
Abstract
Liquid water is a requirement for biochemistry, yet under some circumstances it is deleterious to life. Here, we show that liquid water reduces the upper temperature survival limit for two extremophilic photosynthetic microorganisms (Gloeocapsa and Chroococcidiopsis spp.) by greater than 40°C under hydrated conditions compared to desiccated conditions. Under hydrated conditions, thermal stress causes protein inactivation as shown by the fluorescein diacetate assay. The presence of water was also found to enhance the deleterious effects of freeze-thaw in Chroococcidiopsis sp. In the presence of water, short-wavelength UV radiation more effectively kills Gloeocapsa sp. colonies, which we hypothesize is caused by factors including the greater penetration of UV radiation into hydrated colonies compared to desiccated colonies. The data predict that deserts where maximum thermal stress or irradiation occurs in conjunction with the presence of liquid water may be less habitable to some organisms than more extreme arid deserts where organisms can dehydrate prior to being exposed to these extremes, thus minimizing thermal and radiation damage. Life in extreme deserts is poised between the deleterious effects of the presence and the lack of liquid water. Key Words: Deserts—Extremophiles—Stress—High temperatures—UV radiation—Desiccation. Astrobiology 17, 309–318.
1. Introduction
E
High temperature is known to cause protein aggregation and denaturation in cyanobacteria (Inouse et al., 2001). Although they have a range of responses when metabolically active to cope with heat stress, such as the production of heat shock proteins (Suzuki et al., 2006), the upper temperature limit for most cyanobacteria in a hydrated state is around 50°C (Casteilli et al., 2009).
Ultraviolet radiation also has deleterious effects on cyanobacteria, particularly in extreme deserts where during the day organisms accumulate UV radiation damage. Using the desert-dwelling cyanobacterium, Chroococcidiopsis, Cockell et al. (2008) showed that these cells were killed within a day by unattenuated surface fluxes of UV radiation. These data demonstrate the importance of habitats that attenuate UV radiation, such as the interior and subsurface of rocks. UV radiation, particularly short-wavelength UVB (280–320 nm) radiation, directly damages proteins and nucleic acids (Quesada and Vincent, 1997) and causes indirect damage through the production of reactive oxygen species (Schulz and Scherer, 1999). The photosynthetic apparatus of cyanobacteria is bleached and destroyed by UV radiation (Castenholz and Garcia-Pichel, 2000).
It is generally assumed that, in deserts where organisms are exposed to physical extremes such as extremes of temperatures and UV radiation, the presence of liquid water is beneficial to life (e.g., Lange et al., 1990; Pointing et al., 2006). Insofar as metabolism (such as repair processes) and the growth and replication of organisms depend on the presence of liquid water, water must be available, even if only transiently, for life-forms to subsist.
However, it is known that, at the biochemical level, water is not always beneficial to life. Hydrolysis reactions are testament to the potentially deleterious role that this solvent can play at the molecular level (Ball, 2007). In this paper, we show how liquid water can also be deleterious to life at the organismal and ecological level, and we show that water defines a window of habitability in extreme deserts that is caused by a trade-off between the presence of liquid water for beneficial biochemical reactions and the role of liquid water in exacerbating cellular temperature and UV radiation–induced damage. We discuss predictions that arise from these results.
2. Methods
2.1. Organisms
Two extremophilic cyanobacteria were used for the investigations.
Chroococcidiopsis strain 029 (isolated from cryptoendolithic growth in Nubian sandstone in the Negev Desert): This Gram-negative non-spore-forming cyanobacterium, which is adapted to life in hot deserts, has been studied previously in the laboratory in a variety of contexts and belongs to a genus reported in diverse hot and cold deserts around the world (Friedmann, 1980; Grilli Caiola et al., 1996; Billi, 2009; Billi et al., 2010; Cockell et al., 2011; Baqué et al., 2013). The cells do not form large colonies, but cells sometimes exist as multiple cell aggregates containing 3–6 cells, including tetrads.
Gloeocapsa sp.: This Gram-negative non-spore-forming cyanobacterium was isolated in our laboratory by exposing rocks obtained from cliff faces in Devon, UK, to conditions in low Earth orbit on the outside of the Biopan orbital satellite and the International Space Station (Olsson-Francis et al., 2010; Cockell et al., 2011). The Gloeocapsa species studied here has a characteristic colonial growth habit whereby cells multiply into an amorphous mass held together by polysaccharide containing several hundred to thousands of cells. Colonies achieve sizes of several millimeters after 3–4 months' growth in liquid media or on the surface of agar plates. In some experiments, these native colonial forms were studied. In other experiments, these colonies were gently broken up into individual cells and cell clumps by homogenization in a 7 mL borosilicate homogenizer (Fisher Scientific, UK), taking care not to rupture the cells. These cells are referred to as “separated cells.” Separation was confirmed under bright field microscopy as described for microscopy below.
Both organisms were cultivated in BG (Blue-Green)-11 liquid medium (Rippka et al., 1979) to prepare cells for the experiments. For the enumeration of cell viability, organisms were grown on BG-11 agar plates containing 2% w/v Bacteriological agar No 1 (Oxoid, UK). All cultures and plates were kept under a light intensity of 50 μmol/m2/s at a temperature of 21°C for 2 months, unless otherwise stated. The organisms were grown under continuous light. This is observed to allow for good growth (unpublished data), and it is observed to be satisfactory for the growth of other cyanobacteria (e.g., Moore et al., 2007).
For experiments in which desiccated cells were used, cultures of cells were aliquoted into Eppendorf tubes and placed into a desiccation jar containing silica gel. Cells were desiccated for 4 days until water within the tubes had been removed and cells formed a dried pellet on the bottom of the tube. We confirmed the desiccated state by observing that there was no residual water associated with the cells under bright field microscopy (see below) and that the cells had lost turgor associated with the hydrated state.
2.2. Microscopy
In some experiments, cells were observed by microscopy. All cells were observed with a Leica DM4000B fluorescence microscope (Leica Microsystems, Germany). Images were acquired with a Leica DFC450C camera. Autofluorescence of cyanobacterial pigments was observed under a rhodamine filter set (510–560 nm, emission 590 nm).
In some experiments, metabolic activity was assessed by examining the activity of esterases. Esterases are a group of intracellular enzymes present in all types of cells that are involved in cell membrane formation. Esterase activity was assessed by the ability of the cells to intracellularly cleave fluorescein diacetate (FDA) to the fluorescent product, fluorescein (e.g., Battin, 1997; Regel et al., 2002). FDA (Sigma Chemicals, UK) was dissolved in acetone at a concentration of 5 mg/mL and then added to cell suspensions to a final working concentration of 5 μg/mL. Cells were incubated at room temperature in the dark for 15 min before examination under the microscope. For FDA visualization, cells were excited at 485 nm with a bandpass of 22 nm and observed at the emission wavelength of 530 nm (bandpass 30 nm). Esterase activity is indicated by cells glowing a bright green color. Inactive cells displayed a red coloration. In cases where cell metabolic viability was quantified, cells were scored as “fluorescing” when they showed a bright green coloration. Esterase activity was expressed as the percentage of “enzyme inactive” cells (red cells) in the observed population.
LIVE/DEAD stain was used according to the manufacturer's instructions (Invitrogen, UK). The kit was used with a 1:1 ratio of SYTO 9 and propidium iodide. Cells were observed under fluorescence microscopy as for FDA analysis (above).
2.3. Experiments
2.3.1. Thermal stress
To investigate the effects of thermal stress on cell viability, Chroococcidiopsis sp. and Gloeocapsa sp. (native colonies) were aliquoted into Eppendorf tubes. Cells were either maintained in BG-11 medium or desiccated. Tubes of cells were placed into a Grant GBD2 heat block (Grant Instruments, UK) at defined temperatures. Following thermal stress, viable cells were enumerated on BG-11 plates, and the loss of viability was determined by expressing the cell viability as a ratio of the number of viable thermally stressed cells to the control (nonthermally stressed, either hydrated or desiccated) cells calculated as a percentage.
In Experiment 1, cells were exposed to temperatures from 40°C to 90°C for 15 min to evaluate the effects of temperature on hydrated and desiccated Chroococcidiopsis sp. (at 5°C increments) and Gloeocapsa sp. (at 40°C, 60°C, and 80°C) cells. In this set of experiments, the colonial growth form of Gloeocapsa sp. was used. We examined fewer temperature points, as our objective was to determine whether, like Chroococcidiopsis, hydration resulted in a lower temperature threshold for thermal stress.
In Experiment 2, the effects of thermal stress in Gloeocapsa sp. were further investigated under hydrated conditions to determine with greater resolution the temperature at which thermal inactivation of cells occurs. In this experiment, the viability of separated, hydrated cultures was tested after heat shock treatment at varying temperatures of 2°C increments between 40°C and 50°C, for varying time increments of 2, 5, 10, 15, and 30 min.
To determine how thermal stress can affect enzymatic activity, the esterase assay was employed. Quantification of the number of metabolically inactive cells was carried out after thermal stress at 40°C, 60°C, and 80°C for both Chroococcidiopsis and Gloeocapsa sp. after different time periods of thermal stress (5, 10, 15, and 30 min).
To determine whether loss of cell membrane integrity might account for loss of viability under thermal stress in hydrated cells, LIVE/DEAD stain was employed. Cells stained with LIVE/DEAD were evaluated after 15 min thermal stress at 40°C, 45°C, 50°C, 55°C, 60°C, and 70°C.
2.3.2. Ultraviolet radiation
To investigate the effects of desiccation and liquid water availability on UV radiation damage, colonies and separated cells of Gloeocapsa sp. were placed into 4 cm diameter glass dishes and exposed to short-wavelength (254 nm) UV radiation by using a 3 W UV lamp (UV Products, California, USA) at a distance of 4 cm. UV radiation of wavelength 254 nm is not found in the natural environment, but in this experiment we use it as a proxy for general damage caused by natural short wavelengths of UV radiation. Colonies were either submerged in a ∼2 mm deep layer of BG-11 for hydrated cells or dried overnight on the bottom of the glass disc for desiccation experiments. Following UV exposure, the desiccated cells were rehydrated. Colonies and separated cells were either plated directly onto BG-11 agar plates as described above or examined for esterase activity under the microscope as described above.
2.3.3. Freeze-thaw stress
To determine whether desiccation prior to freezing provides an advantage to cells, aliquots of Chroococcidiopsis sp. were prepared in Eppendorf tubes. One set comprised 250 μL hydrated cells, and one set of desiccated cells was prepared by desiccating 250 μL of the same culture for 4 days in a desiccator. These aliquots were stored at −20°C, and every 24 h they were warmed to 21°C under laboratory conditions prior to being refrozen. After each freeze-thaw cycle, a triplicate of both hydrated and desiccated cells was removed and plated.
After 10 freeze-thaw cycles, aliquots of hydrated and desiccated cells were exposed to 15 min thermal stress at 40°C, 45°C, and 50°C to determine whether freeze-thaw cycles increased susceptibility to thermal stress.
2.3.4. Long-term (159 months) desiccation and thermal stress
A 1-month-old culture of Chroococcidiopsis sp., grown under the conditions described in Section 2.1, was plated into BG-11 agar to form a lawn of cells. The plates were incubated as described above for enumeration. Under aseptic conditions, the plate lids were removed and the cells allowed to desiccate over a period of 2 days. The lid was replaced on the plates containing the desiccated sheet of agar and cells, and the sheets were stored in darkness under laboratory conditions for 159 months (February 2003 to May 2016) with a temperature of between 15°C and 21°C and a relative humidity of between 20% and 60%.
After 159 months of desiccation, the status of the cells was examined on BG-11 to determine whether any of the cells remained viable. They were also examined for the presence of green pigmentation, autofluorescence, and esterase activity (as above). To study the effects of thermal stress after long-term desiccation, segments of the desiccated sheets were rehydrated, and cells were exposed to 15 min of thermal stress at 40°C, 45°C, and 50°C. Samples of desiccated sheet were also exposed to 70°C, 90°C, and 100°C to determine whether cells could survive long-term desiccation and transient extreme thermal stress.
2.4. Statistical methods
In all experiments, triplicates of each condition were examined. Student's t test was used to determine significant differences between given treatments with a p value of <0.1, which is considered to be statistically significant.
3. Results
3.1. Thermal stress experiments
The effects of thermal stress on Chroococcidiopsis sp. survivability at different temperatures in a desiccated and hydrated state is shown in Fig. 1. Desiccated cells of Chroococcidiopsis sp. showed survival to 90°C, but hydrated cells were completely killed at 60°C. In the desiccated state, between 45°C and 55°C, the number of colony-forming units showed an increase in the number of viable cells.
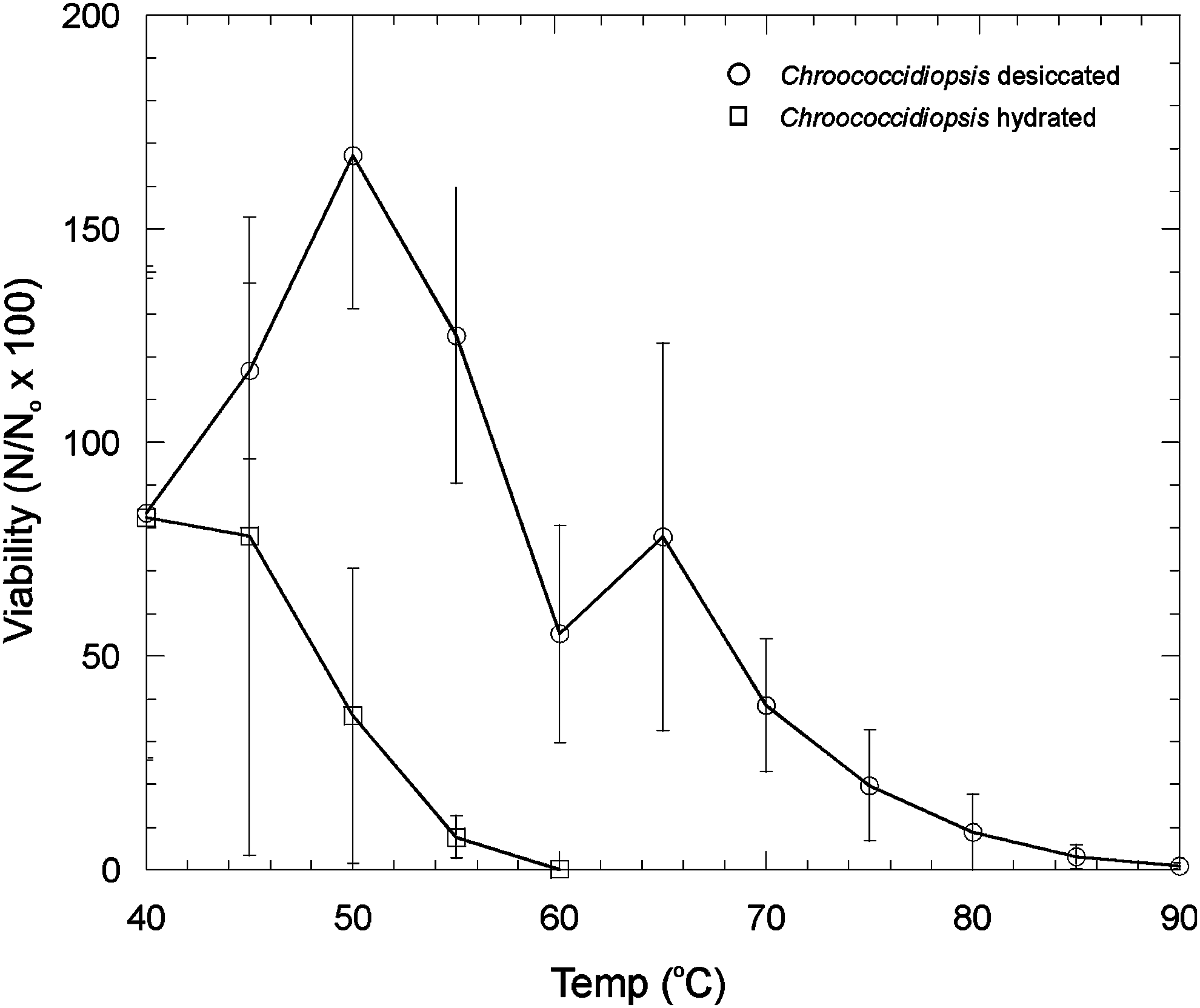
The effects of thermal stress on Chroococcidiopsis viability at different temperatures in a desiccated and hydrated state. Error bars are the standard deviation of triplicate measurements. Viability is expressed as follows: Number of viable cells after the given treatment (N)/number of viable cells in the control (N o) × 100%.
The effects of thermal stress on Gloeocapsa sp. survivability at different temperatures in a desiccated and hydrated state are shown in Fig. 2. Gloeocapsa sp. cell survival under desiccated conditions was observed up to 80°C, but no hydrated cells survived at 60°C.
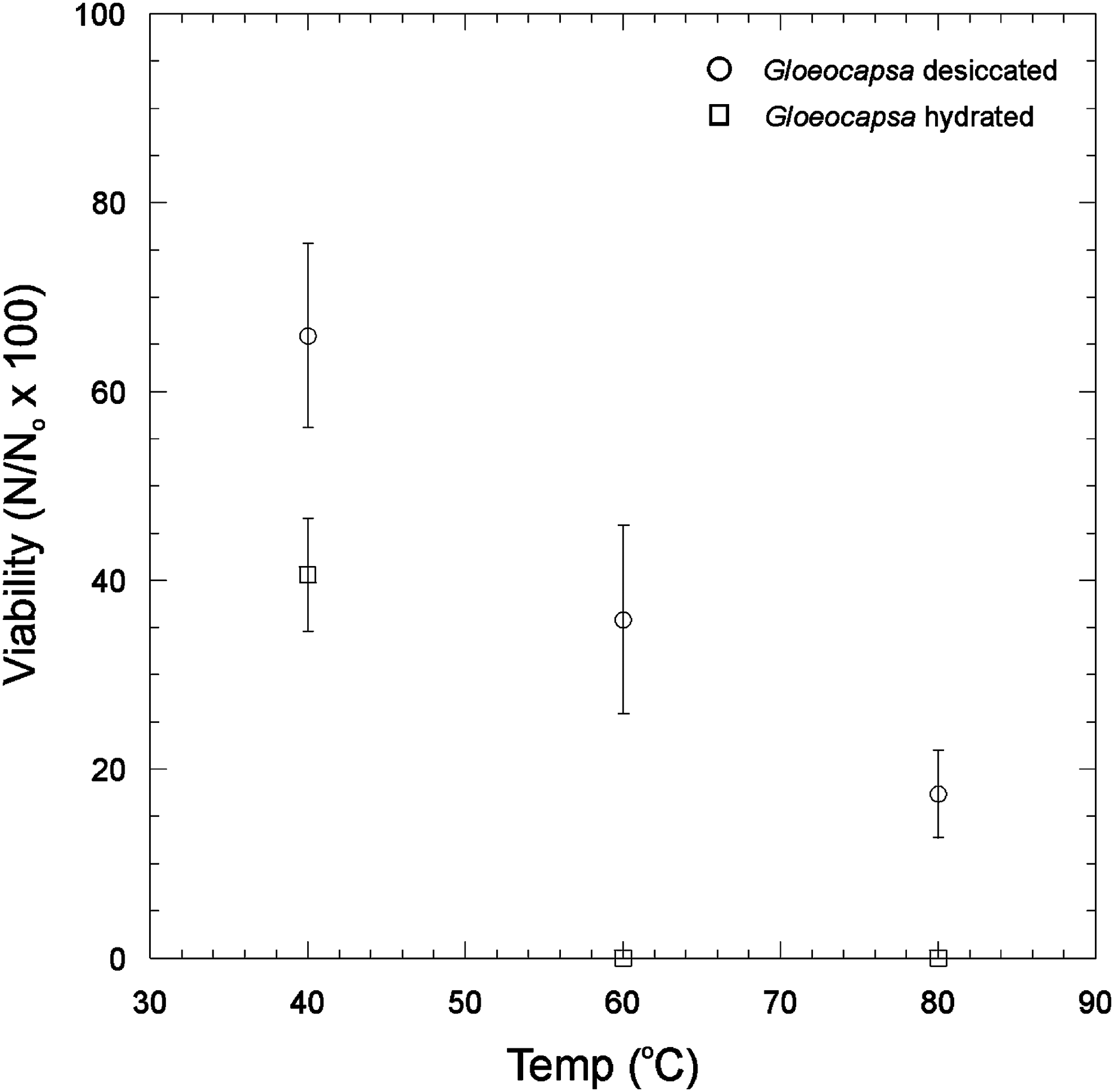
The effects of thermal stress on Gloeocapsa sp. on survivability at different temperatures in a desiccated and hydrated state. Error bars are the standard deviation of triplicate measurements. Viability is expressed as follows: Number of viable cells after the given treatment (N)/number of viable cells in the control (N o) × 100%.
In Fig. 3, the results of experiments to investigate the thermal stress of Gloeocapsa sp. at 2°C increments between 40°C and 50°C are shown. After 30 min thermal stress at 40°C and 42°C, there was no significant difference in viability compared to an untreated control (Student's t test; p = 0.98). At 44°C, there was no significant difference to an untreated control after 10 min of thermal stress (t test, p = 0.47), but viability dropped significantly for 15 min (p < 0.1) and 30 min stress (p < 0.1). At 46°C, significant loss of viability was observed after 10 min and for longer times thereafter (p < 0.1 for 10 min and all time points thereafter). At both 48°C and 50°C, a significant drop in viability compared to the untreated control was observed after 5 min stress (p < 0.1). Some cells remained viable after 30 min stress at 50°C.
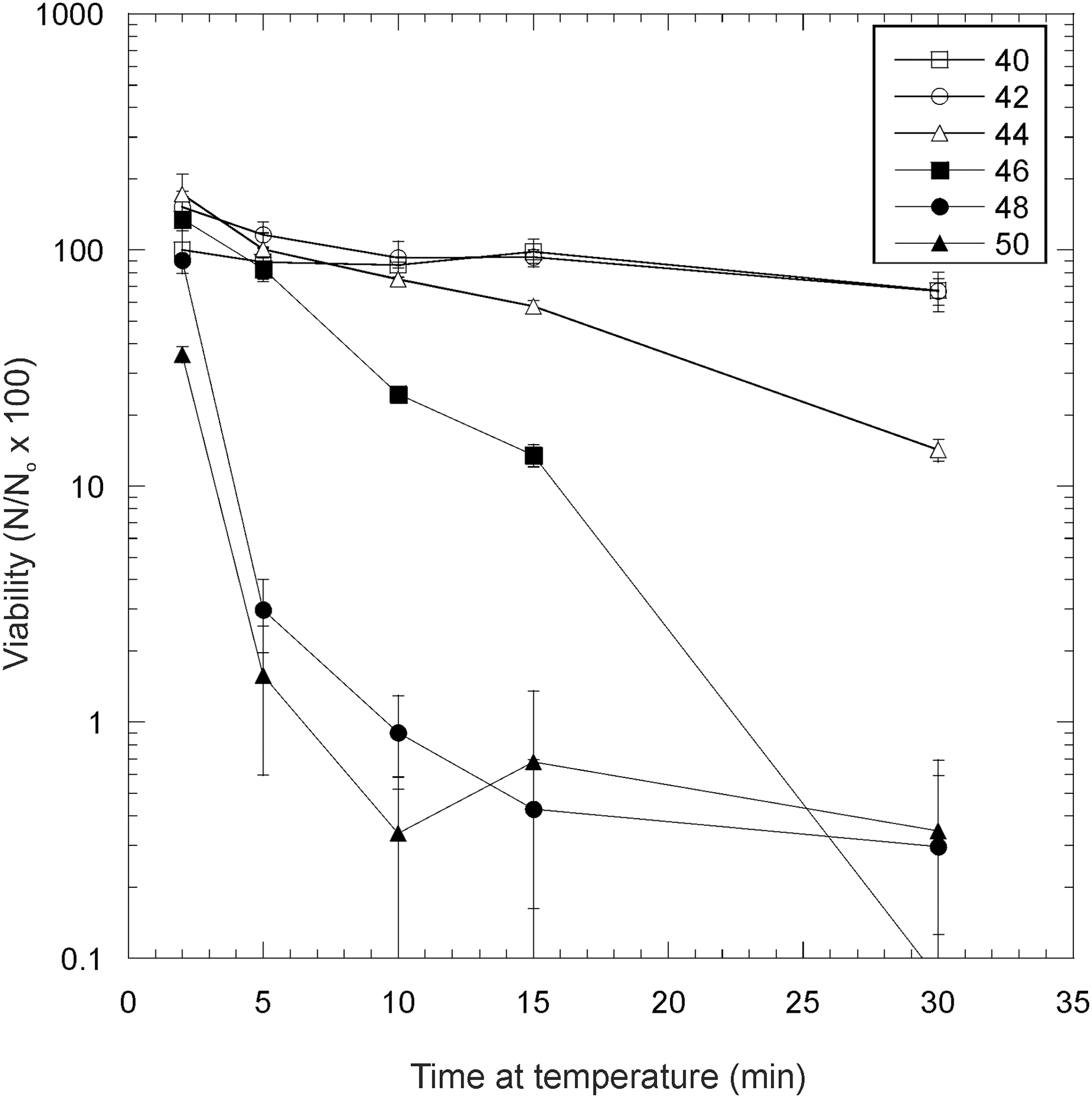
The effects of thermal stress on hydrated Gloeocapsa sp. at 2°C increments between 40°C and 50°C. Error bars are the standard deviation of triplicate measurements. Viability is expressed as follows: Number of viable cells after the given treatment (N)/number of viable cells in the control (N o) × 100%.
In Fig. 4, the effects of thermal stress on the esterase activity of Chroococcidiopsis sp. at 40°C, 60°C, and 80°C are shown. At 80°C, after all time points, there was a large reduction in measurable enzyme-active cells at 5 min and greater periods of thermal stress, which was statistically significant (t test, p < 0.1). The control was the percentage of enzyme-inactive cells in unstressed cells. Figure 5 shows the corresponding data for Gloeocapsa sp. Thermal stress at 60°C and 80°C caused a large reduction in measurable enzyme-active cells after 5 min and for all longer time periods of thermal stress, which was statistically significant (p < 0.1). In both Gloeocapsa sp. at 40°C and Chroococcidiopsis sp. at 40°C and 60°C, there was a statistically significant (t test, p < 0.1) reduction in the number of enzyme-inactive cells at 30 min. One possible explanation for this latter observation is that thermal treatment can enhance the uptake of FDA into a small proportion of cells that show inactivity not because of inactive enzymes but because of insufficient uptake of FDA. This might occur by partially solubilizing or weakening the extracellular polysaccharide layer.
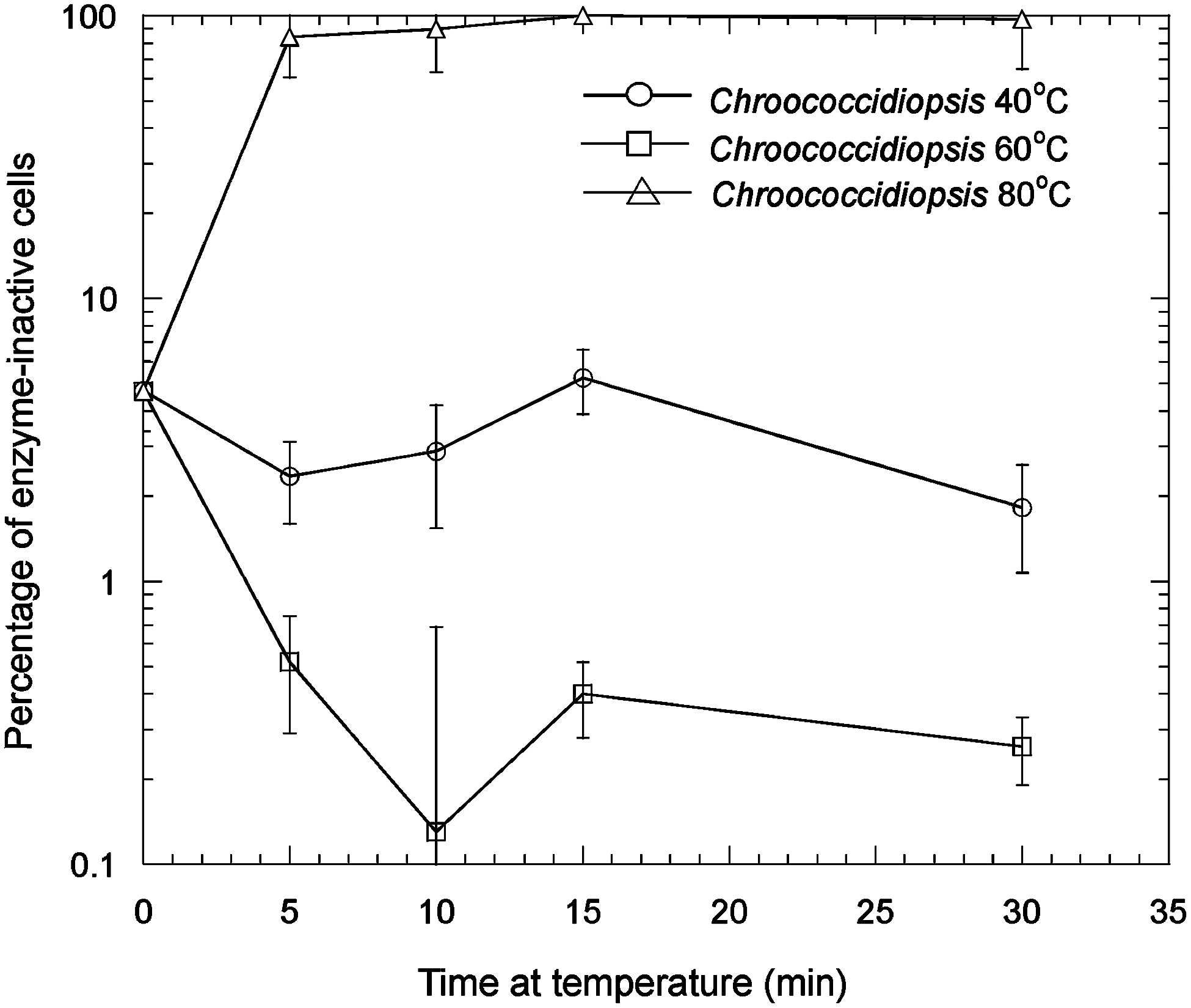
The effects of thermal stress on the esterase activity of hydrated Chroococcidiopsis sp. at 40°C, 60°C, and 80°C. Error bars are the standard deviation of triplicate measurements. Number of viable cells after the given treatment (N)/number of viable cells in the control (N o) × 100%.
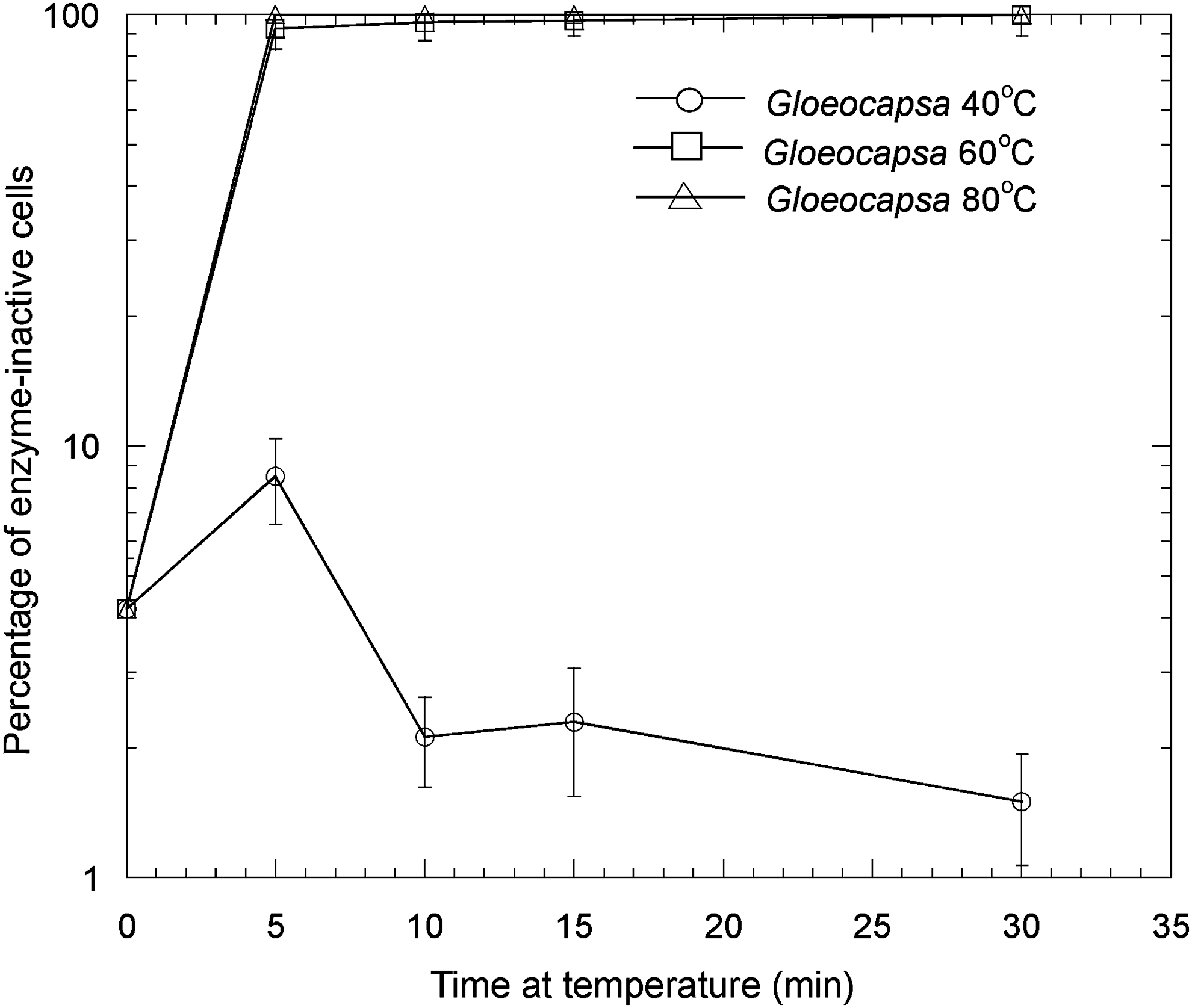
The effects of thermal stress on the esterase activity of hydrated Gloeocapsa sp. at 40°C, 60°C, and 80°C. Error bars are the standard deviation of triplicate measurements. Number of viable cells after the given treatment (N)/number of viable cells in the control (N o) × 100%.
LIVE/DEAD stain was employed to investigate the effects of thermal stress on cell membrane integrity. The percentage of cells showing viability according to the test was not statistically different in cells exposed to 15 min thermal stress at 40°C (89.7 ± 4.0% compared to the controls, which had 90.5 ± 5.3% viability), 45°C (89.6 ± 5.2%), and 50°C (91.7 ± 3.0%) (Student's t test, p > 0.1). However, at 55°C (81.3 ± 9.2% viability) there was just significantly less viability [p < 0.1 (0.09)], and at 60°C (29.0 ± 17.6%) there was also significantly less viability compared to controls (p < 0.05). No cells showed viability according to the test at 70°C.
3.2. UV radiation
In Fig. 6, the effects of UV radiation on the viability of Gloeocapsa sp. are shown for separated cells (isolated single cells and small cell clumps) of the organism and colonial forms of the organisms under desiccated and hydrated conditions. At 5 min and higher exposure times, there is a statistically significant difference between all treatments (t test, all values p < 0.1). Hydrated single cells showed higher UV tolerance than desiccated single cells. Desiccated colonies of cells had the greatest UV tolerance of all the growth forms and conditions. After 2 h exposure, only colonial forms survived, with desiccated colonies showing a significantly higher survival than hydrated colonies (t test, p < 0.1).
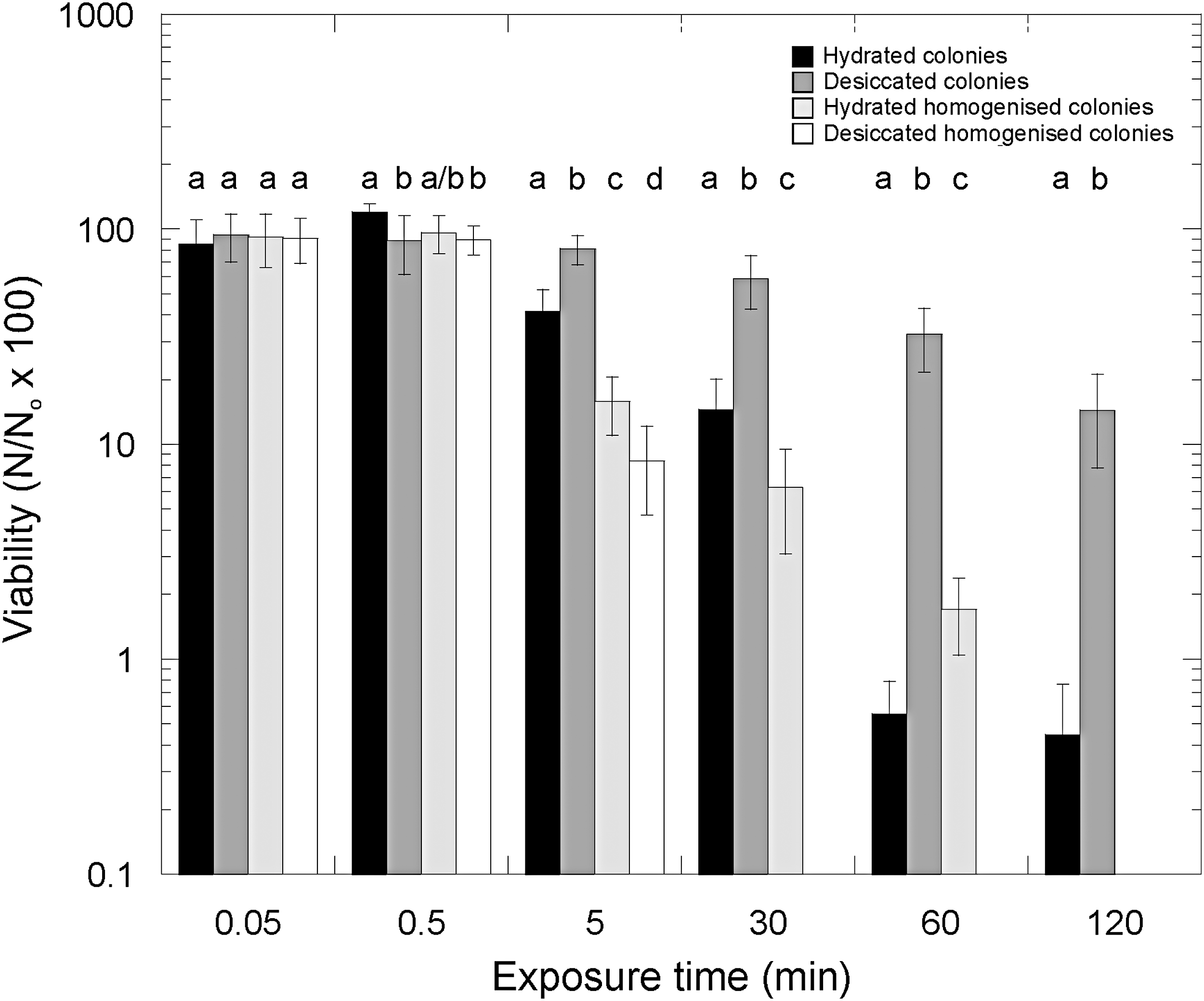
The effects of UV radiation (254 nm) on the viability of Gloeocapsa sp. for separated (isolated single cells here shown as homogenized colonies) organisms and colonial forms under desiccated and hydrated conditions. Letters above columns cluster columns with no statistically significant difference (p > 0.1) for each time point. Error bars are the standard deviation of triplicate measurements. Number of viable cells after the given treatment (N)/number of viable cells in the control (N o) × 100%.
Figure 7 shows representative images of the organism in colonial growth form taken under epifluorescence microscopy following FDA treatment. UV radiation has the effect of killing the cells on the outer layer of the colonies, but cells interior to the colony show enzymatic activity after 2 h. By contrast, colonies in which cells had been separated to expose interior cells were all killed after 2 h.
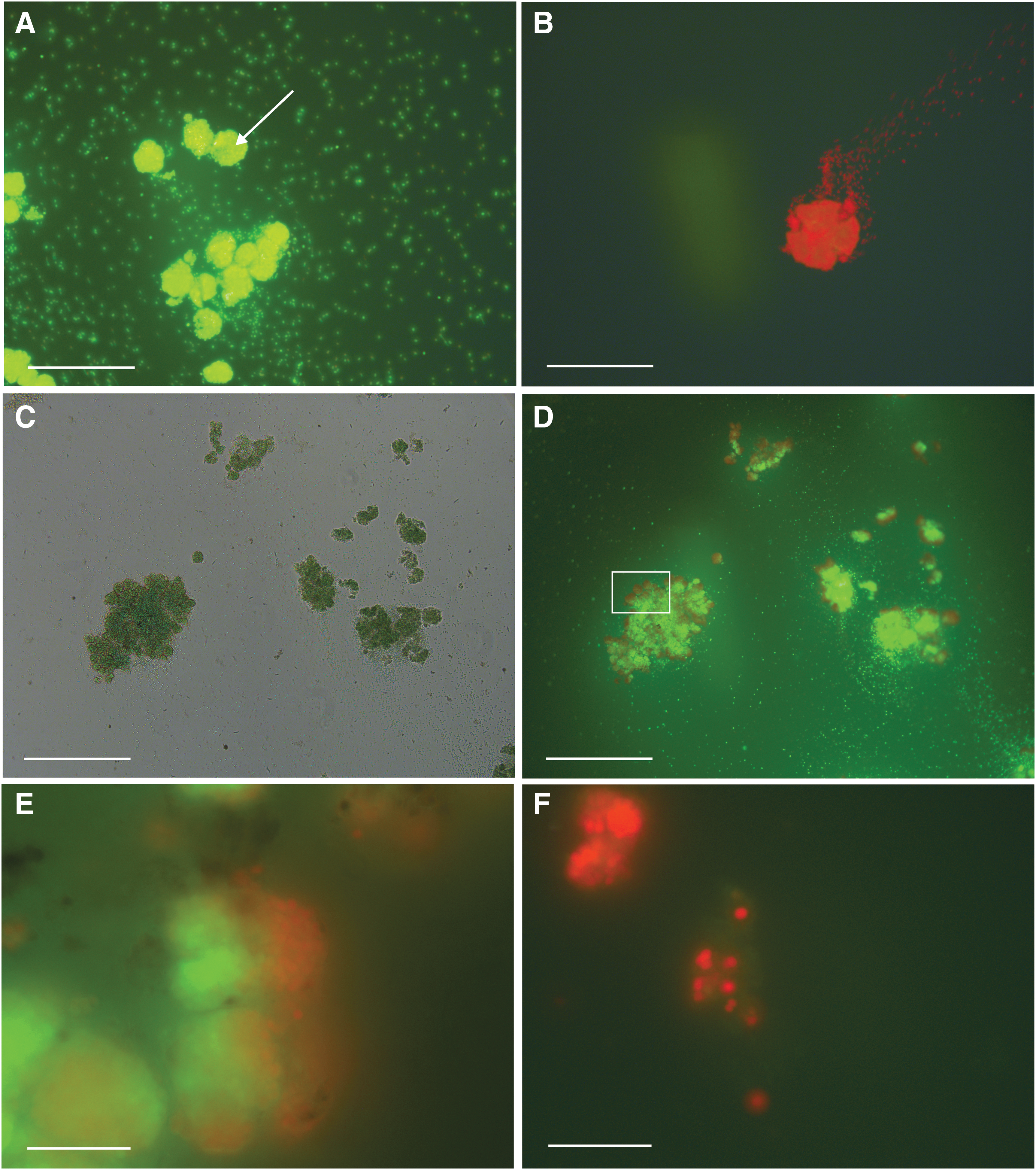
The effects of UV radiation on desiccated Gloeocapsa sp. cells. (
3.3. Freeze-thaw stress
Freeze-thaw stress was deleterious to both desiccated and hydrated Chroococcidiopsis sp. cells after 10 cycles (Fig. 8); however, hydrated cells had a much greater loss of viability. The mean viability of hydrated cells after 10 freeze-thaw cycles was 0.29 ± 0.17% of hydrated controls not exposed to freeze-thaw cycles. The mean viability of desiccated cells after 10 freeze-thaw cycles was 54.5 ± 29.7% of desiccated controls. Both of these losses of viability were statistically significantly different from that of the controls (p < 0.1). In the first three freeze-thaw cycles, the number of estimated viable cells in desiccated samples was greater than that of the controls, which we attribute to disruption of tetrads and multicellular growth aggregates.
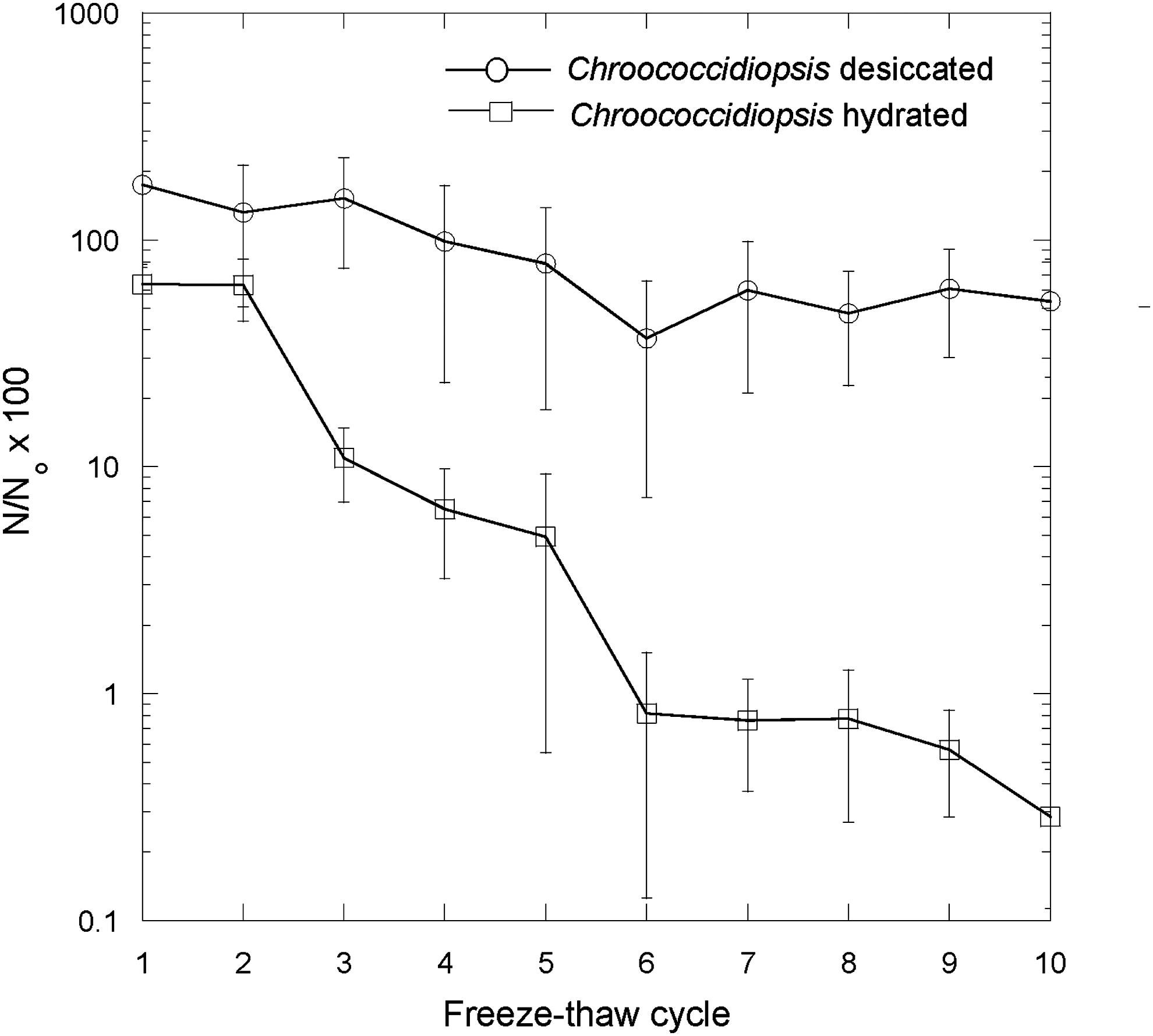
Effects of successive freeze-thaw cycles on the loss of viability of desiccated and hydrated Chroococcidiopsis sp. compared to hydrated and desiccated controls (cells prior to the first freeze-thaw cycle). Number of viable cells after the given treatment (N)/number of viable cells in the control (N o) × 100%.
After 10 freeze-thaw cycles, the thermal stress response of the cells after 15 min at 40°C, 45°C, and 50°C was qualitatively similar to that of cells unexposed to freeze-thaw (Fig. 1). The viability of hydrated cells, as indicated by Student's t test, that were subsequently exposed to stress at 40°C (0.26 ± 0.10%) and 45°C (0.34 ± 0.10%) after 10 freeze-thaw cycles was not statistically lower than the viability of cells that underwent 10 freeze-thaw cycles but were not exposed to heat stress. However, cells exposed to 15 min of 50°C stress had significantly lower viability (0.032 ± 0.05%, p < 0.1). Desiccated cells exposed to 15 min at 40°C (6.16 ± 1.21%), 45°C (3.92 ± 2.74%), and 50°C (5.29 ± 1.66%) after 10 cycles of freeze-thaw showed no significant loss of viability compared to cells that underwent 10 freeze-thaw cycles but were not exposed to heat stress (Student's t test, p < 0.1).
3.4. Long-term desiccation and thermal stress
Cells retained viability after 13 years and 3 months' desiccation (Fig. 9). After this time, cells were visualized as cell masses retaining green pigmentation and autofluorescence caused by phycobiliproteins (Fig. 9A–9C). Some cells also demonstrated esterase activity (Fig. 9D). After exposure to thermal stress at 40°C, cells showed no statistically significant loss of viability compared to controls (Student's t test), but cells exposed to 45°C and 50°C showed a significant (Student's t test, p < 0.1) loss of viability compared to controls (1.22 ± 2.11% and 2.1 ± 3.00% of controls, respectively), showing that long-term desiccation enhanced sensitivity of cells to thermal stress at 45°C compared to a fresh hydrated culture, which normally showed no statistically significant loss of viability at 45°C compared to controls.
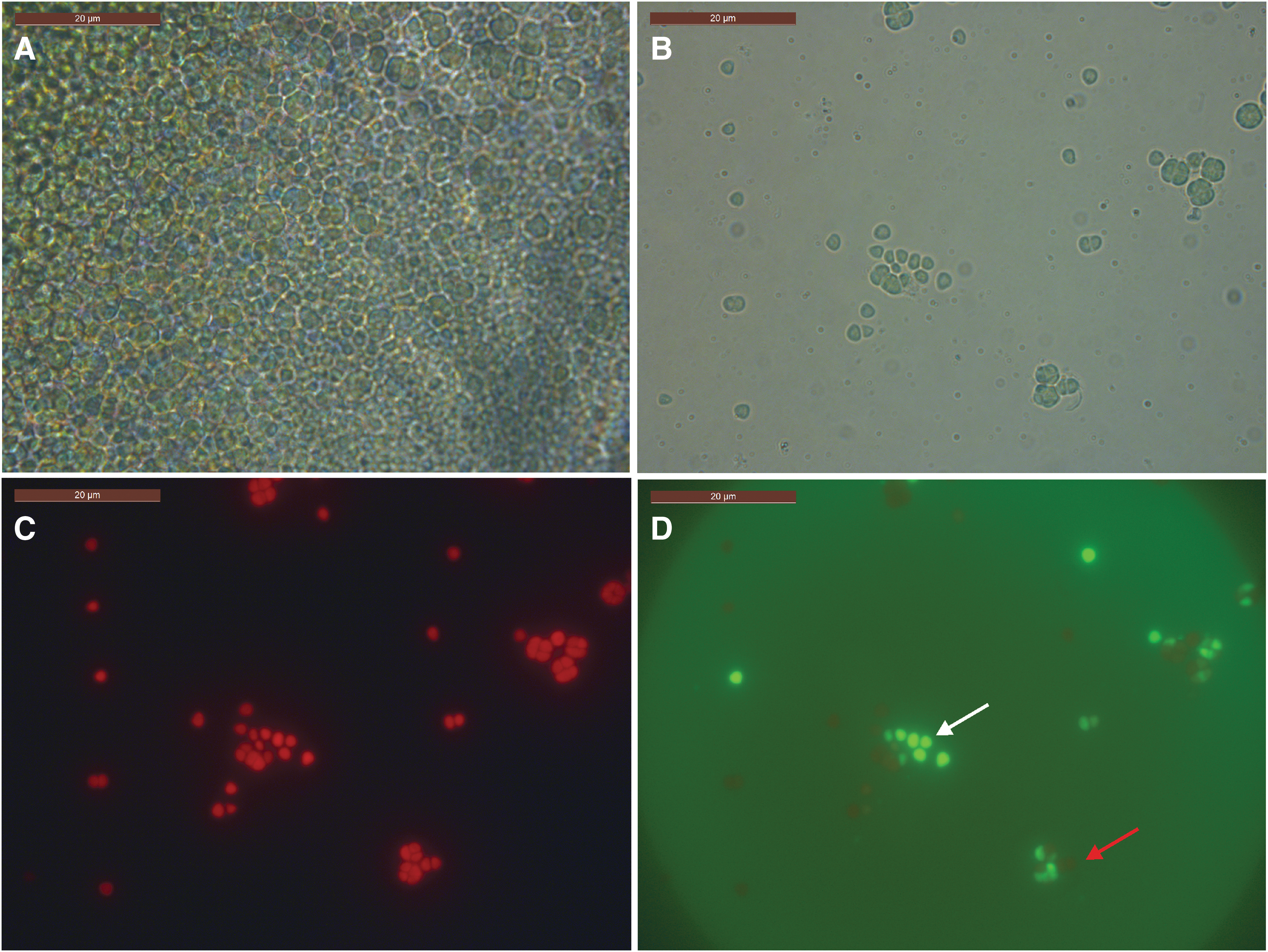
Chroococcidiopsis sp. exposed to 13 years and 3 months' desiccation. (
Long-term desiccated cells were successfully grown after exposure to 15 min at 70°C and 90°C but with viability less than 0.1% of controls. No cells survived 100°C (data not shown).
4. Discussion
Extreme hot and cold deserts are known to be uninhabitable to many microorganisms owing to extreme desiccation and, in equatorial deserts, accumulated solar (UV) radiation damage in surface environments (Nienow et al., 1988). Extreme temperature fluctuations impose additional stresses on desert organisms (Friedmann, 1980). The requirement for liquid water to allow growth and biochemical repair pathways (Mottl et al., 2007) leads to the paradigm that liquid water is beneficial to life and that deserts where liquid water is more available are therefore generally more habitable (Warren-Rhodes et al., 2006).
In this work, we show how liquid water can be deleterious in extreme deserts by increasing the damaging effects of certain physical extremes. Liquid water, therefore, defines a window of habitability in extreme environments where its beneficial uses in growth and reproduction are balanced against the advantages of its absence when those same organisms suffer exposure to extreme conditions, the damaging biochemical effects of which are exacerbated by liquid water.
The presence of liquid water increased the mortality of two different extremophilic cyanobacteria under high temperature stress. We found that the threshold at which cells were completely killed was increased by ∼40°C when cells were desiccated. Under desiccated conditions, thermal energy will be less effectively transferred to biomolecules through their kinetic energy and thereby less likely to cause disruption of essential molecular activity. However, at 50°C and 70°C even desiccated Chroococcidiopsis sp. showed some loss of viability after several hours of thermal stress.
Under hydrated conditions, it was found that the thermal range within which loss of viability occurs was very narrow. For example, after 30 min at 42°C, Gloeocapsa sp. cells showed no appreciable loss of viability. However, viability began to be lost after the same time period in cells exposed to 44°C, whereas those at 48°C and 50°C had lost more than 90% viability after 5 min. These data are consistent with the thermal limit of plant tissues. In Ranunculus glacialis, irreversible damage was done to the plant's chloroplasts at 45–46°C. This damage was attributed to the thermal damage of membranes manifested as the swelling of thylakoid membranes and general changes in organelle ultrastructure (Larcher et al., 1997). The mediation of heat shock through changes in membrane integrity has been observed in other organisms including microorganisms (Cossins, 1994; Vigh et al., 1998), and the response to heat shock can involve modifications to membrane composition (Balogi et al., 2005). However, our data, acquired by using LIVE/DEAD stain as a proxy for membrane integrity, show that cells apparently remain viable according to this test even after 15 min thermal stress at 60°C, after which no colonies can be cultured on BG-11 plates. This shows that cell membrane integrity remains even after cell viability is lost, which suggests that the thermal cause of loss of viability in Chroococcidiopsis sp. is not gross disruption of membrane integrity but some other component of the cell. The data we obtained suggest that LIVE/DEAD stain, at least in some cyanobacteria, may not be a reliable indicator of cell viability.
Investigations in which FDA was used to examine enzymatic activity of thermally stressed hydrated cells are consistent with the idea that denaturation of proteins can play a role in loss of viability (Inouse et al., 2001). Under hydrated conditions, within 5 min at 60°C, enzymatic activity was completely lost in Gloeocapsa sp. At 80°C, similar results were observed in Chroococcidiopsis sp.; however, esterase activity was still observed at 60°C, which suggests that not only might there be differences in susceptibility of enzymatic activities in different species of cyanobacteria, but also the loss of viability can occur within the cells before all enzymatic activity is completely lost. This is consistent with observations by Cockell et al. (2005), who studied Chroococcidiopsis under a simulated martian UV flux and found that cells lost viability while retaining esterase activity. The cause of cell death in cyanobacteria during thermal stress may involve oxidative stress, protein denaturation, and loss of membrane integrity (Cossins, 1994; Vigh et al., 1998; Balogi et al., 2005); however, the narrow temperature range in which cell viability is lost suggests that critical degradation of a few specific molecules is responsible for determining the threshold of thermal stress. The lack of this effect in desiccated cells could be accounted for by the more rigid nature of molecules that are less easily denatured and the lack of a milieu to generate and move around reactive oxygen species or to mediate fluid deterioration in membrane integrity.
Intriguingly, we observed an apparent increase in viable Chroococcidiopsis sp. cell numbers after thermal stress up to 55°C in desiccated cells (Fig. 1) and after the first three freeze-thaw cycles (Fig. 8). This finding might be attributed to the growth pattern of the cells. They frequently form tetrads during growth, and one plausible explanation for the results is that desiccation or freeze-thaw causes separation of the cells, which would lead to higher cell numbers when determined by plate counts compared to the controls in which tetrads and other multicell aggregates will generally appear as a single colony. As the organism has a doubling time of approximately 2–10 days, depending on growth conditions, the observations are not accounted for by cell growth during the course of the experiment.
We also investigated the influence of liquid water in UV radiation damage. We found that separated cells of Gloeocapsa sp., when exposed to UV radiation, were more quickly killed when desiccated compared to hydrated cells, yet in a colonial growth form, we observed the opposite effect. Cells had greater viability after UV irradiation when in the desiccated state compared to the hydrated state.
When exposed to UV radiation as single cells, hydrated cells have the advantage that they can actively repair accumulating damage, compared to desiccated cells, which cannot actively repair this damage that results in more rapidly accumulated lethal levels.
In contrast, in desiccated Gloeocapsa sp. colonies, cells on the upper part of the colony protect those at greater depth as shown by the esterase enzymatic assay. This is explained by attenuation, by both direct absorption and scattering, of UV radiation through the cell material. Even after 2 h exposure to 254 nm radiation, there were still viable cells in the interior of the colonies. However, when colonies are hydrated, the cells are less compact, and we hypothesize that UV radiation more effectively penetrates the colony interior and kills the cells, which leads to a lower viability. When hydrated, colonies were also liable to move around in the fluid by convection, leading to more effective exposure on the outside of the colonies. In extreme natural environments, cyanobacteria usually adopt biofilm growth habits where cells form a coherent layer either within rocks, under them, or in desert crusts (Belnap and Lange, 2001). Thus, the data we obtained with colonies are likely to be representative of cells in the natural environment. One other way in which liquid water can be damaging to cells under UV radiation stress is by producing photolysis products such as reactive oxygen species, products that can be catalyzed through the photo-Fenton reaction (Barrett and Baxendale, 1960; García-Fernández et al., 2012). This is likely to be one mechanism for damage induced under hydrated conditions.
The deleterious effects of water are also observed at low temperatures. Hydrated Chroococcidiopsis sp. were more susceptible to damage caused by freeze-thaw cycles than desiccated cells. One possible cause of this damage is the formation of ice crystals that could damage cell membranes or intracellular components. Previous research has suggested that, under natural situations, cells might vitrify, which would mitigate or prevent damage caused by ice crystal formation (Clarke et al., 2013). Nevertheless, the results show how liquid water can be deleterious in exacerbating the destructive effects of freeze-thaw.
We found that Chroococcidiopsis sp. desiccated for 13 years and 3 months retained viability and autofluorescence, and some cells retained enzymatic activity. They were also able to tolerate short-term 90°C thermal stress similarly to fresh desiccated cells. Chroococcidiopsis are known to tolerate desiccation (Potts, 1999) and have been recovered from 30-year-old hypoliths. The organism is also known to possess a range of physiological and biochemical properties, such as thick cell envelopes, that enhance desiccation resistance (Grilli Caiola et al., 1996; Billi, 2009) and a susceptibility to desiccation that can lead to death in some cells, but not in others (Billi, 2009). However, we found that when the cells were rehydrated they were more susceptible to thermal stress, showing that long-term desiccation compromises the ability of cells to deal with the deleterious effects of liquid water.
These experiments show that life in extreme deserts is precariously poised between two extremes. In one extreme, a lack of liquid water threatens cell death by accumulated unrepaired damage. In the other extreme, the presence of liquid water exacerbates certain physically and chemically induced damage such as extreme temperature and UV radiation stress.
These observations result in some predictions about the habitability of extreme deserts. In deserts where liquid water is present contemporaneously with high temperatures and UV radiation, damage to some organisms will be greater; thus deserts with these attributes will be less habitable to them compared to deserts where cells have a chance to desiccate prior to exposure to these extremes. For example, a desert where water is available in the form of fog in the early morning might allow for cell growth and replication. The dissipation of the fog prior to midday high temperatures and maximum UV radiation flux would allow cells to desiccate and better survive these extremes. A desert where liquid water persists in microhabitats until the afternoon or where precipitation occurs shortly before the maximum of UV irradiation and high temperatures, which would leave cells hydrated during these extremes, would expose them to greater stress. In addition, at the other end of the temperature scale, these data illustrate that, for deserts that experience diurnal freeze-thaw cycles, those that are characterized by more extreme conditions whereby cells are desiccated prior to freezing may be more habitable than deserts where greater liquid water availability causes cells to enter freeze-thaw cycles in a hydrated state. Thus, these observations predict the possibility of deserts that are less habitable to certain organisms than deserts that are more extreme and have lower liquid water availability. Some of the best places to isolate novel thermal and radiation-tolerant extremophiles or polyextremophiles with which to investigate new biochemical pathways are not necessarily the most extreme dry deserts. Instead, deserts where organisms must adapt to the extremes caused by physical stress combined with the presence of liquid water may provide more interesting insights into microbial stress responses and adaptations.
Finally, these experiments provide an example of how water is not always beneficial for life. Although the solvent has long been known to mediate damaging chemical reactions in life including hydrolysis reactions (Ball, 2007), these data show how the damage caused by the presence of liquid water under certain extremes might manifest at the ecological scale. Under the most challenging high-temperature conditions to be found in hot deserts (or, more speculatively, planets at the end of the habitable lifetimes transitioning into a greenhouse state; O'Malley-James et al., 2015), life is involved in a precarious balance in using its available energy and capabilities of adaptation to deal with the deleterious effects of both the lack and presence of liquid water.
Footnotes
Acknowledgments
H.L. received summer student funding support from the School of Physics and Astronomy. The Microbiology Society provided the Harry Smith Vacation Studentship for T.S.