
Abstract
The potential habitability of an exoplanet is traditionally assessed by determining whether its orbit falls within the circumstellar “habitable zone” of its star, defined as the distance at which water could be liquid on the surface of a planet (Kopparapu et al., 2013). Traditionally, these limits are determined by radiative-convective climate models, which are used to predict surface temperatures at user-specified levels of greenhouse gases. This approach ignores the vital question of the (bio)geochemical plausibility of the proposed chemical abundances. Carbon dioxide is the most important greenhouse gas in Earth's atmosphere in terms of regulating planetary temperature, with the long-term concentration controlled by the balance between volcanic outgassing and the sequestration of CO2 via chemical weathering and sedimentation, as modulated by ocean chemistry, circulation, and biological (microbial) productivity. We developed a model that incorporates key aspects of Earth's short- and long-term biogeochemical carbon cycle to explore the potential changes in the CO2 greenhouse due to variance in planet size and stellar insolation. We find that proposed changes in global topography, tectonics, and the hydrological cycle on larger planets result in proportionally greater surface temperatures for a given incident flux. For planets between 0.5 and 2 R ⊕, the effect of these changes results in average global surface temperature deviations of up to 20 K, which suggests that these relationships must be considered in future studies of planetary habitability. Key Words: Planets—Atmospheres—Carbon dioxide—Biogeochemistry. Astrobiology 18, 469–480.
1. Introduction
A
Our knowledge of the geochemical cycles that have controlled atmospheric composition through the history of our home planet remains incomplete, but careful studies have elucidated what appear to be the controlling factors in the carbon cycle over the lifetime of Earth (Berner, 1991, 2006; Sleep and Zahnle, 2001; Hayes and Waldbauer, 2006), and the resulting modeled changes in CO2 concentration and global surface temperature appear reasonable when compared to what direct evidence is available (Royer et al., 2004; Mills et al., 2014). In this paper, we adapt a model of Earth's carbon cycle to infer the temperature and habitability for Earth-like planets of different sizes and star properties.
Earth appears to have maintained habitable (0–70°C) conditions at its surface for as much as 4 billion years (Gyr) in spite of significant increases in solar luminosity over this period (Sackmann et al., 1993; Rushby et al., 2013). This is due to the effect of increased solar forcing (or other non-CO2 temperature forcing) on the balance between the drawdown of atmospheric CO2 via the weathering of silicate minerals, which is temperature dependent, and the tectonic and volcanic emission of CO2 to the atmosphere, which operates independently of surface temperature. This silicate weathering feedback, first postulated by Walker et al. (1981), has most likely led to a steady reduction in atmospheric CO2 concentration over the lifetime of Earth. Direct estimation of early Earth CO2 concentration from geological proxy records is difficult, but efforts to do so have often suggested upper limits (see review in Feulner, 2012), which generally do not provide the required radiative forcing to maintain liquid water at the planetary surface. By contrast, numerical models based on the carbonate-silicate cycle generally predict higher concentrations (e.g., pCO2∼100 PAL; Sleep and Zahnle, 2001) in line with the strong negative temperature feedbacks present—without liquid water the key CO2 sink is removed and the atmospheric concentration can increase unbounded.
It should be briefly noted that determinations of radiative forcing of large amounts of CO2 remain under active debate (Halevy et al., 2009), and incorporations of newly measured laboratory parameters have continuously modified the “radiative effect” that a given amount of CO2 will have. Differential incorporation of various physics (e.g., line absorptions at high temperatures, collisionally induced absorption, pressure broadening) involved with radiative forcing of high-CO2 atmospheres has therefore allowed various re-computations of the “circumstellar habitable zone” (e.g., Kopparapu et al., 2013, 2016; Airapetian et al., 2016) in a manner that makes it difficult to directly compare studies and their conclusions over time.
Debate around the CO2 greenhouse/surface temperature relation will continue, but it is generally accepted that on an Earth-like planet with active tectonics, gradual warming of the host star will cause atmospheric CO2 to decline over time. In this paper, we investigate the significance of the carbonate-silicate weathering feedback in modifying habitable lifetimes relative to previously developed simple energy balance approaches, focusing on predicting biogeochemically self-consistent CO2 concentrations and their evolution through planetary history. We extend the model to assess the first-order effect of planet size on this process and determine how the drawdown of CO2 over geological time effects the reorganization and eventual termination of life on Earth and other potentially life-bearing planets of varying sizes.
2. Methodology
A carbon cycle model following Sleep and Zahnle (2001) (henceforth “SZ01”) is constructed to compute levels of atmospheric carbon dioxide (pCO2) and globally averaged surface temperature for a given incident flux in the range 0.75–1.25 S ⊕, where 1 S ⊕ corresponds to the present-day incident flux at the top of the atmosphere (1367 W m−2). This model is constructed of four carbon reservoirs: Ocean/Atmosphere (R oa), Oceanic Crust (R oc), Continental Sediment (R cs), and the Mantle (R man) (see Fig. 1). Here, “continental sediment” indicates both the carbonates and organics present in the continental crust as well as the C-bearing sediments on the continental shelves, which are geologically homologous (SZ01). Carbon is transferred through the system by a series of fluxes, constrained by the assumption of a steady-state preindustrial carbon cycle. The temporal resolution of the model is of the same order as the complete equilibration time of carbon in the ocean atmosphere system (∼106 yr).
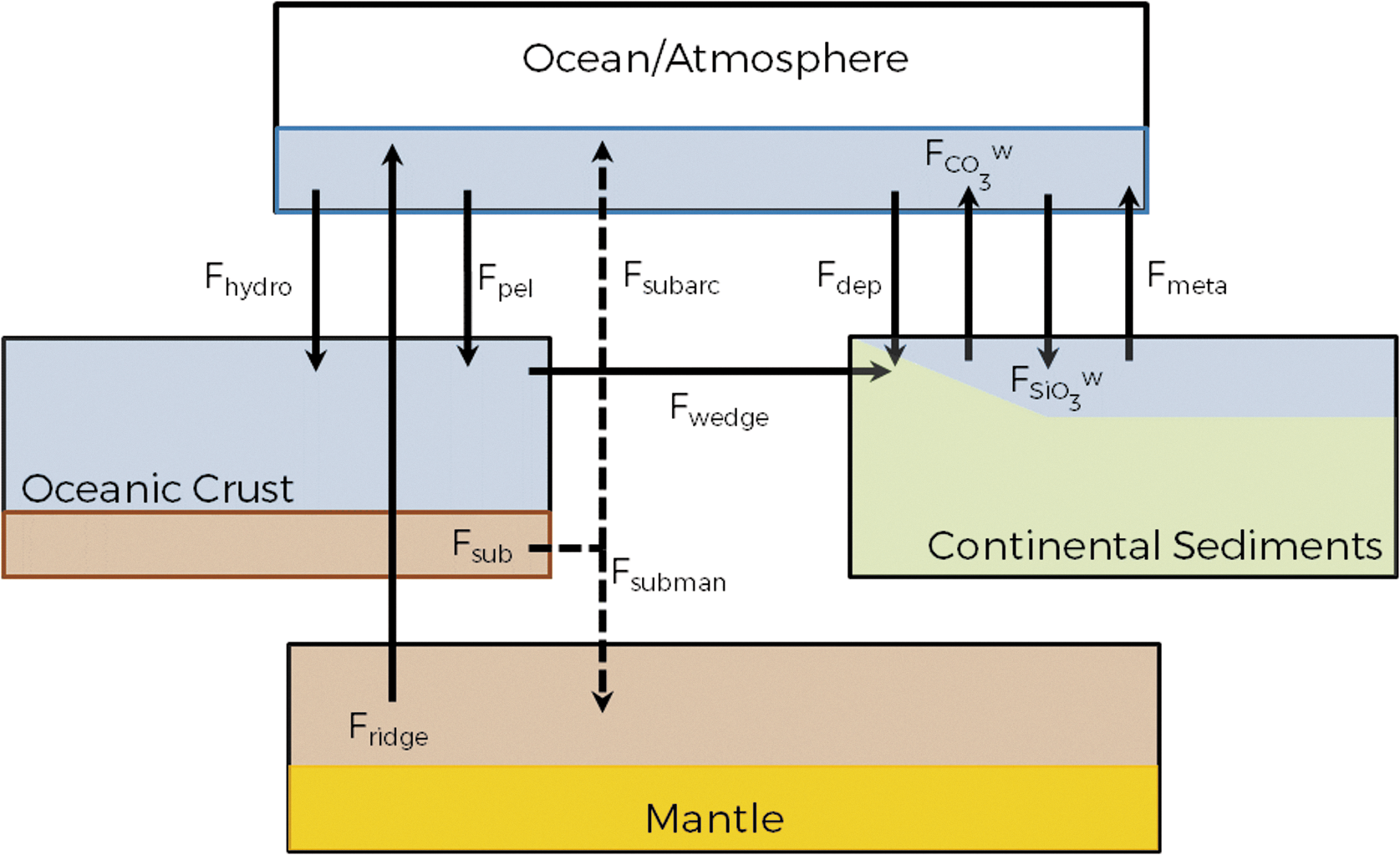
Model schematic illustrating carbon reservoirs and fluxes.
Planetary surface temperature is computed from a 1-D radiative-convective climate model with an Earth-like atmospheric water vapor profile, derived from a 1 bar, cloud- and ozone-free, N2-dominated model atmosphere, which includes CO2 and H2O absorption coefficients from the HITRAN and HITEMP line-by-line databases (Haqq-Misra et al., 2008; Kopparapu et al., 2013).
To determine pCO2, carbon is separated between the ocean and the atmosphere by a partitioning constant (φ), which, assuming a preindustrial pCO2 of 280 ppm, is 0.016 (Bergman et al., 2004). However, Omta et al. (2011) suggested an approximately exponential dependence between atmospheric CO2 and average ocean temperature, so we include an additional scaling that alters the partitioning of CO2 between the ocean and the atmosphere as a function of temperature. This allows for the model to account for large changes in ocean temperature, for example those that would occur toward the end of the planet's habitable lifetime as solar output increases steadily, on the partitioning of carbon species between the ocean and the atmosphere. Their scaling produces an approximate 4% increase in fractional atmospheric CO2 per 1 K increase in ocean temperature, relative to present-day ocean temperatures. To ensure mass balance, atmospheric volume was normalized to atmospheric scale height, which was computed as a function of g under the assumption of an Earth-like dry air gas constant and mean atmospheric temperature of 288 K.
While planetary habitability is a complex and multifaceted concept that is not easily reducible to single parameter or metric, for the purposes of this work we consider habitability to be constrained to first order by average surface temperature and available pCO2 for oxygenic photosynthesis by C3 and C4 terrestrial organisms (Cockell et al., 2016). Surface temperatures of between 273 and 343 K are applied as lower and upper bounds; 273 K representing a limit for “Snowball” conditions and 343 K representing the upper limit, given that beyond this many of the proteins involved in photosynthesis begin to degrade (Rothschild, 2007). Excluding the survival of certain extremophilic organisms on Earth, which have been reported to endure temperature extremes ranging between 253 and 395 K, these limits encapsulate the habitats of most life on the planet (Junge et al., 2004; Takai et al., 2008). However, setting an arbitrary habitable constraint at 273 K may be too limiting in terms of the stability of the climate, as a strong positive ice-albedo feedback will most likely result in an entirely glaciated planet up to a global average temperature of 278 K (Pierrehumbert, 2010). But conversely, it appears that Earth may have strayed below this lower limit during two distinct periods around 2.3 Ga and 700 Ma (Hoffman et al., 1998; Kirschvink et al., 2000), not necessarily to the detriment of the evolving biosphere.
Photosynthetic primary producers on Earth require atmospheric carbon dioxide to produce organic compounds, producing oxygen as a by-product of this process. Low levels of atmospheric CO2 will inhibit these organisms' metabolic function and will therefore also affect levels of atmospheric oxygen crucial for the metabolisms of most eukaryotic organisms. The vast majority of terrestrial carbon capture is performed by plants utilizing either the C3 or C4 photosynthetic pathway (representing 89% of plant species, combined) (Bear et al., 2016). The effective pCO2 limit for these organisms is ∼150 and ∼10 ppmv, respectively (Lovelock and Whitfield, 1982; Caldeira and Kasting, 1992; O'Malley-James et al., 2013, 2014). Below these limits, photosynthesizers that use these carbon concentration mechanisms become less able to effectively capture carbon from the atmosphere in order to build biomass. Given the importance of these organisms as primary producers in the terrestrial biosphere, we conclude that the biosphere would effectively undergo a significant reorganization following their extinction, possibly resembling that of the anoxic, early history of Earth.
2.1. Planet mass-radius relationships
An understanding of the interiors of exoplanets requires (at a minimum) knowledge of their mass and density. Radial velocity techniques allow for planet mass to be estimated (up to a degeneracy in the orbital inclination) based on the gravitational effect that the orbiting world has on its host star. By using transit photometry, the radius of an orbiting planet can be estimated based on the amount of light that the planet obscures, and its mass can be determined if the composition of the interior of the planet is known (Jenkins et al., 2002). However, radius and mass estimates are not sufficient to identify a planet's interior composition, with the possible exception being at the extremes in density—very low-density planets are likely to be dominated by H and He, and very high-density planets are likely composed primarily of Fe (Sotin et al., 2010). Scaling relationships between planet radius and mass can be used to constrain geodynamic processes operating on exoplanets to first order, if a series of simplifying assumptions are made regarding bulk composition, surface gravity, geothermal heat flux, as well as mantle and core radii (Seager et al., 2007; Sotin et al., 2010). As mass is a product of the volume of the planet and its density, it scales with the approximate cube of the radius, modified by the effect of the bulk composition of the interior. Although some uncertainty exists as to the upper radius limit for rocky worlds (e.g., Rogers, 2015), we choose an optimistic upper limit of 2 R
⊕ and a lower limit of 0.5 R
⊕. We then adopt the mass-radius relationships presented by Barnes et al. (2015):
By scaling the normalized size of the crustal reservoirs of carbon using these relationships, the effect of increasing mass on carbon storage and transfer between these reservoirs can be quantified.
2.2. Volcanism
Some mode of volcanic outgassing that enables atmospheric volatile replenishment is considered to be a necessary prerequisite for long-term planetary habitability due to the ability of volcanic outgassing to significantly alter the composition of planetary atmospheres (Kasting and Catling, 2003; Korenaga, 2012). However, it is currently unclear as to whether the tectonic regime observed on Earth is typical of terrestrial planets of similar size, or indeed how these processes will change with planet size (Kite et al., 2009). Volcanism is expected to be a widespread characteristic of rocky planets resulting from partial melting of their silicate mantle under extremes of pressure, but contention exists over the form of thermal convection that would dominate on massive planets and to what alternative form their tectonic regimes may take (O'Neill and Lenardic, 2007; Valencia et al., 2007; Kite et al., 2009).
Regardless of the mechanism of convection, internally generated heat must escape, following the laws of thermodynamics. Assuming that the abundance of radiogenic elements in a planet is proportional to its mass, then the radiogenic heat flux (Q) varies as a function of planet mass, according to Sotin et al. (2010):
where Q
⊕ is the geothermal heat flux on Earth. While acknowledging considerable uncertainty regarding the rate of geothermal heat loss during the Archean, this model follows the precedent of SZ01 (as well as Franck and Bounama, 1999; Lowell and Keller, 2003; Hayes and Waldbauer, 2006) in assuming a time-dependent decay function in the form
where t 0 describes the time of planet formation. This parameterization predicts heat loss rates from early Earth between 4.5 and 3 times that of present day. This relationship has a significant control on the likely geochemical and climatic evolution of the planet as it is a key driver in many of the flux terms associated with the carbon cycle. In geodynamic models, the seafloor spreading rate is found to closely approximate Q 2 and has a strong control over the rate of subduction of oceanic crust into the mantle, and by implication the amount of carbon returned to the ocean/atmosphere system via arc volcanism (F subarc) as well as via ridge degassing (F ridge), metamorphism (F meta), and the rate of seafloor hydrothermal carbonatization (F hydro) (Franck et al., 1999; SZ01), which are defined in Table 1 and Fig.1.
Davies and Davies (2010).
Trenberth et al. (2007).
2.3. Weathering
Silicate weathering and its dependence on temperature and atmospheric CO2 are an integral driver that controls long-term planetary climate. A negative feedback between the rate of weathering of silicate rocks, the surface temperature of the planet, and the partial pressure of carbon dioxide in the atmosphere operates to regulate pCO2 over geological time (>106 yr); as surface temperatures increase (i.e., due to steadily increasing solar luminosity), rates of continental weathering increase, thereby drawing down CO2 and reducing the overall radiative forcing (Walker et al., 1981). Studies find that the rate of CO2 sequestration via continental silicate weathering is a strong function of temperature and a weak function of pCO2. Our formulation of this process, following SZ01, is
Here, β is an exponent that controls the sensitivity of the reaction to the partial pressure of atmospheric carbon dioxide and the rate of carbonic acid formation (held at 0.3 for this work [SZ01]), and B is a dimensionless weathering calibration parameter suggested by Walker et al. (1981) to account for runoff (B = 13.7). The additional parameter κ simulates the effects of biotic enhancement of weathering, where terrestrial organisms via the action of their roots, metabolisms, or exuded organic acids increase the rate of weathering of silicate minerals beyond what would be expected in an abiotic environment (Schwartzman and Volk, 1989; Berner, 1997; SZ01). The delivery of bicarbonate ions and silicate cations to the ocean is thought to be strongly dependent on the supply of material via erosion, and by precipitation and runoff (West et al., 2005; Hartmann et al., 2014). The additional scaling parameter R E in Eq. 4 represents our first-order expectation that evaporation rates will scale with the fraction of planetary surface area covered by ocean. R E captures both variable ocean/continental fractions and associated carbon partitioning between oceanic and terrestrial environments, as well as evaporation rates from those surfaces. A variable ocean/continental fraction is given in this zero-D formulation as a parameter that determines the contrast between the albedo of the ocean and the land surface, which in turn affects the energy balance at the surface. The initial carbon inventory of the ocean and continental reservoirs is then proportionally partitioned relative to the steady-state, present-day Earth case.
A comparison with the COPSE geochemical model (Bergman et al., 2004) was carried out under conditions of differing biotic weathering enhancement (κ), and our modeled pCO2 output demonstrated good agreement with that model over the Phanerozoic (Fig. 2a). COPSE has a greater temporal resolution and includes a sulfur (S), phosphorus (P), and partial iron cycle, important for accurately representing the global oxygen cycle, which produces more-detailed, high-resolution pCO2 data, as output is constrained by comparison to δ34S and δ13C isotopic records. As it is clear from this comparison that the model developed here lacks the ability to resolve highly temporally variable CO2 and temperatures associated with the Phanerozoic, mean values of CO2 and temperature from COPSE (split at the Permo-Carboniferous) have also been included for a more appropriate comparison. Based on this comparison, we hold κ at 0.5 for this model, representing a doubling of terrestrial weathering rates by biology. Furthermore, our model output demonstrates reasonable agreement with available geochemical proxy data from the work of Feulner (2012). Figure 2b (modified from Feulner, 2012) displays pCO2 as a function of S/S ⊕ generated from default model conditions, as well as estimates of atmospheric carbon dioxide from other sources. Disparity between this work and that of Rosing et al. (2010) can be explained by their underestimate of surface albedo.
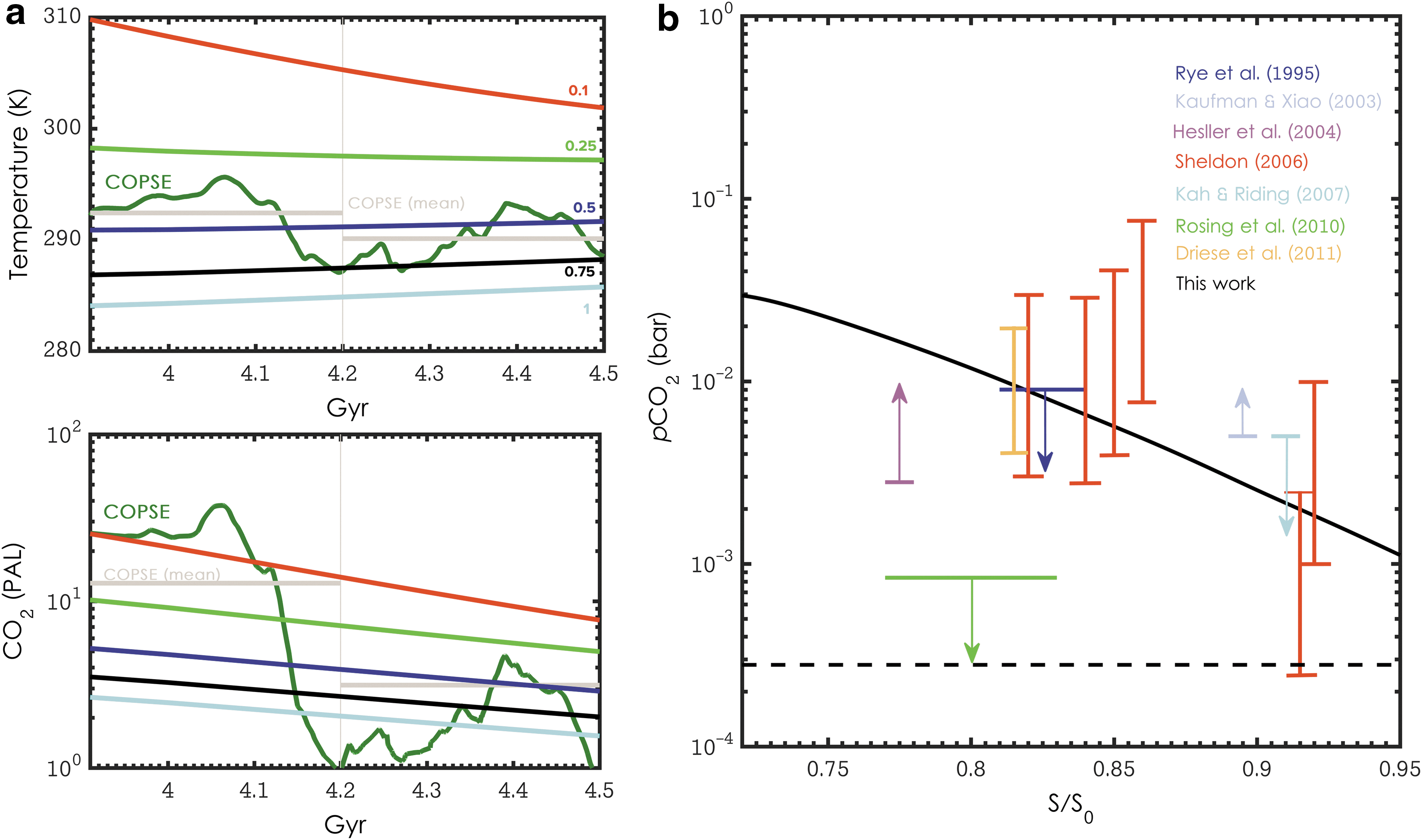
Ensemble of sensitivity analyses. (
3. Results
When coupled with a stellar evolution model that predicts luminosity over time, our model allows for estimation of the duration of habitable conditions on Earth-like worlds. For example, by using the stellar evolution data from the model presented by Rushby et al. (2013) and setting an upper temperature limit of 343 K, the habitable period of an Earth-like world around the Sun can be quantified (Fig. 3a). For a 1 R ⊕ planet at 1 AU (e.g., Earth), this limit is approximately 6.23 Gyr after planet formation, or 1.69 Gyr from present day. Additionally, atmospheric CO2 falls below the C3 and C4 plant compensation limit after ∼5.37 and ∼5.89 Gyr, respectively, which would put increasing stress on primary producers dependent on oxygenic photosynthesis for energy production (Fig. 3b). The carbonate-silicate buffer becomes ineffective at this stage, as CO2 is not returned to the atmosphere/ocean system at an equivalent rate as it is being removed due to the lower geothermal heat flux, ocean-floor spreading, and tectonic outgassing rates.
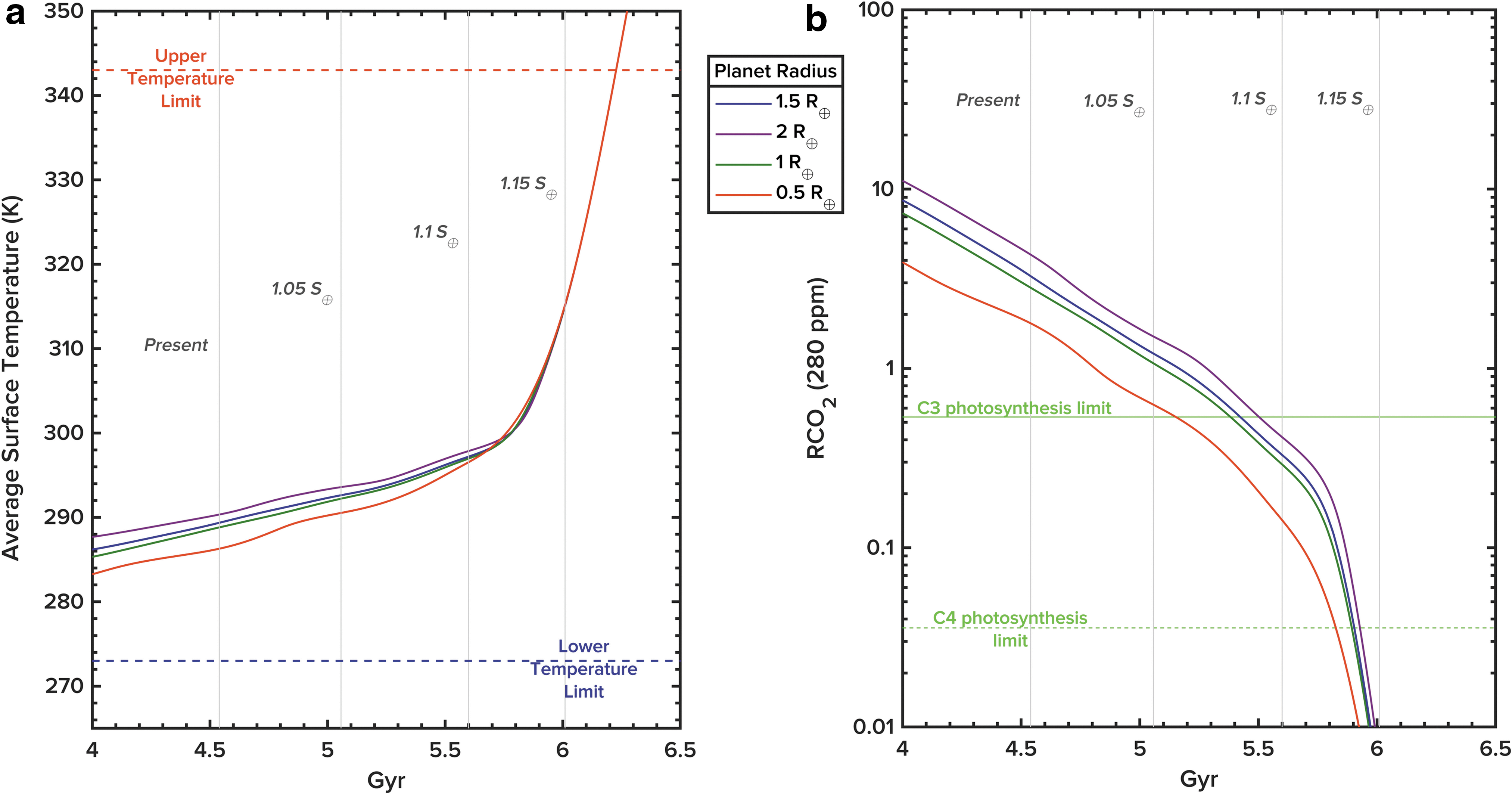
(
Notably, these “photosystem” compensation limits are reached some time before the model predicts temperatures to be above the habitable maximum, suggesting that considering “habitability” merely as a product of incident flux and surface temperature is too limited and neglects the inherent complexities of the interconnected planetary system. That considered, regardless of planet size (between 0.5 and ∼2 R ⊕) a “runaway greenhouse” event (set at the limit when outgoing longwave radiation is greater than incoming shortwave radiation) occurs in our model when incident flux on the planet is between ∼1.13 and 1.15 S ⊕. We note that there is a very slight planet size dependence on the timing of the “moist greenhouse” transition that occurs just prior to full runaway (beginning around ∼1.1 S ⊕). However, the radiative effect of these slight differences is negligible as CO2 remains uniformly low across the range of planet sizes at this stage because the extreme enhancement in silicate weathering at very high insolation overwhelms minor differences in CO2 content and outgassing rates. Larger planets are, however, warmer during the early stages of the simulation when incident flux is lowest and CO2 concentration highest, but given the limitations of this model in terms of representing the likely surface and atmospheric conditions of the early history of a terrestrial planet (e.g., the hydrogen/helium paleoatmosphere, a “magma ocean” phase, or a period of heavy bombardment), this result is likely not fully representative of the actual conditions we may expect during this period. Our primary focus is the stage at which high stellar flux and secularly decreasing pCO2 begin to dismantle the carbonate-silicate buffer, which in the Sol-Earth system is toward the end of the Sun's main sequence lifetime but several billion years before its red giant phase.
This model predicts that planets with larger radii tend to have higher average surface temperatures for a given incident flux during their habitable lifetimes (Figs. 3a and 4). For example, a 2 R ⊕ planet receiving the same incident flux as the present Earth is ∼5 K warmer on average than an Earth-sized world (Fig. 4). The reasons for this are manifold: higher geothermal fluxes (that also persist for longer after the planet's formation due to a more significant radiogenic inventory) leading to greater rates of outgassing of CO2, and subduction and seafloor spreading contributing to higher average pCO2 values for larger planets (Figs. 3b and 5). Terrestrial weathering fluxes to the oceans are also more significant due to greater chemical weathering of continental materials, somewhat but not entirely counteracting the increased outgassing and leading to more rapid cycling of carbon through the planetary system overall.
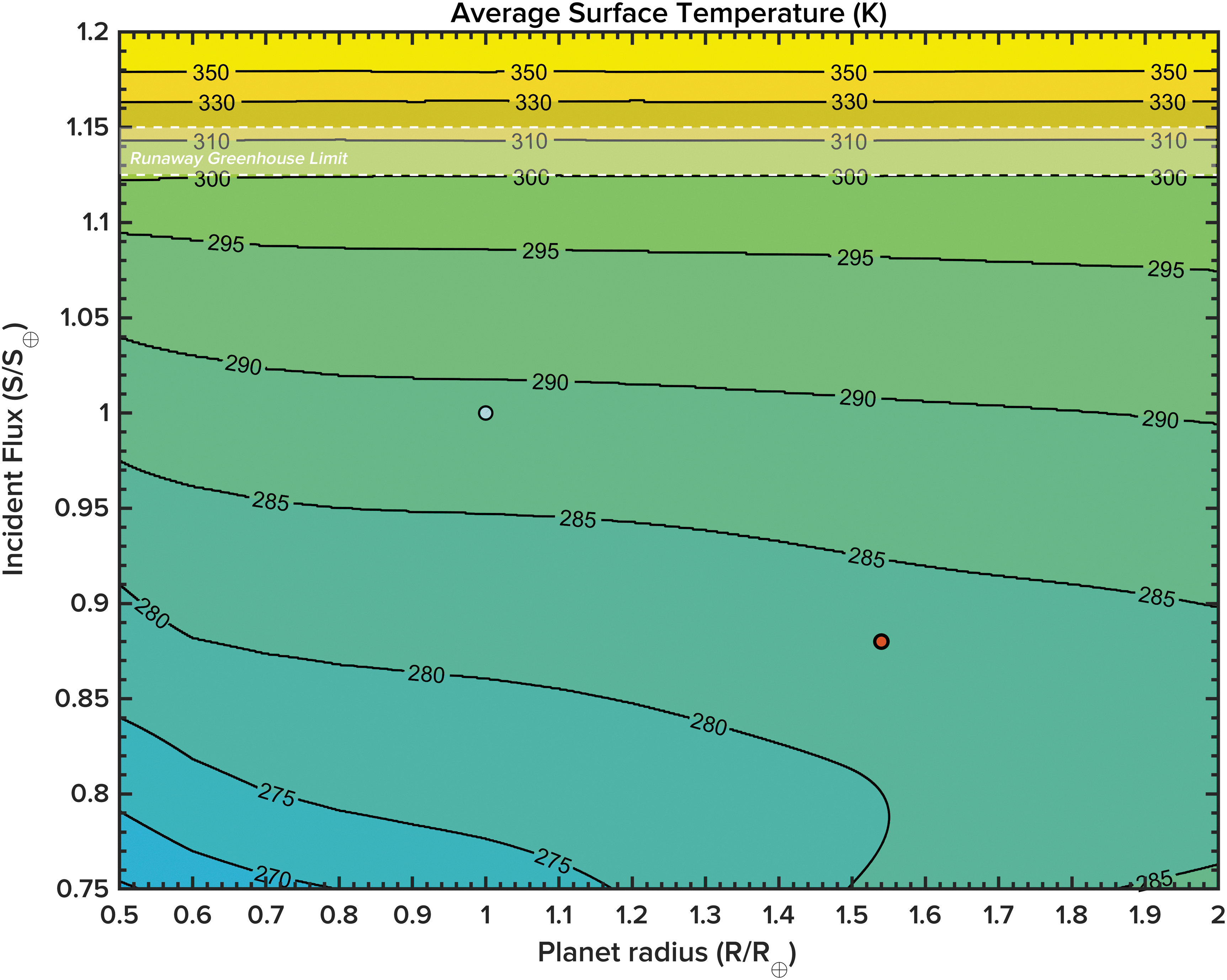
Contours of average surface temperature in K as a function of planet radius (in Earth-relative units) and incident flux (relative to present-day Earth). Also displayed are current Earth (blue marker) and GJ 667Cc (red marker).
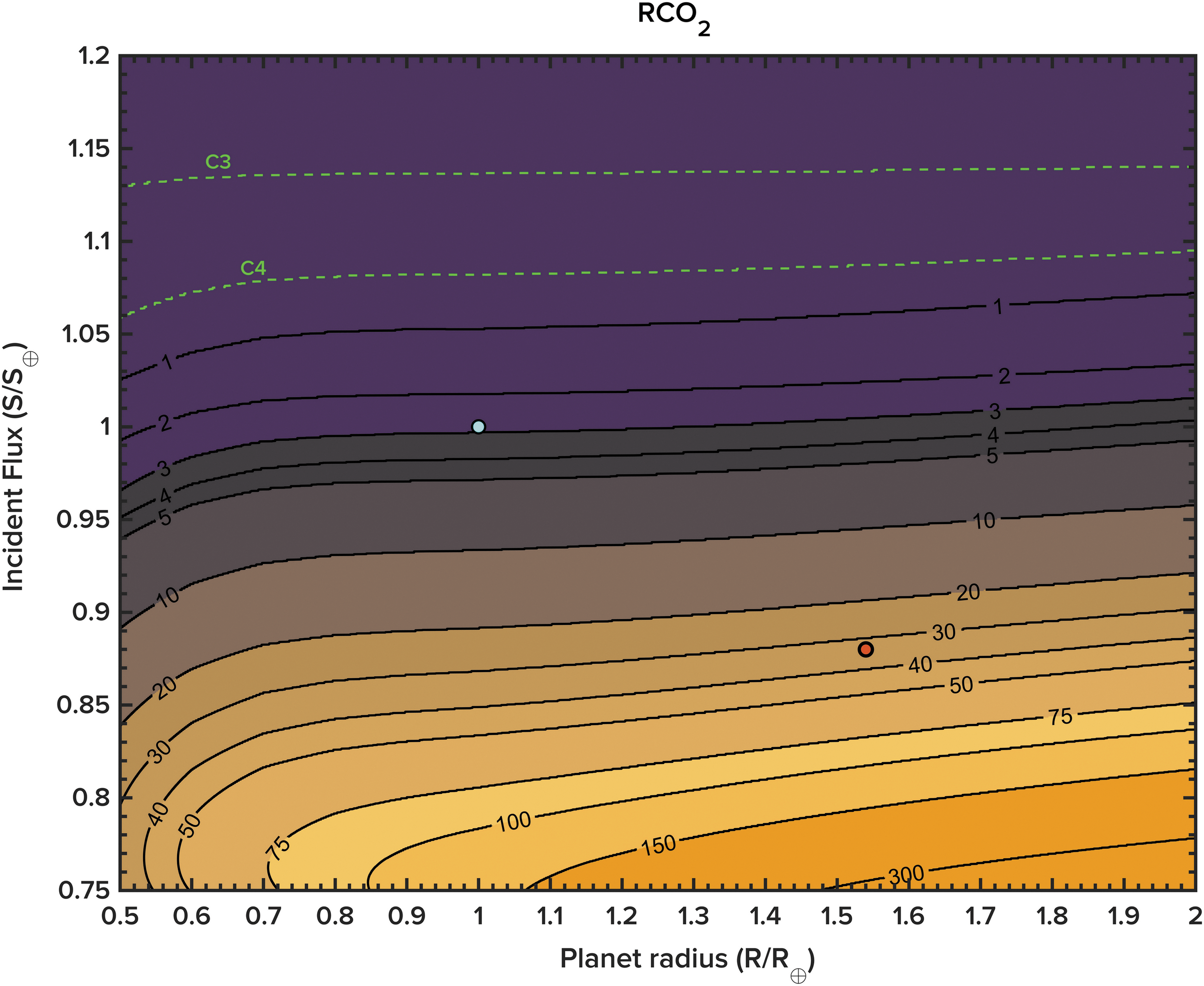
Contours of atmospheric CO2, in preindustrial units (where 1 RCO2 = 280 ppm), as a function of planet radius (in Earth-relative units) and incident flux (relative to present-day Earth). Also displayed are the C3 and C4 photosynthesis limits.
Atmospheric CO2 levels on larger planets remain higher for a given incident flux despite this enhanced terrestrial weathering. They also remain above the carbon fixation limits of photosynthesizing primary producers for proportionately longer, which would in turn allow for a biosphere based on oxygenic photosynthesizers to persist for longer than on smaller planets. As an example, this model suggests that a 2 R ⊕ planet at 1 AU around the Sun would maintain pCO2 levels above the C3 and C4 photosystem limits for ∼5.5 and ∼5.93 Gyr, respectively. This is 130 and 40 million years longer than may be expected on an Earth-sized planet at the same separation. As the “runaway greenhouse” limit discussed above seems to be mass-independent (over the range of planetary masses considered in this work), the primary control that planet size exerts on long-term habitability is through the recycling of carbon through the planetary system over geological time.
Conversely, planets smaller than 1 R ⊕, due to their smaller radiogenic inventories and associated heat flux, experience less vigorous rates of outgassing, subduction, seafloor spreading, and terrestrial weathering. A 0.5 R ⊕ planet is approximately 2 K cooler than a 1 R ⊕ planet at a radiative flux of 1 S ⊕. At lower incident flux, this effect is especially significant: at 0.75 S ⊕ (approximately equivalent to the flux incident on Earth during the early history of the Solar System), the temperature difference between a 0.5 R ⊕ and 1 R ⊕ planet is closer to 8 K, and nearly 20 K between 0.5 R ⊕ and 2 R ⊕. To further illustrate this, if Mars was ocean-covered at Earth's current position, it would be in a Snowball state, even if the planet was allowed all biogeochemical cycling process available to Earth. This is partly due to the fact that the atmospheres of these smaller worlds are also proportionately depleted in carbon, which is removed at a greater rate (assuming burial continues up to the limit of the extinction of a C3 and C4 biosphere) due to the earlier cessation of geophysical activity that prevents the return of previously subducted carbon to the atmosphere. While not captured in this particular model, this process would be further exacerbated by the fact that atmospheres of smaller worlds are more rapidly stripped away by high-energy stellar particles. C3 plants on an Earth of half the radius, but at the same orbital separation, would become stressed by lack of atmospheric carbon ∼5.1 Gyr after planet formation. In terms of atmospheric carbon availability for photosynthesis habitability, planet size matters.
Applying this model to known exoplanets requires the assumption that these worlds will resemble Earth in many of their physical and chemical characteristics. At this stage, this assumption remains defensible, given the limited information available to astrobiologists on the interiors and atmospheres of small, potentially rocky worlds. Allowing this first-order assumption allows for re-evaluation of planetary habitability in some cases. For example, also plotted in Fig. 4 (red marker) alongside Earth is GJ 667Cc, a possibly terrestrial planet (R ⊕ = 1.54, S ⊕ = 0.88) discovered orbiting a nearby red dwarf star (Bonfils et al., 2013). The effective temperature for this planet, assuming an albedo of 0.3, is in the range of 227–247 K, depending on uncertainties in its orbital semimajor axis and stellar parameters. Adopting the flux and size estimates given above, and assuming an Earth-like carbonate-silicate cycle, ocean/continent fraction, and CO2 greenhouse, GJ 667Cc could maintain a significant, biogeochemically self-consistent CO2/H2O greenhouse (assuming the same relative initial conditions as Earth), with a predicted surface temperature buffered closer to 283 K. This value is significantly warmer than previous estimates, rendering this world potentially more “habitable” than previously considered, if the various “Earth-like” assumptions hold.
This case study can be taken a step further, should the relative age of a planet's star be known. GJ 667Cc orbits a small, red dwarf star (M ⊙ = 0.31) that has an estimated main sequence lifetime of 288 Gyr (Rushby et al., 2013), well in excess of the current age of the Universe. The implications of this long main sequence evolution are the possibility that GJ 667Cc may have a habitable period on the order of 65 Gyr, excluding the potentially deleterious effects of the frequent flaring to which these small stars are prone, as well as the effects of tidal locking and tidal heating, which may be considerable and increasingly important as internal heat sources diminish over time (Shields et al., 2016). However, the age of its host star is poorly constrained at >2 Gyr, and therefore any estimate of the current duration of habitable conditions on GJ 667Cc is subject to considerable uncertainty.
4. Discussion
This work presents a simple biogeochemical model that seeks to represent the evolution of the biosphere-planet system over geological and astronomical time, with particular focus on the carbonate-silicate cycle due to its significant effect on the surface, interior, and atmosphere of the planet. Larger planets tend, on average, to be warmer than smaller planets under a given stellar flux, with this effect especially pronounced at lower insolation when the atmospheric concentration and radiative effect of CO2 are more significant. By using temperature and CO2 concentrations as a first approximation of habitable conditions, it seems that the complex interplay between these components produces an uncertain conclusion: habitability is strongly variable in time and also falls along a spectrum as opposed to a clear binary distinction between “uninhabitable” and “habitable.” Conditions exist where, while average surface temperatures may fall within the traditional habitable boundaries, other factors act to reduce the overall habitability of the planet.
The upper temperature boundary considered here represents the endmember case for habitability, but before this limit is reached, the terrestrial biosphere will undergo significant and likely dramatic reorganization, in terms of spatiotemporal distribution, habitat and resource availability, and metabolic stress exacerbated by falling pCO2 levels, which results in a form of “photosystem-limited habitability” set by this CO2 compensation limit. The planet enters this state toward the end of its habitable lifetime as surface temperatures increase under a brightening star, and the action of terrestrial weathering draws down CO2 at a rate faster than it can be returned to the atmosphere by volcanic sources, the driver of which (geothermal heat flux) is also diminishing with time as the planet's radiogenic inventory becomes depleted. Primary producers crucial for supporting the planet's biosphere would become CO2-stressed, and this would eventually result in the collapse of the (terrestrial) biosphere long before the planet becomes too warm. O'Malley-James et al. (2014) described the changes likely to occur in the trophic hierarchy of the terrestrial biosphere of a planet in a state of CO2 stress: below 150 ppm CO2, desiccation-tolerant CAM and C4 plants would briefly dominate in isolated refugia (subterranean caves, high-altitude environments), possibly followed by a period of microbial anoxygenic photosynthesis when CO2 falls below 10 ppm. Below 1 ppm CO2, the end of all photosynthetic life is expected, and this juncture would represent a terminal extinction event for the biosphere, even if surface temperatures remained within habitable limits (O'Malley-James et al., 2014). It should be noted that we assume a “business as usual” scenario and that, given the timescales over which this CO2 decline takes place, it may be feasible, but outside the scope of this paper, to suggest that evolutionary adaptations may take place to extend the life span of a terrestrial photosynthetic biosphere to a low-CO2 world.
Furthermore, this work has demonstrated that a planet's size affects the maximum limit of habitable conditions through geophysical and atmospheric processes, one that can be, to some degree of accuracy, derived from the mechanics of carbon cycle as we currently understand it. Larger terrestrial planets are predicted to have warmer average surface temperatures and more carbon in their atmospheres through the control that planet mass has on various elements of the carbonate-silicate cycle. Planetary mass–dependent effects include a greater and more persistent geothermal flux that drives subduction and seafloor spreading at an accelerated rate, and for longer, on more massive terrestrial planets. This results in higher average pCO2 values for larger planets, which in turn generates a more effective greenhouse. However, the mass of a planet seems to have little effect on the eventual initiation of a runaway greenhouse event in this case; models of atmospheric photochemistry that take into consideration surface pressure and pressure broadening on larger worlds would likely suggest otherwise and would represent a potential future improvement to this particular model.
An implication of these results is that we are ∼70% of the way through the habitable lifetime of Earth (4.54 of 6.23 Gyr). We can begin to contrast and compare this value with that of known exoplanets, if the age of their star is known and the assumptions outlined above are adopted, to determine the comparative temporal habitability of worlds across the Galaxy. As demonstrated by the case of GJ 667Cc (a potentially rocky 1.54 R ⊕ planet in the orbit of a red dwarf binary), planets in the orbit of small stars with long main sequence lifetimes may considerably eclipse Earth in this measure. Despite the fact that this concept is often overlooked in studies of planetary habitability, the duration and relative timing of habitable conditions on terrestrial planets also have pressing implications for atmospheric evolution and future studies concerned with the spectroscopic detection of biosignature gases (Caldeira and Kasting, 1992; Watson, 2008; O'Malley-James et al., 2013, 2014; Rushby et al., 2013).
Furthermore, the nature of main sequence stellar evolution (i.e., growing in luminosity over time) also presents the possibility of a once-cold planet that formed beyond the outer edge of the habitable zone “entering” the habitable zone during the course of its host star's main sequence lifetime. Rushby et al. (2013) suggested that this may occur for Mars in our solar system but that it is likely too small and geologically inactive for habitable conditions to arise during the late stages of the Sun's main sequence lifetime. The possibility of the reactivation of a carbonate-silicate cycle on a planet undergoing secular stellar warming has not been studied in detail, but given that active plate tectonics is required to drive the subduction and outgassing of carbon from the planet's interior, it may not be possible to invoke this as a possibility. To further obfuscate the possibility of a late-stage reactivation of planetary carbonate-silicate cycling, the importance of mantle hydration as a mechanism for facilitating and maintaining plate tectonics over geological time remains unclear (Korenaga, 2010, 2012). It is possible that a drier mantle and lower associated viscosity will make the initiation of plate tectonics easier, but maintaining a continental crust may require interaction between the hydrosphere and solid Earth over geological time (Korenaga, 2010, 2012).
Attempting to quantify the likely lifetime of the biosphere of habitable planets has been a key area of research in astrobiology, and the concept remains extremely pertinent to astrobiology and SETI campaigns. Arguably, providing an estimate for the maximum life span of a planet's biosphere goes beyond a diagnostic investigation of the operation of feedback processes on the planet. This limit provides a terminal limit for biological evolution and speciation, with some bearing on certain parameters contained within the Drake equation, such as the fraction of life-bearing planets on which intelligent life emerges, as well as the duration of the civilizations formed by that life. Watson (2008), drawing on the earlier work of Carter (1983) and Szathmáry and Maynard-Smith (1995), developed a probabilistic model that attempts to quantify the timing and likelihood of several critical (defined broadly as only occurring once in Earth's history) steps in evolution en route to intelligent observer species. A key parameter in these calculations is the habitable period of the planet, which exhibits a strong control over the expected timing on these critical steps. Their broad conclusions are that only a planet with a long habitable period (>109 yr) could support complex life, and intelligent life may require longer still, as these critical steps are incredibly unlikely—on the order of 10−9 yr−1 (Watson, 2008). Therefore, both future SETI studies as well as those searching for “non-intelligent” life should prioritize exoplanets that satisfy both criteria of spatial and temporal habitability.
The idealized planetary system depicted here represents a first approximation of the geochemical cycling of carbon on a terrestrial world with a differentiated oceanic and continental crust. Assumptions implicit in the operation of the carbonate-silicate cycle are, however, a limitation to this work; it is not known at this stage whether analogous processes will operate on terrestrial exoplanets, even those with oceans and active tectonics, nor how planetary composition and density will scale with size and what effect the formation history of the system will have on the eventual composition of the planetary body. Furthermore, our understanding of the operation of this cycle on Earth reveals potential biological amplification and mediation at several junctures, including the fixation and burial of inorganic carbon in the ocean and the acceleration of mechanical erosion on the surface by the action of roots and exuded organic acids (Lenton and Watson, 2004; Mills et al., 2014). Additionally, plants return a large proportion of carbon-rich organic matter for burial upon their deaths, and throughout their life cycle represent a significant interface between the geosphere and biosphere. Furthermore, the marine ecosystem supported by cyanobacterial photoautotrophs will likely persist for some time following the extinction of the terrestrial biosphere. The carbon capture and concentration mechanisms of these organisms, as well as the behavior of CO2(aq), ensure that the oceanic carbon cycle will continue to operate, albeit under conditions of increasing stress and ecosystem reorganization. This is an area of study into which future iterations of this model will attempt to make ground. Separating biotic and abiotic processes implicit in the carbonate-silicate cycle and understanding their relative influence of the evolution of atmospheric CO2 therefore remain research priorities in Earth system and exoplanetary system science alike.
5. Conclusion
This study is arguably one of the first to attempt to incorporate a self-consistent carbon cycle and radiative-convective climate model to determine how planet size affects the long-term cycling of carbon, as well as how these differences may alter the surface temperature of potentially habitable terrestrial planets. Here, we demonstrate that both average planetary surface temperature and atmospheric carbon dioxide increase with increasing radius, and that we might expect a larger planet to be proportionately warmer and CO2-rich for a given incident flux. This model can also be used, in conjunction with stellar evolution data, to refine estimates of the duration of habitable conditions on a planet with similar carbonate-silicate cycling and compositional and geophysical characteristics to Earth, which is predicted to remain within habitable temperature bounds for approximately a further 1.7 Gyr. However, the abundance of pCO2 will decline secularly toward this limit, potentially starving terrestrial photosynthesizers of a suitable source of energy and limiting planetary habitability through their inability to support a complex eukaryote biosphere as primary producers.
Footnotes
Acknowledgments
A.J.R. would like to recognize the support of a Dean's Fellowship at the University of East Anglia, under which most of this work was produced, as well as an appointment to the NASA Postdoctoral Program at NASA Ames Research Center, administered by Universities Space Research Association under contract with NASA, where the model was updated and paper compiled. B.J.W.M. acknowledges a University of Leeds Academic Fellowship. A.J.R. would like to thank Christian Clanton and Jessica Lewis for their useful notes on the early manuscript, as well as the invaluable feedback from two anonymous reviewers that strengthened the final version.
Conflict of Interest
The authors report no conflicts of interest.