
Abstract
In the search for microfossils of early life on Earth, the demonstration of biogenicity is paramount. Traditionally, only syngenetic structures with cellular elaboration, hollow sheaths/cell walls, and indigenous kerogen have been considered bona fide fossils. Recent reports of inorganically preserved microfossils represent a shift from this practice. Such a shift, if accompanied by a robust set of biogenicity criteria, could have profound implications for the identification of biosignatures on early Earth and extraterrestrial bodies. Here, we reaffirm the conventional criteria by examining aggregates of inorganic filaments from the Pilbara region of Western Australia. These aggregates are preserved in bedded chert, and the filaments measure up to 1 μm in diameter and 100 μm in length. The aggregates superficially resemble kerogenous microbial fabrics and mycelial organisms. However, the filaments consist of manganese oxide, lack cellular elaboration, and show no evidence for hollow sheaths or cell walls. We conclude that the filaments are fibrous minerals of abiotic origin. The similarities between these pseudofossils and some filamentous fossils highlight the need for strict application of the conventional criteria for recognizing microfossils. In the absence of kerogen, morphologically simple structures should, at least, show evidence of cellular features to be considered bona fide fossils. Key Words: Fossil—Manganese oxide—Pilbara—Precambrian—Pseudofossil. Astrobiology 18, 249–258.
1. Introduction
B
Paleontologists traditionally use a set of stringent criteria that, when applied in hierarchical series, allows for the differentiation of fossils from pseudofossils (Hofmann, 1971, 1972, 2004; Muir et al., 1977; Muir, 1978; Schopf and Walter, 1983; Buick, 1990; Cady, 2000; Hofmann and Farmer, 2000; Brasier et al., 2002, 2005; Brasier et al., 2004, 2006; Schopf et al., 2005; Sugitani et al., 2007; Wacey, 2009). These criteria typically require that the microstructures in question occur in petrographic thin sections prepared from sedimentary or low-grade metasedimentary rocks, equal or exceed in size the smallest known microorganisms, and co-occur with other fossils of similar morphology (Buick, 1990). Carbonaceous materials are common components of fossils (Muscente et al., 2017a), and extracellular walls and sheaths are the microbial characters with the greatest potential for preservation (Knoll, 1985); so according to convention, true microfossils also possess kerogen, hollow sheaths/walls, and cellular elaboration (Buick, 1990). By design, these criteria establish a high burden of proof, which reduces the risk of false positives in the search for ancient life by ensuring that products of abiotic mineralization processes are not mistaken for biogenic structures (García-Ruiz et al., 2003, 2017).
The conventional criteria for differentiating fossils from pseudofossils do not adequately accommodate fossils that have been preserved (replicated by replacement) as oxide, carbonate, silicate, and other minerals devoid of carbonaceous material (Cady, 2000). These fossils, which occur in rocks of various ages (Hein et al., 2000; Kilias et al., 2007; Fernández-Remolar and Knoll, 2008; Ivarsson et al., 2010, 2015; Chi Fru et al., 2013), are preserved via complex taphonomic pathways involving authigenic and diagenetic mineralization (Muscente et al., 2015a, 2017b). Because mineral formation does not guarantee cellular replication or preservation of organic matter (i.e., carbon compounds produced or processed by life in the past), some mineralized fossils lack kerogen, hollow sheaths/walls, and/or cellular elaboration, and do not satisfy the conventional criteria. As a result, when it comes to such microfossils, the high burden of proof limits opportunity for scientific discovery in astrobiology.
Two end-member categories of microfossils can be envisioned on the basis of their preservational styles. “Organically preserved microfossils” are fossilized microorganisms, which are preserved as kerogen or other carbonaceous material. Such fossils have hollow lumina bounded by sheaths or cell walls (and may or may not have cellular elaboration). “Inorganically preserved microfossils,” in contrast, are microfossils composed of oxide, carbonate, silicate, and/or other minerals, which are devoid of carbonaceous material. Purely abiotic processes can generate microstructures that resemble mineralized microbes (García-Ruiz et al., 2003). These microstructures, however, do not represent remains or traces of life and should not be considered fossils, even if they contain carbonaceous material that condensed on the structures after their formation (García-Ruiz et al., 2003). Instead, these microstructures should be considered as “pseudofossils.” Distinguishing “inorganically preserved fossils” from these abiotically produced pseudofossils presents a significant challenge. In many cases, such a distinction cannot be made with confidence, and the problematic microstructures are termed “dubiofossils” (Hofmann, 1972; Sugitani et al., 2007).
Recent works describing inorganically preserved microfossils in Archean (Dodd et al., 2017) and Paleoproterozoic (Bengtson et al., 2017) rocks are suggestive of a shift in thinking, deemphasizing the importance of kerogen, in the search for fossils of simple microbial organisms. This shift from organic to inorganic fossils may ultimately broaden our vision in the search of early terrestrial and extraterrestrial life, but it also introduces a risk of false positives. Safe to say, it should be accompanied by corresponding advancement in methods and criteria for recognizing bona fide microfossils, lest it result in false positives detrimental to astrobiology as a whole. Although methods development remains an active area of research in Precambrian paleontology, most work on this topic focuses on organic, rather than inorganic, microstructures and on determining whether or not they are fossils (Schopf et al., 2002, 2005; Schiffbauer et al., 2012). Developing new approaches to distinguishing inorganically preserved microfossils from pseudofossils should be a top priority, as this pursuit has implications for searching for ancient life on Earth and throughout the Solar System (Cady, 2000).
Of all inorganically preserved fossils and biogenic structures, those preserved as oxides may have the most promise in astrobiology. Microscopic and macroscopic organisms preserved as iron and manganese oxides affirm that oxide mineral formation can replicate morphology and record biosignatures (Hein et al., 2000; Kilias et al., 2007; Fernández-Remolar and Knoll, 2008; Ivarsson et al. 2010, 2015; Chi Fru et al., 2013; Zhou et al., 2015). Notably, this process occurs in environments where hydrothermal vents represent the source of iron and manganese (Hein et al., 2000; Kilias et al., 2007; Chi Fru et al., 2013). Given that life may have evolved in hydrothermal systems (Martin et al., 2008) and ancient sedimentary rocks of hydrothermal origin may contain the oldest fossils on Earth (Dodd et al., 2017), oxide minerals in such environments throughout the Solar System represent prime targets in the search for extraterrestrial life (Vance et al., 2007; McKay et al., 2008; Waite et al., 2017). Future Mars missions—particularly, potential sample return missions—could also focus on oxide minerals. Rover data indicate that martian sedimentary rocks have a low potential for preservation of organic matter (Sumner, 2004), which does not bode well for the existence of organically preserved microfossils. Iron and manganese minerals, however, occur broadly on the martian surface (Lanza et al., 2016) and, from a practical standpoint, represent convenient and appealing targets in the search for biogenic structures. Indeed, martian hematite concretions may represent products of microbial processes and could conceivably contain physical remains of microorganisms (Squyres et al., 2004). Of course, the merit of pursuing research on such minerals as well as their potential value to astrobiology ultimately depends upon the robustness of obtainable data as evidence of life.
Several aspects of manganese make its oxides an attractive target for astrobiological research. First, manganese is the third most common heavy metal behind iron and titanium (Post, 1999), and given its occurrence on Mars (Lanza et al., 2016), its oxides represent a practical focus for investigations of rocky planets and moons. Second, like iron, Mn2+ is sourced from hydrothermal vents albeit at variable concentrations (Von Damm, 1990), and although the kinetic limits imposed by redox conditions are greater for manganese than iron oxidation (Canet et al., 2005), manganese oxides are common in both deep (Marchig et al., 1999) and shallow (Canet et al., 2005; Fitzgerald and Gillis, 2006) hydrothermal systems. Lastly, certain groups of bacteria (Butterfield et al., 2013) and fungi (Tang et al., 2013) mediate oxidation of Mn(II) to Mn(III/IV), resulting in formation of Mn(III/IV) oxides on their external surfaces. In addition, manganese oxide formation can lead to fossil preservation by means of mineralization of shells (Hein et al., 2000) and potentially microbes as well (Kilias et al., 2007; Ivarsson et al. 2010, 2015). Thus, multiple pathways exist for the preservation of fossils in association with manganese oxide minerals.
In this report, we investigate filamentous manganese oxides with the goals of (1) sharpening the astrobiological toolkit employed in exploring oxide minerals for evidence of ancient life and (2) developing approaches to differentiating inorganically preserved microfossils from pseudofossils. Our work focuses on filamentous manganese oxide structures from the “Pilbara Manganese Province” of Western Australia (De la Hunty, 1963). By providing data on these microstructures from a variety of analytical techniques, we highlight the plethora of instruments available for studying manganese oxide minerals and draw attention to the conventional criteria for recognizing microfossils. The structures share similarities with proposed microbial fabrics (Schopf et al., 2015) and putative anastomosing mycelia-like fossils (Bengtson et al., 2017) in Paleoproterozoic rocks, in that they sometimes resemble cobweblike aggregates of slender threadlike filaments. Even so, these similarities do not guarantee the biogenicity of any of the structures, which were preserved via different pathways. On the contrary, the similarities highlight the need for robust recognition criteria—the presence of cellular elaboration and hollow sheaths/cell walls—and the potential for false positives when these criteria are not satisfied. In this context, our work on filamentous oxide minerals highlights the urgency of developing and ground-truthing criteria for differentiating inorganically preserved fossils from pseudofossils.
2. Geological Context, Materials, and Methods
The sample in this study is reposited in the Virginia Tech Department of Geosciences and was collected by S. Xiao on the 2012 International Geological Congress field trip WA-2 as a float near the “Awramik locality” (21.067116°S, 119.411341°E)—the Awramik State Geoheritage Reserve R44711 (Grey et al., 2010)—in North Pole Dome of the Pilbara region (Supplementary Fig. S1; Supplementary Information are available online at
Billets of the sample were cut, polished, and mounted to petrographic glass slides to make 30 μm thick petrographic thin sections. Thin sections were examined via transmitted light microscopy and transmitted light photomicrographs of the filaments and associated minerals were collected. In addition, two- and three-dimensional images of selected filaments were acquired by confocal laser scanning microscopy (CLSM) using an Olympus FV1200 microscope (Olympus, Inc., Shinjuku, Japan) housed at the University of Cincinnati. The system used a 488 nm laser, a 60× oil immersion lens (NA = 1.42), and a confocal aperture of 65 μm. The filaments fluoresced weakly with 488 nm excitation. Both fluorescence and reflected light images were produced, but the reflected light images were superior due to the weak fluorescence of the material (cf. Czaja et al., 2016). Two-dimensional images were rendered by using the Fluoview software package FV10-ASW (v.3.01; Olympus, Inc., Shinjuku, Japan). Three-dimensional images and animations (Supplementary Animation S1) were created and deconvoluted to remove optical and confocal artifacts by using the software package Huygens Essentials version 17.04 (Scientific Volume Imaging, The Netherlands) (cf. Schopf and Kudryavtsev, 2009).
Raman spectra of the filaments were collected with a Horiba T64000 system (Horiba, Inc., Edison, NJ) attached to an Olympus BX41 microscope at the University of Cincinnati. Spectra were acquired with a 50× long working distance objective (NA = 0.50) and 514.5 nm excitation from an Innova 90CFreD Ar+ laser (Coherent, Inc., CA). Each spectrum was accumulated for 30 s to achieve an acceptable signal-to-noise ratio. Data were collected and analyzed with the LabSpec software package (v.5; Horiba, Inc., Edison, NJ). No processing of the spectra was required.
The thin sections were also imaged and elementally mapped by using two scanning electron microscopy (SEM) systems: (1) a Hitachi TM3000 tabletop thermionic (tungsten filament source) scanning electron microscope with polepiece backscattered electron (BSE) silicon drift detector (SDD) and Bruker XFlash compact energy-dispersive X-ray spectroscopy (EDS) detector housed in the Virginia Tech Department of Geosciences and (2) a FEI Quanta 600F low vacuum environmental scanning electron microscope with a field emission gun electron source, polepiece BSE SDD, and BSE and secondary electron (SE) Everhart–Thornley detector (ETD) housed in the Virginia Tech Institute of Critical Technology and Applied Science Nanoscale Characterization and Fabrication Laboratory (see Muscente and Xiao, 2015, for additional technical details on this equipment). Images of the thin sections, which were not coated prior to SEM and EDS analyses, were acquired at 12 mm working distance and at various accelerating voltages between 15 and 30 keV. Two types of images were acquired: BSE images with high compositional contrast based on atomic number (Z) were acquired by the SDDs on both systems, and SE images with high topographic contrast were acquired by the ETD on the Quanta 600F system. EDS elemental mapping and semiquantitative point spectra of elemental concentration analysis were conducted at 15 keV, and the scan live times were 400–500 s with ∼8000 counts/s. Given the operating conditions, the tungsten filament electron source, and the count rate, the dead time should be within an acceptable range (∼10–40% live time) for elemental mapping analysis. The EDS data were processed with the Quantax 70 Micro-analysis Software by BrukerTM, designed for the Hitachi TM3000 scanning electron microscope.
We further examined the ultrastructure and composition of the filaments using bright field (BF) transmission electron microscopy (TEM), BF scanning transmission electron microscopy (STEM), and high-resolution TEM-based EDS. For these analyses, we prepared an in situ cross section through an aggregate of filaments (Supplementary Animation S2) as an ultrathin (∼100 nm thick) foil liftout using dual-beam focused ion beam electron microscopy (FIB-EM), following procedures used in previous studies of filamentous microstructures (Muscente et al., 2015b). All FIB-EM work was conducted using a FEI DualBeam Helios 600 NanoLab equipped with a field-emission gun electron source, ETD, STEM detector, and focused Ga+ ion beam column for controlled, nanoscale material addition and excavation (Schiffbauer and Xiao, 2009). An aggregate of filaments, located at and below a polished surface of a thin section, was targeted for milling. Prior to preparation of the ultrathin foil, the thin section was coated with 10 nm thick Pt–Pd, and a thin (∼1 μm thick) layer of Pt was deposited on the surface of the thin section above the aggregate. Milling of the thin section was then conducted with the Ga+ ion beam at 30 kV accelerating voltage. The working distance was adjusted periodically, and throughout the FIB-EM work, SE images were acquired by using the ETD at 5 keV accelerating voltage (Muscente and Xiao, 2015). After the cross section was exposed via milling, it was removed from the slide with a micromanipulator probe, secured to a copper TEM grid, and ion polished, producing a ∼100 nm thick ultrathin foil for TEM and STEM imaging. The BF-TEM, BF-STEM, and associated EDS analyses of the ultrathin foil were conducted at 200 keV using a JEOL 2100 TEM with thermionic source and large solid angle EDS detector, and the resulting data were processed with the JEOL Analysis Program software package. All TEM-based EDS elemental maps were acquired at 200 keV with a scan time of ∼26 min (dwell time, 2 ms; resolution, 1024 × 768 pixels) and were processed with the JEOL AnalysisStation software package.
3. Results
The sample consists of ferromanganiferous chert bedded with ferruginous layers (Fig. 1a, 1b). Ferruginous layers primarily consist of iron oxide minerals (hematite and goethite) and bedded siderite, which in some places contain void-filling siderite, calcite, and manganese oxide cements. Chert layers or bands similarly contain void-filling iron oxides (Figs. S2a–S2g, S3a–S3t) and manganese oxides (Fig. S2d–S2j, S3f–S3y) with botryoidal siderite cements (Figs. S4a–S4f, S4j–S4n, S5) in addition to multiple generations of void-filling and crosscutting calcitic and siliceous cements (Figs. S2–S4), including coarse-grained crystalline and fine-grained microcrystalline quartz (Fig. 1e, 1f). Some of the calcite, which may represent primary or early diagenetic sedimentary carbonate, is rimmed by manganese oxides and surrounded by microcrystalline quartz and later calcitic cements (Figs. 1c, 1d, 2a). Iron oxides are generally surrounded by microcrystalline quartz and void-filling carbonates (Figs. S2e, S3k–S3o) but in some places have well-developed rims, representing admixtures of iron oxides, manganese oxides, calcite, and silica (Figs. S2f, S2g, S3p–S3t). Manganese oxides occur as textureless masses (Fig. S2a, S2c) and masses of fibers with curved and branching microstructures (Fig. S2j).

Light and confocal laser scanning microscopy. (

Electron microscopy and EDS. (
Void-filling calcite (Figs. 1c, 1d, 2a), coarse-grained crystalline quartz (Fig. 1e–1h), and fine-grained microcrystalline quartz (Fig. 1e–1h) contain “cobweblike” aggregates of filaments (30 nm to 1 μm in diameter; up to 100 μm in length), which are rarely found in isolation. These aggregates range from tens (Fig. 1i) to hundreds (Fig. 1g) of micrometers in thickness and occur abundantly in chert crosscut by generations of coarse-grained quartz (Fig. 1g, 1h). In some places, they encrust botryoidal siderite (Fig. S4c–S4f) and primary or early diagenetic carbonate (Figs. 1c, 1d, 2a) and are truncated by euhedral crystals (Fig. S4g–S4i), which have been replaced by quartz but may have originally been barite.
Observed in plain light images of petrographic thin sections, the individual filaments appear opaque to slightly translucent compared to surrounding transparent siliceous and calcareous minerals of the matrix. They resemble curved, threadlike fibers (Fig. 1i–1k) and appear with possible branching configurations (Fig. 1j) in random orientations (Fig. 1i–1k), which resemble those of the fibers in the manganese oxide masses (Fig. S2j). However, we did not observe any branching structures in electron image or in three-dimensional renderings of the structures generated via CLSM, and furthermore, the CLSM images show that the filaments in the aggregates have roughly parallel orientations (Animation S1). Compositional contrast in BSE-SEM images indicates that the filaments consist of a material of higher Z than calcite (Fig. 2a) or silica (Fig. 2b). A representative Raman spectrum (Fig. 3) of a filament surrounded by chert (Fig. 1g) contains a major peak for quartz at 463 cm−1 and other minor quartz peaks (Kingma and Hemley, 1994) as well as doublet peaks for manganese oxide, possibly todorokite, at 598 and 630 cm−1 (Julien et al., 2003; Laetsch and Downs, 2006).
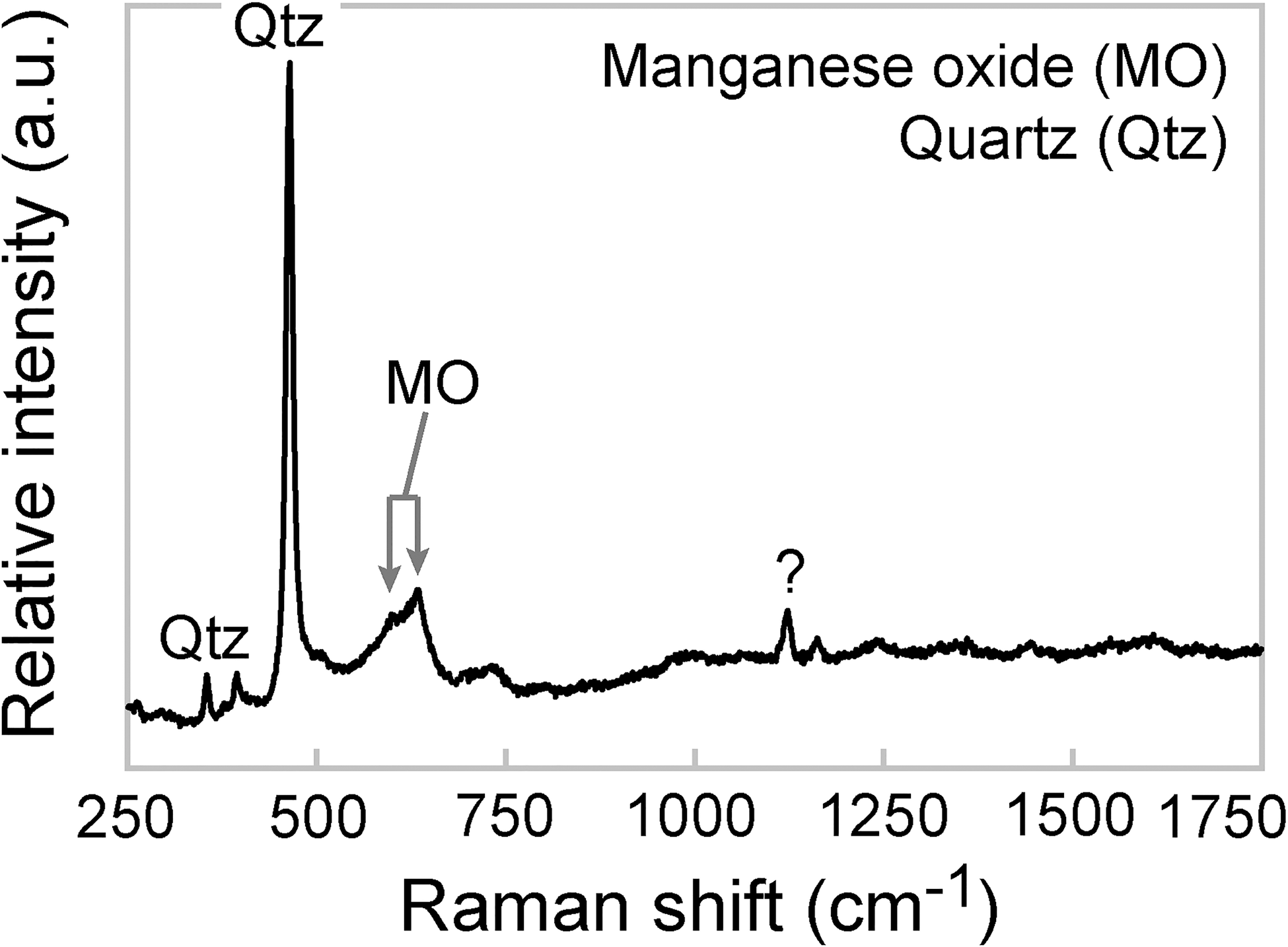
Representative Raman spectrum of a filament. The filament is composed of manganese oxide (MO), likely todorokite, and is embedded in a quartz matrix (Qtz). One minor peak near ∼1120 cm−1 (labeled with a question mark) has not been identified. The spectrum was acquired from the point indicated by the arrow in (g) of Fig. 1.
Preparation of the sample with the FIB (Fig. S6) resulted in damage to the filaments and significant redeposition of milled material. Nonetheless, the FIB work facilitated preparation of an ultrathin foil containing both transverse and longitudinal sections of filaments (Fig. 2c), which, evidently, consist of stacked layers of crystalline material, range in thickness between tens and hundreds of nanometers (Fig. 2d–2i), and are crosscut by microcrystalline quartz grain boundaries (Fig. 2d). Mass-thickness contrast in BF-STEM (Fig. 2c) and BF-TEM (Fig. 2d–2i) images of the cross sections affirms that the filaments consist of a higher-Z material than silica. No other materials were observed in the ultrathin foil, except for a globular platinum-rich material that was redeposited during milling and concentrated around the filaments (Fig. 2f). Data from TEM-based EDS analyses indicate that the filament material consists primarily of manganese and oxygen (Fig. 2j–2m). Small carbon and platinum peaks were measured in the filaments, but these signify surface contaminants that became concentrated on the structures via sputtered material redeposition. Silicon was also recorded, though this element signal likely emanates from surrounding and permeating quartz. Evidently, as shown in BF-TEM images, the manganese oxide did not form within cellular walls, like pyrite found within bona fide microfossils of the Strelley Pool Formation (Wacey et al., 2011). Instead, the filaments are comprised entirely of the mineral, which, like many manganese oxides (Ostwald 1986; Julien et al., 2003; Xu et al., 2010), exhibits ultrastructural layering (the filaments do not represent individual crystals, but rather, stacks of crystalline material).
4. Discussion and Conclusions
Overall, the multiple textural, geochemical, and mineralogical lines of evidence presented here indicate that the aggregate structures in the quartz and calcite cements of the Pilbara sample consist of nonrandomly oriented manganese oxide filaments. Based on the paragenesis of the minerals, we infer a hydrothermal (metasomatic) rather than a weathering origin for the filament aggregates. Ferromanganiferous cherts and ferruginous layers were deposited in alternation, and calcite, silica, and iron and manganese oxides precipitated within cracks and voids in the beds. The filaments, evidently, formed and grew on preexisting surfaces, such as siderite botryoids and sedimentary/diagenetic carbonates, indicating that they postdate those minerals. However, unlike manganese oxides that form as surface weathering products (Xu et al., 2010), the filaments did not form and grow on fracture surfaces and bedding planes, like manganese dendrites. Crosscutting relationships, instead, provide evidence for the antiquity of the structures. Various minerals grew after the filaments, including euhedral (possibly barite) crystals that truncated the filaments as well as the calcite and quartz matrixes that formed around the aggregates. The aggregates are crosscut by boundaries of grains in fine-grained microcrystalline quartz (Fig. 2d) and later-formed, fabric-destructive coarse-grained quartz (Fig. 1g, 1h). Given that the chert bands consist of filament-containing microcrystalline quartz, the petrographic data indicate that at least some of the filaments formed prior to, or concomitantly with, the surrounding silica layers, perhaps under the influence of hydrothermal metasomatism. The iron oxides, indeed, possess well-developed rims composed of admixtures of calcite, silica, and iron and manganese oxides, which may have formed via simultaneous solution and deposition of the minerals. Taking into consideration that todorokite can form as a result of processing of other manganese-bearing species (Finkelman et al. 1974), the filaments may similarly represent alteration products resulting from dissolution and reprecipitation of the manganese oxides occurring as textureless masses. In any case, the paragenesis suggests that at least some of the filaments predate the chert bands and therefore most likely represent syngenetic features, as opposed to Recent or ancient weathering products.
Because our sample was collected from float, it is impossible to know its origin. The presence of manganese oxides in our sample strongly suggests that the rock was deposited following the Paleoproterozoic oxygenation event (Komiya et al., 2008). The main manganiferous unit in the region is the Mesoproterozoic Manganese Group, which outcrops several hundred kilometers to the east of the North Pole area (Jones et al., 2013; Jones, 2017). Additionally, several hundred kilometers to the southwest, the Paleoproterozoic Turee Creek Group contains Mn-rich (7.4 wt % MnO) banded iron formation with siderite and chert (Williford et al., 2011). In general, the manganese deposits of the Manganese Group are hydrothermal in origin with a late supergene overprint (Ostwald, 1993; Jones et al., 2013). The banded manganese deposits of the Turee Creek Group have received less attention, but geochemical data from authigenic pyrites in the Turee Creek Group are suggestive of hydrothermal fluid (Williford et al., 2011), so the deposits may similarly reflect hydrothermal input of manganese.
Because they contain no kerogen and exhibit no evidence of cellular elaboration or hollow sheaths/cell walls, and TEM analyses show that some of the microstructures are much smaller (∼30 nm in diameter) than the smallest microorganisms alive today, we interpret the filaments described herein as abiotic mineralogical pseudofossils, not fossils. In general, fibrous and asbestiform forms of manganese oxides, like todorokite, are common in nature (Finkelman et al. 1974; Ostwald 1986; Post and Bish 1988; Mohapatra et al. 1996; Gutzmer and Beukes, 2000; Xu et al., 2010; Jones et al., 2011; Johnson et al., 2016), and their formation does not require biological influence, as demonstrated by hydrothermal synthesis of manganiferous nanowires in autoclaves (Wei et al., 2005; Teng, 2009). Thus, by themselves, the filaments in our sample do not represent compelling evidence of life.
The filament aggregates notably resemble putative anastomosing fossils preserved as calcite/phyllosilicate minerals in a basalt of the ∼2.4 Ga Ongeluk Formation in South Africa (Bengtson et al., 2017) as well as kerogenous microbial fabrics within cherts of the ∼1.8 Ga Duck Creek Formation and ∼2.3 Ga Turee Creek Group in Western Australia (Schopf et al., 2015), interpreted as sulfur-cycling fossil communities (Schopf et al., 2015) on the basis of similarities to modern analogues (Gallardo and Espinoza, 2007). All three of these examples consist of slender threadlike filaments. The largest filaments in this study are close in diameter to the smallest filaments described from the kerogenous microbial fabrics (∼1 μm) and are generally thinner than the mycelia-like filaments (∼2–3 μm) in the Ongeluk basalt. Nonetheless, in all these cases, filaments occur in “cobweblike” aggregates. The morphological similarities among the structures as well as their broad similarities to modern fungi and microbial mats, however, are insufficient support of biogenicity. On the contrary, the superficial similarities among the compositionally distinct structures highlight the potential for false positives in this work. To deal with this issue, astrobiology and Precambrian paleobiology must compel a hefty burden of proof and encourage rigorous application of recognition criteria to new material. In this light, only the Duck Creek and Turee Creek microbial fabrics, which consist of kerogen and exhibit an array of cellular morphologies (Schopf et al., 2015), pass the biogenicity test. Future work should aim to put forth innovative recognition criteria for identifying inorganically preserved microfossils. In the meantime, cellular structure and size should be considered essential criteria when organic matter is not preserved, as these features are known in bona fide fossils conserved in oxides (Fernández-Remolar and Knoll, 2008).
Footnotes
Acknowledgments
The research was supported by NASA Exobiology and Evolutionary Biology Program (NNX15AL27G). A.D. Muscente is supported by the Keck Foundation–funded project “The Co-Evolution of the Geo- and Biospheres: An Integrated Program for Data-Driven Abductive Discovery in Earth Sciences.” The sample was collected on a 2012 International Geological Congress field trip led by Arthur Hickman and Martin van Kranendonk. We thank Dr. Hickman and Dr. Charlotte Hall of the GSWA for their assistance in facilitating our sampling near the Awramik State Geoheritage Reserve R44711. We additionally thank two anonymous reviewers for feedback, which greatly helped us improve the quality of this report.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.