
Abstract
Small-scale terracing (microterracettes) is a surface geomorphic feature that recurs under a range of environmental settings, such as those existing in high to low temperature geothermal springs and evaporitic environments, through the single or combined action of physicochemical agents and microbiological processes. Such morphology can also be observed in a confined sector of the Sabkha Oum Dba, which is an inland sabkha of the Western Sahara (Morocco), where field and laboratory investigations revealed that they primarily depend on the accumulation of naviculoid diatoms. Through their biofilm production ability, these benthic diatoms are able to stabilize surface morphologies and make organic alveolar frameworks where the precipitation of low Mg calcite occurs in areas subjected to active oxygenic photosynthesis. Because microterracettes arise in a specific set of environmental conditions, they have environmental significance and, thanks to a high fossilization potential due to mineral precipitation, they can be an effective source of biomorphological and chemical evidence for life. The relationship with aqueous environments, considered to be widespread on Mars especially during a period of intense hydrologic activity as in the late Noachian and Hesperian periods, make the understanding of surficial processes useful (such as the formation of microterracettes) whose formation is frequent in terrestrial analogues for martian environments, such as ephemeral saline continental lakes (sabkhas) and related to the products of bacterial and eukaryotic life, as in the case of biofilms, in search for similar life forms beyond Earth.
1. Introduction
M
The contribution of microbes to the development of surface micromorphologies can also be highlighted by their ability to induce mineral precipitation. This creates a substantial increase in their preservation potential in the geological record (e.g., Noffke, 2010; Noffke et al., 2013a). The geomorphic role of microorganisms can, therefore, be interpreted as a direct expression of their vital activities across different spatiotemporal scales. Stromatolites represent the most prominent and investigated example of this microbe–sediment expression.
As a rapidly growing research field, microbial geomorphology is becoming a tool for understanding how the interactions between microbiota and their physical environments may develop an effective capacity to produce biosignatures across the entire range of the geological record on Earth and elsewhere (Seckbach and Walsh, 2008; Grosch and McLoughlin, 2015).
Among microorganisms active in shaping surface geomorphologies, those able to produce biofilms have a role of particular relevance because of the ability of a biofilm to attach firmly to a substrate and, as a consequence, to induce physical and chemical interactions with it (Meyer-Reil, 1994; Gorbushina and Krumbein, 2000; Paterson et al., 2003; Mujah et al., 2017). This ability—and its macroscopic expression: the formation of microbial mats—has been largely preserved through time (e.g., Noffke et al., 2013b). In close connection with the formation of surface morphologies (and their fossilization potential) is the cohesive nature of biofilms, which may bind together loose mineral or skeletal particles at the sediment surface (Neumann et al., 1970; Winsborough, 2000; Paterson et al., 2008; Grabowski et al., 2011).
On Earth, the combined action of microbial mats and the mineral precipitates that they induce atop and within microbiota and sediment enables the formation and stabilization of macro- and microscale diagnostic morphologies in habitats including coastal lakes, lagoons, and sabkhas. The astrobiological relevance of these habitats, which are believed good terrestrial analogues for martian environments, is primarily due to their relationship with water and the microbial communities of extremophiles, and to some preservation potential in the fossil record. During the present-day phase of the investigation of the martian surface, these morphologies might also reveal a useful comparative and diagnostic potential (for a sabkha/playa lake example, see Noffke, 2015).
Although life in a biofilm is a condition that primarily typifies prokaryotes—for example, many bacteria develop biofilms as a survival and development strategy (e.g., Costerton et al., 1995)—it also characterizes certain eukaryotes, such as algae and fungi. This is a life strategy implemented by benthic eukaryotic microorganisms, especially when they inhabit extremely harsh environments (e.g., Brake and Hasiotis, 2008) where they may directly secrete their own mucilage to produce a biofilm in which to be hosted.
Examples of particularly demanding environmental conditions are acid mine drainage, such as the well-known case of the Río Tinto (Aguilera et al., 2007) or radioactive contaminated environments (Zirnstein et al., 2012). In acid mine drainage systems, photosynthetic eukaryotes (e.g., diatoms) have been found as biofilm producers and, as a consequence, are actively involved in the formation of organosedimentary structures such as stromatolites (Brake et al., 2004; Brake and Hasiotis, 2010). Some meiofaunal eukaryotes, such as microgastropods and arthropods, may use the product of mat-forming microbiota as a source of food and for microenvironmental protection (e.g., Gerdes, 2003).
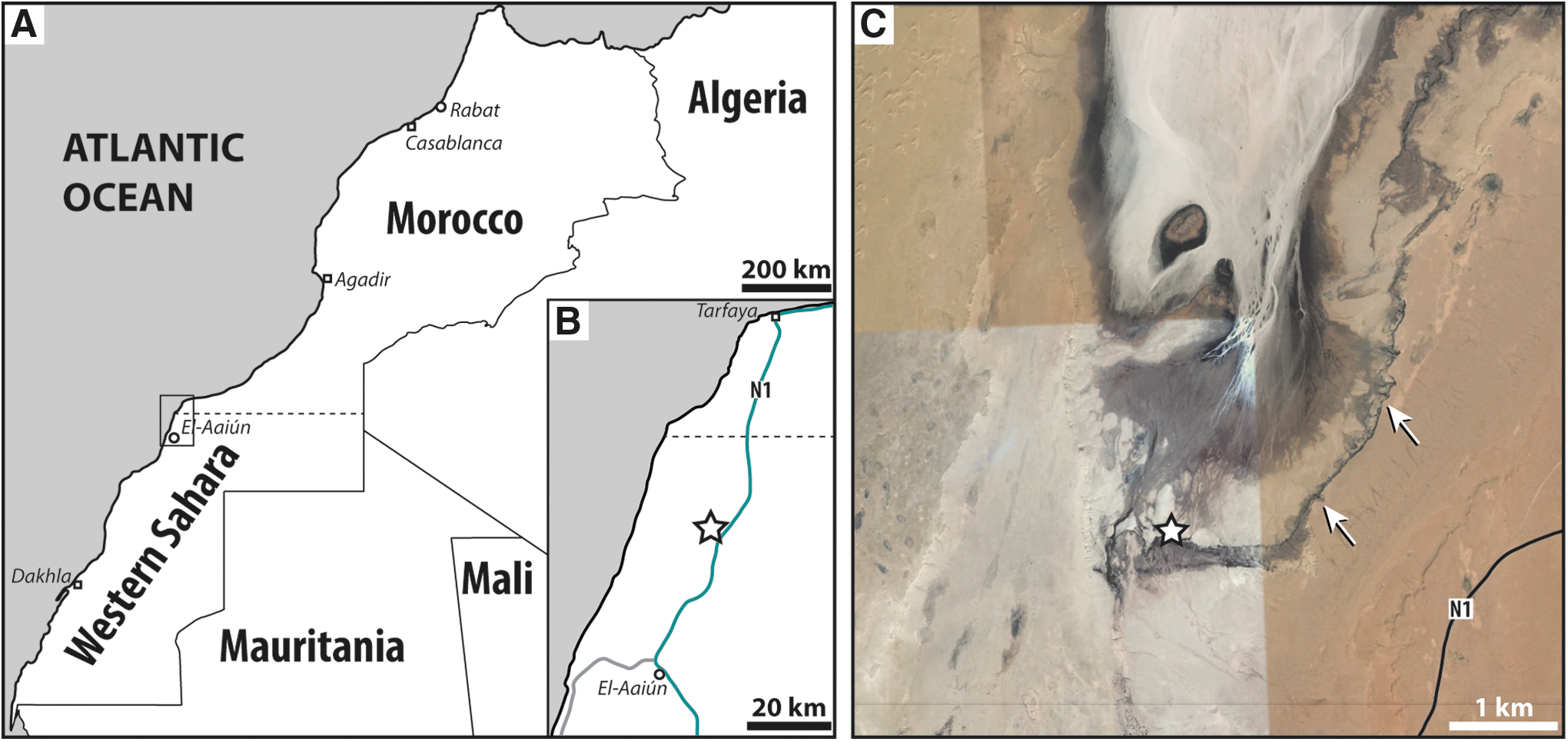
The case discussed in this article refers to the formation of microterracettes—a widespread surface small-scale morphology, the formation of which may also depend on the contribution of mat-forming microorganisms—that is spectacularly developed in a sector of the Sabkha Oum Dba in Western Sahara, Morocco (Fig. 1), an arid site affected by the continuous emission of moderately salty neutral/slightly alkaline groundwater at the ground surface (Table 1). The reason for this constant groundwater supply is unclear; a possible explanation might be the topographic level of the sabkha surface, that is lower than the surrounding region. In this confined sabkha environment, different microhabitats coexist, and the relationships between microorganisms and the physical environment can be directly observed in an easily accessible, active ecosystem.
Small-scale terracing occurs under different terrestrial aquatic settings, such as in high to low temperature geothermal springs associated with the formation of silica or carbonate deposits, and evaporite environments.
In this article, the term “microterracette” has been used after the classification into three size categories of Fouke et al. (2000) for terraced travertine at Yellowstone National Park, where they used “microterracettes” for the smaller size category. In general, their origin and development depend on a number of physical factors, including water salinity and temperature, and the contribution of the biotic component. Since they may represent loci for biotic–abiotic interactions and, accordingly, may be sites for hosting microbiological communities, microterracettes can also preserve bioinduced morphologies as well as other biomarkers. The latter possibility, and similarly to other morphologies arising from microbes–mineral interactions, allows for their potential use even in an astrobiological perspective (e.g., Röling et al., 2015).
Another reason justifying an astrobiological role for these morphologies from clastic–evaporite sabkha environments relies on the abundance of evaporite deposits (carbonates, sulfates, and chlorides) on Mars (Zheng et al., 2013). At least for some of these deposits, confined aquatic settings similar to those on Earth have been hypothesized (e.g., Metz et al., 2009; Grotzinger et al., 2015). A late Noachian and Hesperian age for these martian evaporites (Carr and Head, 2010), namely during a phase of extensive hydrological activity, is also notable.
2. Microterracettes in Natural Environments
Terracettes and microterracettes are recurring geomorphic landforms that are products of physical (Jettestuen et al., 2006) or biological agents, or the interaction of the two. In hot and cold water geothermal environments, a combination of water- and bioinduced processes can lead to their formation.
During the interaction between microbes and their surroundings, the cohesive action of microbial agents, possibly combined with (bio)mineral precipitation, may concur to stabilize this landform. Microterracettes, in particular, have been described from different geomicrobiological systems, forming under harsh conditions such as acidic mine drainage (Sánchez-Espanã et al., 2007) and a variety of geothermal areas, for example, Yellowstone National Park, USA (Hammer et al., 2010; Beam et al., 2016), El Tatio, Chile (Jones and Renaut, 1997; Nicolau et al., 2014), and Dallol Crater, Danakil Depression, Ethiopia (Waltham, 2010), with different scales and spacings.
Microterracettes also develop in low temperature, nearly neutral/slightly alkaline (this study) or alkaline springs (Brecon Beacons National Park [Wales], Ian Crawford, University of London, pers. comm.). They also have been detected in the fossil record, for example, in the Quaternary warm spring deposits of the Skoura region (Ouarzazate depression, Morocco). The mentioned examples of microterracette morphologies, however, differ in their mineral composition, being composed, depending on the sites, by iron (oxy)hydroxide, siliceous sinter, sodium chloride, and calcareous travertine (Fig. 2A–D). As for active geothermal sites, microterracette constructions have been detected in overflow areas with the persistent action of running water. Present-day examples include those observed at El Tatio and Yellowstone geyser fields (Fig. 2B, D), where microterracette surfaces (topped by lilypad-type stromatolites) are formed around domal mounds derived from continuous hot water emissions (Guidry and Chafetz, 2003; Nicolau et al., 2014). Similar geomorphic features characterize the travertine microterracing at Mammoth Hot Springs, Yellowstone (Fouke et al., 2000; Fouke, 2011), and fossil examples of travertines from different hydrothermal settings (e.g., Jorge-Villar et al., 2007; Fiorelli et al., 2012), including the Skoura region (Fig. 2A).

Examples of fossil and modern (active) microterracette morphologies.
The microterracette slopes at El Tatio and Yellowstone may vary from steep (nearly vertical) to a low-angle gradient. Their uppermost surfaces often exhibit small depressions bounded by minidam pattern (sensu Pentecost, 2005) and, therefore, able to retain some water so as to become a hospitable environment for living cyanobacteria, algae, and other microorganisms. Since both biological and physicochemical factors may play a role in the formation of microterracing landforms, processes of mineral precipitation can also contribute to enhancing the fossilization potential of associated organic remains. Because of their abundance in geothermal environments—at Yellowstone, for example, microterraced surfaces represent the most ubiquitous small-scale surface morphology (Pursell, 1985; fide Winsborough et al., 1994)—and the different microbial interactions that they may host, these landforms can be excellent repositories of organic biosignatures. In other instances of geothermal spring systems, such as at the hot spring of the Dallol Crater (Fig. 2C), centimeter-scale microterracettes appear to be solely the product of mineral (halite) precipitation in a very dynamic water flow system.
3. Study Area, Samplings, Methods
3.1. The Sabkha Oum Dba
The Sabkha Oum Dba is located 35 km north of El Aaiún, Western Sahara (Morocco), 20 km from the Atlantic Ocean, at about 5 km along a track west of the N1 national road (Fig. 1A–C). It is a confined and elongated area of ∼70 km2 (5 km wide × 15 km long). This inland sabkha overlies the Neogene-Quaternary succession of the Tarfaya Basin (El Jorfi et al., 2015).
Despite a relatively limited size, the sabkha hosts a number of different subenvironments related to the evaporation of saline waters (salt flats), eolian deposits (sand dunes), deflation areas, and ephemeral streams. Much of the surface of the sabkha, however, consists of flat areas with clay and sand deposits, which are often encrusted by thick halite crusts. Because of the steep margins that almost completely surround the Sabkha Oum Dba, especially on its eastern side (Fig. 1C), the easiest access road is from the south, and in the southernmost sector of the sabkha, at the entry point into the basin of a small stream, is located the study area (Figs. 1C and 3). This confined sector hosts a peculiar set of water- and salt-related subenvironments.

3.2. Physicochemical environment and sampling
Morphologically, the study area includes a vertical cliff, ∼8 m high × 36 m wide (Fig. 3), that partially delimits the southern margin of the sabkha depression from the surrounding desert region (Fig. 1C). Cold and relatively salty groundwater (salinity of 21,000–24,000 ppm total salts, Table 2) emerges about 500 m south of the cliff, and feeds a shallow stream to form a small waterfall at the cliff. Several shallow pools (10–30 cm deep), permanently fed by the water emission to form a stream, can be observed near the edge of the cliff. Shallow pools of standing water (up to 1 m deep) occur at the base of the cliff where they are surrounded by bushes of halophytes belonging to the genera Artrochnemum, Kochia, and Spartina (Fig. 3). Immediately beyond the waterfall, the stream flows and disperses a few hundred meters to the north within the sabkha, thereby leaving thin white salt deposits. During all of the field campaigns (in May and October 2012, and February 2014), groundwater was observed to supply the stream and its ecosystem.
Concentrations are in mg/L.
Altogether, the investigated field area at the base and top of the cliff includes the waterfall, water ponds, areas covered by water spray, and adjacent areas. All of these environments host a wealth of living/partially mineralized and completely mineralized microbial communities. They are embedded in millimeter- to centimeter-thick microbial mats with colors varying from light-dark brown to cream (Fig. 4A). Where the microbial mat is thicker, it comprises 3–4 cm of laminated partially mineralized mat (Fig. 4B). The living microbial mats with ongoing mineral precipitation (Fig. 4A–C), and then heavily encrusted and, lastly, mineralized (Fig. 4D–F), as well as other surface landforms induced by the interaction between microbes and physical factors, are set on a substrate consisting of eolian deposits (sands) cemented by calcium carbonate and calcium sulfate precipitation (Fig. 4B). Tomographic investigations in this sector of the Sabkha Oum Dba revealed that this type of eolian deposit reaches an overall thickness of at least 15–20 m (Sabbadini, 2014).

The microterracettes are the prevailing geomorphic landforms in two distinct areas (Fig. 3): (1) near the base of the cliff, an area labeled OD1 (coordinates: 27°27′69.6″N, 13°03′02.8″W), where rock surfaces are permanently wet by water falling from the waterfall; and (2) at the top eastern portion of the cliff, an area labeled OD3 (coordinates: 27°27′72.2″N, 13°03′06.5″W), where the rock surface permanently receives water spray from the waterfall. The OD1-microterracette morphologies occur in areas subjected to the action of water flowing in a shallow sheet, on top of thick and living/partially mineralized (mineral precipitation was active at the time of sampling) microbial mats (Figs. 4A and 5A). In contrast, the OD3-microterracettes were only observed where the available water (at least at the time of sampling) was limited to waterfall spray. Here the microterracette landforms were completely mineralized (Fig. 5B, C). The range of pH and temperature, repeatedly measured on waters during several days during the field survey and sampling, provided the following data: pH of 7.9–8.1 and 17–23°C were recorded during the day and a pH of 7.5–7.8 and 11–12°C at night (Table 1).

Samples for this study were collected in February 2014 and comprise (1) portions of living/partially mineralized (and then air dried) microbial mats from the OD1 area, (2) samples of mineralized surface micromorphologies (from the OD1 and OD3 areas), (3) rock samples from the sedimentary substrate (eolian sandstones), (4) water samples.
3.3. Methods
Field analysis included the identification of biosedimentological features and direct observation of the relationship between physical and biotic components in an active natural system. Electrical conductivity, pH, and temperature were measured in the field by using an XS Instruments pH 70 portable laboratory.
The samples of living/partially mineralized (air dried) microbial mats were investigated through freshly broken mat portions to observe both the surface and the millimeter- to micrometer-scale (mixed microbial/mineral) stratigraphy, by combining transmitted light and scanning electron microscopy (SEM and environmental scanning electron microscopy [ESEM]) analyses.
Samples of mineralized surface micromorphologies (microterracettes) and the eolian sandstone of the substrate were investigated through uncovered polished petrographic thin sections (7.5 × 4.5 cm2) and polished slabs under transmitted and reflected light microscopy by using a Wild M8 stereomicroscope and a Zeiss Axioplan binocular optical microscope equipped with a Nikon digital camera Head DS-Fi2. ESEM observations were performed on uncoated samples with an FEI Quanta 200, under low vacuum conditions, with an accelerating voltage of 20–25 kV, which followed the preliminary analyses carried out on coated samples with an SEM Jeol JSM-5400 instrument. The mineral and elemental compositions were determined with bulk X-ray diffraction (XRD) on two powdered samples from the OD1 area using a Philips PW 1480 X-ray diffractometer, whereas the elemental analyses were performed in the OD1 and OD3 areas with an OXFORD-SATW light elements X-ray spectroscope (SEM-EDX). For the water analyses, samples were collected in plastic bottles. In the laboratory, atomic absorption spectroscopy and ion chromatography (Thermo Scientific S Series and Metrohm 881 Compact IC pro instruments) were used for the determination of the major cations and anions (Table 2), respectively.
4. Microterracettes of the Sabkha Oum Dba: Field and Laboratory Results
4.1. Field observations
Microterracettes, either partially or completely mineralized, are common landforms in the cliff–waterfall area. At the base of the cliff (Fig. 5A), this surface morphology develops as arc-shaped ridges reaching a maximum height of a few millimeters on top of living/partially mineralized microbial mats, and extends over a substrate of cemented eolian sands. It developed at the transition between the water cascade itself and the surrounding areas were exposed to some water spray. The regime of the water flow and the gradient steepness are the physical factors that, combined with mineral and biological components, seem to determine the development of this landform. Microterracettes comprise a system of partially anastomosing, arc-shaped ridges delimiting microareas ranging from tens of square millimeters up to a few square centimeters (Fig. 5A–C). Although the height of the ridges ranges from <1 mm to several millimeters, in case of steeper gradients, it can also reach a development up to 1 cm (Fig. 5B, C). Depending on their size, ridges can allow a certain amount of water to be trapped between contiguous microterracettes. Under the action of laminar water flow, the ridges develop a relatively continuous morphology (Fig. 5B). In contrast, under a less orderly (turbulent) flow regime, and/or with steeper gradients, ridges become much less continuous (Fig. 5C). In case of extremely steep slopes, the microterracettes turn into a system of partially overlapping lobes oriented along the direction of a maximum gradient (Fig. 5D). In both OD1 and OD3 areas, microregions that are delimited by ridges are often ornamented by a kind of mineral network produced by small dense aggregates of Mg calcite precipitates (Fig. 5B). A similar microcrystalline precipitation was also observed on top of the living/partially mineralized microbial mats and this readily documents the effect of early mineralization stages (Fig. 6A–D).

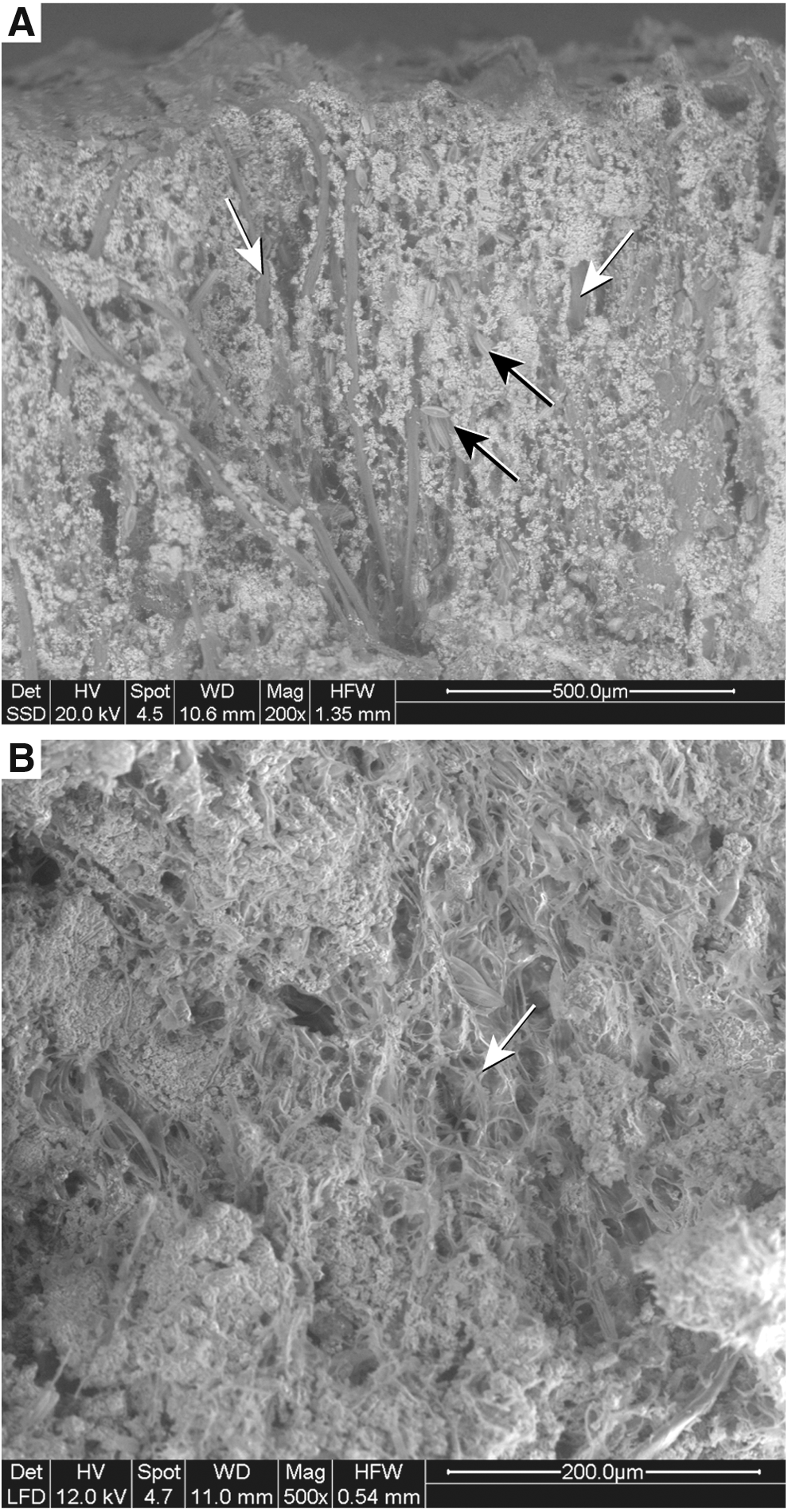
OD1 area: SEM micrographs on top of living (air dried)/partially mineralized microbial mats in a sector covered by the formation of microterracettes
4.2. Optical and scanning electron microscopy observations
Light and electron microscopy investigations of the samples collected from areas covered by partially and completely mineralized microterracette morphologies suggest a formation mechanism. The main microbiological agent that contributed to these surface landforms appears to be dense aggregations of mucus-secreting pennate diatom frustules. Filamentous microorganisms (cyanobacteria) and macroorganisms (especially hydrobid gastropods) and their metabolic products (fecal pellets) are also embedded in the microbial mats (Fig. 4C–F). Besides the concentrations of mineral precipitates of calcium carbonate (low Mg calcite), calcium sulfate (especially gypsum), calcium magnesium carbonate (dolomite), and sodium chloride (halite), revealed by XRD and energy dispersive X-ray spectroscopy (EDX) data analyses from the collected samples, the abiotic components also include fine-grained (wind-blown?) dispersed sediment particles derived from clay minerals.
In the OD1 area, the surface of light brown and thick (up to 3–4 cm) living/partially mineralized microbial mats growing at the base of the waterfall (Figs. 4A, B and 5A) revealed a system of microterracettes delimited by ridges that transition into one another with a lateral continuity of a few millimeters (Figs. 5A and 6A). In these surface morphologies, which appeared homogeneously compact, viscous, and sticky to the touch, variable amounts of surface mineral precipitation, especially calcium carbonate, were ongoing at the time of sampling (Fig. 4C). A feature readily observable during analysis under the microscope was abundant strongly lanceolate and elliptical lanceolate diatom frustules that, including their associated slime (Fig. 7A, B), appear to be a primary constituent of fresh microbial mats. Most of the diatoms belong to the pennate, naviculoid genera Berkeleya and Mastogloia (Figs. 6B–D and 7A–E). These two genera abound in the living microbial mats of the OD1 area, whereas a further common pennate genus Ulnaria (Fig. 7F)—also referred to as Synedra in the previous pertinent literature—was detected in the OD3 area. Here, the needle-shaped Ulnaria is also associated with Mastogloia on top of fully mineralized microterracette morphologies (Fig. 5B, C), where the only available water present, during the field investigation, was from some discontinuous spray of the waterfall.

ESEM micrographs.
SEM/ESEM analyses performed on the surfaces of living/partially mineralized (air dried) microbial mats from the OD1 area reveal extremely high concentrations of frustules mostly belonging to the needle-shaped Berkeleya (Figs. 6D and 7B, C). Whereas in the inter-ridge surface of microterracettes, these frustules display a predominantly random arrangement (Fig. 6C, D), the ridges themselves appear largely composed of iso-oriented densely stacked accumulations of Berkeleya frustules (Fig. 6B). They are arranged in concentrations perpendicular to the current direction observed in the field and, therefore, providing a greater resistance to the current flow and, in this manner, favoring frustule accumulation. The result of this iso-oriented concentration is ridge formation, the asymmetrical profile of which (Fig. 6A, B) shows a steep slope on the down-current direction and, therefore, readily indicates the water flow direction. The naviculoid diatom Mastogloia is the second most abundant diatom genus in the living/partially mineralized OD1-microterracette-forming mats (Fig. 7B, D, and E). Its elliptic-lanceolate frustules have been detected in localized concentrations of a few individuals (Fig. 6D).
The microbial mats, if viewed in vertical cross section just beneath the microterraced surface, display closely packed palisades of large cyanobacterial trichomes (a single trichome is ∼50 μm large), heavily encrusted by micrite precipitation, in a prevailing vertical (phototactic) alignment (Fig. 8A). Cyanobacterial palisades are also visible in completely mineralized rocks (Fig. 4D, E). Again in the vertical section, and beneath the cyanobacterial palisades, an alveolar framework—with membranous morphologies (Fig. 8B) for which elemental (EDX) analysis indicates an organic composition—has alveoli becoming the loci of calcium carbonate precipitation organized as microcrystal aggregates that only partially fill the alveoli and, therefore, leaving a residual high porosity.
OD1 area: ESEM micrographs of the alveolar framework visible in vertical cross section, with normal stratigraphic orientation, on top and just beneath the microterraced surface of living (air dried)/partially mineralized microbial mat.
In the living microbial mats of the OD1 area, upon which microterracette morphologies were being formed, elemental (EDX) and SEM analyses revealed that the distribution and accumulation of mineral precipitates largely occurred in the still fresh gelatinous mat. Whereas single or small crystal aggregates of calcium sulfate, dolomite rhombs, and sodium chloride coatings locally occur, and can also be recognized on a morphological basis, low-Mg calcium carbonate is by far the main mineral precipitate.
Micritic calcium carbonate covers large sectors of mucilage biofilms (Fig. 9A, B) with microcrystalline (<1 μm diameter) aggregates and, therefore, it becomes the main agent for in vivo encrustation/mineralization of the organic structures (Fig. 9C, D). Recurring features observed atop the fresh mucilage biofilm also include alignments of micritic aggregates that gradually encrust organic membranes and strands with micritic covering arranged according to the linear flow of thin film of water (Fig. 9A). Precipitation of microcrystalline calcium carbonate takes place within the slime on the surface (Fig. 9E, F)—the cohesive nature of which enables the single particles (diatom frustules, bacterial cells, macrobenthos, aggregates of mineral precipitates, sand, and silt grains) to be held together—and in the underlying cyanobacterial palisades and alveolar framework.
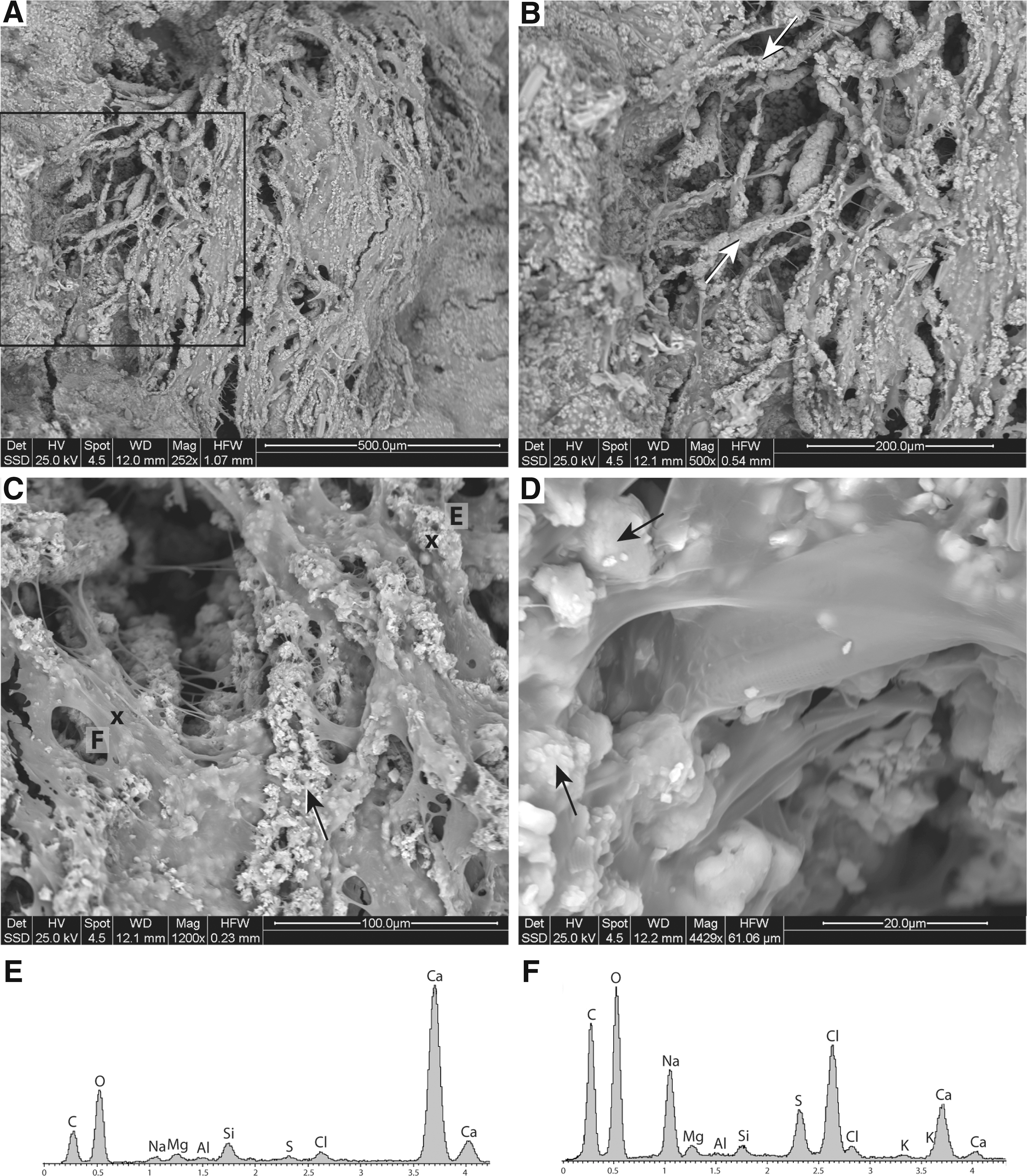
OD1 area: ESEM micrographs of the alignments of micritic aggregates on top of the partially mineralized mucilage biofilm in the microterracette areas (in
Locally, mineral precipitates also include microcrystal aggregates of calcium sulfate that give rise to microcrinkled (rough) surfaces produced by clusters of small-sized cauliflower-like structures also described elsewhere in arid regions (e.g., Stivaletta and Barbieri, 2009). Simulations of microcrinkled surfaces are also produced by silt and clay-sized particles (detrital material) when coated by sodium chloride.
Unlike the OD1 area, where microterracettes can be either partially or completely mineralized (Figs. 4A and 5A, D), in the OD3 area these morphologies are only completely mineralized in low-Mg calcium carbonate (Fig. 5B, C). There, the only available water at the time of sampling was from the spray of the nearby waterfall, and by narrow shallow rivulets that are deemed responsible from breaking up the continuity of the ridge (Fig. 5C). In this mineralized area Ulnaria, a mat-forming commonly early colonizer is the diatom genus most frequently recovered (Fig. 7F).
5. Sabkha Oum Dba and Biofilm Producers: A Discussion
Whether living in the water column or on a firm surface, the slime produced by single-cell organisms, whether they are prokaryotes or eukaryotes, displays a number of ecological functions (Wotton, 2004) with significant effects at the scale of physical microhabitats and with respect to sediment stabilization (Gerbersdorf and Wieprecht, 2015). In the areas of the Sabkha Oum Dba covered by microterraced surface morphologies, the contribution of mat-forming organisms, particularly diatoms, in their formation is crucial. Together with filamentous cyanobacteria—that in certain mineralized surfaces at Oum Dba exhibit the typical closely packed palisades and tufts of iso-oriented colonies—these diatoms are responsible for the formation of considerable amounts of mucilage of up to >1 cm in living/partially mineralized mats. The known symbiotic interactions between diatoms and cyanobacteria (e.g., Snoeijs and Murasi, 2004; Amin et al., 2012) allow the two groups of microorganisms to have a mutual advantage from their own mucilage. Furthermore, the microbial mat surfaces can subsequently be hardened by precipitation crusts of calcium carbonate (and other minerals) around the microbial cells (cyanobacterial trichomes) as a by-product of their respiration processes. Despite alleged difficulties in preserving diatom frustules in saline lakes (Flower and Ryves, 2009), at Oum Dba the formation and satisfactory preservation of surface micromorphologies appear to be the consequence of the existence of the mucilage, and the mineral events that they promote, in an environment based on oxygenic photosynthesis determined by cyanobacterial metabolism. Large (trichome width >50 μm) mat-forming filamentous cyanobacteria, similar to those described from the Sabkha Oum Dba, occur in a wide range of coastal and hypersaline environments (e.g., Stal, 2007; Whitton and Potts, 2007).
Despite the evident contribution of physical (hydrological) agents, the high abundance of Berkeleya valves in living/partially mineralized portions of the microbial mats of the OD1 area indicates that environmental conditions were suitable for the development of this taxon and its organization into colonies of a large number of individuals. Similar to many other benthic diatom genera, Berkeleya exudes amounts of extracellular polymeric substances (EPS), the main biofilm component in which these microalgae stay embedded during their life cycle (e.g., Hoagland et al., 1993). The abundance of residual biofilm in the inter-ridge areas of the sampled materials indicates that biofilm-producing diatoms actively contributed to the formation of the living mats (Figs. 6D and 7A, B). Differently, in correspondence of ridges, where the concentration of Berkeleya frustules is maximized and mostly attributed to physical transport (Figs. 6B and 7C), the residual portions of biofilm appear particularly scarce.
Furthermore, Mastogloia is an active producer of biofilm and has known symbiotic relationships with other biofilm producers, such as other motile diatoms and mucilage-secreting cyanobacteria (Snoeijs and Murasi, 2004; Adams et al., 2012). Therefore, it has an important role as a sediment biostabilizer (Awramik and Riding, 1988). Mucilage produced by diatoms also has a cohesive function on muddy sediments and may biostabilize loose sediments at the sediment–water interface by increasing the erosion threshold (Awramik and Riding, 1988; Decho, 1990, 2000; Winsborough, 2000; Stal and de Brouwer, 2003; de Brouwer et al., 2005). This biostabilization is an important prerequisite for the preservation and, ultimately, the fossilization of surface morphologies induced by microorganisms (e.g., Viles, 2012). In building microterracettes, therefore, the diatom slime could also play a preliminary (premineralization) function of biostabilization. This has been observed at Oum Dba (Fig. 5A).
Because the morphologies produced by microbial mats degrade rapidly, their preservation potential can be greatly increased by an early mineralization process. Along with biological products (i.e., biofilms, diatom valves, cyanobacterial filaments, shells, and excretory materials from microinvertebrates), mineral precipitates are primary components of the microterracette morphologies at Oum Dba. Similar to microbial mats in other modern ecosystems around the world (Dupraz et al., 2009), in this sabkha CaCO3 is the main precipitating mineral, with a remarkable potential of inducing their preservation as lithified mats. In this type of natural system, the microenvironment suitable for CaCO3 formation is determined by the metabolic activity of microorganisms, such as aerobic respiration and/or sulfate reduction, associated with environmental conditions that may influence CaCO3 precipitation (e.g., Dupraz et al., 2009, 2013; Kaźmierczak et al., 2015). The abundance of EPS might also have a positive impact on mineralization since the EPS matrix can promote the nucleation and growth of carbonate minerals by binding Ca2+ and Mg2+ ions (Dupraz et al., 2004, 2013; Ercole et al., 2012; Gerbersdorf and Wieprecht, 2015). At the OD1 site, the surfaces of fresh and living microbial mats exhibit microcrystal aggregates of calcium carbonate (Fig. 4C) that seem particularly concentrated where there is a greater production of mucilage (Fig. 7A). This confined area of the sabkha exhibits a complete transition from non- or poorly lithified surface morphologies and mat (gelatinous mat, Fig. 5A) up to completely mineralized morphologies (Fig. 5D).
The change in pH values, with a consequent effect on mineral precipitation, is the presumable main factor that has also changed the redox potential (Eh values) between day and night (Table 1), with increasing Eh measured at sunlight, as a result of oxygen production during photosynthesis, and a marked decrease of Eh values at night (see also Friedman and Foner, 1982).
At Oum Dba, the three most abundant diatom genera are all capable of secreting extracellular mucilage, which also supplies nutrients to meiofaunal grazers. This is especially the case for the populations of small hydrobid gastropods that crowd and graze the living mats (Fig. 4C) and that also leave traces of movement and rest. Shells of hydrobid gastropods often appear trapped in the CaCO3 precipitates (Fig. 4F). In addition to being a meiofaunal food source, mucilage biofilm appears to help protect them from desiccation, especially in arid environments (e.g., Gerdes, 2003). This may further explain the local high abundances of hydrobids and their excretory products.
6. Microgeomorphologies of the Sabkha Oum Dba as Astrobiological Tool
In different geological issues, high-resolution image spectroscopy has greatly benefited from the use of miniaturized instruments, with the spatial resolution of a centimeter or less and unexpected new insights (Greenberger et al., 2015). Recently developed miniaturized imaging spectrometers—such as, for example, the multispectral microscopic imager (Núñez et al., 2014), ultra compact imaging spectrometer (Van Gorp et al., 2014; Fraeman et al., 2016), laser ablation mass spectrometer (Neubeck et al., 2016), and Raman imaging spectrometer (Hutchinson et al., 2014)—have been specifically designed for future missions in martian environments and are expected to achieve submillimeter spatial resolutions. This new generation of analytical tools can investigate extremely small-scale subjects, such as single-grain/crystal compositions or micromorphologies, and is of particular interest for astrobiology because of the possibility to identify aqueous mineral compositions and textures as well as structures possibly related to a biological origin or any other feature able to assess past (or present) signs of habitability. A useful preliminary tool to promote the use of these rapidly expanding techniques in future planetary missions includes the study and interpretation of several terrestrial microgeomorphologies produced with the contribution of life forms—in particular the microbial-based morphologies—that can be potentially present on planetary bodies that have (or have had) Earth-like environmental conditions, beginning with Mars, based on its geological and climatic history, and the potential to host life as we know it.
Anywhere on Earth where some liquid water (at least transient) exists, microbes can interact with physical substrates and produce suites of morphologies potentially preservable in the geological record. Besides stromatolites, the so-called microbially induced sedimentary structures (Noffke et al., 1996)—that is, widespread microbial structures and other morphologies that document the interaction between microbiota and physical environment developed in shallow siliciclastic shelf settings—have been extensively investigated because of the abundance of fossilized remains that retain structures diagnostic of some biological activity. Astrobiological potential is not only limited to the direct products of the interactions between microbial organisms and physical environment. Traces and ichnofabrics from different biological behaviors, for example, represent a wide field of knowledge still largely unexplored in terms of astrobiological application (Baucon et al., 2017).
In the case study described herein, the astrobiological relevance is primarily connected with its habitat conditions that is among those hypothesized on Mars during the Noachian/Hesperian transition, a period in which the climate and hydrological cycle may have allowed fluvial–lacustrine conditions with plausible possibilities of being habitable (e.g., Grotzinger et al., 2014; Wordsworth, 2016). A biological component mainly represented by cyanobacteria and benthic diatoms also characterizes other alleged environmental analogues for Mars at the end of Noachian, as are high-altitude hypersaline lakes (e.g., Cabrol et al., 2007) or hydrothermal fields (e.g., Ruff and Farmer, 2016).
The issue of the significance of diatoms in astrobiology is, however, rather debated, and although they represent a relatively complex form of life, diatoms have long been involved in the discussion over panspermia (Hoover et al., 1986). Diatoms are able to inhabit the widest range of terrestrial extreme environments (Battarbee et al., 2001), and their role, after their recovery in the upper atmosphere, has also been reconsidered. An alleged provenance from the space, claimed for diatom frustules found in the stratosphere (Wainwright et al., 2013), indeed, could also be explained by merely terrestrial events, such as the upward transport of explosive volcanism (Van Eaton et al., 2013). Similarly, the recovery of diatom frustules in carbonaceous meteorites assumed free from terrestrial contamination (Wickramasinghe et al., 2013) is also a hypothesis welcomed with skepticism. A still persistent relationship of diatoms with travel through space, however, seems rather dependent on their ability to produce heavy resting spores (e.g., Härnström et al., 2011) and of their fossil frustules to survive destruction during hypervelocity impact experiments (Burchell et al., 2014).
There is a wide interest in the existence of ancient lakes on Mars (Cabrol and Grin, 2010), and dry lake deposits, in particular, which have been hypothesized since the time of the Viking mission, make sabkhas interesting analogues to at least some of the sulfate and carbonate deposits that on Mars are presumed derived from lacustrine environments (Ori, 2010). In dry lakes, the formation of salt deposits mainly occurs by the evaporation of standing bodies of water, such as in arid environments, or in hydrothermal evaporite systems (such as at Yellowstone and Dallol). In these environments, water and salts can be colonized by halophilic microorganisms, which may also contribute to the formation of mineral precipitates (such as biomineralized carbonates) and produce distinctive micromorphologies that can reveal their biological origin, as in the case described in the Sabkha Oum Dba.
Overall, based on the type of habitat, the biological communities, the formation of bioinduced and preservable micromorphologies, and the precipitation of evaporitic minerals, such as carbonates and sulfates, the Sabkha Oum Dba can be considered a site of remarkable astrobiological significance.
The question of the nature and preservation of biosignatures, and the means for their identification in an astrobiological perspective, is still an open issue for which diverse approaches have been suggested (e.g., Westall et al., 2015). The case of evaporitic lacustrine environments—which on Earth thrive with a wealth of microorganisms and have a significant microbial habitability potential (Barbieri, 2013; Westall et al., 2013)—deserves careful consideration because of the abundance of salts on the martian surface and shallow subsurface, and some of them may have been deposited in this type of lacustrine environment. This is the case, for example, of the Gale Crater, the landing site of the Mars Science Laboratory, where, on the basis of the discovery of calcium sulfate veins in fracture fills, evaporitic sabkha-like settings have been hypothesized (Grotzinger et al., 2014, 2015; Nachon et al., 2014).
7. Conclusions
The activity of prokaryotic and eukaryotic microorganisms, favored by suitable physical processes, can produce distinctive surface/shallow subsurface geomorphic features of astrobiological relevance in case the environment, and its geological and biological attributes, approach those expected on Mars. This is the case of the microterracing described herein, which is originated by the interplay between the physical conditions in a confined area of a continental sabkha permanently fed by salty groundwater and mat-forming benthic diatoms and cyanobacteria. This small-scale geomorphic process appears enhanced and stabilized by the early cohesive action of the mineral precipitation promoted by metabolic activities (i.e., intense biofilm production and aerobic respiration) of microorganisms, which has contributed to the massive preservation, during and after burial, of diatom frustules and sheathed cyanobacterial colonies. Microterracettes, in addition to representing themselves a morphological biosignature determined by water flow, mineral precipitations, physical (slope) gradients, and microbiological action, can also represent a potential repository for other structures and remain directly derived from prokaryote/eukaryote communities.
The astrobiological significance of natural environments hosting microterracette morphologies is related to the existence, during phases of the geological history on the martian surface, of hydrated evaporite and/or hydrothermal evaporite systems from presumably confined (lacustrine?) environments, which are the potential conditions under which similar surface morphologies may have been formed.
Footnotes
Acknowledgments
We would like to thank Brahim Abounacer for field assistance, the late Bruno Capaccioni for water geochemistry analyses, Patrick Kociolek for diatoms identification, Carlo Ferrari for recognition of the halophyte plants, Giorgio Gasparotto for his help with the mineralogical determinations, Daniele Scarponi for recognition of the hydrobid gastropods, Massimo Tonelli for ESEM assistance, Fabio Gamberini for rock processing, and two anonymous reviewers for their constructive comments. This study was funded by MIUR PRIN 2012 and the University of Bologna (RFO 2014–2015).
Author Disclosure Statement
No competing financial interests exist.