
Abstract
In-depth knowledge regarding the biological effects of the radiation field in space is required for assessing the radiation risks in space. To obtain this knowledge, a set of different astrobiological model systems has been studied within the STARLIFE radiation campaign during six irradiation campaigns (2013–2015). The STARLIFE group is an international consortium with the aim to investigate the responses of different astrobiological model systems to the different types of ionizing radiation (X-rays, γ rays, heavy ions) representing major parts of the galactic cosmic radiation spectrum. Low- and high-energy charged particle radiation experiments have been conducted at the Heavy Ion Medical Accelerator in Chiba (HIMAC) facility at the National Institute of Radiological Sciences (NIRS) in Chiba, Japan. X-rays or γ rays were used as reference radiation at the German Aerospace Center (DLR, Cologne, Germany) or Beta-Gamma-Service GmbH (BGS, Wiehl, Germany) to derive the biological efficiency of different radiation qualities. All samples were exposed under identical conditions to the same dose and qualities of ionizing radiation (i) allowing a direct comparison between the tested specimens and (ii) providing information on the impact of the space radiation environment on currently used astrobiological model organisms. Key Words: Space radiation environment—Sparsely ionizing radiation—Densely ionizing radiation—Heavy ions—Gamma radiation—Astrobiological model systems. Astrobiology 17, 101–109.
1. The Radiation Field in Space and Its Hazards to Biological Systems
T
2. Interaction of Radiation with Biological Matter
There are two alternative ways of interaction of radiation with biological key substances, such as proteins, RNA, and DNA: either by direct energy absorption (direct radiation effect) or via interactions with radicals, for example, produced by radiolysis of cellular water molecules (indirect radiation effect) (Kiefer et al., 1996) (Fig. 1). Water is the main constituent of all living systems. Therefore, in irradiated cells, most of the energy is absorbed by water molecules, which are either excited or ionized and later split. The primary products are H•, •OH, H2O+ and electrons (Hutchinson, 1985). All these species possess unpaired electrons, thus being highly reactive free radicals.
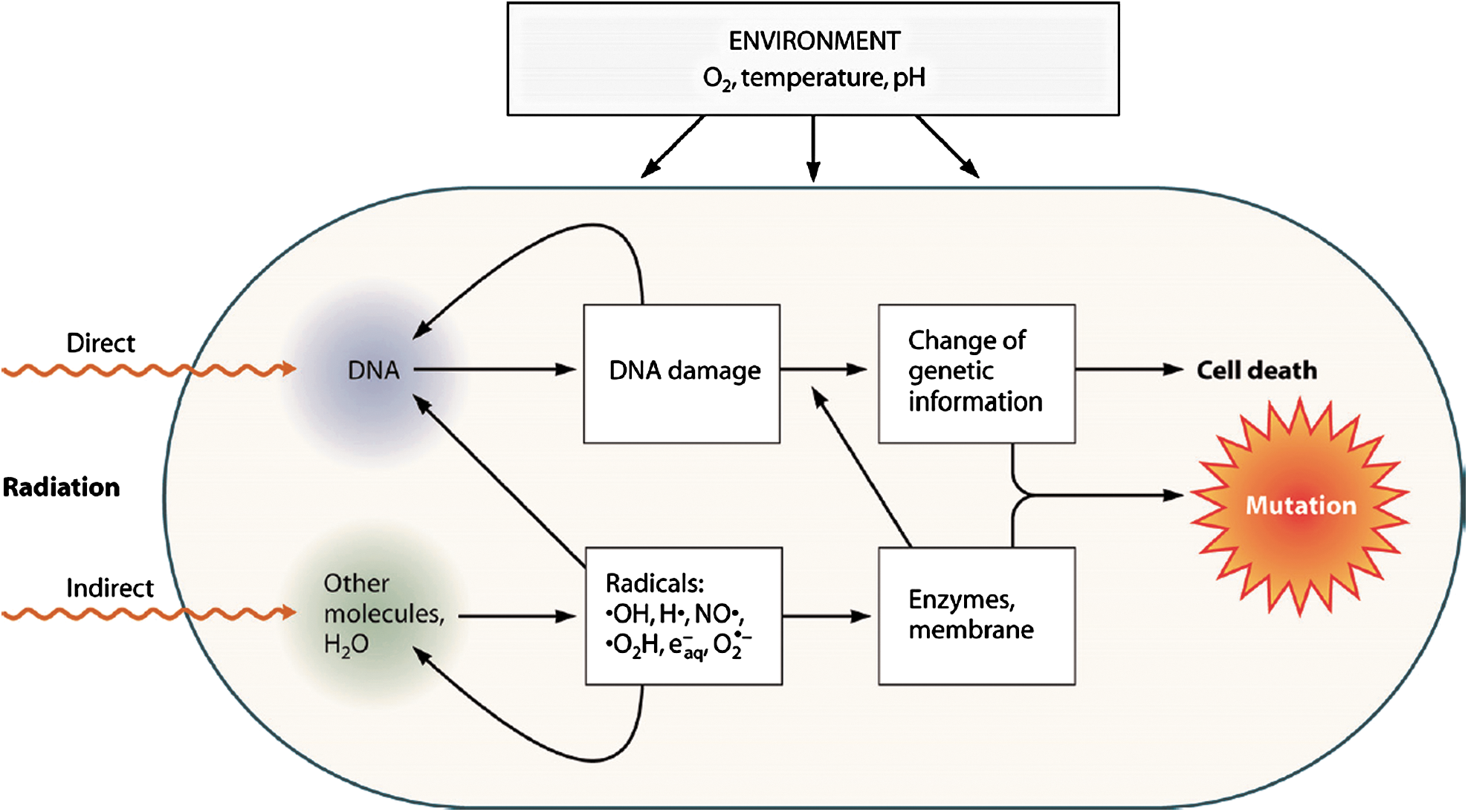
3. Relative Biological Effectiveness of Ionizing Radiation
Ionizing radiation is measured in the SI unit of absorbed dose per mass unit, the gray (Gy), with 1 Gy equal to the net absorption of 1 J in 1 kg of water (please note that with the term dose absorbed dose is meant). The biological effectiveness of radiation largely depends on the local energy distribution, the linear energy transfer (LET). Therefore, different qualities of radiation can have different biological effectiveness, even at the same physical dose, measured in grays. With increasing density of ionization; that is, with increasing LET, the number of changed molecules along the path of a particle increases, leading to a condensation of radiation effects in cells (Kiefer et al., 1996; Horneck et al., 2002, 2006; Durante and Cucinotta, 2011). Densely ionizing radiation, such as the heavy ions and α particles of radiation in space, produces clusters of ions and radicals that are very close together (Fig. 2). If radiation as a beam passes through the target DNA, then the lesions induced by low-LET radiation are located far from each other, whereas those induced by high-LET radiation are densely packed around the beam track (Fig. 2). These latter lesions (locally multiplied damaged types and sites), also called “clustered DNA damage,” are thought to be one of the major causes of the lethal and mutagenic effects of ionizing radiation in space (Goodhead, 1994, 1999; Sutherland et al., 2000). Clustered DNA damage is produced in high yields after exposure to high-LET radiation, for example, Fe ions (Sutherland et al., 2000; Lomax et al., 2002; Asaithamby and Chen, 2011).
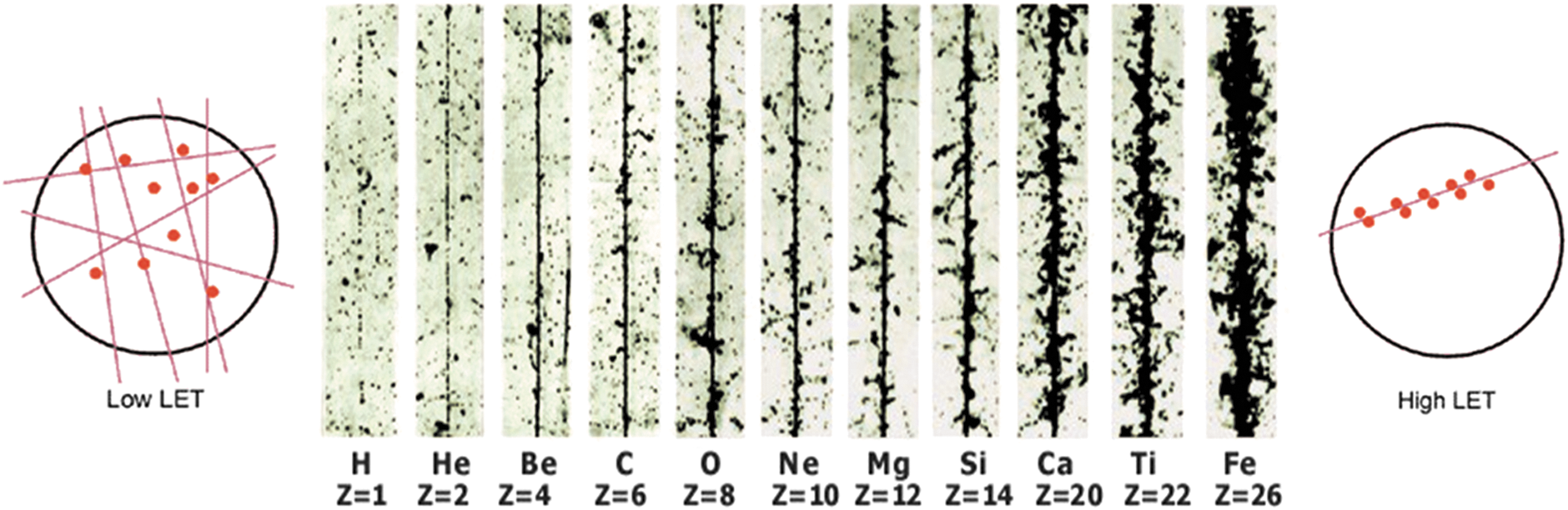
Track structure of different ions of cosmic radiation (from protons to iron ions). Modified image showing (in the center) tracks in photo-emulsions of different nuclei of the primary cosmic radiation. For biological radiation effects, the efficiency of a radiation type increases as the ion density along the tracks increases [Modified from Fig. 7-06 from Horneck et al. (2006); with kind permission of Springer (Springer eBook)]. The left and right parts visualize the ionization potential and tracks of low- and high-LET radiation through a cell [source: Radiation Effects Research Foundation (Hiroshima and Nagasaki, Japan)
This dependence of the biological effectiveness on LET is described by the relative biological effectiveness (RBE) (Butts and Katz, 1967; Edwards, 2011). RBE is the ratio of the physical doses (in Gy) of the test radiation and, for example, X-rays, leading to the same biological effect. The RBE value can be different for different biological systems, depending on their stage in the growth cycle and other environmental factors, such as the oxygen content (Fig. 1) (Kiefer et al., 1996; Horneck et al., 2002, 2006). By averaging over a variety of RBE values for the same LET value, one obtains the quality factor Q, which serves as the biological weighting function of ionizing radiation. For the low-LET X-rays and γ rays, Q is equal to 1. For high-LET radiation, the absorbed dose D needs to be multiplied with Q, resulting in the dose equivalent, H:
with H = the dose equivalent, Q = the quality factor, and D = the absorbed dose. The SI unit for the dose equivalent is sieverts (Sv).
4. Biological Effectiveness of Radiation in Space
4.1. Biological weighting of the particulate components of radiation in space
Galactic cosmic rays (GCR) originate from outside our solar system, being produced in previous cataclysmic events such as supernovae explosions (Simpson, 1983; Wilson et al., 1991). They consist of 98% baryons and 2% electrons. The baryonic component is composed of 85% protons (hydrogen nuclei), with the remainder being α particles (helium nuclei) (14%) and heavier nuclei (about 1%) (Ferrari and Szuszkiewicz, 2009). When GCR enter our solar system, their energies must be high enough to overcome deflection by the magnetic fields of the solar wind (Simpson, 1983; Nymmik, 2006).
The heavier nuclei component of the GCR comprises particles of high (H) charge (Z) and high energy (E) (HZE), which are defined as cosmic ray primaries of charges Z > 2 and of energies high enough to penetrate at least 1 mm of spacecraft or of spacesuit shielding (NCRP, 1989; Cucinotta et al., 2003; Horneck et al., 2006; Durante and Cucinotta, 2011). Although those HZE particles (also called heavy ions) only contribute to roughly 1% of the flux of GCR, they are considered as a potential major concern to living beings in space, especially for long-term missions beyond the protection of Earth's magnetosphere (Durante and Cucinotta, 2011). The reason is their high RBE and Q value, resulting in a high dose equivalent value H. Figure 3 shows the percent contributions from individual GCR elements for their particle flux, dose, and dose equivalent contributions at solar minimum. Whereas the flux of the HZE particles is lower by orders of magnitude than that of, for example, protons, this difference shrinks when the absorbed dose D is considered, which is proportional to Z2. If we consider the dose equivalent H, then the contribution of iron ions exceeds that of protons: Whereas iron ions are only 0.02% abundant considering the flux, they contribute 20% to the dose equivalent H, that is, the biologically effective dose (Cucinotta et al. 2003); for comparison, the contribution of protons to the dose equivalent is only about 9%.
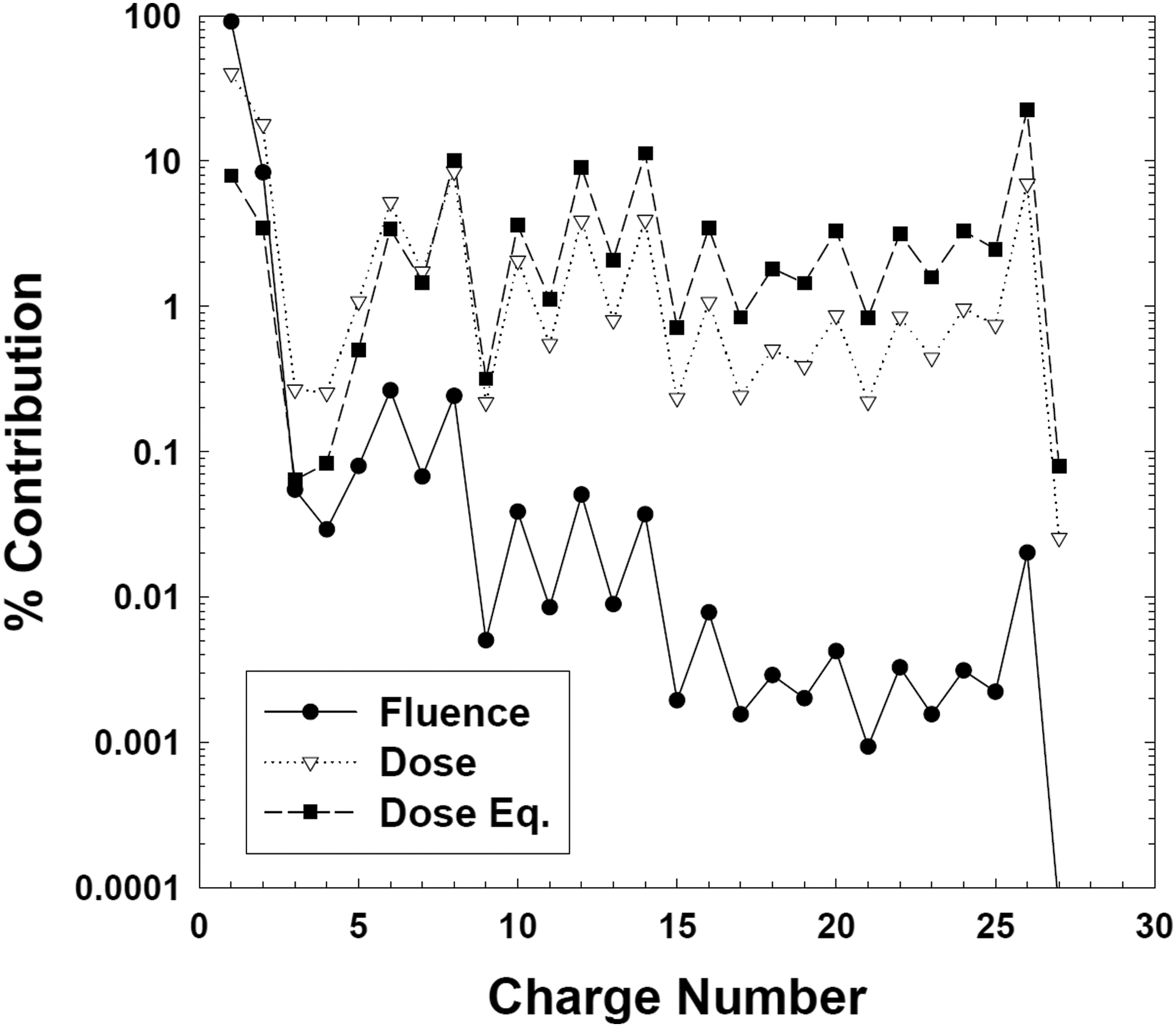
Percent contributions from individual GCR elements for the particle flux and fluence (filled circles), radiation dose (open triangles), and dose equivalent (filled squares) at solar minimum [from Horneck et al. (2006; Fig. 7-04); with kind permission of Springer (Springer eBook)].
4.2. Experiments in space
Radiobiological studies in space dealing with the biological effects of HZE particles of GCR are confronted with the challenge of the very low flux of this particulate radiation: The maximum total particle flux of GCR at solar minimum has been estimated to be about 4 particles/cm2 s (NCRP, 1989), and HZE particles contribute to about 1% of this. To overcome this difficulty, methods have been developed to localize each penetrating HZE particle with regard to the biological system under investigation and to determine its relationship to potential biological effects along its path. One example is the Biostack experiments, which consisted of monolayers of selected biological objects, fixed in position and sandwiched between visual nuclear track detectors (Fig. 4). This arrangement allowed localizing each HZE particle's trajectory in relation to the biological specimens and correlating the biological effect observed with the HZE particle parameters (Bücker, 1975, reviewed in Horneck, 1992). The evaluation of the effects observed in bacterial spores, plant seeds, and animal embryos demonstrated that single HZE particles induce significant biological perturbations, although with varying efficiency. These effects included gross somatic mutations, severe morphological anomalies, disturbance of development, or complete inactivation. However, as a consequence of the intense labor required for those Biostack experiments, they were not continued after the space shuttle era.
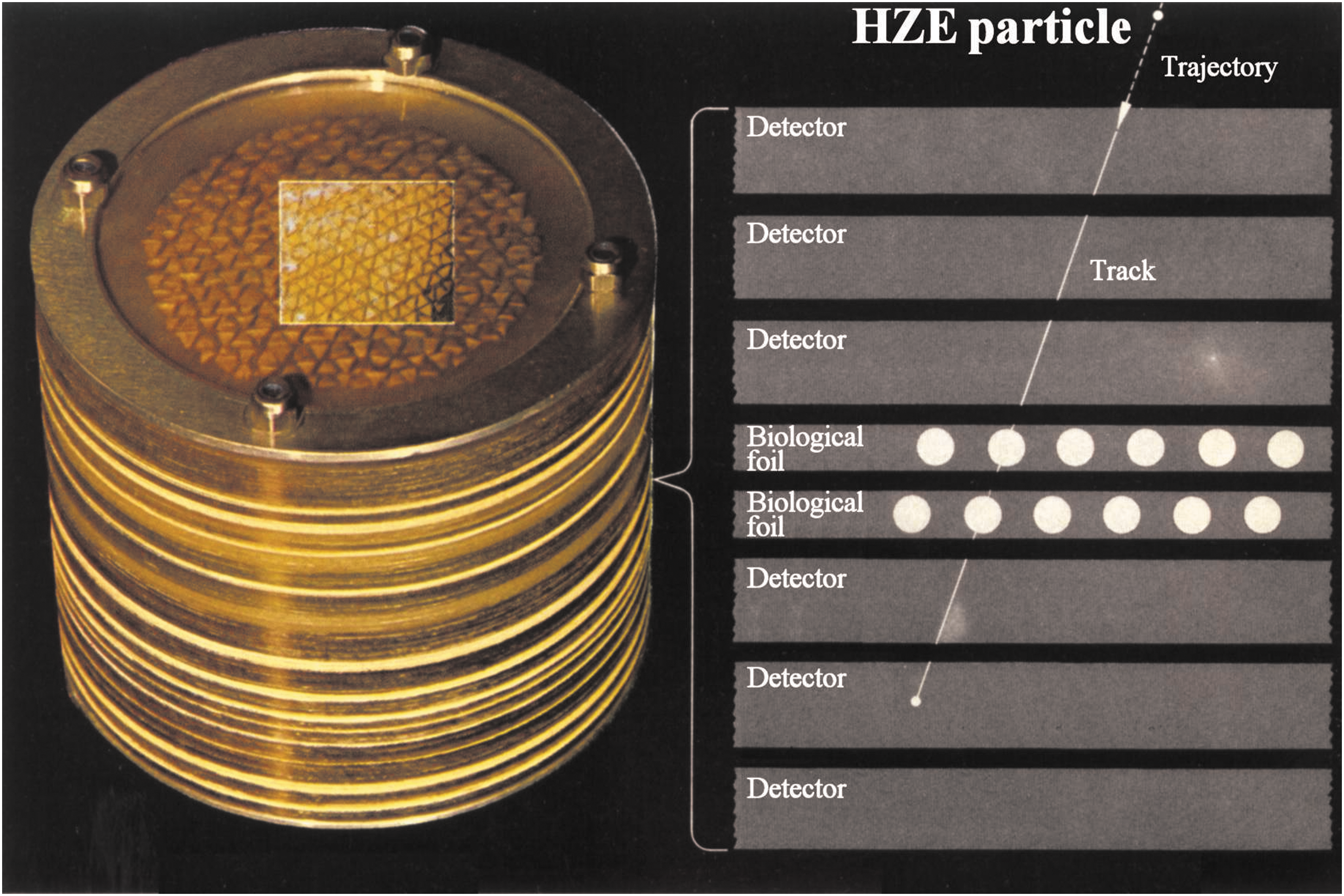
Biostack concept to localize the track of HZE particles of GCR in relation to biological test systems [from Horneck et al. (2006; Fig. 7-10); with kind permission of Springer (Springer eBook)].
4.3. Laboratory experiments mimicking components of radiation in space
In 2012, we established a consortium (called STARLIFE) of various international laboratories and research groups (coordinated by Ralf Moeller) in order to determine and investigate a selection of previously and presently used as well as planned astrobiological model systems for ground-based ionizing radiation experiments (Table 1). Several of those systems have been “space veterans,” which had already participated in space exposure experiments (Rabbow et al., 2012, 2015). In those studies, most organisms have demonstrated high resistance to the non-ionizing parameters of space, such as space vacuum, extraterrestrial solar electromagnetic radiation (especially UV radiation), and extreme temperature fluctuations. However, in those missions, the doses of GCR were in the milligray range, too low to detect any radiobiological effect, if only global analyses were performed. Therefore, it was one of the objectives of STARLIFE to study the responses of those test systems to increased doses of heavy ions, mimicking representatives of the HZE component of GCR; however, it should be kept in mind that the doses and dose rates used in the experiments were much higher than those of natural GCR. From 2013 to 2015, a total of six irradiation campaigns were conducted. They included four irradiations with heavy ions at the Heavy Ion Medical Accelerator in Chiba (HIMAC) facility of the National Institute of Radiological Sciences (NIRS) in Chiba, Japan, using helium (150 MeV/n), argon (500 MeV/n), and iron (500 MeV/n), covering an LET range in water from 2 to 200 keV/μm and doses up to 3 kGy. For comparison, low-LET radiation was provided at the German Aerospace Center (DLR, Cologne, Germany) with X- rays up to 15 kGy and at Beta-Gamma-Service GmbH (BGS, Wiehl, Germany) with γ rays up to 113 kGy (Table 2). Various biological endpoints (e.g., vitality, survivability, cell proliferation, damage induction) have been investigated with a combination of different biochemical and molecular biological methods [e.g., colony formation assays, most probable number, vitality staining, different microscopic analysis, Raman spectroscopy, PCR/RT-PCR, metabolism, and key biomarker stability (Table 1)]. They allowed gaining a better understanding and broader spectrum of the effects of GCR on astrobiological model systems. The individual reports of the STARLIFE group are enclosed in this special issue.
Performed at the HIMAC (but outside the STARLIFE irradiation campaigns).
All the dosimetry and experimental settings/measurements were performed by DLR or BGS scientists/engineers, respectively.
Footnotes
Acknowledgments
The authors are very grateful to all NIRS (HIMAC), BGS, and DLR technicians, operators, coworkers, and students for assistance during the irradiation campaigns. We express our sincere gratitude to Hisashi Kitamura for his excellent support and help throughout the years. R.M., M.R., S.L., T.B., and C.E.H. were supported by the DLR grant FuE-Projekt “ISS LIFE” (Programm RF-FuW, Teilprogramm 475).
Author Disclosure Statement
No competing financial interests exist.
(Other) Members of the STARLIFE Research Group and Their Affiliations
Yuko Kawaguchi, Shin-ichi Yokobori, and Akihiko Yamagishi: School of Life Sciences, Tokyo University of Pharmacy and Life Sciences, Horinouchi, Tokyo, Japan.
Petra Rettberg: Astrobiology Research Group, Radiation Biology Department, Institute of Aerospace Medicine, German Aerospace Center (DLR), Cologne, Germany.
Cyprien Verseux, Mickael Baqué, Riccardo Cifariello, Claudia Fagliarone, and Daniela Billi: Department of Biology, University of Rome Tor Vergata, Rome, Italy.
Claudia Pacelli, Laura Selbmann, Laura Zucconi, and Silvano Onofri: Department of Ecological and Biological Sciences, University of Tuscia, Viterbo, Italy.
Igor Shuryak: Center for Radiological Research, Columbia University, New York, USA.
Annette Brandt, Theresa Backhaus, Joachim Meeßen, and Sieglinde Ott: Institute of Botany, Heinrich-Heine-University (HHU), Düsseldorf, Germany.
Reiner U. Jänicke: Laboratory of Molecular Radiooncology, University of Düsseldorf, Düsseldorf, Germany.
Rosa de la Torre, Ana Zélia Miller, and M. Luisa Martín-Cerezo: Departamento de Observación de la Tierra, Instituto Nacional de Técnica Aeroespacial (INTA), Madrid, Spain.
Beatriz Cubero: Instituto de Recursos Naturales y Agrobiología de Sevilla—CSIC, Sevilla, Spain.
Ute Böttger: Institute of Optical Sensor Systems, German Aerospace Center (DLR), Berlin, Germany.
Jean-Pierre de Vera: Institute of Planetary Research, German Aerospace Center (DLR), Berlin, Germany.
K. Ingemar Jönsson: School of Education and Environment, Kristianstad University, Kristianstad, Sweden.
Andrzej Wojcik: Department of Molecular Biosciences, The Wenner-Gren Institute, Stockholm University, Stockholm, Sweden.