terrestrial electrophic microbes might reveal an ability to live off of secondary electrons instead of a cathode. A simple experiment that could be performed in a vacuum chamber to test our hypothesis is shown in Fig. 3. Instead of using a current directly from a battery or other energy source, a small layer of metal may be irradiated by electrons or atomic ions to produce secondary electrons for use by the organisms. More insulating materials can be used in other experiments to ensure the organisms are being powered by the secondary electrons. If an electrophilic terrestrial psychrophile is found, then the same experiment can be repeated using water ice as the irradiated substrate.

Possible experimental setup to test the direct electrophy mechanism. A vacuum chamber is used to simulate surface conditions on airless bodies and allow the passage of an electron beam emitted from an electron gun. The beam hits a surface material suitable to generate secondary electrons. On the other side of this surface lies a small batch of terrestrial electrophilic organisms lying in their optimal environmental conditions, the only incoming energy source being secondary electrons. Sustenance of these organisms would help determine if attenuated particle radiation from exogenic sources such as planetary magnetospheres could be used by extraterrestrial life as an energy source.
5. Conclusion
We have presented and discussed two hypothetical mechanisms through which microorganisms may use free electrons as an energy source, while presenting some challenges to the proposed mechanisms. The first mechanism, “direct electrophy,” involves direct uptake of secondary electrons from incoming particle or photon radiation. As an upper limit, one could expect a biomass of about 10−5 g cm−2 to use such a metabolic strategy. The second, “indirect electrophy,” focuses on the indirect use of secondary electrons through a fluorescing compound (e.g., N2 ice) which generates visible light to drive photosynthesis (fluorosynthesis). Direct electrophy would require fewer evolutionary steps and may be a more likely mechanism than fluorosynthesis-based indirect electrophy. While direct electrophy could sustain a substantial biomass, indirect electrophy suffers from a flux issue: the photosynthetically active photons from the Sun reaching even distant worlds such as Kuiper belt objects could deliver more photons than those obtained by irradiating water ice. However, the high-energy photons or particles that would aid in fluorosynthesis could penetrate relatively deeper under the surface. In addition, more fluorescent compounds such as N2 ice could better drive fluorosynthesis.
Although we proposed laboratory experiments to test our hypothesis, the only indisputable way to do so would be to explore and sample the potential habitats on and near the surfaces of icy worlds and other planetary bodies. The exploration of the surfaces of these bodies might reveal the presence of such organisms much closer to the surface than previously hypothesized.
Footnotes
Acknowledgments
The authors thank the Josep Comas i Sola International Summer School in Astrobiology, the Instituto Nacional de Tecnica Aeroespacial (INTA), and the NASA Astrobiology Institute's Summer School Scholars program for partially funding this work. We especially thank V. Meadows for reviewing the initial manuscript. K. Stelmach thanks the NASA Earth and Space Science Fellowship program for helping fund his graduate work when these ideas were developed. T. Vick-Majors acknowledges the American Association of University Women for dissertation funding during the preparation of this manuscript. T.M. Hoehler and R. Barnes provided critical creative suggestions during the early phases of this project. D. Allu-Peddiniti, A. Angermeyer, C. Harman, B. Farcy, P. Gao, B. Lam, and A. Rodriguez helped with early idea development. Lastly, three anonymous reviewers greatly helped improve the quality of this manuscript.
Disclosure Statement
No competing financial interests exist.