
Abstract
The BIOMEX (BIOlogy and Mars EXperiment) is part of the European Space Agency (ESA) space mission EXPOSE-R2 in low Earth orbit, devoted to exposing microorganisms for 1.5 years to space and simulated Mars conditions on the International Space Station. In preparing this mission, dried colonies of the Antarctic cryptoendolithic black fungus Cryomyces antarcticus CCFEE 515, grown on martian and lunar analog regolith pellets, were subjected to several ground-based preflight tests, Experiment Verification Tests, and Science Verification Tests (SVTs) that were performed to verify (i) the resistance of our model organism to space stressors when grown on extraterrestrial rock analogs and (ii) the possibility of detecting biomolecules as potential biosignatures. Here, the results of the SVTs, the last set of experiments, which were performed in ultraviolet radiation combined with simulated space vacuum or simulated martian conditions, are reported. The results demonstrate that C. antarcticus was able to tolerate the conditions of the SVT experiment, regardless of the substratum in which it was grown. DNA maintained high integrity after treatments and was confirmed as a possible biosignature; melanin, which was chosen to be a target for biosignature detection, was unambiguously detected by Raman spectroscopy.
1. Introduction
One of the main scientific challenges of our time is to discern whether life existed, or still exists, on Mars or on other planets or satellites of the Solar System. The related question of “habitability” is another challenge to investigate (Horneck et al., 2016). As reviewed by Cockell et al. (2016), the investigation of the origin of life on Earth, its persistence on the planet since it appeared, and the search for evidence of life on other planets aid in our understanding of those conditions that allow life to occur.
Astrobiologists strive to understand the limits of habitability and define requirements for metabolic activity and reproduction in planetary environments (Cockell et al., 2016). A second main focus in astrobiology is the definition of significantly distinguishable biosignatures; that is, molecules that indicate the presence of putative extinct or even extant life (de Vera et al., 2012; Horneck et al., 2016). Melanins are promising biosignature candidates; they are distributed among all the kingdoms of life, even if synthesized through different pathways, suggesting an early emergence in the course of evolution. Melanins confer on organisms a number of useful characteristics that allow them to adapt to extreme environments. In addition, melanized fungi are highly resistant to heat, cold, extreme pH, ultraviolet (UV) radiation, ionizing radiation, heavy metals, salt, and dryness (Gadd and de Rome 1988; Gunde-Cimerman et al., 2000; Dadachova and Casadevall, 2008; Gorbushina et al., 2008; Onofri et al., 2008; Selbmann et al., 2008, 2011; Sterflinger et al., 2012).
Melanins also possess unique physical, chemical, paramagnetic, and semiconductive properties (Meredith and Sarna, 2006) that act as possible energy harvesting pigments (Dadachova et al., 2007; Gessler et al., 2014). This double role of protection and energy transduction may have been crucial for microbes in the early history of life on Earth when radiation was higher than today.
Extreme environments on Earth, which have previously been thought to be incompatible with active life, are perfect models for studying the limits of life and habitability. Among them, the McMurdo Dry Valleys in Antarctica are the coldest hyperarid desert environments on Earth (Cowan et al., 2014) and represent a terrestrial analog of Mars. In this location, where the environmental conditions limit the colonization of rocky surfaces, endolithic life and cryptoendolithic life in particular are the predominant life form (Friedmann, 1982), representing the last chance for life before extinction (Friedmann and Weed, 1987). Recently, the geographical limits for endolithic life in Victoria Land were defined (Zucconi et al., 2016). Endolithic communities are of astrobiological significance because putative life on Mars would have adopted an endolithic habitus before its potential extinction, when conditions on Mars became harsher and harsher (Friedmann, 1986).
Our astrobiological model organism, the cryptoendolithic black fungus Cryomyces antarcticus CCFEE 515, from the McMurdo Dry Valleys, is able to resist extreme and long-term desiccation, a wide temperature fluctuation, and high doses of radiation (Onofri et al., 2004, 2008, 2012, 2015; Selbmann et al., 2011).
For all these reasons, it is an excellent eukaryotic model for astrobiological space research and was selected for the Lichens and Fungi Experiment (LIFE) experiment where it survived outside the International Space Station (ISS) under dried conditions and exposed to space for 18 months (Onofri et al., 2012). In the frame of the same experiment, it was also exposed to simulated martian conditions (95% carbon dioxide, UV >200 nm) and cosmic radiation. The LIFE experiment results show that C. antarcticus maintained good viability, with minimal DNA damage (Onofri et al., 2015).
However, some open questions remain in the wake of the LIFE experiment, and therefore the BIOlogy and Mars EXperiment (BIOMEX) project aims to aid our understanding of the following: (i) the survival of microorganisms grown on martian and lunar rock analogs in space and Mars conditions simulated in space; (ii) stability and degradation levels of space-exposed pigments, secondary metabolites, and cell surfaces in contact with terrestrial, lunar, or martian analog mineral environments, with the intent to possibly identify space-resistant biosignatures with regard to the degree of degradation of the organisms and their secondary metabolites due to a variety of space parameters (radiation, vacuum, and Mars-CO2-gas). C. antarcticus was one of the microorganisms selected for the BIOMEX experiment, and this organism was also exposed outside the ISS on the EXPOSE-R2 facility for 1.3 years (introduction article de Vera et al., 2019).
BIOMEX aims to test the resistance and stability of biomolecules, as well as the endurance of selected extremophiles under space and Mars-like conditions (de Vera et al., 2012). In particular, the experiment aims to define a list of reliable biosignatures (biomolecules including pigments) and build up a biosignature database. To mimic real planetary conditions, microorganisms were grown on lunar regolith analog rocks such as anorthosite (Mytrokhyn et al., 2003) and on two Mars regolith analog mixtures, namely phyllosilicatic Mars regolith simulant (P-MRS, Early Mars) and sulfatic Mars regolith simulant (S-MRS, Late Mars); the latter reflect two separate evolutionary epochs associated with environmental changes on Mars (Böttger et al., 2012).
In preparation for the mission, several preflight tests were performed to test and select the most promising organisms for this astrobiological experiment, and to optimize the sample preparation and improve the hardware integration procedures, two sets of tests (Experiment Verification Tests [EVTs] and Science Verification Tests [SVTs]) were used to assess the resistance of selected organism to the abiotic stressors experienced under space and Mars-like conditions.
In the EVTs, simulated space stressors (e.g., UV radiation, vacuum, and temperature variation) were applied separately to the microorganisms to verify the effects of each single condition. Differently, all stressors were applied at the same time during SVTs and on samples integrated in the same hardware used in the space experiment.
The results of the EVTs, which simulated the martian atmosphere, extreme temperature, vacuum, and UV radiation, show a high survival rate of C. antarcticus, as well as the preservation of the fungal DNA (Pacelli et al., 2017c).
In this study, we report and discuss the results of the last set of experiments, the SVTs, a dress rehearsal of the mission as planned, providing a combination of UV radiation and simulated space vacuum, and simulated martian conditions. The postexposure viability was tested by both cultural and molecular methods as previously and successfully applied on samples of the EVTs (Pacelli et al., 2017c). Ultrastructural damage was investigated by transmission electron microscopy (TEM), and metabolic activity was evaluated by the use of 2,3-bis(2-methoxy-4-nitro-5-sulfophenyl)-5-[(phenylamino) carbonyl]-2H-tetrazolium hydroxide (XTT assay) (Kuhn et al., 2003). After exposure to the SVT conditions, the detectability of melanin, our target biosignature molecule, was checked by Raman spectroscopy.
2. Materials and Methods
2.1. Organisms and cultivation conditions
Cryomyces antarcticus CCFEE 515 was isolated by R. Ocampo-Friedmann from sandstone collected at Linnaeus Terrace (Southern Victoria Land) by H. Vishniac in the Antarctic expedition 1980–1981.
To best mimic lunar and martian rocks, substrata were prepared as follows: malt extract agar (MEA) was mixed in Petri dishes with Antarctic sandstone, which is the original substrate (OS), and with lunar analogs (L), which mainly consist of anorthosite. To mimic two distinct martian soil compositions that represent soil from two different martian epochs, colonies were grown on the P-MRS (Early Mars) and S-MRS (Late Mars). The mineral compositions are reported in Table 1. Fungal colonies were grown for 3 months at 15°C on these substrata, obtaining superficial colonies that were well exposed outside the substratum; they were cut to fit within the wells of exposure carrier (12 mm diameter) (Fig. 1b), and then dehydrated under sterile conditions. All samples were prepared, such that they maintained their integrity in the exposure facility EXPOSE-R2 according to the optimized protocol reported by Pacelli et al. (2017c).

Mineralogical Composition of Sulfatic and Phyllosilicatic Mars Regolith Simulants and Lunar Analog Anorthosite (Lunar) in Weight/Volume % (Modified from Böttger et al., 2012)
P-MRS = phyllosilicatic Mars regolith stimulant; S-MRS = sulfatic Mars regolith simulant.
2.2. Ground-based simulations
The SVTs were performed at the Planetary and Space Simulation facilities (PSI) at the Institute of Aerospace Medicine (German Aerospace Center, DLR, Köln, Germany). SVTs are designed to ensure that all samples are appropriately prepared to successfully withstand hardware integration, conditions experienced during the mission, and postflight deintegration. The application of mission-equivalent space parameters allows for testing the resilience of samples toward the extreme environmental conditions of space and simulated Mars exposure.
The EXPOSE-R2 hardware and the scheme of sample accommodation are reported in Figure 1a–c. Samples were accommodated in wells with a diameter of 12 mm, within square aluminum alloy carriers, 76 mm side. Following the accommodation plan scheduled for the EXPOSE platform, SVTs allowed only one replicate per sample (de Vera et al., 2012).
To simulate space-like test conditions, the samples that were grown on OS and lunar analogs were exposed to vacuum (10−5 Pa) and cycling temperatures between −25°C (16 h in the dark) and +10°C (8 h during irradiation), in combination (space irradiated samples) with polychromatic UV (200–400 nm) radiation produced by the solar simulator SOL2000. The dose of 570 MJ/m2 was reached by running the SOL2000 for 99 h at 1370 W/m2. In parallel, Mars test parameters were simulated by low temperature (−25°C), Mars atmosphere (95.55% CO2, 2.70% N2, 1.60% Ar, 0.15% O2, ∼370 ppm H2O; Praxair Deutschland GmbH), and Mars-like pressure of 103 Pa for S-MRS and P-MRS analogs, in combination (Mars irradiated samples) with the same radiation, as described above. Neutral density filters (0.1%) were used to attenuate radiation in all tests performed; all conditions were simulated for a period of 28 days.
The applied fluency corresponds to the long-term space of 1 year of exposure outside the ISS, as estimated from previous EXPOSE data and simulations (Rabbow et al., 2012, 2015). It should be noted that space parameters cannot be fully mimicked in the laboratory (space vacuum and complex radiation environment). For example, deep UV, that is, solar UV radiation <200 nm, cannot be simulated over a lengthy period of time in the laboratory, which is why the SOL2000 delivers UV >200.
Below the irradiated samples, an identical set of samples (space dark samples and Mars dark samples) was kept in the dark and experienced all simulation parameters except UV radiation exposure. Controls (CTR) were kept at DLR in the dark at room temperature. The exposure conditions are summarized in Table 2.
Exposure Conditions During the Scientific Verification Tests
UV = ultraviolet.
3. Survival Assessment
3.1. Cultivation test
After rehydration for 3 days in 1 mL of physiological solution (NaCl 0.9%), the treated colonies were diluted to a final concentration of 50,000 cells/mL, and 0.1 mL of suspension was spread on Petri dishes supplemented with MEA (five replicates). Plates were incubated at 15°C for 3 months, and colony forming units were counted.
3.2. Propidium monoazide assay
This test was performed by adding propidium monoazide (PMA; Biotium, Hayward, CA) at a final concentration of 200 μM to rehydrated fungal colonies. PMA will penetrate only damaged membrane cells, crosslink to DNA after light exposure, and thereby prevent polymerase chain reaction (PCR). DNA extraction, purification, and quantitative PCR (qPCR), which were used to quantify the number of fungal Internal Transcribed Spacer (ITS) ribosomal DNA fragments present in both PMA-treated and nontreated samples, were performed according to the work of Onofri et al. (2012). Before qPCR, DNA was quantified by using the QUBIT system and diluted at the final concentration of 2 ng. All tests were performed in triplicate. qPCR-PMA results were obtained by comparing the intensities with a standard curve prepared with different concentrations of the target gene (range 102–108 copies; statistical significance: R 2 = 0.99).
3.3. Determination of metabolic activity by XTT assay
Colorimetric assay of cellular viability, namely XTT assay, was performed according to the protocol in the work of Kuhn et al. (2003); the XTT is converted to a colored formazan in the presence of cell metabolic activity. After SVT treatments, colonies of C. antarcticus were rehydrated in 1 mL of Malt extract (30 g/L). After 10 days, fungal cells were washed, suspended in phosphate-buffered saline, and placed into 96-well plates, three wells for each condition. The XTT assay was performed by adding 54 μL XTT (10 mg/mL)/menadione (2 mM) to each well. Plates were covered with foil and incubated, in agitation, at room temperature (24°C). Formazan product in the supernatant was detected by measuring the optical density at 492 nm (Labsystem Multiskan, Franklin, MA) after 2, 3, 4, and 12 h of incubation.
3.4. Statistical analyses
Means and standard deviations were calculated. Values obtained for each analogue (OS, L, S-MRS, and P-MRS) were normalized to each control, respectively. Statistical analyses were performed by one-way ANOVA, and pairwise multiple comparison procedure (Tukey test) was carried out with the statistical software SigmaStat 2.0 (Jandel).
4. DNA Integrity Revealed by PCR-Based Assays
The integrity of DNA template was assessed by amplifying different DNA gene lengths extracted from fungal colonies after treatments. DNA was extracted from colonies with the Nucleospin Plant kit (Macherey-Nagel, Düren, Germany) following the protocol optimized for fungi (Onofri et al., 2012). Before amplification, DNA was quantified with a QUBIT system and diluted at the same concentration (2 ng/μL).
ITS and large subunit-coding sequences (LSU) amplification were performed by using BioMix (BioLine GmbH, Luckenwalde, Germany), and adding 5 pmol/ng of each primer and 20 ng of template DNA at a final volume of 25 μL. Random amplification of polymorphic DNA (RAPD) was performed by using BioMix, and adding 5 pmol of the primer and 1 ng of template DNA at a final volume of 25 μL. The primer used for RAPD was GGA7 (GGA GGA GGA GGA GGA GGA GGA) (Kong et al., 2000). The amplification was carried out by using MyCycler Thermal Cycler (Bio-Rad Laboratories GmbH, Munich, Germany) equipped with a heated lid.
ITS, LSU, and RAPD amplification conditions were those as reported in the work of Pacelli et al. (2017c). Band intensity was measured and compared by using ImageJ software (Schneider et al., 2012).
5. Cell Integrity Through TEM
After rehydration, control and irradiated colonies were prepared for TEM, according to the protocol of Pacelli et al. (2017c), and the image acquisition was performed at the Center for High Instruments, Electron Microscopy Section of University of Tuscia (Viterbo, Italy).
6. Raman Confocal Spectroscopy
Raman measurements were performed in the Astrobiology Raman Lab at the DLR Institute of Planetary Research (Berlin) with a confocal Raman microscope (Witec alpha300 R system,
7. Results
7.1. Survival: Cultivation and PMA tests
Germination ability was not significantly different for the controls of each analog (OS, L, S-MRS, and P-MRS) compared with data obtained on Malt Agar alone, suggesting that substrates did not exert any reliable toxic effect (data not shown). C. antarcticus retained significant colony-forming ability after SVT treatments (Fig. 2), showing a similar trend of survival in all the substrates tested.
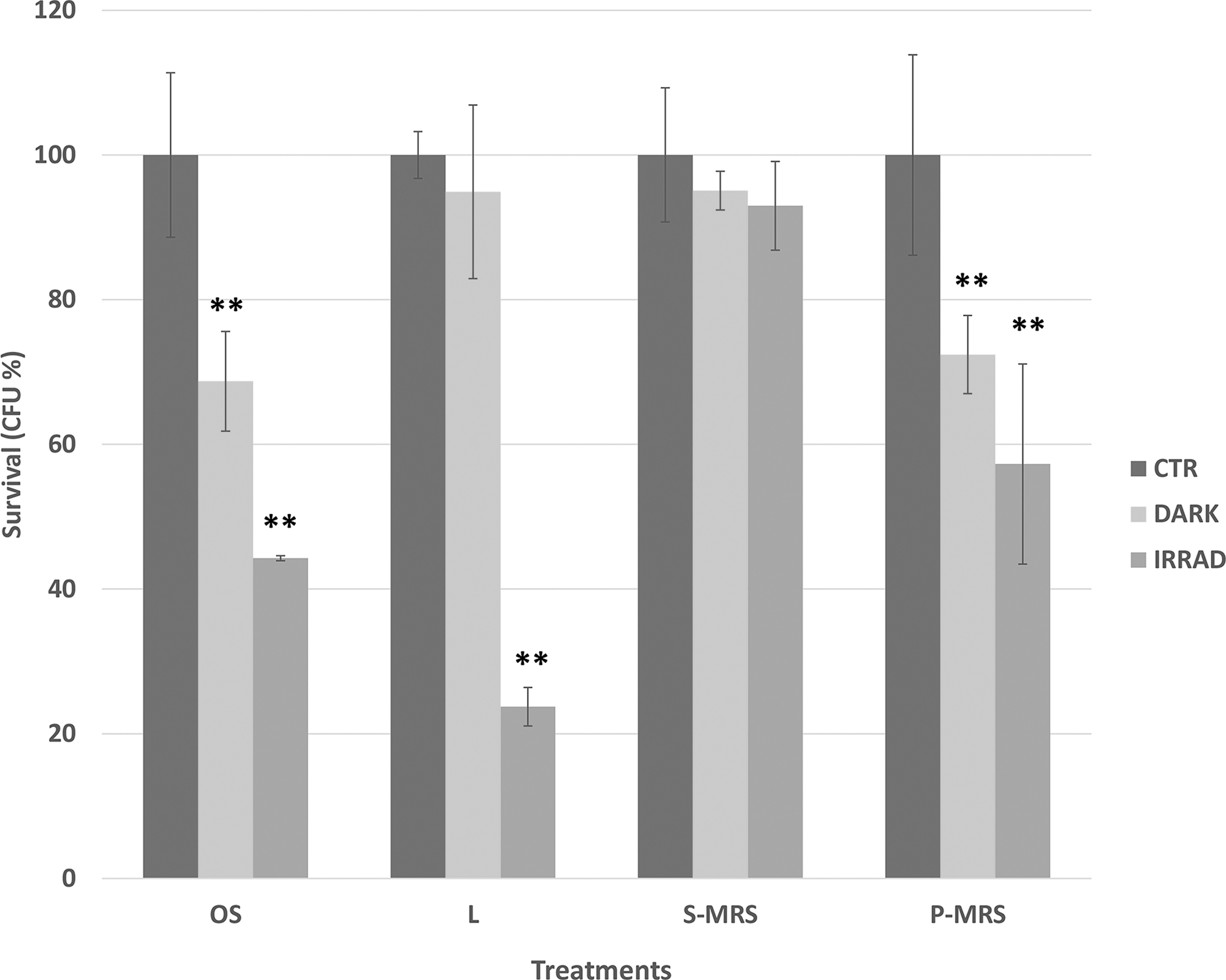
Cultivation test after SVT treatments: percentages of CFU of C. antarcticus on different substrates, relative to controls on the same substrate. OS, original substrate (sandstone); L, lunar analogs; S-MRS, sulfatic Mars regolith simulant; P-MRS, phyllosilicatic Mars regolith simulant. Significant differences were calculated by Tukey's test with **p < 0.001. CFU, colony-forming unit; SVT, Science Verification Test.
In general, radiation clearly represents an additional stress to the cell compared with that related to vacuum or Mars atmosphere conditions alone. For example, a decrease of 32% of the vitality was measured in OS dark samples compared with OS laboratory control, and an additional decrease of 22% was recorded for irradiated samples. Similar trends were obtained for L (same conditions) and P-MRS (Mars atmosphere only and irradiated). Differently, for S-MRS samples no statistically significant differences were observed for Mars dark samples (martian atmosphere) or for irradiation treatments compared with the control. Yet, for all substrates, a considerable capacity of survival was still maintained; 44%, 23%, 88%, and 57% of survivors were recorded for irradiated colonies grown on OS, L, S-MRS, and P-MRS, respectively.
Viability, specifically the integrity of the plasma membrane, was tested through PMA treatment followed by qPCR (Fig. 3). The PMA test also indicated that irradiation treatments, including vacuum plus radiation (for OS and L) and Mars atmosphere plus radiation (for S-MRS and P-MRS), were the most deleterious for fungus, especially on OS and L; beside, a higher percentage of viable cells (measured as cells with intact membrane) was observed compared with the results of the cultivation test. No significant differences between treated samples and control were obtained on S-MRS, corroborating the cultivation test.
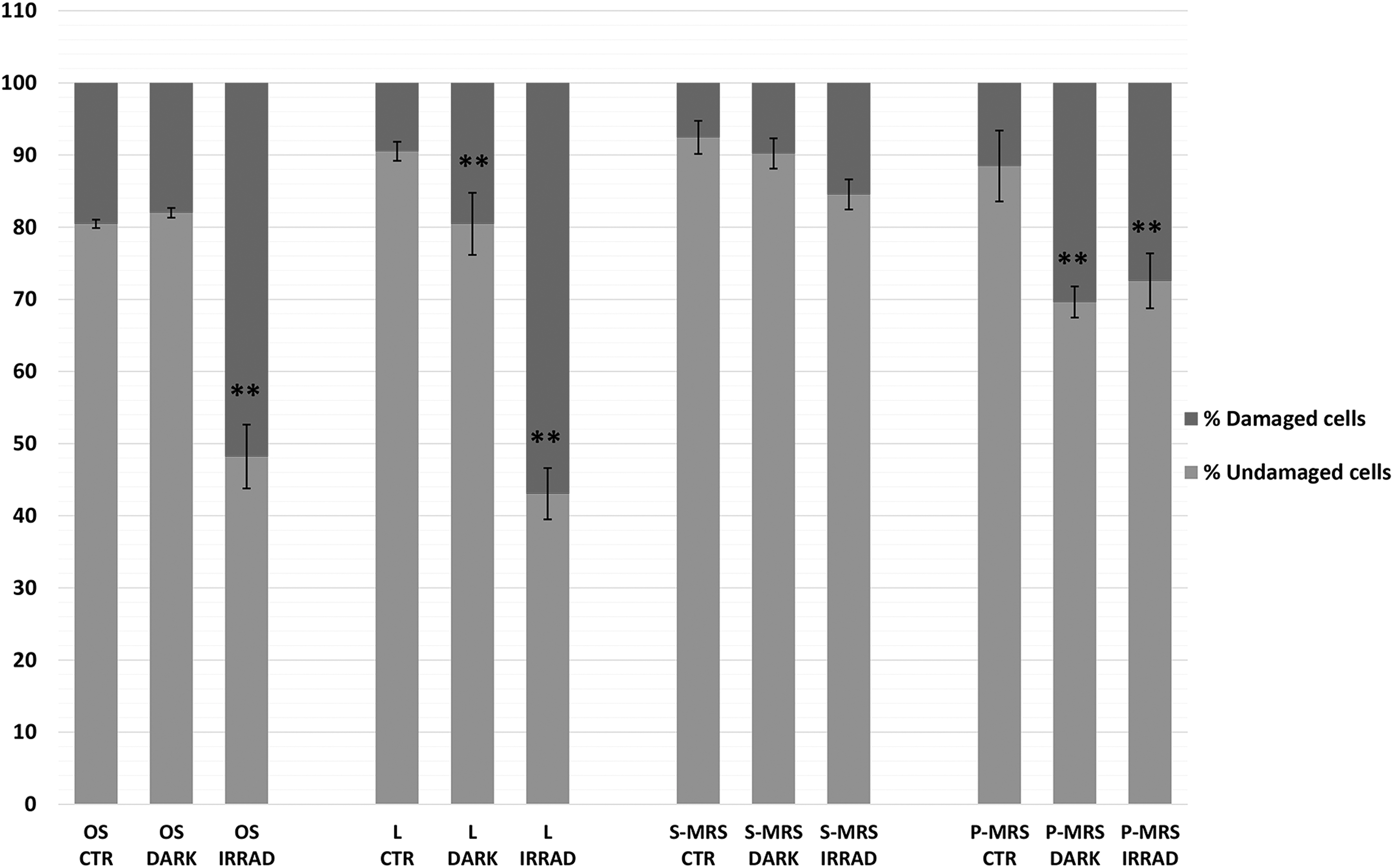
Results of PMA assay coupled with qPCR after SVT treatments in colonies grown on different substrates. Percentages of C. antarcticus cells with damaged membrane (dark gray), and corresponding percentages of cells (light gray) with intact membrane. Significant differences were calculated by Tukey's test with **p < 0.001 PMA, propidium monoazide; qPCR, quantitative polymerase chain reaction.
7.2. Metabolic activity analyses
For the XTT analysis, the most representative results were obtained after 4 h and are shown in Figure 4. The XTT activities decreased accordingly with the intensity of treatments from the dark samples to the irradiated samples for colonies grown on OS, lunar, and P-MRS; lunar irradiated samples showed again the highest decrease. The fungus grown on S-MRS retained the same metabolic activity (92%, no statistical difference from control) for dark samples and a slight decrease (80%) in the irradiated sample. All the results were again in general agreement with the cultivation tests.
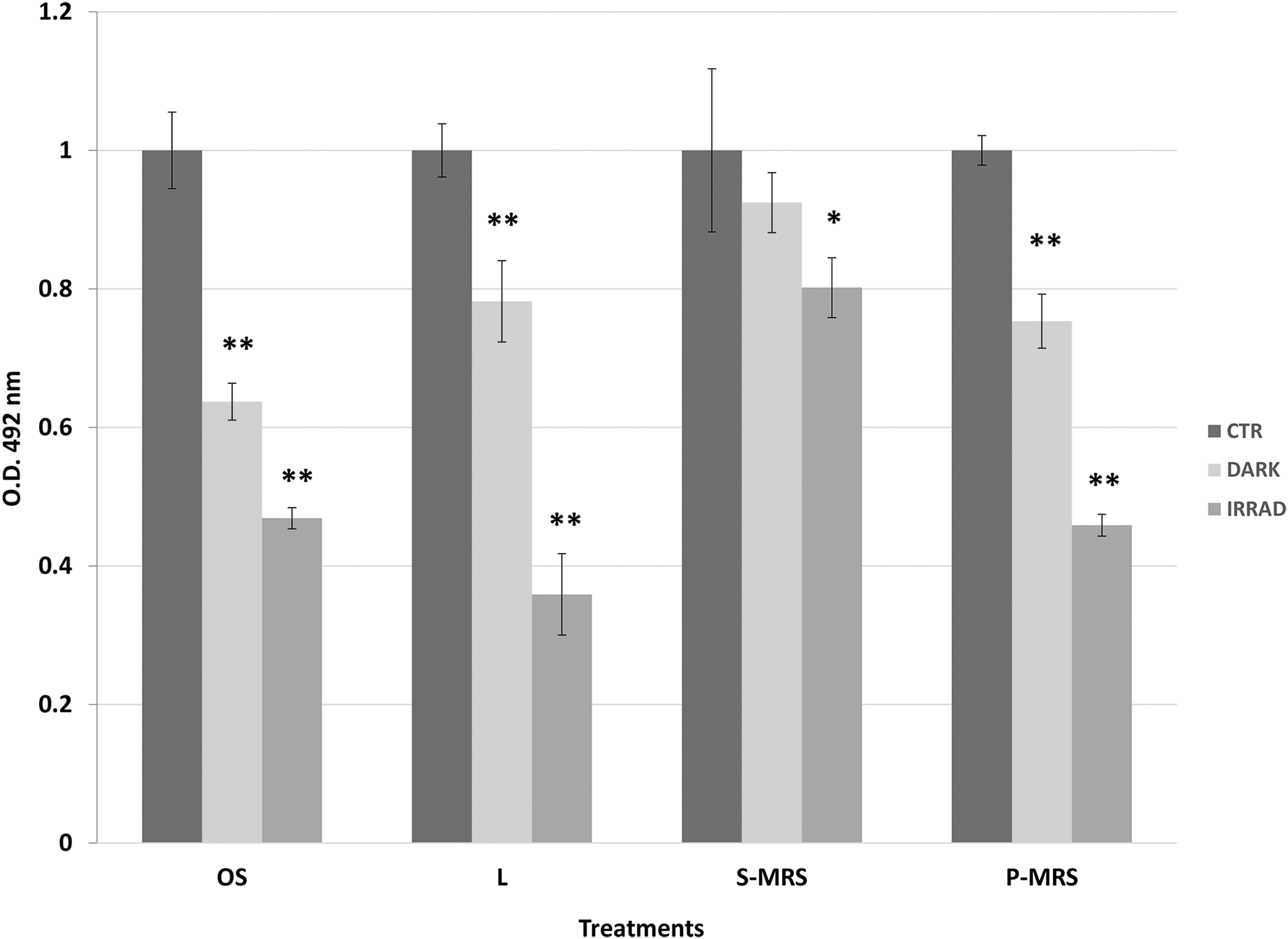
Effects of irradiation on metabolic activity of C. antarcticus cells by XTT assay after SVT treatments. OD 492 nm = optical density at 492 nm. Samples were normalized with the control, and significant differences were calculated by Tukey's test with *p < 0.05 and **p < 0.001.
7.3. DNA damage
The integrity of genomic DNA in treated samples was tested by assessing its ability to serve as a PCR template after SVT treatments. All the amplifications worked out for the ITS and LSU region (Fig. 5a–c). In particular, for the 700 bp gene length, a reduced intensity of PCR bands was recorded in UV irradiated samples, except for S-MRS (SM in Fig. 5a), where the gel analyses measured ∼85% relative density of the band for both irradiated and dark samples (Fig. 5a). For the 1600 bp gene length, the disappearance of the sole irradiated lunar sample was observed (Fig. 5b). For 2000 bp fragments, the band disappearance of lunar and P-MRS (PM in Fig. 5) irradiated samples was recorded, and 3% of band intensity was obtained for OS irradiated sample; beside ∼85% of intensity was still maintained in irradiated and dark S-MRS samples (Fig. 5c). The RAPD profiles were preserved in all the conditions tested (Fig. 5d). A reduction of intensity was measured for L and P-MRS irradiated samples only, demonstrating an overall good preservation of the whole fungal genomic DNA.
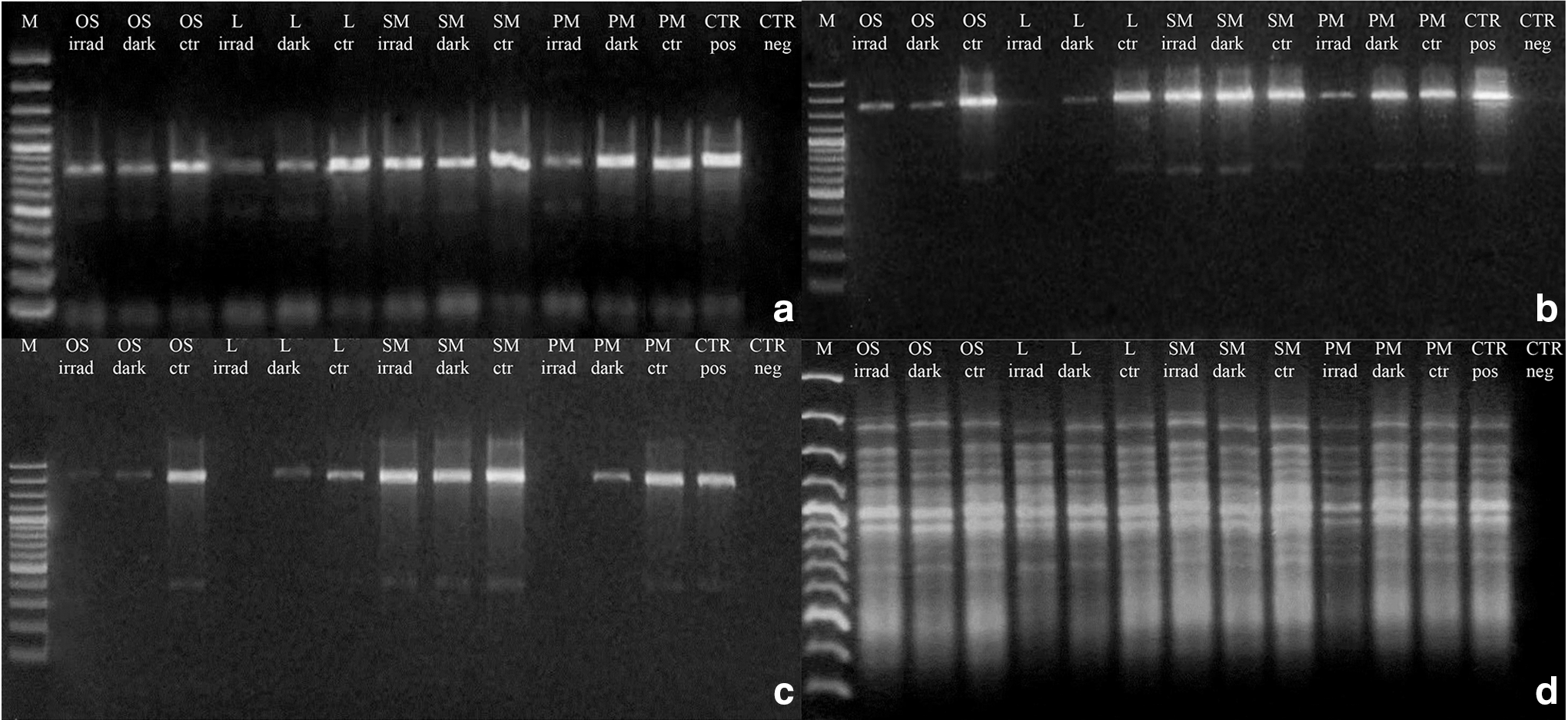
Assessment of the DNA damage on C. antarcticus after SVT treatments; single-gene PCR
7.4. Ultrastructural damage
TEM observations were performed on irradiated samples, where the effects on vitality and DNA damage were the most pronounced compared with the laboratory controls (Fig. 6). The ultrastructural damage in the cells of C. antarcticus was visible in all the irradiated samples, mostly in terms of loss of coherence and organization of the cytoplasm and cell membrane integrity. This was particularly evident in OS and L samples where the organelles were not visible in some cases and cell membranes were discontinuous (Fig. 6b, d, arrows). The same damage was also observed in P-MRS and S-MRS samples but to a lesser extent in terms of the number of cells involved, particularly for the latter (Fig. 6f, h, arrows). These results are again consistent with the data from previous tests where the colonies grown on S-MRS were less affected by the treatment.
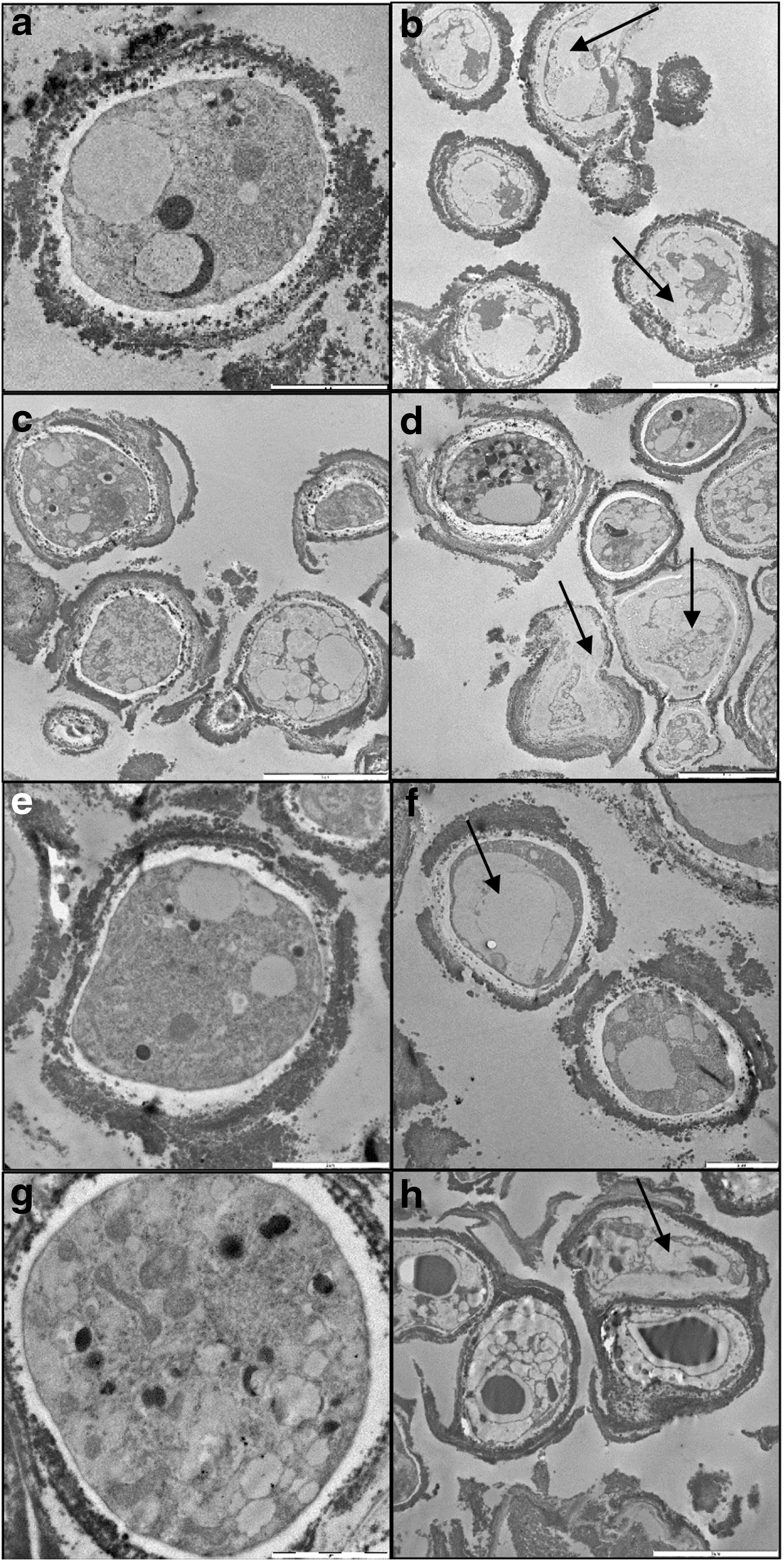
TEM micrographs. Laboratory control and irradiated microcolonies of C. antarcticus on original substrate
7.5. Raman spectroscopy
As for TEM, the Raman spectroscopy was performed on irradiated samples and laboratory controls only. Minerals from the analogs (results not shown) and melanins from the fungal colonies were well detectable in both control samples and irradiated samples (Fig. 7). In particular, the Raman spectrum of melanin in C. antarcticus was identified on irradiated samples on OS, S-MRS, and P-MRS substrates (Fig. 7a, c, d). The melanin-specific Raman peaks were 1349 and 1599 cm−1 (Fig. 7d). In contrast, the irradiated lunar samples gave a high fluorescence, so that any other peaks, including melanin, that might have been there, were undetectable (Fig. 7b).

Melanin Raman spectra (peaks 1349 and 1599 cm−1) obtained from C. antarcticus grown on
8. Discussion
The study of microbial communities adapted to the harshest conditions on Earth, and the assessment of limits for life on our own planet, could help garner insights into the potential for other worlds to support life and determine whether life, as we know it, may exist elsewhere in the galaxy (Saffary et al., 2002). In this work, we investigated the ability of the extremophilic black fungus C. antarcticus to survive and resist stressing conditions by simulating the space and Mars-like environment experienced during the preflight SVTs of the BIOMEX experiment. To mimic real space conditions, fungal colonies were exposed to stressors expected over a year of exposure outside the ISS, as estimated by Rabbow et al. (2012), and grown on lunar and martian rock analogs.
The described experiments aimed to investigate fungal resistance with regard to survival, metabolic activity, DNA integrity, and ultrastructural damage. After exposure, fungal survival was assessed by testing both the colony-forming ability and qPCR coupled with PMA. The metabolic activity and the ultrastructural damage were analyzed by XTT assay and TEM, respectively, to corroborate the survival results. The preservation of biosignatures, namely DNA and melanins, was evaluated by using PCR-based assay and Raman spectroscopy.
The results reveal good viability after the SVT treatments. None of the conditions tested caused complete inhibition of growth or metabolic activity, with a higher viability under Mars analog conditions compared with space conditions. The highest resistance was obtained from colonies grown on S-MRS analogs, where we did not record any statistical difference between dark and irradiated samples compared with the laboratory controls. Previous ground-based experiments, namely EVTs in the frame of BIOMEX too, showed the highest survival of Buellia frigida and C. antarcticus when grown on P-MRS analogs (Meeßen et al., 2015; Pacelli et al., 2017c), whereas Baqué et al. (2013) and Johnson et al. (2011) reported a higher survival of the cyanobacterium Chroococcidiopsis sp. when mixed with S-MRS regolith.
Overall, these results are of utmost importance, indicating that none of the analogs seemed to interfere with the survival of the biological models we use to discern the limits of life and the habitability of other planets such as Mars.
TEM observations were performed after the treatment to gain insight into the potential damage to the ultrastructure. The ultrastructural analyses confirmed the survival results; even if some injured cells were undoubtedly present in the treated samples, intact cells were also observed and were more frequent in S-MRS, where the survival and metabolic activity were the highest.
These results are consistent with the analyses on DNA damage. Amplicons were obtained from almost all samples for the shortest length analyzed. The disappearance of DNA was recorded only for lunar and P-MRS irradiated samples, and still 3% of band intensity was obtained for OS irradiated; yet, the fingerprinting analyses were successful for all samples, showing a good preservation of the whole genome.
These findings are in accordance with previous analyses performed in EVT ground tests, in the frame of BIOMEX (Pacelli et al., 2017c), and further support the use of DNA, an unequivocal proof of life, as a biosignature. In fact, the DNA extraction was efficient even for colonies grown on regolith analogs with no interference in the PCRs. Recent discoveries of nucleic acids or their precursors within meteorites (Martins et al., 2008; Schmitt-Kopplin et al., 2010; Callahan et al., 2011; Cooper et al., 2011) and in interstellar space (Hollis et al., 2000) could steer the development of life toward these biomolecules. Thus, it makes sense to search for RNA- or DNA-based life within potential habitable zones and on Mars. New approaches to search for life in the universe for the ExoMars mission include PCR-based methods for targeting ribosomal RNA genes. DNA sequencing (Isenbarger et al., 2008; Carr et al., 2013) is also one possibility to be used for the search for life.
Previous studies of C. antarcticus have demonstrated this organism's high capacity to resist, when dehydrated, the extreme stresses of simulated space and exposure to martian conditions (Pacelli et al., 2017c), real space conditions (Onofri et al., 2012, 2015), and high doses of ionizing radiation up to 55.81 kGy and even α particle (Pacelli et al., 2017b, 2017d).
The key factor that allows microorganisms to survive extreme environmental conditions is anhydrobiosis (Crowe et al., 1992; Billi et al., 2000; Kranner et al., 2008); for melanized fungi, this protective function is undoubtedly supported by the abundant presence of melanin in the outer cell wall layers (Gorbushina, 2003). Radiation-resistant melanized fungi can even survive in contaminated nuclear reactors (Zhdanova et al., 2000). More recently, it was demonstrated that both melanized Cryptococcus neoformans and C. antarcticus strains showed higher survival after high acute radiation doses under physiological conditions, compared with the nonmelanized strains (Pacelli et al., 2017a). The high viability of the fungus in this study can be explained, in part, by the anhydrobiotic and resting state of the fungus and by the presence of melanin in cell walls. How long these organisms might retain their viability in the interplanetary environment remains a guess for now and a challenge for future research efforts; the analyses of real space experiments will hopefully offer new insights.
Raman spectroscopy was used to detect the presence and persistence of melanin, our target molecule as a potential biosignature, after SVTs exposure, given that a Raman spectroscope is among the miniaturized facilities planned as part of the Pasteur payload in the European Space Agency (ESA)-Roscosmos ExoMars mission (Vago et al., 2006). Anorthosite, the main component of lunar regolith (Mytrokhyn et al., 2003; Kozyrovska et al., 2006), and P-MRS and S-MRS analogs of minerals formed during the Noachian (Early Mars) and Hesperian/Amazonian (Late Mars), respectively (Böttger et al., 2012), were used to simulate real exposure of putative biosignatures to an extraterrestrial environment (de Vera et al., 2012).
A set of measurements along strict intervals of time and number of repetitions was performed to optimize the detection of minerals and biological markers and reduce the disturbing effect of cosmic rays (Böttger et al., 2012). Mineral spectra in the tested samples were obtained (results not shown). Melanins were dominant in the spectrum gained from fungal colonies. Melanin spectra were obtained for both control and irradiated samples of OS, P-MRS, and S-MRS; despite the fact that melanin peaks were similar to those of amorphous carbon (1352 and 1613 cm−1) (Böttger et al., 2012), it was possible to distinguish the presence of our target unambiguously, in both irradiated and control samples (Fig. 7). The Raman approach on lunar irradiated samples was affected by the presence of high fluorescence, covering all Raman peaks, including melanin.
The same problem with high fluorescence in lunar regoliths after irradiation was already observed in other experiments within BIOMEX (M. Baqué, personal communication). Moreover, it was already reported that radiation might lead to degradation of Raman biosignatures (Dartnell et al., 2012). The effect of fluorescence of minerals on Raman spectra points out the need for taking into account their geological and chemical characteristics when developing biosignature databases.
9. Conclusions
As a preparative phase of the BIOMEX experiment of the EXPOSE-R2 space mission onboard the ISS, dried cells of C. antarcticus CCFEE 515 were grown on lunar and martian regolith analogs and were exposed to simulated Mars and space conditions. Results demonstrate that C. antarcticus retains metabolic activity and is even capable of forming colonies, after SVT treatment on extraterrestrial regolith analogs without significant DNA or ultrastructural damage. These results are essential to substantiate and validate analyses from space-exposed samples, and to prepare the BIOMEX experiment and future space experiments.
The high resistance of this fungus justifies its use in a space experiment to test its responses in situ to the real conditions of outer space. It is expected that the fungus has the capability to cope with the complex space environment and simulated martian conditions. Melanin is proposed as a valid biosignature, given that the use of Raman spectroscopy with parameters foreseen for the ExoMars 2020 mission is adequate for the analysis of melanin before and after irradiation. DNA, due to its resistance in simulated space and Mars conditions, may be indicated as a possible good biosignature as well. Further investigation will be needed to improve the appropriate parameter set for Raman spectroscopic analysis of melanin. Future analyses with regard to the search for biosignatures in our samples will focus on gas chromatography mass spectrometers, as one such instrument will be included among the miniaturized facilities slated for the Pasteur payload of the ExoMars mission.
Footnotes
Acknowledgments
We thank the European Space Agency (ESA) for funding SVTs. We also thank the Italian Space Agency (ASI) for co-funding the research (BIOMEX MCF Experiment on ISS for tracking biosignatures on martian and lunar rock analogs and E-GEM GeoMicrobiology for Space Exploration grants), the Italian National Program of Antarctic Researches (PNRA) and Italian National Antarctic Museum “Felice Ippolito” (MNA) for funding the collection of Antarctic samples CCFEE. We thank Dr. Ruth A. Bryan, PhD, for the thorough English revision.
Author Disclosure Statement
No competing financial interests exist.