
Abstract
Observed features such as recurring slope lineae suggest that liquid water may exist on the surface and near-subsurface of Mars today. The presence of this liquid water, likely in the form of a brine, has important implications for the present-day water cycle, habitability, and planetary protection policies. It is possible that this water is formed, at least partially, by deliquescence of salts, a process during which hygroscopic salts absorb water vapor from the atmosphere and form a saturated liquid brine. We performed laboratory experiments to examine the ability of Bacillus subtilis (B-168) spores, alone or mixed with calcium perchlorate salt (Ca(ClO4)2), to form liquid water via deliquescence under Mars-relevant conditions. Spore survival after exposure to these conditions was examined. An environmental chamber was used to expose the samples to temperature and relative humidity (RH) values similar to those found on Mars, and Raman microscopy was used to identify the phases of water and salt that were present and to confirm the presence of spores. We found that B-168 spores did not condense any detectable water vapor on their own during the diurnal cycle, even at 100% RH. However, when spores were mixed with perchlorate salt, the entire sample deliquesced at low RH values, immersing the spores in a brine solution during the majority of the simulated martian temperature and humidity cycle. After exposure to the simulated diurnal cycles and, in some cases, perchlorate brine, the impact of each environmental scenario on spore survival was estimated by standard plate assay. We found that, if there are deliquescent salts in contact with spores, there is a mechanism for the spores to acquire liquid water starting with only atmospheric water vapor as the H2O source. Also, neither crystalline nor liquid Ca(ClO4)2 is sporicidal despite the low water activity. Key Words: Raman microscopy—Mars—Planetary protection—Salts—Water activity. Astrobiology 17, 997–1008.
1. Introduction
T
The case for current habitability on Mars is a challenging topic to address due to the cold temperatures and dry environmental conditions. Generally, the surface of Mars is too cold, too dry, or both, thus making it difficult for terrestrial organisms to thrive (Rummel et al., 2014). However, salts discovered on the martian surface and in the subsurface provide a pathway for the formation and stability of salt solutions in current environmental conditions (Chevrier et al., 2009; Gough et al., 2011, 2014; Nuding et al., 2014, 2015; Martín-Torres et al., 2015; Ojha et al., 2015). Evidence for the presence of liquid salt solutions (brines) on current Mars was recently reported (Ojha et al., 2015). Liquid water is essential to life as we know it, and thus water cycle processes on Mars today, at both macro and micro scales, must be better understood to determine the potential for habitability.
The presence of perchlorates at the Phoenix landing site and potentially in the Gale Crater region near the Mars Science Laboratory (MSL) landing site may affect localized liquid water stability (Glavin et al., 2013; Leshin et al., 2013; Ming et al., 2014). Perchlorates have been shown to absorb water vapor and deliquesce from a crystalline solid into a liquid brine under present-day martian temperature and RH conditions (Chevrier et al., 2009; Gough et al., 2011, 2014; Nuding et al., 2014). If liquid salt solutions are present at landing sites, it is important to understand the interaction of potential perchlorate brines with terrestrial microorganisms.
Laboratory investigations can serve as a proxy for in situ studies on Mars. Nicholson et al. (2012) found that the presence of perchlorate in Mars analog soil extracts does not inhibit spore germination into vegetative cells under present-day Mars atmospheric conditions (Nicholson et al., 2012). Because spores are not impacted, the possibility of contamination of Mars via spores hitchhiking on spacecraft cannot be eliminated (at least not by this short exposure study).
Calcium perchlorate specifically has been reported at the Phoenix (Hecht et al., 2009; Kounaves et al., 2014) and the MSL (Glavin et al., 2013) landing sites; so, an investigation of spore interaction with this particular perchlorate salt is necessary. Our previous investigations demonstrated the extreme hygroscopic properties of Ca(ClO4)2 and its ability to persist as a metastable liquid down to 1% RH at Mars-relevant temperatures (Nuding et al., 2014). This study describes the interaction of Bacillus subtilis spores (B-168 strain) with Ca(ClO4)2 and water vapor under simulated Mars-relevant temperature and RH diurnal cycles.
Here, we seek to answer two main questions: (1) Does perchlorate salt provide a mechanism for spores to acquire and interact with liquid water under martian conditions? (2) Does the immersion of spores in a saturated or even supersaturated perchlorate brine adversely affect spore survival? To answer these questions, Raman microscopy and an environmental chamber allowed us to visually and spectrally analyze the composition and phase transitions of B. subtilis spore and Ca(ClO4)2 mixtures when they are exposed to present-day martian diurnal cycle temperature and humidity conditions. We then used standard plate assay analysis to determine whether B. subtilis spore survival was adversely affected by a briny, low-temperature, low water activity environment.
2. Materials and Methods
2.1. Sporulation of B. subtilis
Purification of B. subtilis spores was carried out as reported elsewhere (Osman et al., 2008). Briefly, nutrient broth sporulation medium was used to induce sporulation from the various isolates (Schaeffer, 1965; Nicholson and Setlow, 1990). A single purified colony of the strain to be sporulated was inoculated into liquid nutrient broth sporulation medium. After 2 to 3 days of incubation at 32°C, the cultures were examined in wet mounts on an Olympus BX-60 phase-contrast microscope to determine the level of sporulation.
Spores appeared as phase-bright bodies when viewed with the phase-contrast microscope. Once the number of free spores in the culture was 90% greater than the number of vegetative cells, the culture was harvested, and the spores were purified. Spores were purified by treatment with lysozyme and washing with salt and detergent (Nicholson and Setlow 1990). Cultures were harvested by centrifugation (5000 rpm, 10 min, 4°C). The pellets were washed with 1 M KCl/0.5 M NaCl and incubated for 60 min at 37°C in Tris-HCl (50 mM, pH 7.5) containing lysozyme (50 μg/mL) to degrade immature spores, any remaining vegetative cells, and cell-wall debris. The spores were cleaned by additional centrifugation (10,000 rpm, 10 min, 4°C) and washing sequentially with NaCl (1 M), deionized water, sodium dodecyl sulfate (0.05%), TEP buffer (50 mM Tris-HCl, pH 7.5, 10 mM EDTA, 2 mM phenylmethylsulfonyl fluoride), and, finally, three washes with deionized water. The resulting purified solution contained >99% of spores that were resuspended in sterile deionized water, heat-shocked (80°C for 15 min), and stored at 4°C in glass tubes.
2.2. Preparation of B. subtilis and Ca(ClO4)2 solutions
The B. subtilis spores prepared had an initial concentration of 1 × 107 spores per mL of sterile water. Ca(ClO4)2•4H2O was purchased from Sigma Aldrich (99% purity). To investigate the impact of Ca(ClO4)2, 100 μL of the spore solution (1 × 107 mL−1) was mixed with 3 mL of a 1% Ca(ClO4)2 solution. The control solution that did not contain Ca(ClO4)2 contained 100 μL of the spore solution (1 × 107 mL−1) diluted in 3 mL of sterile water. The test solutions were nebulized individually at room temperature by using high-purity nitrogen, and droplets were deposited onto a quartz disk.
2.3. Experimental setup
To determine the impact of the martian temperature and RH environment on B-168 spores with and without Ca(ClO4)2, the spores were exposed to martian temperature and RH conditions while being probed with a combination of Raman spectroscopy and optical microscopy. A Raman microscope equipped with an environmental chamber was used to visualize B-168 spores and detect liquid water uptake by the spore and/or perchlorate when exposed to Mars-relevant temperature and RH conditions. Specifically, a Nicolet Almega XR Dispersive Raman spectrometer was outfitted with a Linkam THMS600 environmental chamber, a Linkam automated temperature controller, and a Buck Research chilled-mirror hygrometer (Baustian et al., 2010; Gough et al., 2011). Raman spectra and line maps were generated with a double-frequency Nd:YAG laser at 532 nm (1 μm spectral resolution).
Each quartz disk sample was placed directly on the silver block where the temperature was controlled by a combination of cooling by using a continuous flow of liquid nitrogen and resistive heating. Initially, high-purity dry N2 continuously flowed through the chamber to ensure a dry environment with RH <1%. To control RH in the chamber, a separate and adjustable stream of N2 passed through an H2O vapor bubbler before combining with the dry flow. After passing through the chamber, the flow was directed to a frost point hygrometer for measurement of water vapor. Frost point measurements, specifically dewpoint, from the hygrometer and sample temperature measurements from the platinum resistance sensor allow for determination of the RH at the sample. A diaphragm pump pulling at a rate of 1 L/min was attached to the outlet of the hygrometer to ensure a constant airflow through the chamber independent of any variability in flow rate through the H2O vapor bubbler. Each experiment simulated martian diurnal cycle temperature (223–273 K) and RH (0–100%) conditions with a heating/cooling rate of 1.2 degrees (K)/minute. Mars subsurface models report a range of heating/cooling from 0.02 to 0.08 K/minute (Nuding et al., 2014). In all experiments, temperature was varied while holding H2O partial pressure constant. This process allowed for fine-tuned RH control at lower temperatures. The estimated uncertainty in the RH values reported due to the accuracy of the instrumental apparatus is ±1% RH.
Once the spore solutions were nebulized, the quartz disk that contained the sample was placed in the environmental chamber and analyzed visually and spectrally. Figure 1 shows microscopic images (50 × objective) with correlating Raman spectra at 60% RH for pure Ca(ClO4)2 (Fig. 1A), pure spore (Fig. 1B), and spore/perchlorate mixture (Fig. 1C) samples, respectively. In Figure 1A, a large Ca(ClO4)2 particle is shown in the aqueous phase (indicated by the broad O-H stretch) with the signature perchlorate peak at 936 cm−1 (Nuding et al., 2014). Although the Raman spectrum shown in Figure 1A is of the largest droplet, other much smaller (1 μm) aqueous Ca(ClO4)2 droplets are also seen in this image. The pure spore sample (Fig. 1B) shows 1 μm B-168 spores. Several of the spores formed a cluster as seen in the center of the image. The spores are spectrally fluorescent, but a signature peak is detectable at 1085 cm−1 (Hutsebaut et al., 2006). No O-H stretch was detected, indicating that the spores were not adsorbing a measurable amount of H2O at this RH. Although no visual or spectral evidence of liquid water uptake is present, the spatial resolution of the Raman signal (spot size = 1 μm) is on the same scale as the size of the spores (∼1 μm). The mixing of spores with 1% Ca(ClO4)2 (Fig. 1C) resulted in several spores aggregating to form a cluster in the center of a very large (40 μm) perchlorate crystal. It was commonly observed that the spores formed larger clusters when particles were generated from a mixture of perchlorate and spores, compared with particles generated from a solution consisting only of spores.
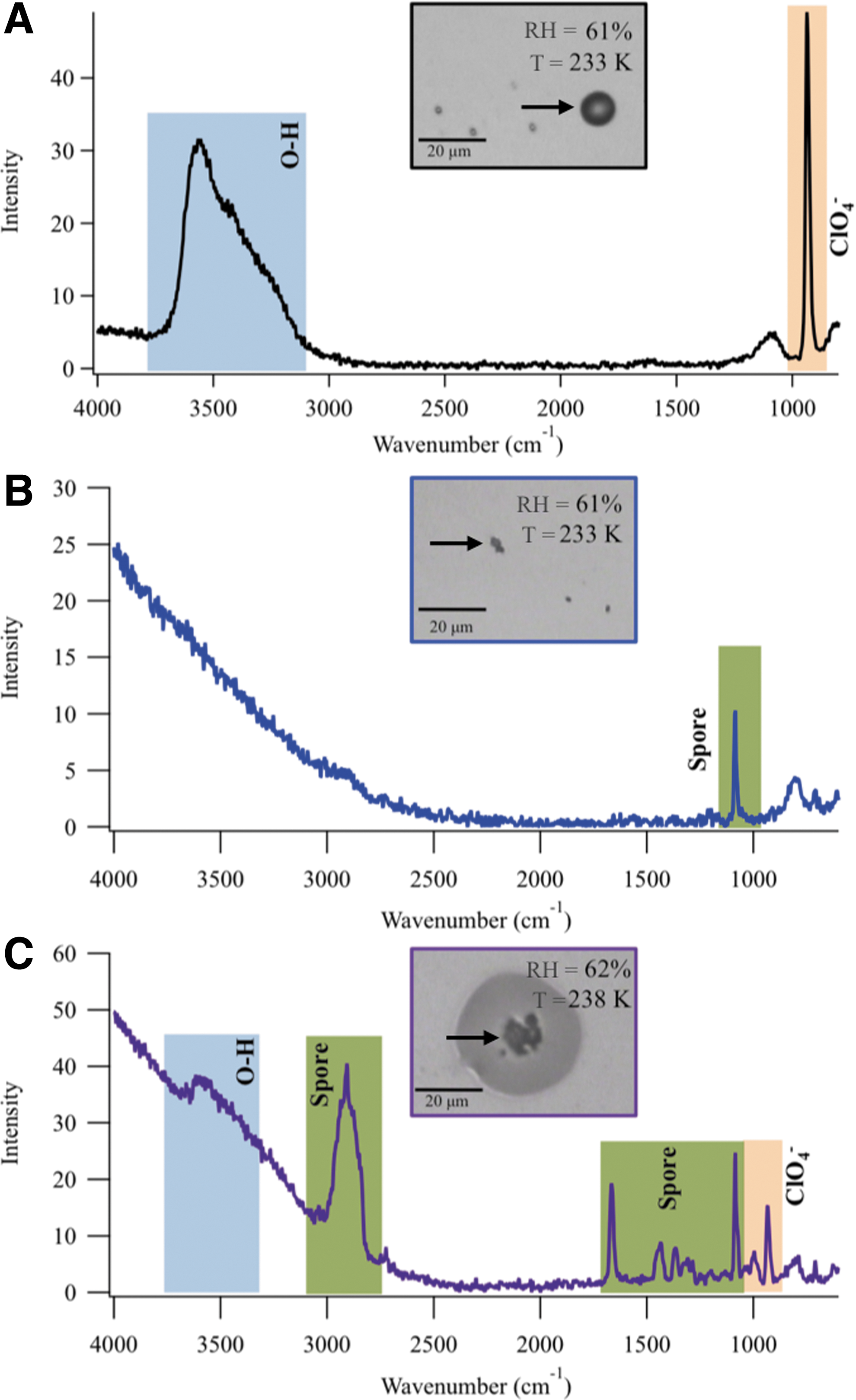
Overall, these combined particles were typically 50 μm, but particles were as large as 150 μm in diameter in some cases. Due to the variability in particle size, four samples were generated and analyzed to reduce uncertainty of results. The larger concentration of spores in the spore/perchlorate mixture allowed for not only clear identification of the 1085 cm−1 spore peak but also detection of peaks at 2908, 1665, 1434, and 1365 cm−1. The difference in intensity of the spore-related peaks between the Raman spectra in B and in C could be due to differences in the water content of the environment surrounding the spores (Kong et al., 2012). Several of these spore-related peaks are similar in location (within 10 cm−1) to known dipicolinic acid peaks (Kong et al., 2012). This species comprises a large amount of the dry weight of bacterial spores. Ca(ClO4)2 in the aqueous phase is identified by the perchlorate peak at 936 cm−1. In addition, liquid water is clearly present as evident by the O-H stretch (3500–3635 cm−1) (Baustian et al., 2010).
It should be noted that, in previous studies of spores, researchers using Raman spectroscopy have detected changes that occur during germination.
Kong et al. (2010) found that information about a spore's germination state can be obtained from the refractility (brightness of a phase-contrast image) and the release of key biological compounds from the spore, as monitored with Raman spectroscopy. We did not collect such data here, nor do we have any reason to believe that the spores would have begun germination during the experiments.
2.4. Bacterial survival
To determine how the viability of spores was affected by the presence of deliquescent perchlorate salts when under martian conditions, we exposed each type of sample (B. subtilis spores only (control) and B. subtilis spores + salt) to either one or six simulated Mars diurnal cycles. A new sample was made for each case. After exposure to the martian diurnal temperature and RH cycles, each quartz disk was placed in a sterile tube. To analyze the impact of each environmental scenario on spore survival, the surviving spore population was estimated by standard plate assay (Osman et al., 2008). The quartz disks were placed into 5 mL of sterile water and sonicated for 2 min to dissociate the attached spores from the substrate. After sonication, sample aliquots were then transferred into petri dishes, and cultivable spores were enumerated by the pour plate technique with trypticase soy agar (TSA) as the growth medium (32°C for 3–7 days) (Osman et al., 2008). This was repeated a total of eight times, four disks for B-168 spore samples and four disks for Ca(ClO4)2/B-168 spore samples.
3. Results
3.1. Exposure of pure B. subtilis spores to martian temperature and RH environment
The control spores (no perchlorate) were cycled through an accelerated Mars diurnal temperature and RH environment to discern whether they could condense any water vapor. Figure 2 shows optical images taken during a B-168 cooling cycle experiment. Each image is a snapshot of the sample as the temperature was cooled from 273 K and the RH was simultaneously raised from 0% RH. The first image (Fig. 2A) shows the small, 1 μm B-168 spores with a few larger clusters of spores. As temperature was cooled and RH was increased in a Mars-relevant manner (Fig. 2B, D), there was no spectral evidence of condensation of liquid water from the vapor phase. In addition, no changes of any sort were visually observed during any cycles performed on the pure B-168 spores. This was confirmed by Raman spectroscopy; as shown in Figure 1, no O-H stretch or other spectral change was observed in the B-168 spore spectra as temperature was decreased and RH was increased, even after multiple cycles. This confirms that the spores alone were not hygroscopic under martian conditions, at least at the detection limits of our spectral technique.
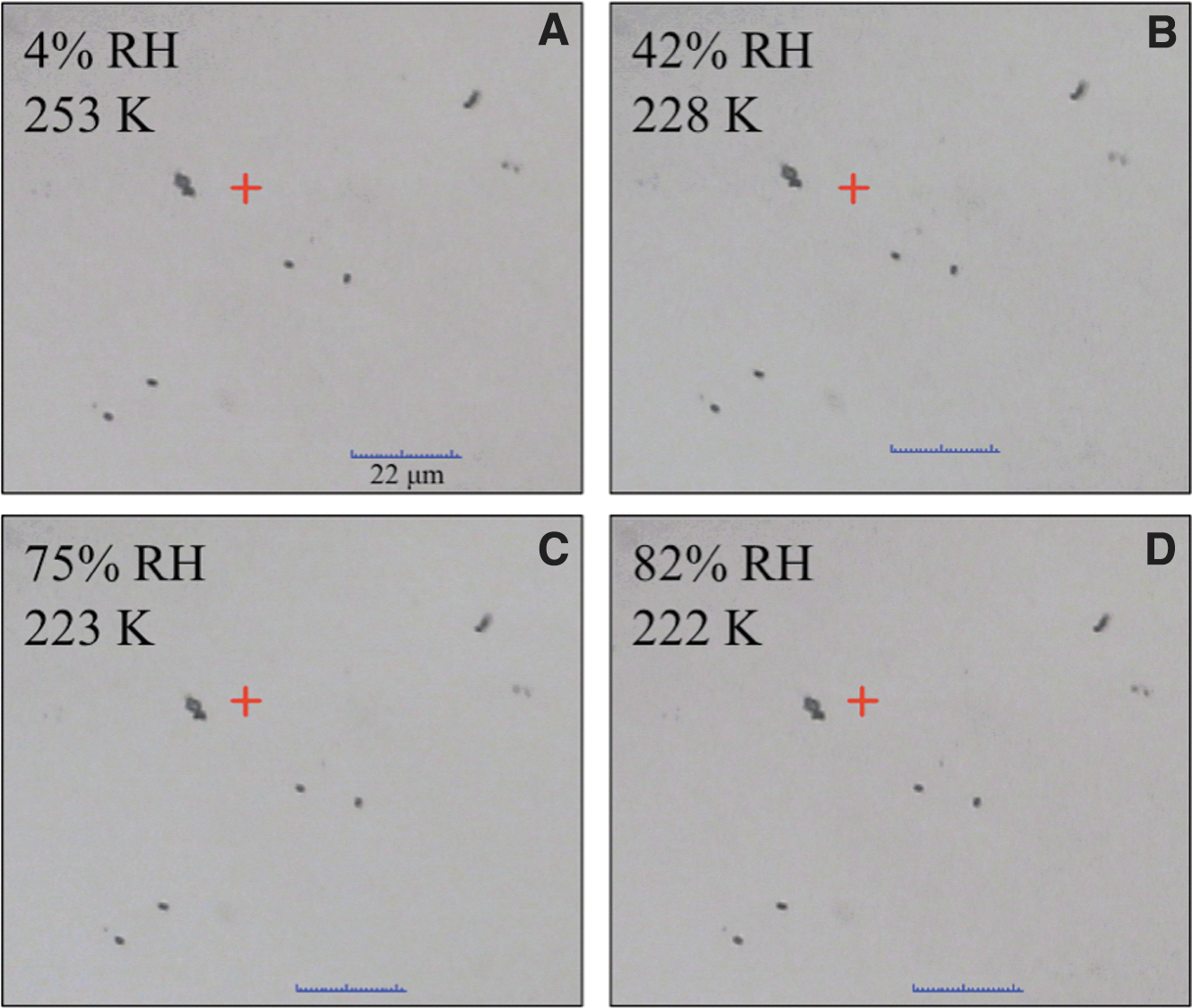
Microscope images taken during a simulated martian diurnal cycle experiment on B-168 spores. The first image shows spores at 4% RH and 253 K. There appear to be single spores (∼1 μm in size) and small clusters of several spores. Temperature is decreased at 1.2 K/min to reach 228 K and 42% RH, where no visual change is observed. As RH continues to increase to 82% RH (at 222 K), there is no spectral or visible evidence of condensation of water onto the spores.
3.2. Exposure of B. subtilis spores and Ca(ClO4)2 mixture to martian temperature and RH environment
We found that when Ca(ClO4)2, a highly deliquescent salt, was mixed with B-168 spores there was formation of liquid water around the spore due to deliquescence of the salt. The salt used here, Ca(ClO4)2, is particularly deliquescent. Experimental results with Ca(ClO4)2 alone reveal a deliquescence relative humidity (DRH) of 10% and an efflorescence (transition from aqueous solution to solid crystalline) relative humidity (ERH) as low as 1% RH at 0°C (Nuding et al., 2014).
Figure 3 shows a series of images of a Ca(ClO4)2/B-168 spore sample taken throughout a simulated diurnal cycle. This experiment started at 1% RH and 0°C (Fig. 3A), with two spore/Ca(ClO4)2 particles in the field of view. For the cooling cycle (images in top row), the temperature was decreased at 1.2 K/min. The larger particle on the left side of the image contains three dark regions that consist of a cluster of B-168 spores surrounded by crystalline Ca(ClO4)2. The smaller 10 μm particle on the right of the image also shows the dark region of spores in the center of the particle surrounded by Ca(ClO4)2 in the crystalline phase. As RH was increased (by decreasing the temperature at constant water vapor partial pressure), the Ca(ClO4)2 deliquesced as expected by 45% RH (Fig. 3B), surrounding the spores in a saturated aqueous salt solution. This aqueous solution persisted up to very high RH (∼114% RH) and low temperatures (223 K), as seen in Figure 3C.

Microscope images taken during a simulated martian diurnal cycle of a mixture of Ca(ClO4)2 and B-168 spores. The experiment starts at 1% RH and 273 K
Next, the RH was lowered and the sample was warmed, as shown in the lower three images, Figure 3D–F (bottom row). The RH decreased going from right to left in these images. The particle was still aqueous at 50% RH (Fig. 3D). By 8% RH and 253 K (Fig. 3E), the aqueous Ca(ClO4)2 solution had effloresced, recrystallizing into crystalline Ca(ClO4)2 around the spores. No further visual changes were observed as the RH was lowered to 0% RH and the temperature was warmed to 273 K (Fig. 3F).
To confirm the phase of the Ca(ClO4)2 surrounding the B-168 spores under different conditions, we utilized Raman spectroscopy. Figure 4 is a Raman line map (1 μm resolution) of a B-168 spore surrounded by calcium perchlorate at 223 K and RH = 114% (same particle and environmental conditions as in Fig. 3C).
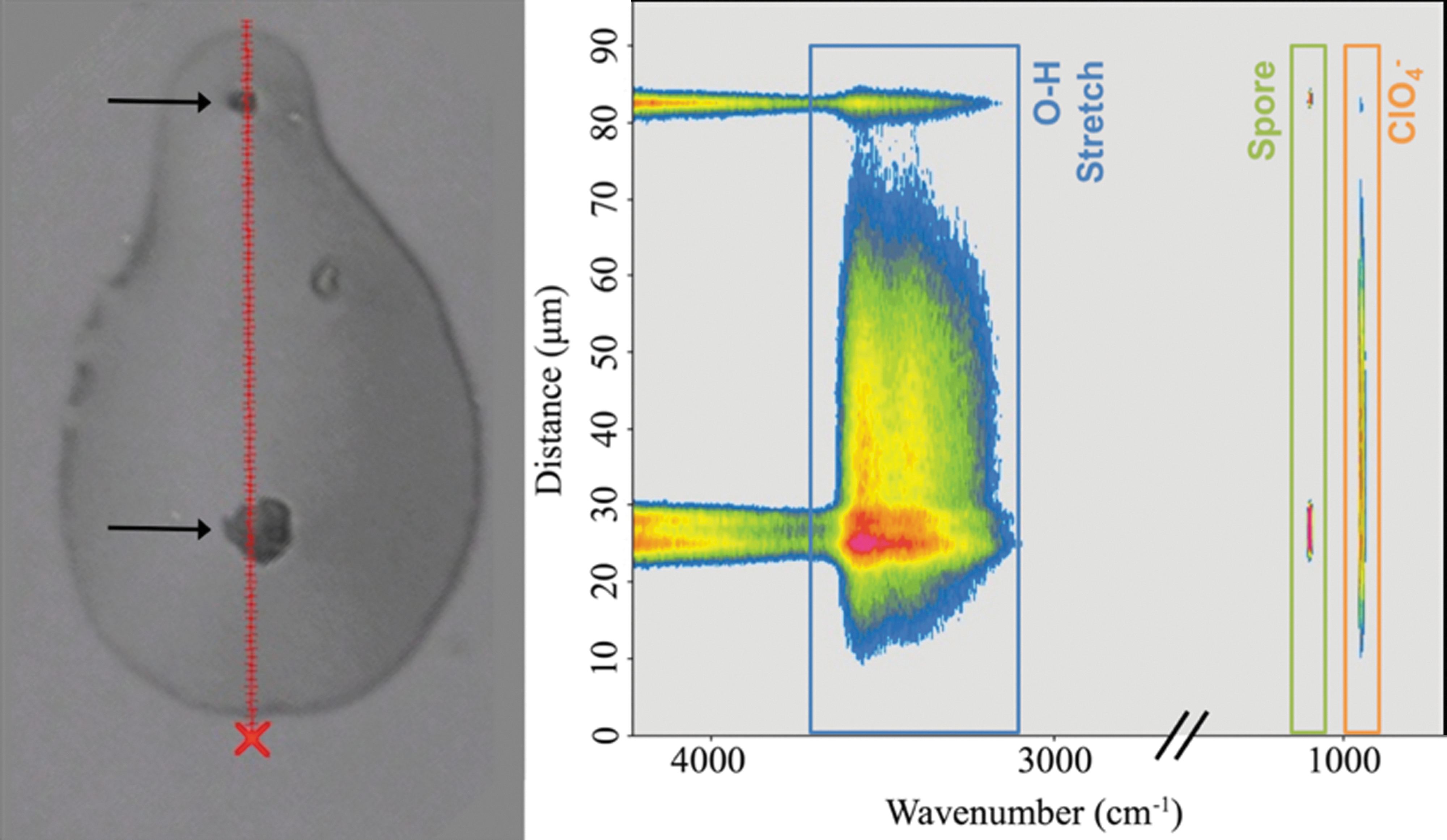
A Raman line map of a B. subtilis and aqueous Ca(ClO4)2 particle at 223 K and RH = 114% (same particle and conditions as in Fig. 3C). This particle is 80 μm long in the y-direction, with the optical image on the left and the Raman line map on the right at the same scale. The optical image shows clumps of dark spores (see arrows) surrounded by aqueous Ca(ClO4)2, with each tick mark on the red line representing 1 μm. The fluorescent features of the spore are apparent between 3700 and 4000 cm−1, correlating with the visible location of the spore in the optical image. A peak at 1085 cm−1 is present in the line map only where the spore is located spatially. The O-H stretch (centered at 3500 cm−1) and solvated ClO4 − stretch (938 cm−1) are present throughout the majority of the length of the entire particle and indicate that an aqueous perchlorate solution surrounds the spores.
Each of the 80 tick marks on the red line in the microscope image represents a location (each 1 μm apart) at which a Raman spectrum was taken. Those corresponding Raman spectra are shown in the right-most panel, a 1D Raman line map. The distance scale on the y-axis of the microscope image and the y-axis of the Raman line map are identical, meaning that each horizontal slice of the Raman map contains the Raman spectrum (with intensity on the color axis) of whichever tick mark lies directly to the right. Warmer colors represent higher Raman intensity. The line map in Figure 4 shows that the Ca(ClO4)2 stretch (938 cm−1) was present throughout the entire particle under these conditions (223 K and RH = 114%). There is an evident liquid water signature shown by the O-H stretch (centered at 3500 cm−1) detected throughout the entire particle with the B-168 spore stretch (1085 cm−1) detected in the two darker regions of the particle in the microscopic image (see arrows). Although this peak location is different than that reported elsewhere (Hutsebaut et al., 2006), the difference in temperature (of at least 30 K) may account for some of the 70 cm−1 offset. The line map confirms that the dark regions in the image were the B-168 spores, which were surrounded by a Ca(ClO4)2 aqueous solution.
In addition to images, Raman spectra were obtained at multiple points during each single-cycle and six-cycle experiment to investigate possible compositional changes of the Ca(ClO4)2 and B-168 spore particles throughout the cooling and warming cycles. Figure 5 is a representative example of an increasing RH/cooling cycle. The experiment started at 0% RH at 273 K (red line). The spore particles were very fluorescent. Because the Raman laser was centered on a spore-rich region of the particle, a large spectral baseline was observed at higher wavenumbers.
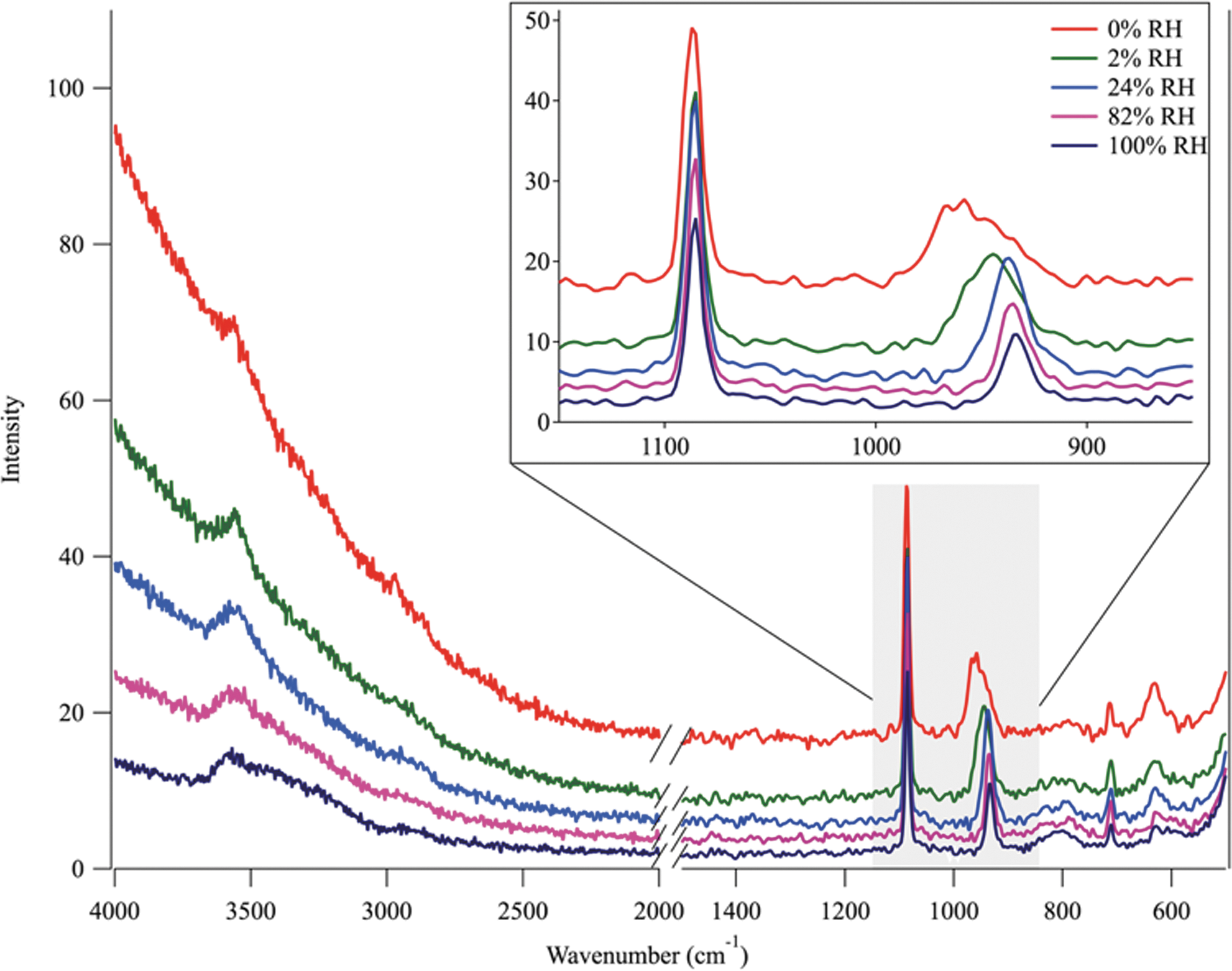
The Raman spectra of a Ca(ClO4)2 and B-168 sample during a cooling cycle that starts at 0% RH at 273 K (red line). The spectra are fluorescent because they are taken in a spore/perchlorate region of the particle. Although the O-H stretch is mostly masked by fluorescence at 0% RH, the experiment begins with hydrated crystalline Ca(ClO4)2, as indicated by the peak centered at 956 cm−1. The B-168 spore peak is centered at 1085 cm−1 and does not shift throughout the entirety of the experiment. As RH begins to increase, an immediate shift of the ClO4 − peak is observed (green line) to 946 cm−1, indicating a change in hydration state. As RH continues to increase, the fluorescence decreases, and by 24% RH (blue line) the ClO4 − peak shifts to the aqueous peak centered at 938 cm−1. The O-H stretch broadens and becomes more prominent as the fluorescence decreases, showing a broad shoulder between 3200 and 3600 cm−1. Once the ClO4 − deliquesces, the aqueous O-H stretch is observed (and remains unchanged) up to RH values higher than 100%.
Although the O-H stretch is masked by the fluorescence at lower RH, the experiment began with hydrated Ca(ClO4)2 as indicated by the peak centered at 956 cm−1 (Nuding et al., 2014). The B-168 spore peak centered at 1085 cm−1 does not shift throughout the entirety of the experiment. As RH was increased, an immediate shift of the ClO4 − peak was observed (green line) to 946 cm−1, indicating a change in hydration state. As RH continued to increase, the fluorescence decreased, and by 24% RH (blue line) the ClO4 − peak shifted to the wavenumber characteristic of an aqueous perchlorate (938 cm−1) signifying deliquescence. The O-H stretch broadened and became more prominent as the fluorescence decreased, showing a broad shoulder between 3200 and 3600 cm−1. Once the ClO4 − deliquesced, no further ClO4 − peak shift occurred in the spectra. The Raman spectra confirmed the visual observations from the optical images; the aqueous Ca(ClO4)2 solution surrounded the spores once the environmental conditions favored the aqueous salt phase.
The warming portion of each diurnal cycle began by increasing the sample temperature at 1.2 K/min, thus decreasing the RH. Figure 6 indicates that at 50% RH and 233 K an aqueous Ca(ClO4)2 solution was present around the B-168 spores. Spectral features of liquid water (O-H stretch), perchlorate (aqueous ClO4 − peak), and spore are all visible here. As RH continued to decrease to 20% RH (teal), the shoulder on the O-H stretch began to shrink, but no shift in the ClO4 − peak was observed, indicating that an aqueous solution was still present. At 5% RH (orange), the ClO4 − peak shifted to 946 cm−1, indicating that a phase change to a crystalline solid had occurred. Finally, as the temperature was increased to 273 K at 1% RH, the ClO4 − peak shifted to 956 cm−1, indicating that the hydrated crystalline solid present at the beginning of the simulated cycle had re-formed. This same deliquescence and efflorescence process (and significant hysteresis) was observed in all experiments. The same DRH and ERH values were also observed in samples exposed to six diurnal cycles as well.
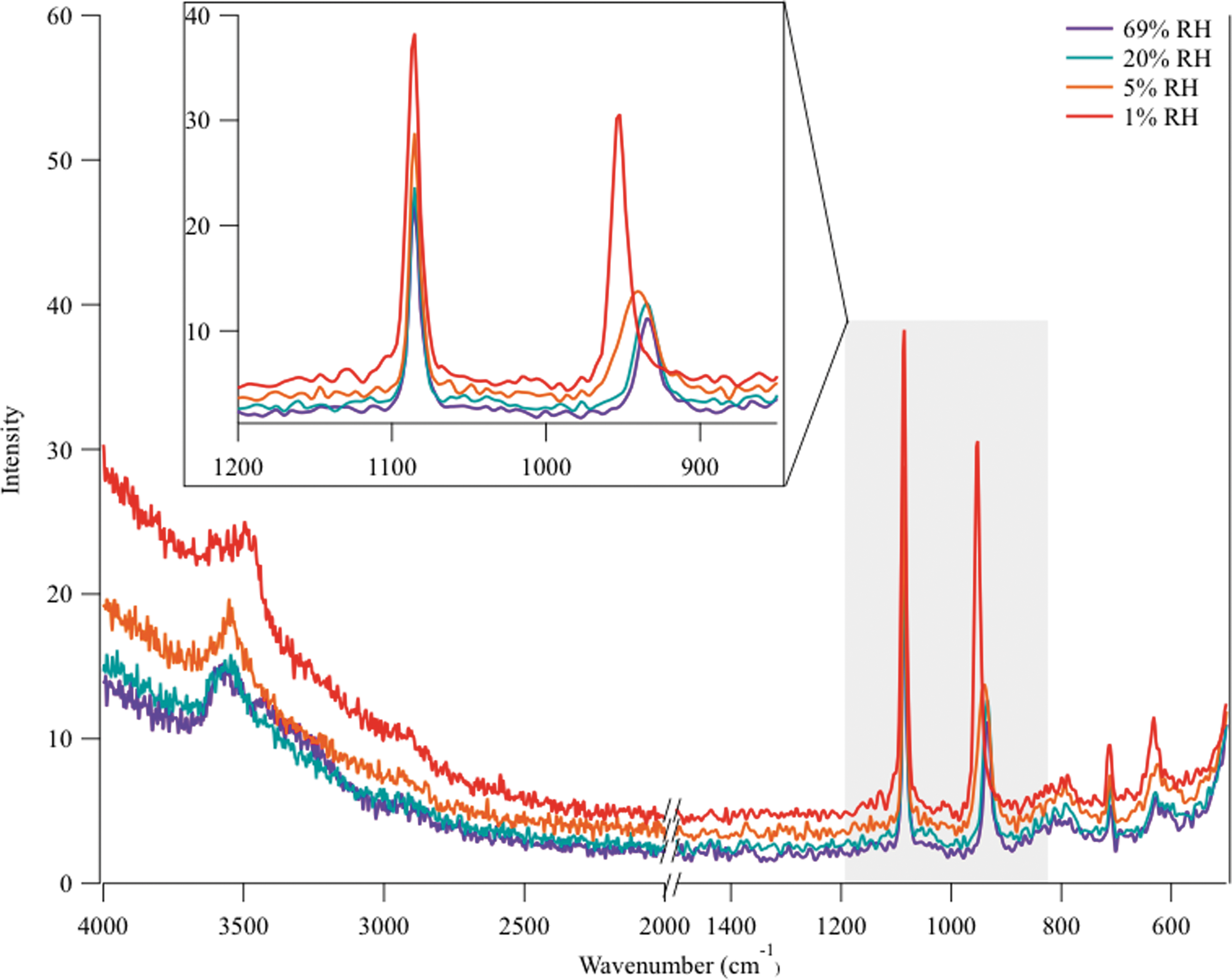
Warming cycle (decreasing RH) of a single Ca(ClO4)2 + B-168 spore particle. Beginning at 100% RH, the sample was heated at a rate of 1.2 K/min, which lowered RH. The purple spectrum (69% RH) contains the O-H stretch, spore peak, and aqueous ClO4 − peak. As RH continues to decrease to 20% RH (teal), the shoulder on the O-H stretch begins to shrink, but no shift in the ClO4 − peak is observed, indicating that an aqueous solution is still present. At 5% RH (orange), the ClO4 − peak shifts to 946 cm−1, indicating that a phase change has occurred. By 273 K and 1% RH, the ClO4 − peak shifts to 956 cm−1, indicating the formation of a hydrated crystalline solid.
3.3. Influence of perchlorate on the survival of B. subtilis spores
To understand the impact of exposure to Mars temperature and RH conditions and contact with Ca(ClO4)2 salts and solutions on survival of the spores, a control sample for each solution was analyzed. The spores in the control sample were not exposed to any martian temperature and RH cycles. The control B-168 samples were nebulized onto the quartz disk, as previously described, from a solution either with or without perchlorate salts. The samples were then immediately placed into a sterile tube and stored at 275 K. These results were used as the baseline for comparison to B-168 samples exposed to single diurnal cycles and six diurnal cycles, and samples were mixed with Ca(ClO4)2.
Figure 7 shows a cell survival comparison between B-168 (control) and B-168 + Ca(ClO4)2 mixtures in the most extreme and Mars-relevant case: exposure to six martian diurnal cycles. Figure 7 represents the average number of colony-forming units (CFU) per μL for each case. Spores were exposed to temperatures as low as −50°C and an RH environment that varied between 0% and 100% RH. The B-168 spore survival, reported at 215 ± 4 CFU/μL (1 SD), did not change (within uncertainty) when the spores were mixed with Ca(ClO4)2. The B-168/Ca(ClO4)2 mixture contained 243 ± 34 CFU/μL (1 SD). To mitigate the effect of various sized clumps of B-168/Ca(ClO4)2 on the quartz wafers seen in Figure 3, the samples were resuspended in liquid and vigorously vortexed before plating on TSA for colony count assay. The B-168 and Ca(ClO4)2 mixture sample results demonstrate that exposure to solid and aqueous Ca(ClO4)2 under martian diurnal conditions had no measurable effect on B-168 survival.
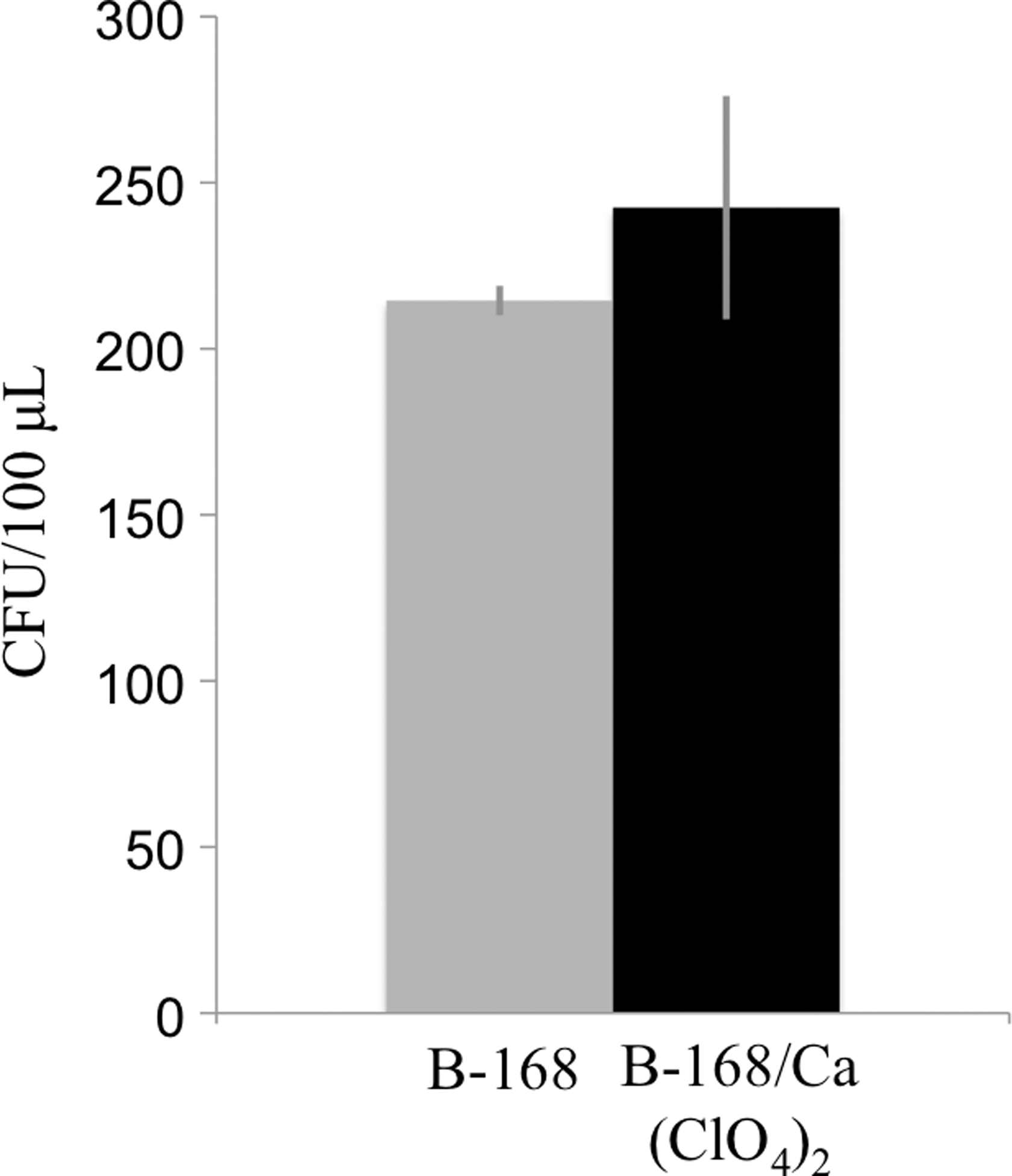
The survival of B. subtilis spores with and without Ca(ClO4)2 after exposure to six martian diurnal cycles. The B-168 spore survival, reported at 243 ± 34 CFU/μL (1 SD) whereas the B-168 spore plus Ca(ClO4)2 mixture contained 215 ± 4 CFU/μL (1 SD). CFU, colony-forming units.
4. Discussion
We showed in section 3.1 that B. subtilis spores alone are unable to absorb measureable water from the atmosphere even under very high humidity conditions. Spores are not known to be hygroscopic; therefore, it was not surprising that condensation of water was not visually or spectrally observed during any 0–100% RH cycles performed on the pure B-168 spores. It was previously known, however, that perchlorate salts, whether pure or in a salt mixture, can readily absorb water vapor from the surrounding environment and deliquesce into a stable, aqueous solution (Chevrier et al., 2009; Gough et al., 2011, 2014).
We show here that, if perchlorate is present in contact with a spore, this hygroscopic salt can provide a pathway for the formation of liquid water in contact with the spore. Specifically, our results indicate that the B-168 spores are surrounded in a deliquescence-derived aqueous Ca(ClO4)2 solution down to extremely low RH values at Mars-relevant temperatures. Even when in contact with spores and even after multiple-phase transition cycles, the Ca(ClO4)2 was observed to deliquesce and effloresce at the relative humidity values previously reported by Nuding et al. (2014), who investigated the deliquescence of pure Ca(ClO4)2. Because the RH at the martian surface typically cycles between very low (<10% RH) and saturation (100% RH) diurnally (Savijarvi, 1999; Zent et al., 2010), it is likely that some deliquescence of Ca(ClO4)2 would occur and that any spores in contact with salts may then be in contact with, or immersed in, brine. The concentrated salt brine that is formed by deliquescence can then potentially interact with the spore or vice versa, although our experiment was not able to prove or disprove an interaction.
We found that the B-168 spores were tolerant to exposure to Ca(ClO4)2 both as a crystalline solid and as a saturated (or even supersaturated) aqueous solution. In all cases examined, after the spores were later germinated, there was no significant change in viable spore count due to the perchlorate exposure. This lack of change in germinated spore count due to salt exposure may not be surprising, as Schuerger et al. (2012) demonstrated that a Mars analog soil containing sodium perchlorate did not result in a significant decline in recovered viable spores of B. subtilis. However, a soil analogue rich in other salts, including chloride salts, did result in a 2.5-fold decrease in B. subtilis survival after 7 days (Schuerger et al., 2012).
To our knowledge, this study is the first to report the effect of spores on perchlorate deliquescence or the effect of a saturated perchlorate brine on the survival of a spore. In each case, the effects were not measurable; the low temperature uptake of water vapor by calcium perchlorate such that a saturated brine was formed did not seem to be affected by the presence of one or more spores. In addition, the survival of the spores was not measurably affected by contact with, or complete immersion in, crystalline or aqueous calcium perchlorate salt, or by exposure to low temperatures. Because this complete immersion of a spore in an aqueous solution was not observed with the spore-only samples, perchlorate deliquescence is, thus, a potential pathway for potential spores to interact in a nonsporicidal way with liquid water in the martian environment. In this study, we did not investigate the germination of spores in a perchlorate brine. More work will need to be done regarding the feasibility of spore germination in saturated perchlorate brine (especially at low temperatures), the effect of salt concentration during spore germination, and other potential biological responses of the spore to this particular perchlorate-rich environment.
5. Conclusion
Observations and model simulations suggest that liquid water exists on the surface or in the near-subsurface of present-day Mars, likely in the form of brines (Renno et al., 2009; McEwen et al., 2011, 2014; Chevrier and Rivera-Valentin, 2012; Martín-Torres et al., 2015; Ojha et al., 2015). Ca(ClO4)2, a hygroscopic salt discovered at recent landing sites, is the best candidate for brine formation. It is known that landed Mars missions such as Phoenix and MSL do contain extreme-tolerant terrestrial microorganisms (Benardini et al., 2014; Venkateswaran et al., 2014), and the presence of liquid water can impact the survival of the spores, their return to a vegetative state, and potentially their transport, if the liquid water is moving.
Here, we experimentally showed that Ca(ClO4)2 deliquescence provides a pathway for spores to interact with liquid water in the martian environment. When exposed to RH values that would be found on Mars, the spores without associated perchlorate were not exposed to any liquid water at any point in the simulated cycle. Samples that contained both spores and perchlorate, however, were observed to deliquesce at low RH values. This allowed the spores to be immersed in liquid water in the form of Ca perchlorate brine during a significant fraction of the simulated martian sol.
For the duration of the studies performed, we found that the survival of B-168 spores was not impacted by the addition of Ca(ClO4)2 when compared with the control (no salt present). The results from this study have implications for potential spores that may land on Mars: Spores in close proximity to perchlorate salts may be able to interact with water formed via salt deliquescence, with only water vapor as the source of H2O. This brine would not be sporicidal despite its high salt concentration and low water activity. In contrast, the deliquescence cycle, and gravitational or capillary effects, could provide a mechanism for transport of microorganisms in the martian environment. Future studies could investigate the survival and possible germination and proliferation of spore-forming bacteria under Mars conditions in the presence of nutrients as well as a deliquescent salt.
Footnotes
Acknowledgments
The research described in this publication was carried out in part at the Jet Propulsion Laboratory, California Institute of Technology, under a contract with the National Aeronautics and Space Administration. The authors acknowledge the technical assistance of Parag Vaishampayan at the Jet Propulsion Laboratory. Part of this research was funded by a NASA Research Announcement Research Opportunities in Space and Earth Sciences (NRA ROSES) grant to K.J.V. Part of this research was also funded by a Mars Fundamental Research Program grant to M.A.T. The authors would also like to acknowledge the NASA Earth and Space Science Fellowship NNX13AO60H.
Author Disclosure Statement
No competing financial interests exist.