
Abstract
As many of the methanogens first encountered at hydrothermal vents were thermophilic to hyperthermophilic and comprised one of the lower roots of the evolutionary tree, it has been assumed that methanogenesis was one of the earliest, if not the earliest, pathway to life. It being well known that hydrothermal springs associated with serpentinization also bore abiotic methane, it had been further assumed that emergent biochemistry merely adopted and quickened this supposed serpentinization reaction. Yet, recent hydrothermal experiments simulating serpentinization have failed to generate methane so far, thus casting doubt on this assumption. The idea that the inverse view is worthy of debate, that is, that methanotrophy was the earlier, is stymied by the “fact” that methanotrophy itself has been termed “reverse methanogenesis,” so allotting the methanogens the founding pedigree. Thus, attempting to suggest instead that methanogenesis might be termed reverse methanotrophy would require “unlearning”—a challenge to the subconscious! Here we re-examine the “impossibility” of methanotrophy predating methanogenesis as in what we have termed the “denitrifying methanotrophic acetogenic pathway.” Advantages offered by such thinking are that methane would not only be a fuel but also a ready source of reduced carbon to combine with formate or carbon monoxide—available in hydrothermal fluids—to generate acetate, a target molecule of the first autotrophs. And the nitrate/nitrite required for the putative oxidation of methane with activated NO would also be a ready source of fixed nitrogen for amination reactions. Theoretical conditions for such a putative pathway would be met in a hydrothermal green rust-bearing exhalative pile and associated chimneys subject to proton and electron counter gradients. This hypothesis could be put to test in a high-pressure hydrothermal reaction chamber in which a cool carbonate/nitrate/nitrite-bearing early acidulous ocean simulant is juxtaposed across a precipitate membrane to an alkaline solution of hydrogen and methane. Key Words: Green rust—Methanotrophy—Nitrate reduction—Emergence of life. Astrobiology 17, 1053–1066.
A likely impossibility is always preferable to an unconvincing possibility.
1. Introduction
M
Under this early view, pores in these mounds were the “culture chambers” in which life emerged (Russell et al., 1989, 1994). It had been further ventured, following Degens, that hydrothermal minerals would have “catalyzed” the synthesis of “peptides, polysaccharides, lipids, and nucleic acids, which would have been discharged to the hydrosphere” (Degens, 1979; Russell et al., 1989). This view was elaborated as “metabolism quickens, by many orders of magnitude, oxidation, and reduction reactions on our planet”… (and)… “links can be assumed between the rather slow, low-temperature reactions of geochemistry and the quickened reactions of early biochemistry” (Russell et al., 2003).
Following on from this supposition, Martin and Russell (2007) mooted the idea that microbial methanogenesis—assumed then to have been the most ancient form of archaeal metabolism—had emerged as a “quickening” of the abiotic serpentinization reaction presumed to occur at moderate hydrothermal temperatures. That methanogens occupied one root of the evolutionary tree and appeared to be thermophilic, or even hyperthermophilic, was considered to support the notion (Amend and Shock, 2001; Ciccarelli et al., 2006).
Yet, a nagging and anxious thought regarding the plausibility of these later ideas was: Why would methane be generated in competition with the high concentration of the same volatile in the alkaline effluent? That is, why would methane synthesis be driven against that concentration gradient? In addition, concerns arising from multiple sources—kinetic considerations, isotopic investigations, possible contaminations, biotic sources, and false positives in experiments, as well as phylogenetic analyses—have thrown doubt on the assumption that methanogenesis was foundational to the archael domain (Woese et al., 1990; Kelley and Früh-Green, 1999; Seewald et al., 2006; McCollom and Seewald, 2007; Proskurowski et al., 2008; Shock and Canovas, 2010; Lazar et al., 2012; Paukert et al., 2012; Nitschke and Russell, 2013; Reeves et al., 2014; Suda et al., 2014; McDermott et al., 2015; Seyfried et al., 2015; McCollom and Donaldson, 2016). Most arresting was the fact that Proskurowski et al. (2008) demonstrated an absence of radiocarbon in methane in the Lost City fluids, strongly implying that this methane is not derived from CO2 delivered by percolated and convecting ocean waters, but that any hydrocarbon source of the methane, or the methane itself, was intrinsic to, or had lodged within, the ocean crust and merely been released to, but not generated by, the circulating fluids.
That methane is a major volatile, second only to hydrogen in terrestrial alkaline springs, was already well known when the AVT was first formulated (Moiseyev, 1968; Neal and Stanger, 1984). Convecting, advecting, and/or artesian aqueous fluids fed from surface waters and exhaling from serpentinizing terrestrial ultramafic rocks are always alkaline and always enriched in hydrogen (≤25 mM) and methane (≤3 mM) (Abrajano et al., 1988; Etiope et al., 2012; Greenberger et al., 2015; Konn et al., 2015; Seyfried et al., 2015).
There is sturdy contextual and experimental evidence to suppose that the hydrogen was, and is, generated still through the exergonic oxidation of iron in olivine and, to a lesser degree, in the more recalcitrant orthopyroxene, by water (Coveney et al., 1987; McCollom and Bach, 2009), with recent research possibly adding iron-rich spinels to the mixture (Mayhew et al., 2013). It has also been argued that a proportion of the hydrogen so released goes on to reduce carbonate or bicarbonate, originally present in the same circulating fluid, to methane in a mechanism akin to the Sabatier reaction (Horita and Berndt, 1999; Neubeck et al., 2011; Wang and Gong, 2011; Etiope et al., 2012). Indeed, multiple investigations to determine whether methane could be generated through the reduction of CO2 species have been made and several showed that CO, CO2, or HCO3 − species can be reduced by iron–nickel alloys to form methane under low-temperature (<200°C) alkaline hydrothermal conditions (Horita and Berndt, 1999; Wang and Gong, 2011). Certainly, reduction of such precursors to methane, while facing substantial kinetic obstacles, is thermodynamically favorable (Shock, 1992; Maden, 2000; Lyons et al., 2005; McCollom and Seewald, 2007).
Moreover, we were aware from the calculations of Shock (1992) that serpentinization reactions taking place in off-ridge systems at moderate temperatures of 120–150° are more favorable to the thermodynamic drive to CO2 reduction than those at higher temperature—indeed, the lower the better in terms of the Gibbs free energy though not in terms of kinetics (cf., Herschy et al., 2014).
Yet, the recent article by McCollom and Donaldson (2016) has thrown what was a seemingly straightforward segue from geochemistry to biochemistry in some disarray. In experiments lasting >20 weeks, they failed to observe the generation of methane from CO2, thus placing the idea of methanogenesis as “an observable homologue at hydrothermal vents,” and thereby its foundational status, in doubt (pace Sousa and Martin, 2014). Compounding the uncertainties regarding the original AVT formulation is the suggestion of Windman et al. (2007) that hydrothermal formate, rather than hydrogen, was an initial fuel. Being thus forced to rethink our assumptions led us to attempt to make more sense of the stepped transitions from mineral to life–life that still extracts its inorganic components ultimately from mineral, while still being driven by redox disequilibria (Leduc, 1911; Nitschke and Russell, 2009, 2013; cf., McGlynn, 2017).
2. So What Were the First Fuels and Oxidants?
The original idea that life emerged autogenically into the very first autotrophs has a long pedigree (Traube, 1867; Pfeffer, 1877; Darwin, C, in Darwin, F, 1888; Haeckel, 1892; Mereschkowsky, 1910; Leduc, 1911; Goldschmidt, 1952)—a view beclouded for some 80 years by Oparin's and Haldane's primordial soup (Lane et al., 2010). Autotrophic views resurfaced in the context of submarine hydrothermal vents late last century with, for example, gradients across hydrothermal chimneys inducing the anaerobic utilization of H2 and native sulfur as well as the production and consumption of CH4 (Corliss et al., 1981; Baross and Hoffman, 1985). In a similar vein, Wächtershäuser (1988) called upon a putative pyrite reaction to supply electrons for CO2 fixation. In contrast, Russell et al. (1989) appealed directly to hydrothermally generated organic molecules as well as to hydrogen and methane as fuels.
However, since McCollom and Donaldson (2016) concluded that the reduction of CO2 all the way to CH4 during serpentinization may take thousands of years, we reconsider previous researches that assumed hydrothermal methane to have been variously (i) leached, or cracked, from primary abiotic and/or biotic organic precursors within the ultramafic crustal rocks, (ii) reduced from mantle-derived CO2 in the lower-to-mid crust and trapped thereafter in fluid inclusions, or (iii) even leached from under-thrust sediments and metasediments or Precambrian continental lithosphere. All these studies force a reassessment of what might have been the very first steps to life (Watanabe et al., 1983; Pineau and Mathez, 1990; Proskurowski et al., 2008; McCollom, 2013; Lollar et al., 2014; Suda et al., 2014).
As formate is the only rapidly produced organic molecule through CO2 reduction in conditions at, or simulating those at, an alkaline vent (Lang et al., 2010, 2012; Seyfried et al., 2015), and because McCollom and Donaldson (2016) also threw doubt on the assumption that hydrogen is produced rapidly through the serpentinization reaction, we no longer presuppose an immediate quickening or takeover of the hydrogenase reaction either! Perhaps the biological hydrogenation only came into play after the first steps of metabolism were mounted (Schuchmann and Muller, 2013). Otherwise, following Windman and coworkers' suggestion (Windman et al., 2007), formate could be considered as both a fuel and as another source of carbon.
Although these findings threaten the simple “rocky-roots,” or purely reductive form of the acetogenic pathway (Russell and Martin, 2004; Sousa and Martin, 2014), were CO2 to be reduced to only formate during serpentinization, then we are left with the tributary to the methyl group being derived oxidatively from CH4, although still arriving at the target molecule of the acetyl coenzyme-A pathway, acetate, through methylation of CO (Nitschke and Russell, 2013; Sojo et al., 2015). Thus, in our opinion, abandonment of the methanogenic aspect of the acetyl-coA pathway as the initiator of metabolism leaves only one alternative, namely, a denitrifying methanotrophic acetogenic pathway (DMAP) (Nitschke and Russell, 2013) (Fig. 1). Under this view, hydrothermal methane is considered a fuel for, rather than the exhaust of, emergent life. However, for this scenario to work requires the presence of sufficiently oxidizing electron acceptors.
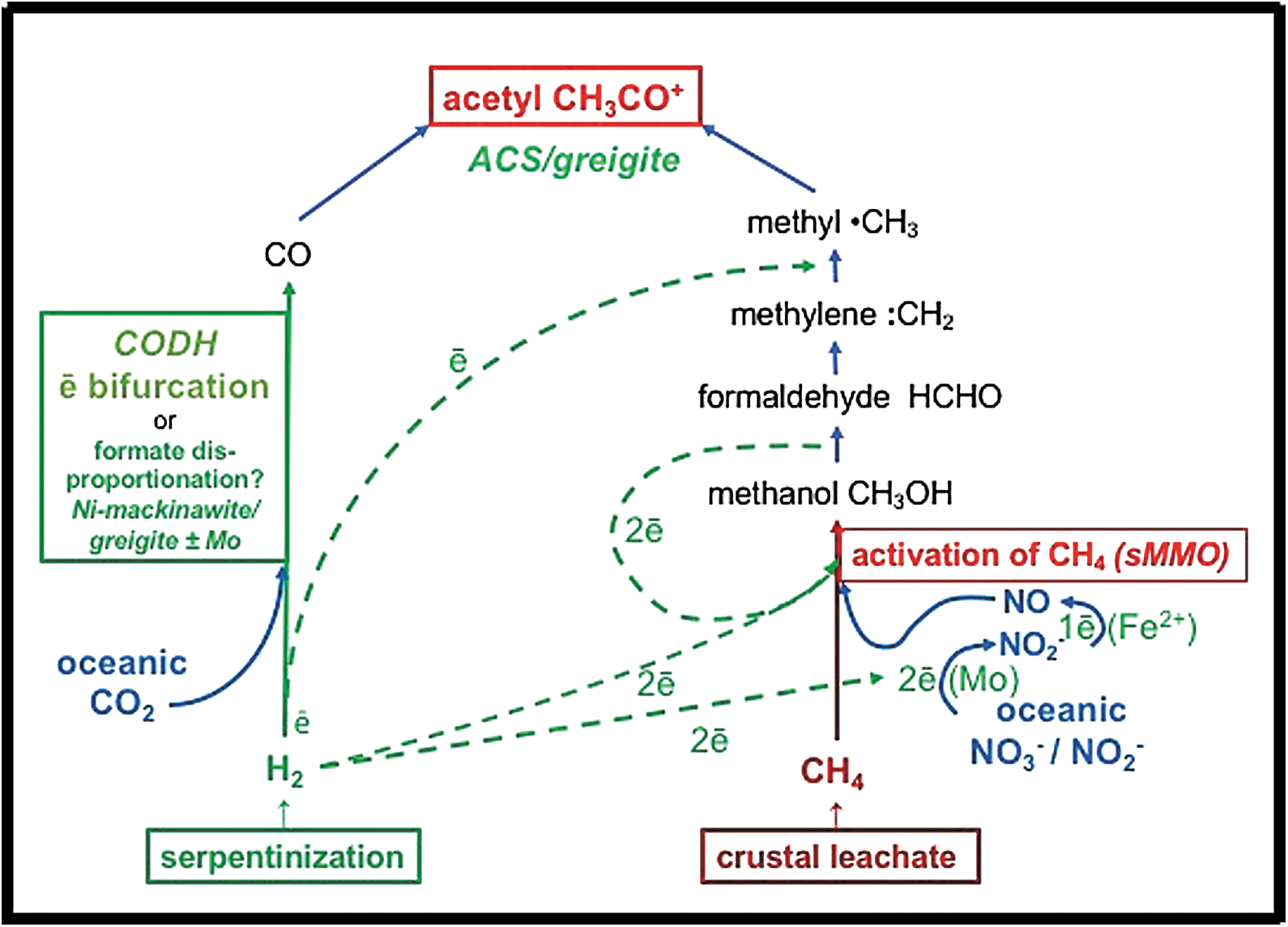
Simplified reaction steps of the putative denitrifying methanotrophic acetogenesis model based on Nitschke and Russell (2013, figures 3–5). Methane is produced by hydrothermal leaching of cracked carbon material previously residing in the crust. Hydrogen and formate are generated through the reduction of water during serpentinization. Hydrogen provides electrons for the reduction of HCO3 − to HCOO−. Formate disproportionates to CO and water as pH drops (Keene, 1993). The reduction of nitrate to NO (Kampschreur et al., 2011) drives the putative oxidation of methane to methanol and the hydrogenation of methylene to a methyl group. The methyl group reacts with the CO to produce activated acetate (Chistoserdova et al., 2009). Although this denitrifying methanotrophic (right hand) path to acetate looks complicated, the high electron mobilities (tunneling and bifurcations) within semiconducting GR allow improvization toward the best, if intricate, pathways and thereby long-range charge transport (Marcus, 1964; Wander et al., 2007; Nitschke and Russell, 2009, 2011, 2013; Ruby et al., 2010; Génin et al., 2012). Water and heat are among the waste products. The active sulfidic centers of metalloenzymes CODH (carbon dioxide dehydrogenase) and ACS (acetyl coenzyme synthase) are affine with the structure of nickeliferous greigite (Russell and Hall, 2006; Cao et al., 2009; Nitschke and Russell, 2013; Bassegoda et al., 2014; Roldan et al., 2015), whereas sMMO (soluble methane monooxygenase) is a di-iron enzyme with similarities to the structure of the more reduced form of GR, fougerite (FeII 4FeIII 2(OH)12CO3•3H2O) (Nitschke et al. 2013). GR, green rust.
Guided by results from geochemistry and extant biology (Mancinelli and McKay, 1988; Ducluzeau et al., 2009; Ettwig et al., 2010; Kampschreur et al., 2011), we consider that nitric oxide, released on the partial reduction of the nitrogen oxyanions nitrate and nitrite in green rust (GR) (“fougerite” sensu stricto, FeII 4FeIII 2(OH)12CO3•3H2O), is the most likely candidate for oxidizing methane in alkaline hydrothermal vent settings (Nitschke and Russell, 2013; Russell and Beckett, 2017). Again, in line with biology, we propose that methane was sequentially oxidized first to methanol and then to formaldehyde and maybe methylene. The view of a crucial role for nitrogen oxyanions and nitric oxide is given credence by estimations of NO pressure around a millibar at times in the Hadean atmosphere as in the work of Wong et al. (2017a)—enough to produce micromolar concentrations of nitrate and nitrite in the then ocean.
The fact that nitrate and nitrite can be reduced to NO with GR (Kampschreur et al., 2011) provides the molecule that, in extant life, is known to serve as oxygen donor in the oxidative formation of methanol (Ettwig et al., 2010). Counter-intuitively, the oxidation of the notoriously unreactive methane to methanol in biology requires a prior overreduction of the catalytic center binding both the methane and the oxygen donating molecule (O2 or NO). The redox potentials of the involved reactions are such that this prereduction can be performed either by electrons released from the subsequent oxidation steps (fed back as methanol is oxidized to formaldehyde and formaldehyde to methylene) or from hydrogen oxidation (Nitschke and Russell, 2013).
The reaction scheme both in biology and in our proposed scenario, therefore, is highly networked and contains numerous (autocatalytic) feedback loops (Fig. 1). We address the pertinence of this networking hereunder. The ultimately generated activated methyl is proposed to react with CO produced by the branch shown on the left-hand side of Figure 1. If, as we have proposed previously, during the emergence of life, this branch resembled that of extant organisms, then the electron donor to CO2 would be molecular hydrogen. However, the reduction of CO2 to CO by H2 is highly endergonic and involves the phenomenon of electron bifurcation in the respective organisms (Buckel and Thauer, 2013). Consequently, we have presumed that electron bifurcation may have played a corresponding role during life's emergence and that the two-electron redox metals molybdenum and tungsten might have been the primordial electron bifurcating agents (Nitschke and Russell, 2013). Recent results on the electrochemical properties of molybdopterin enzymes corroborate the possibility of such scenarios (Duval et al., 2016).
However, that carbon monoxide can also disproportionate from formate—known to be present in the alkaline vent fluids—under the slightly acidic conditions met with on the ocean side of the mound, adds a further (alternative or concomitant) source of CO to the system (Lang et al., 2010). Formate-derived CO possibly provides the kick-starting for the two-pronged reaction scheme inherent in the Wood–Ljungdahl reactions common to methanotrophy, methanogenesis, and acetogenesis. The prize would be the provision of the ammonium ion for amination reactions within the reactor pile and associated chimneys through the concomitant reduction of the nitrate and/or nitrite (Fig. 2) (cf. Mancinelli and McKay, 1988; Haroon et al., 2013; Arshad et al., 2015).
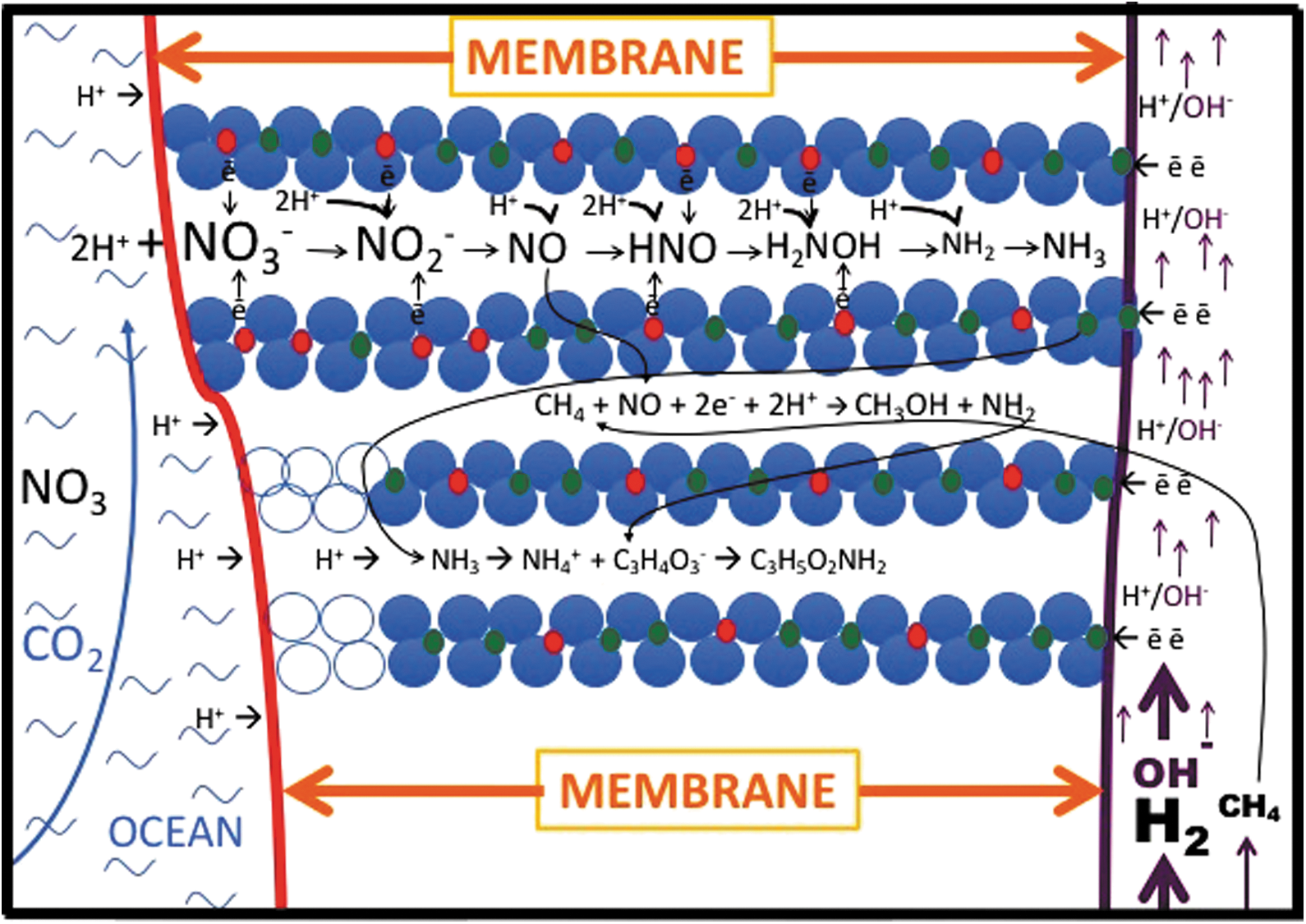
Model of GR as a ready-made difunctional enzyme precursor set in the inorganic membrane wherein it reduces nitrate to aminogen or ammonium between the “brucite” galleries (Trolard and Bourrié, 2012). At the same time, methane would be converted to a methyl group by NO (Kampschreur et al., 2011)—a yet-to-be-tested hypothesis. Flores et al. (2016) showed that in these same circumstances, pyruvate can be aminated to alanine. The inorganic membrane or barrier, represented in this case by GR, separates the alkaline hydrothermal solution on the right, from ocean water on the left. Green rust was precipitated from the vast amounts of metastable iron precursors in the early oceans on meeting alkaline solutions such as those issuing from the vent (Arrhenius, 2003; Mielke et al., 2010; Tosca et al., 2016; Halevy et al., 2017).
3. The Pertinent Disequilibria (Free Energies) and Their Conversion
Baross and Hoffman (1985) foresaw that “a multiplicity of physical and chemical gradients” was to be expected “as a direct result of interactions between extensive hydrothermal activity in the Earth's crust and the overlying oceanic and atmospheric environments.” However, these gradients, especially pH and redox, would have been much more pronounced at putative submarine alkaline springs (Russell and Hall, 1997). There are two autotrophic free energy conversion mechanisms recognized as fundamental to driving life's endergonic operations, the proton motive force (pmf)/chemiosmosis to generate free energy for the cell (Mitchell, 1961, 1976; Boyer, 1975; Harold, 2001), and redox bifurcation whereby two electrons of the same energies may be released to single electron acceptors—one to a high potential acceptor and the other, “hot” electron, to reduce an otherwise well-defended low-potential acceptor (Baum et al., 1967; Wikström and Berden, 1972; Mitchell, 1975, 1976; Kovacs, 1989; Iwata et al., 1999; Thauer et al., 2008; Nitschke and Russell, 2009, 2011, 2013; Kaster et al., 2011; Chowdhury et al., 2016; Peters et al., 2016).
In the AVT, the emergence of the metabolic system was relieved of the necessity to pump protons out of the first compartments or protocells to generate the pmf, because a pre-existing steep proton gradient would have been imposed in the barriers from the acidulous Hadean ocean across to the alkaline interiors. Such a gradient is assumed, following the works of Baltscheffsky (1971), Baltscheffsky and Persson (2014), and Baltscheffsky et al. (1999), to have driven a pyrophosphatase, a primary ion pump with structure recently revealed by Kellosalo et al. (2012) and Tsai et al. (2014). We have appealed to GR as the flexible mineral precursor PPase to produce “the energy currency” of the cell (Lane, 2010; Branscomb and Russell, 2013; Russell et al., 2013, fig. 5). Could the same mineral have played some part in the putative DMAP by oxidizing the methane to a methyl group in these hydrothermal conditions?
For the GR mineral, “fougerite,” to engineer the exergonic oxidation of methane to methanol—a kinetically highly challenging reaction given the stability of the methane molecule—requires an oxygen activated by adjacent ferrous irons within the double layers. Activated nitric oxide, formed as an intermediate in the reduction of nitrate or nitrite to ammonium within the galleries of GR, is the likely candidate (Hansen et al., 2001; Ducluzeau et al., 2009; Kampschreur et al., 2011; Wu et al., 2015; Wong et al., 2017a). Further indications that GR could act as a potential mediator of such a reaction are (1) because of its structural similarity to the metal cofactor of soluble methane monooxygenase and (2) for its variable valence (Nitschke and Russell, 2013; Nitschke et al., 2013; Banerjee et al., 2015). Moreover, the two oxygen atoms forming the diamond center are both derived from O2 (Banerjee et al., 2015). However, as nitric oxide is isoelectric with dioxygen and, moreover, is slightly asymmetric, and has similar interaction properties as O2 (Pilet et al., 2004), it is potentially as strong a candidate as oxygen to activate the methane bound to the metal site for oxidation.
Although GR itself has never been shown to oxidize methane to methanol, a bent mono(μ-oxo)di-nickel anchored within a synthetic zeolite does promote such an oxidation at 150°C (Shan et al., 2014; and see Starokon, et al., 2013). Clearly, a demonstration of methane oxidation by nitric oxide intercalated within GR or other iron-rich double layer hydroxide (DLH) in hydrothermal conditions must be demonstrated for the denitrifying methanotrophic acetogenesis hypothesis to survive (Figs. 2 and 3).
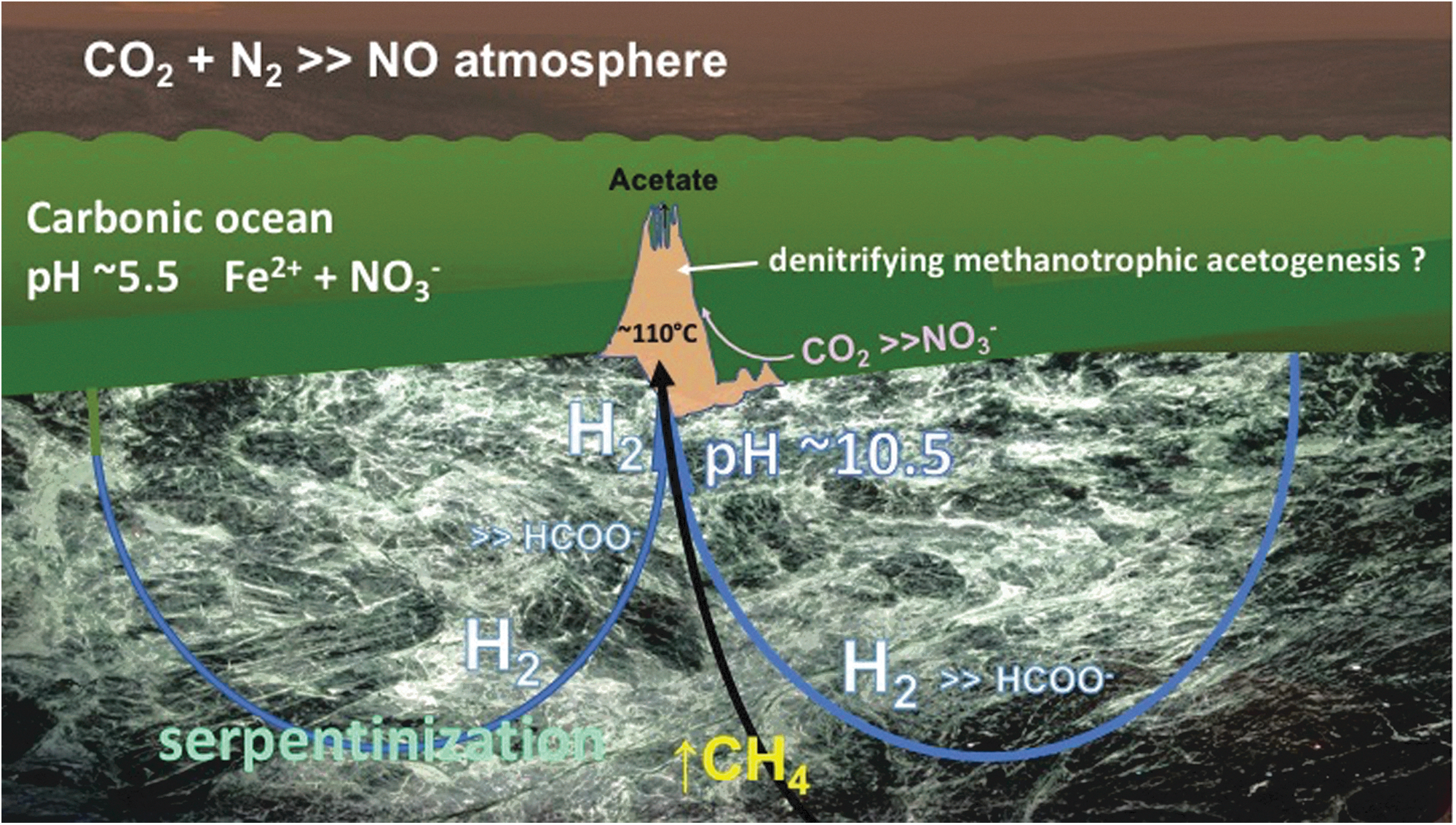
Schematic representation of alkaline hydrothermal vent model for the early emergence of life on Earth through a putative denitrifying methanotrophic acetogenic pathway fed from H2 generated through serpentinization while methane is leached from ambient reduced carbon molecules residing in the crust (Russell and Hall, 1997; Proskurowski et al., 2008; Nitschke and Russell, 2013).
As mentioned, furnishing CO from CO2 in the left-hand branch of Figure 1 is highly endergonic and thus requires redox bifurcation to occur. For this to happen, a molybdenum atom (or atoms) would then need to be hosted in the DLH (Nitschke and Russell, 2009, 2013). We have speculated that a molybdenum sulfide complex [FeO(OH)(MoS4)2]3− (Helz et al., 2014), sequestered in the interlayers of GR or other DLH, would be secured through hydrogen bonding, much as an H-bond network surrounding the pyranopterins in the molybdenum site modulates two-electron redox properties in arsenite oxidase (Itaya et al., 1987; Russell et al., 2014; Duval et al., 2016).
4. Astrobiological Implications
That methane concentrations at alkaline vents and volcanic eruptions are so low on Earth is because of the surprisingly high oxidation state of Earth's mantle throughout geological history; this is surprising because the planetesimals and meteorites from which it accumulated are generally buffered below iron-wustite and contain reduced organic compounds (Shock, 1992; McSween and Huss, 2010; Dale et al., 2012; Mousis et al., 2015). Wood et al. (2006) explained this relatively high state by recognizing the tendency of ferrous iron (mainly in olivine) to disproportionate in the presence of perovskite which is stable at depths below the 660-km discontinuity, that is, at 24 GPa and 1900 K (McCammon, 1997; Wood, 2000; Chudinovskikh and Boehler, 2001; Frost et al., 2004). The mantles of smaller bodies would be at much lower pressure and, therefore, more reduced (Wadhwa, 2008; Dale et al., 2012; Gaillard et al., 2015). The resultant low-oxygen fugacities would explain the preponderance of methane on some of the moons of Saturn (Waite et al., 2009; Bouquet et al., 2015; Glein et al., 2015; Dorofeeva, 2016), possibly Europa (Goodman et al., 2004; Hand et al., 2007; Zolotov and Kargel, 2009) and, perhaps, even early Mars (Wadhwa, 2001; Hirschmann and Withers, 2008; McSween et al., 2009; Blamey et al., 2015; Edwards and Ehlmann, 2015; Hu et al., 2015).
As with the early Earth, the icy moons would, while producing copious reductants, soon face an oxidant crisis. Denitrifying methanotrophic acetogenesis might well be the emergent metabolism on such worlds, with nitrate/nitrite produced in the ice shell through the oxidation of ammonia with hydrogen peroxide entrained from the surface as electron acceptors (Loeffler and Hudson, 2015; Russell et al., 2017).
5. Discussion and Issues Raised
Our main aim of this work is to reconsider our previous expectations of a relationship between the geochemistry of moderate temperature serpentinization with that of early biochemical pathways, namely, was biochemical methanogenesis derived from a rapid and facile geochemical reduction of CO2 to CH4, that is, from abiotic methanogenesis (Martin and Russell, 2007)? It was reasoned that such a reaction had to be rapid enough compared with the product dissipation rate to maintain a self-organizing protometabolic pathway and support continued growth. In other words, it would need to happen within minutes to hours if it were to be the harbinger of enzyme-catalyzed reactions that are generally measured in milli- to microseconds or less (Garrett and Grisham, 2012, page 17).
It follows that the results reported from various laboratories and field sites, which we accept, militate against the idea that biochemical methanogenesis emerged partly through a quickening of geochemical reactions. Moreover, the thermodynamic calculations of Shibuya et al. (2016) estimate the delivery of formaldehyde and acetate from hydrothermal systems generated by serpentinization to be 9.7 × 10−27 and 8.6 × 10−24 molal, respectively. Formaldehyde, which anyway can be difficult to analyze and is a significant contaminant, has not been registered in serpentinization experiments (Borowska and Mauzerall, 1991; Barro et al., 2009). And Lang et al. (2010), analyzing acetate at Lost City, found concentrations between 1 and 35 μmol/kg for which they tentatively assumed a microbial derivation.
Another point at issue is whether hydrogen was the main fuel as we and others have claimed, or whether hydrothermal formate fulfilled that role as suggested by Windman et al. (2007). As mentioned, formate will disproportionate to CO and water on being driven through the inorganic hydrothermal mound toward the acidulous ocean. CO is then available to react with the methyl group produced through the supposed oxidation of methane as already alluded. An alternative, or early derivation, of this tributary to the DMAP would be a reverse of the formate hydrogenlyase (FHL) reaction (Andrews, 1997), whereby the proton gradient (acidulous ocean juxtaposed to the alkaline, hydrogen-bearing interior) would drive the generation of formate from CO2 with the two electrons supplied from H2 or HCOO− through a molybdenum-dosed nickel-rich mackinawite (FeNiS2)n or greigite ([FeNi]3S4)n nanocluster that acts as a hydrogen store and protohydrogenase (Cao et al., 2009; Nitschke and Russell, 2009, 2013; Bassegoda et al., 2014; Yamaguchi et al., 2014; Wang et al., 2015; White et al., 2015; Wilkin and Beak, 2017). Pinske and Sargent (2016) demonstrated unambiguously that FHL enzyme is bidirectional, so giving some strength to this hypothesis (and see Roger et al., 2017).
A theoretical assessment of whether molybdenum-bearing GR can act as the engine to drive the DMAP would require ab initio molecular dynamic simulations to indicate how redox changes in GR involving electron and proton transport compare with methane monooxygenase. From our ideas about the quickening of abiotic serpentinization reductions, it seems imaginable that such a rapid and facile reduction of CO2 to HCOO− could be taken over by emerging life as one step in carbon fixation. Nevertheless, these results also threaten the facile view that hydrogen was the very first fuel for life, that is, the electron donor in a redox chain that can then drive other chemistry (Windman et al., 2007). Contrarily, it is extremely unlikely that CO2 was hydrogenated directly to methane. 12CH4 is presumably distilled, or cracked, from organic molecules previously lodged in crystalline and sulfidic source rocks (Strauss, 1989). So, although it had been commonly considered that archaeal methanogens—exploiting or even discovering the reductive acetyl-coA pathway—occupied the very lowest branches of the evolutionary tree, recent experiments show that this idea can no longer be entertained (cf., Koonin and Martin, 2005; Russell et al., 2005; Martin and Russell, 2007; Wolfe, 2014; Shock and Boyd, 2015).
At lower redox and pH, there is a transition from HCOO− to CO (Keene, 1993). Huber and Wächtershäuser (1997) showed how, given a methane thiol, acetate can be generated by the comparable Monsanto reaction (Crabtree, 1977). Huber and Wächtershäuser (1997) called upon a mechanism proposed in an article by Heinen and Lauwers (1996) to supply the methyl group, supposedly generated through the reduction of CO2 in water in the presence of H2S and FeS at low pH. However, the yields are ∼0.1% or less of the available H2S (as HS−), which itself is only one millimole (and see Schoonen et al., 1999 for a critique). So even supposing all the hydrogen sulfide is converted to methyl sulfide, this would provide <1 nmol/L—hardly enough to satisfy the ∼50 mmol/L of formate to produce activated acetate.
The only way for some form of the acetyl-coA pathway to survive as the first carbon fixation hypothesis is for the methyl group to be provided from a different origin. Alert to this dilemma, it has been speculated that the necessary methyl group was provided by the oxidation of the juvenile methane by Fe3+-rich GR at the alkaline hydrothermal mound (Nitschke and Russell, 2013; Russell et al., 2014; cf., Scheller et al., 2016). This explanation is amenable to experimental falsification (cf., Diaz-Campos et al., 2009). Wu et al. (2013) demonstrated the direct synthesis of acetate from CO2 and methane on a zinc-modified aluminosilicate zeolite, and the idea gleans some further support in that methane can be oxidized to methanol at 160°C on a synthetic iron-bearing zeolite previously calcined in the presence of nitrous oxide at 200–250°C (Starokon et al., 2013).
As we have seen, the initially assumed hydrogenation of CO2 to methane by H2, proposed to then segue into methanogenic metabolism, probably does not occur at significant velocities and yields. Moreover, it is also comparatively poorly suited for rationalizing life's emergence in the framework of the laws of the physical world. As pointed out half a century ago (Schrödinger, 1944), the second law of thermodynamics imposes the requirement that life, and a fortiori its emergence, is possible only as a subset of a larger system featuring strong thermodynamic disequilibria. The formidable entropy decrease inherent in living things can thus be fueled by low-entropy sources from their environment. The entropy of the entire system will then increase despite a local decrease in the living subset.
Although the second law (Boltzmann's entropy law) specifies under which conditions life is, or is not, possible, it does not provide a mechanism to rationalize its emergence. However, recent developments achieved, and the insights gained, within the field of far-from-equilibrium thermodynamics do indicate the kind of mechanism required to bring about the onset of life (Branscomb and Russell, 2013; Branscomb et al., 2017). Systems far from thermodynamic equilibrium have indeed been shown to frequently generate substantially ordered and self-organizing states as they relax toward equilibrium, that is, while dissipating free energy (Prigogine and Nicolis, 1967, 1989).
The fields of rheology, atmospheric sciences, or networked chemical reactions are abound with examples of such “dissipative structures.” These feature many of the defining properties of living systems, and the emergence of a dissipative structure, “life,” under appropriate conditions of high disequilibria appears as a quasi-necessity in the theoretical framework of far-from-equilibrium thermodynamics. In addition to strong free energy gradients characterizing the system, far-from-equilibrium thermodynamics has worked out further important parameters that favor the emergence of dissipative structures: (a) nonlinear equations of motion in reaction phase space and (b) reaction feedback loops (note that mathematically (a) and (b) are often intimately correlated with the high ΔG criterion).
How do the two alternative hypothetical founding metabolic reactions in which methane appears either as an exhaust (methanogenesis) or as a fuel (methanotrophy) fare against the test of the already listed criteria? The reaction hydrogenating CO2 to methane certainly features a substantial negative ΔG under most conditions (i.e., concentrations of reactants and pH values) likely encountered in alkaline hydrothermal vents of the Hadean (Amend et al., 2013). However, this is the total ΔG of the bulk solution, and its high value stems from the significant concentrations of reactants in the reaction mixture.
The individual redox reaction (4H2 + CO2 ↔ CH4 + 2H2O), by contrast, is, under standard conditions, among the least exergonic (ΔEm of about 150 mV, N.B.: ΔGo–nFΔEm) free energy converting reactions exploited by life (Schoepp-Cothenet et al., 2013). That this reaction actually is close to equilibrium is nicely illustrated by the fact that the sequence of redox transitions operating in methanogens in the presence of H2 and CO2, but at low concentrations of CH4, is readily reversed in methanotrophic Archaea when CH4 concentrations become significant (Haroon et al., 2013).
More importantly, both the biological and the posited abiotic reduction of CO2 to methane (Martin and Russell, 2007) proceed through sequential and mutually independent electron donations, in the latter case to, for example, the reduced carbonaceous chondrites (the C1-stony meteorites). From the point of view of far-from-equilibrium thermodynamics, the absence of feedback loops necessary for inducing autocatalysis and hence self-amplification during the approach to equilibrium of a system that was not far from equilibrium to begin with, a priori, does not look favorable for generating a dissipative structure.
The proposed natural pH of alkaline hydrothermal vents (Russell et al., 1989) as well as the recently discovered (Thauer et al., 2008; Chowdhury et al., 2016) flavin-based electron bifurcation reaction, however, is quintessential to entropy-decreasing engines (Cottrell, 1979; Branscomb and Russell, 2013) and, therefore, adds some of the ingredients required for the emergence of self-organizing systems to the reaction scheme advocated by Lane and Martin (2012 and see e.g., Weiss et al., 2016).
Methane-oxidizing reactions, such as, for example, that which is pulled by nitrogen oxides and oxyanions, yield a completely different picture (Fig. 1). The drop in free energy pertaining to their individual redox reactions (ΔEm) exceeds 1 V (Schoepp-Cothenet et al., 2013). Furthermore, they are replete with feedback loops, both at the level of substrates (the nitrogen compounds) and of reducing equivalents (Fig. 1). Since subsequent one-electron oxidation states of methane feature increasingly reducing electrochemical potentials, these can feed back into the initial reactions of methane activation and of the reduction of nitrogen oxyanions. Obviously, the already mentioned entropy-lowering engines employing the pH gradient and of electron bifurcation are expected to also play crucial roles in the scenario of denitrifying methanotrophy (Nitschke and Russell, 2013).
From the already described observations, we conclude that a reaction producing methane as an exhaust does not only appear geochemically compromised but also comparatively poorly suited (although not excluded) for kick-starting life's emergence in the form of a dissipative structure. By contrast, the oxidation of the “fuel” methane by high potential electron acceptors, such as, for example, nitrogen oxides and oxyanions, fulfills several of the criteria laid out by far-from-equilibrium thermodynamics for the emergence of self-organizing systems. A major task for future research into the details of a putative emergence of life driven by denitrifying methanotrophic acetogenesis will be to work out the behavior of reactants and products under different ranges of initial conditions.
6. Résumé and Conclusions
As CO2 may be rapidly hydrogenated to formate, it seems plausible that this reduction was taken over by emerging life as one step in carbon fixation (Seewald et al., 2006; Lang et al., 2010, 2012; Nitschke and Russell, 2013). However, a rapid and total hydrogenation of CO2 to methane is slow and does not produce the necessary metastable intermediates (McCollom and Seewald, 2003). Most of the CH4 at Lost City and other alkaline springs is presumably distilled, or cracked, from organic molecules previously lodged in crystalline and sulfidic source rocks (Strauss, 1989; Proskurowski et al., 2008). This finding is a stumbling block to the acetyl-coA pathway being the first metabolic pathway as negotiated by the methanogens. We, and others, had hitherto considered these archaea to occupy the lowest branches of the evolutionary tree (Koonin and Martin, 2005; Russell et al., 2005; Martin and Russell, 2007; Wolfe, 2014; Shock and Boyd, 2015).
So the serpentinization reaction appears limited to the direct delivery of the fuels H2 and, perhaps, HCOO−/CO—reduced from CO2—to emergent life, whereas the provision of CH4 is through the processing of pre-existing carbonaceous components or by entrainment of the volatile residing, or produced in, the oceanic lithosphere (Watanabe et al., 1983; Pineau and Mathez, 1990; Seewald et al., 2006; Proskurowski et al., 2008; Lazar et al., 2012, 2015; Paukert et al., 2012; McCollom and Seewald, 2013; Lollar et al., 2014; Suda et al., 2014; McDermott et al., 2015; Seyfried et al., 2015; McCollom and Donaldson, 2016).
In detail, we conclude from our critical review:
1. The serpentinization reaction—sometimes alluded to as the “sister” of early metabolism—does not reduce carbon dioxide to methane, nor to methane thiol, at least not rapidly enough for it to be thought of as a precursor to a metabolic reaction (Fig. 3) (Reeves et al., 2014; McCollom and Donaldson, 2016). As we have tried to convey in this article, serpentinization likely was the “mother engine” of life—not its sister (Russell et al., 2013)! Although the serpentinization reaction reduces some of the interacting circulating aqueous fluid to hydrogen, perhaps over a period of hundreds to thousands of years, that reduction too is neither facile nor rapid (McCollom and Donaldson, 2016).
These experimental findings contradict expectations that biochemistry first emerged as parallels to serpentinization reactions, that is, directly from geochemistry (cf., Russell and Hall., 1997; Martin and Russell, 2007; Nitschke and Russell, 2009). Lang et al. (2010) also cast very considerable doubt on the view that acetate may have been an immediate product of serpentinization. These authors found acetate at Lost City to vary nonsystematically with hydrothermal-to-ocean fluid chemistry from 1 to 35 μmol/kg for which they tentatively assumed a microbial derivation. We note also that the calculations of Shibuya et al. (2016) put acetate in Hadean hydrothermal springs at ∼10−23 molal.
In contrast, reduction of CO2 to formate is rapid during serpentinization (timescale of minutes or tens of minutes according to White (2013) and Herschy et al. (2014)). Also, Lang et al. (2010) recorded ∼150 μmol/kg of formate in the end-member hydrothermal fluids issuing from Lost City (and see McCollom and Seewald, 2001, 2003; McDermott et al., 2015). The serpentinization reaction thus appears limited to the direct delivery of HCOO− (reduced from CO2) to emergent life. These results force a rethink of the idea that the classic acetyl coenzyme pathway was the harbinger of life (Russell and Hall, 1997; Russell and Martin, 2004). In this regard, we note that Windman et al. (2007) suggested formate as an alternative source of fuel and reduced carbon, in place of H2 and CO2.
2. In our view, a modified (oxidative) version of the acetyl-coA pathway survives as the first pathway to life, although requiring nitrate and/or nitrite in early oceans as an electron acceptor to oxidize iron in GR as well as hydrothermal methane (Fig. 1). This hypothesized earliest metabolism has been termed “denitrifying methanotrophic acetogenesis” (Nitschke and Russell, 2013; Stern et al., 2015; Wong et al., 2017a). Although there has been some tenuous experimental support for this hypothesis (Russell et al. 2014 and references therein), the conclusion is again forced that an inorganic membrane is required to provide the vectorial redox and pH gradients (chemiosmotic disequilibrium) to drive life's emergence (Russell et al., 1989, 1994) in line with the works of Herschy et al. (2014) and Sojo et al. (2015) (and see Batista and Steinbock, 2015 for future experimental approaches).
In particular, Herschy et al. (2014) demonstrated that a proton gradient acting across thin nickel-doped mackinawite ([Fe>>Ni]S) membranes separating hot (70°C) alkaline (pH 11) solution from an ancient ocean simulant ∼pH 5 and ∼20°C appears also to drive the reduction of CO2 to formate (∼50 μmol/L), although not to acetate. We conclude that dispositive experimentation must demonstrate that nitric oxide, produced from nitrite/nitrate and activated in the interlayers of an Fe2+-rich GR, is called for as oxidant before the DMAP hypothesis could be acceptable (McGlynn et al., 2009; Nitschke and Russell, 2013). To carry out such tests, we envision juxtaposing a cool carbonate/nitrate/nitrite-bearing early acidulous ocean simulant, across the precipitate membrane, to an alkaline solution of hydrogen and methane in a high-pressure hydrothermal reaction chamber.
3. Our general conclusion is that what drove life's emergence was not merely speeding up of chemistry or geochemistry (toward biochemistry) as sometimes assumed. Key to the emergence and the maintenance of all life are specific enzymes, many of which are effectively disequilibria-converting engines (Branscomb and Russell, 2013; Branscomb et al., 2017). These are turnstile-like engines, often housed in membranes that, for example, couple strong redox and pH gradients to drive endergonic reactions (Branscomb and Russell, 2013; Branscomb et al., 2017). Reaction–diffusion systems are another mechanism for driving disequilibria conversions at the submarine alkaline vent (Epstein and Xu, 2016).
Comprising portions of the membrane and the exhalative pile, GR offers a chemically active, and confining, matrix that is not that different from the construct of a cell in which the several potential reactants could compete between diffusion distances and reaction rates, resulting in the self-organization of products as indicated in Figure 2 (Russell et al., 2003; Russell and Beckett, 2017; and see Johnson et al., 2015; Forticaux et al., 2015; Johannessen et al., 2016). A vital requirement for emergence-of-life research is to discover and demonstrate the power of minerals to act as such drivers and facilitators through flexuring, chemical waves, and changes of conformation, effected through redox, acid–base, and hydrolysis reactions (Arrhenius, 2003; Coveney et al., 2012; Hoffmann, 2012; Russell et al., 2013; Branscomb et al., 2016; Epstein and Xu, 2016).
4. With the proviso that “(E)xothermic serpentinization of ocean crust is life's mother engine” (Russell et al., 2013), we entertain the view that, far from being one of the first waste products of metabolism, abiotic methane was likely a fuel of methanotrophy (Evans et al., 2015 and supplementary information). On reflection, it is certainly more logical to see methane as a reductant, one, along with hydrogen—for it to be partially oxidized—thereby providing another portion of the carbon to the first biotic organic molecules (Ducluzeau et al., 2009, 2014; Nitschke and Russell, 2013).
5. An astrobiological implication: That the wet and rocky bodies in the solar system smaller than Earth probably have little or no bridgmanite (perovskite) and, therefore, have more reduced mantles may explain the preponderance of methane on some of the moons of Saturn (Wood et al., 2006; Bouquet et al., 2015; Dorofeeva, 2016; Girard et al., 2016; Gu et al., 2016), possibly Europa (Zolotov and Kargel, 2009) and even early Mars (Wadhwa, 2001; Edwards and Ehlmann, 2015; Hu et al., 2015; Wong et al., 2017b). The challenge then is to consider the availability of electron acceptors on these other worlds (e.g., Nealson, 1997; Russell et al., 2017).
Footnotes
Acknowledgments
We dedicate this contribution to Isik Kanik for his untiring and enthusiastic backing of our work in this area. We also thank Elbert Branscomb, Grayson Chadwick, Laurie Barge, Stuart Bartlett, Christophe Sotin, Steve Vance, and the members of the Keck Institute for Space Studies workshop, Methane on Mars, organized by Yuk Yung, Pin Chen, and Ken Nealson, for discussions. M.J.R.'s research was supported by the National Aeronautics and Space Administration, through the NASA Astrobiology Institute under cooperative agreement issued through the Science Mission directorate; No. NNH13ZDA017C (Icy Worlds) at the Jet Propulsion Laboratory. Copyright 2016. All rights reserved.
Author Disclosure Statement
No competing financial interests exist.