
Abstract
Submarine hydrothermal vents are inhabited by a variety of microorganisms capable of tolerating environmental extremes, making them ideal candidates to further expand our knowledge of the limitations for terrestrial life, including their ability to survive the exposure of spaceflight-relevant conditions. The spore resistance of two Bacillus spp. strains, APA and SBP3, isolated from two shallow vents off Panarea Island (Aeolian Islands, Italy), to artificial and environmental stressors (i.e., UVC radiation, X-rays, heat, space vacuum, hydrogen peroxide [H2O2], and low-pressure plasma), was compared with that of two close phylogenetic relatives (Bacillus horneckiae and Bacillus oceanisediminis). Additional comparisons were made with Bacillus sp. isolated from spacecraft assembly facilities (B. horneckiae, Bacillus pumilus SAFR-032, and Bacillus nealsonii) and the biodosimetry strain and space microbiology model organism Bacillus subtilis. Overall, a high degree of spore resistance to stressors was observed for the strains isolated from spacecraft assembly facilities, with an exceptional level of resistance seen by B. pumilus SAFR-032. The environmental isolate SBP3 showed a more robust spore resistance to UVC, X-rays, H2O2, dry heat, and space vacuum than the closely related B. horneckiae. Both strains (SBP3 and APA) were more thermotolerant than their relatives, B. horneckiae and B. oceanisediminis, respectively. SBP3 may have a novel use as a bacterial model organism for future interrogations into the potential of forward contamination in extraterrestrial environments (e.g., icy moons of Jupiter or Saturn), spacecraft sterilization and, broadly, microbial responses to spaceflight-relevant environmental stressors.
1. Introduction
M
2. Materials and Methods
2.1. Bacterial strains and culture conditions
Environmental samples of sediment and thermal fluid were collected by SCUBA divers in the immediate vicinity of two shallow submarine hydrothermal vents, namely Black Point (latitude 38°38′23″N, longitude 15°06′28″E, depth 23 m) and Campo 7 (latitude 38°37′59″N, longitude 15°06′59″E, 21.3 m depth), located off the Panarea Island (Aeolian Islands, Italy) (Fig. 1). Strain SBP3 was isolated from a sediment sample taken at Black Point and strain APA from a fluid sample emitted at Campo 7 (Gugliandolo et al., 2012). Samples of sediment were collected directly in the immediate vicinity of the emissions by using sterile polycarbonate tubes (Lentini et al., 2014). To minimize the potential contamination by seawater, hydrothermal fluid sampling was carried out by using sterile syringes through special stainless steel probes, which were inserted into the emission orifice (Gugliandolo et al., 1999). At the sampling sites, characteristics of the thermal fluids emitted from Black Point and Campo 7 were temperature 130°C and 60°C, pH 3.0 and 4.92, and conductivity 46.20 and 49.2 mS/cm, respectively (for more details on physical and chemical characteristics of thermal fluids and gas composition, see Maugeri et al., 2009, 2010).
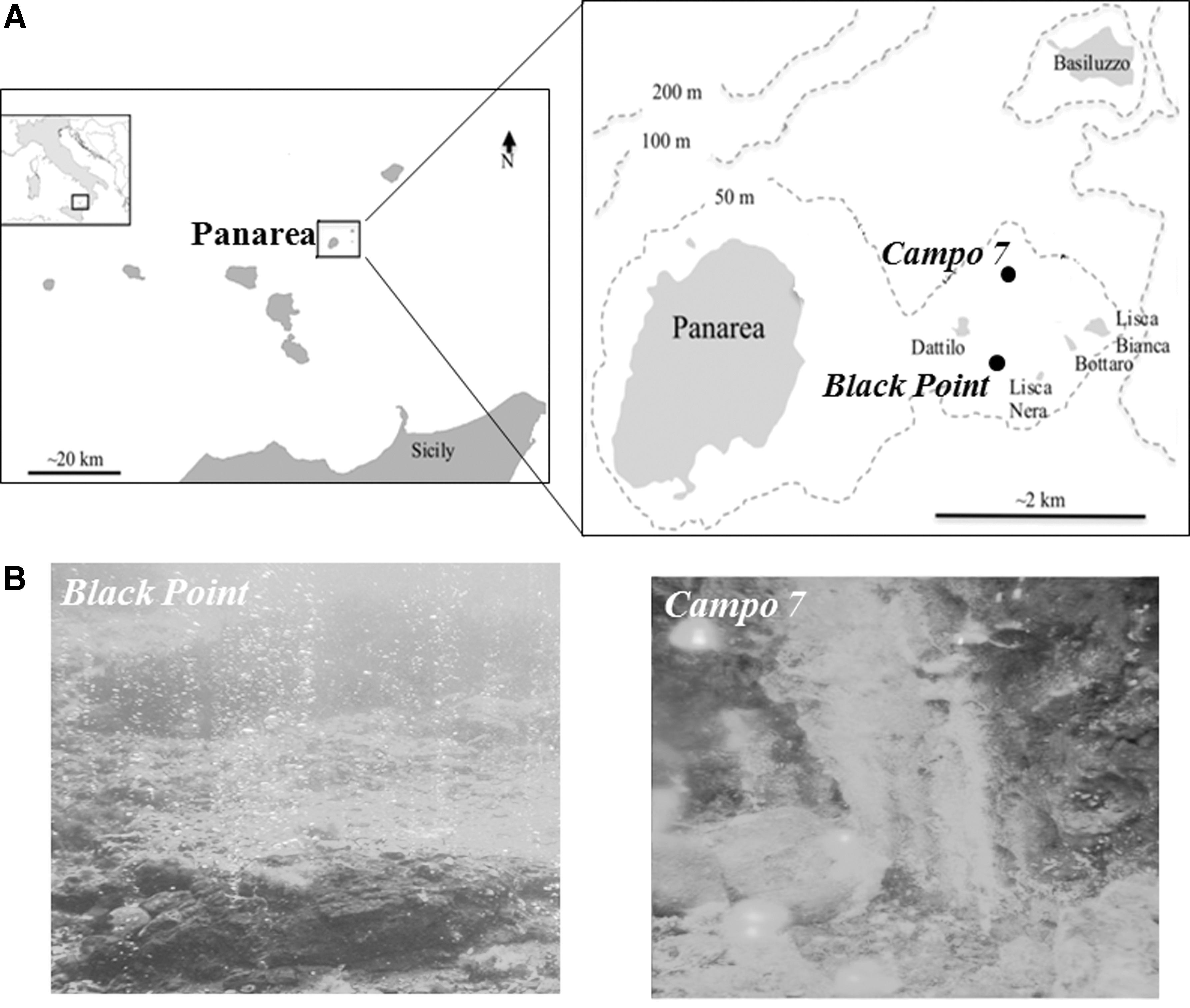
Sampling sites' location (
2.2. Bacterial strain characterization
The isolates were investigated regarding cell morphology and spore production, and tested for oxidase and catalase activity. Temperature and pH range for optimal growth conditions were determined following incubation of the strains for 3 days at 4°C, 37°C, 40°C, 45°C, 50°C, 55°C, 60°C, and 70°C and pH 5.5, 6, 7, 8, and 9 in tryptone soy broth (TSB; Oxoid) at a final concentration of 1.5% NaCl (TSB1.5). Halotolerance was tested after incubation in Bacto nutrient broth (NB; Difco) supplemented with 0%, 2%, 3%, 5%, 6%, and 10% (w/v) NaCl. Optimal growth was evaluated by measuring changes in turbidity at 600 nm with a spectrophotometer (Ultraspec 3000; Amersham Pharmacia Biotech, Freiburg, Germany). Sensitivity to the following antibiotics, chloramphenicol (30 μg), nalidixic acid (30 μg), novobiocin (30 μg), and polymyxin B (300 U), was tested on TSB agar plates. Fatty acids were analyzed as methyl ester derivatives, as described by Kuykendall et al. (1988), and by the Miller method (Miller, 1982). The fatty acid methyl esters were separated and identified with the Microbial Identification System (MIDI) Sherlock version 6.1 (TSBA40 database). Polymerase chain reaction amplification (with the primers: 519F and 926R) and sequencing of the 16S rRNA gene were done at Selekt-ID Biolabs GmbH (Berlin). All sequences were aligned manually with Bioedit (Hall et al., 1999) and MEGA 7 (Kumar et al., 2016) was used for construction of the phylogenetic tree.
2.3. Spore resistance assays
The bacterial strains used are listed in Table 1. Bacterial spores for the isolates were obtained by incubation in liquid Schaeffer Sporulation Medium (containing 0.1% KCl, 0.012% MgCl2, 0.5 mM CaCl2, 0.01 mM MnCl2, 0.001 mM FeSO4, and 8 g/L NB) at 37°C for 72 h in a rotary shaker with vigorous aeration (Link et al., 2004; Nagler et al., 2014). Spore preparations were determined to be essentially free of vegetative cells and to consist of >99% phase-bright spores by phase-contrast microscopy (Nagler et al., 2014) (Fig. 2). Harvested spores were washed 10 times and resuspended in sterile distilled water before storage at 4°C. The level of spore resistance of the Bacillus strains to UVC radiation, X-ray irradiation, H2O2, ultrahigh vacuum (UHV), low-pressure argon plasma, and dry and wet heat exposure was determined as described previously (Moeller et al., 2007, 2012, 2014). In short, either air-dried spore monolayers (106 spores) immobilized on 7-mm-diameter steel disks or spores in aqueous suspension (107 spores/mL) were used for the spore resistance assays. The effects of, and resistance to, UVC radiation on air-dried spore monolayers and spores in suspension were determined as previously described (Riesenman and Nicholson, 2000; Moeller et al., 2007) by using a commercial low-pressure mercury arc lamp (NN 8/15; Heraeus, Berlin, Germany), which emits monochromatic 254-nm UV radiation. For studying the effects of ionizing radiation on spore survival, X-rays (150 keV/20 mA) were generated by an X-ray tube (Gulmay Medical RS225; X-Strahl, Surrey, United Kingdom) (Moeller et al., 2007, 2014). Air-dried spore monolayers were exposed to different doses of X-rays as described previously (Moeller et al., 2014). To measure spore resistance to H2O2, aliquots (50 μL) of spore suspensions (107 spores/mL) in sterile phosphate-buffered saline (PBS) were placed in Eppendorf tubes, together with 500 μL of 10% (v/v) H2O2 solution (Merck KGaA, Darmstadt, Germany). After incubation at room temperature (20°C ± 2°C), 100 μL of each sample was removed at various times and immediately diluted 1:10 with a solution of bovine catalase (100 μg/mL in PBS; Sigma-Aldrich, St Louis, MO) for the purpose of neutralizing the H2O2 (Riesenman and Nicholson, 2000; Moeller et al., 2012). To determine the resistance of air-dried spore monolayers to LPP, a double inductively coupled LPP reactor with argon as process gas (flow rate: 100 sccm, pressure of 10 Pa, and power of 500 Watt) was used as described in Halfmann et al. (2007) and Raguse et al. (2016). Resistance of water-suspended spores to heat treatment (100°C) was measured as previously described by Melly et al. (2002) and Moeller et al. (2012). Here, spore suspensions (107 colony forming unit (CFU)/mL) were placed in Eppendorf tubes and exposed to 100°C for various times (i.e., 0, 1, 2, 4, and 6 min). To measure spore resistance to dry heat stress, air-dried spore samples were exposed to 120°C for different times (i.e., 0, 2, 4, 8, 12, and 16 min), as described in Moeller et al. (2012). For studying the effect of vacuum-induced extreme desiccation, air-dried spore monolayers were exposed for 7 days to UHV produced by an ion-getter pumping system (400 L/s; Varian SpA, Torino, Italy) reaching a final pressure of 10−7 Pa (Moeller et al., 2007). Spore recovery and viability assays were performed following the protocol described in Moeller et al. (2007). In brief, to recover the spores from the steel disks after treatments, the spore monolayers were covered by a 10% aqueous polyvinyl alcohol (PVA) solution, and after air-drying, the spore–PVA layer was stripped off as described previously (Moeller et al., 2007). Spore survival was determined by plating serial dilutions, prepared in sterile distilled water, for CFUs occurring after overnight growth on solid LB or tryptone soy agar medium incubated at the optimal temperature for each strain (Table 1). The surviving fraction was determined from the quotient N/N
0, with N being the number of CFUs of the treated sample and N
0 the CFUs of the nontreated controls. By plotting the logarithm of N/N
0 as a function of each treatment, survival curves were obtained (for details see Supplementary Figs. S1–S14; Supplementary Data are available at https:

Phase-contrast microscopic images of spores of Bacillus subtilis 168
DSMZ = German Collection of Microorganisms and Cell Cultures GmbH (Braunschweig, Germany).
LB = Luria Bertani; NB = nutrient broth; TSA = tryptone soy agar; TSB = tryptone soy broth.
3. Results
3.1. Bacterial strains
Cells of strains SBP3 and APA were found to be Gram-positive, endospore-forming, and rod-shaped bacteria as determined by standard microbial staining and microscopy methods (Venkateswaran et al., 2003; Vaishampayan et al., 2010; Zhang et al., 2010). They were deposited at the German Collection of Microorganisms and Cell Cultures (DSMZ) and have been assigned numbers DSM 103062 (APA) and DSM 103063 (SBP3), respectively. Phylogenetic analysis (Fig. 3) showed that strain SBP3 was strictly related to B. horneckiae (DSM 23495T), whereas strain APA was closely related to Bacillus oceanisediminis (DSM 24771T). The strains showed a 16S rRNA gene sequence similarity value of 99.86% and 99.5% to its nearest phylogenetic neighbor, respectively (Fig. 3). Differential phenotypic characteristics of strains APA and SBP3 in comparison with those of their closest related type strains are reported in Table 2. SBP3 spores exhibited the highest optimal growth temperature (45°C) and favored more alkaline conditions than spores of strain APA and their closest relative strains. Furthermore, both SBP3 and APA spores were more sensitive to lower temperatures, showing no growth at temperatures <25°C, whereas both B. horneckiae and B. oceanisediminis strains grew at temperatures as low as 4°C. APA spores displayed greater sensitivity to antibiotic treatments (e.g., chloramphenicol and novobiocin) than B. oceanisediminis and strain SBP3. Fatty acid percentages were highly concordant with environmental conditions, namely temperature (Table 2).
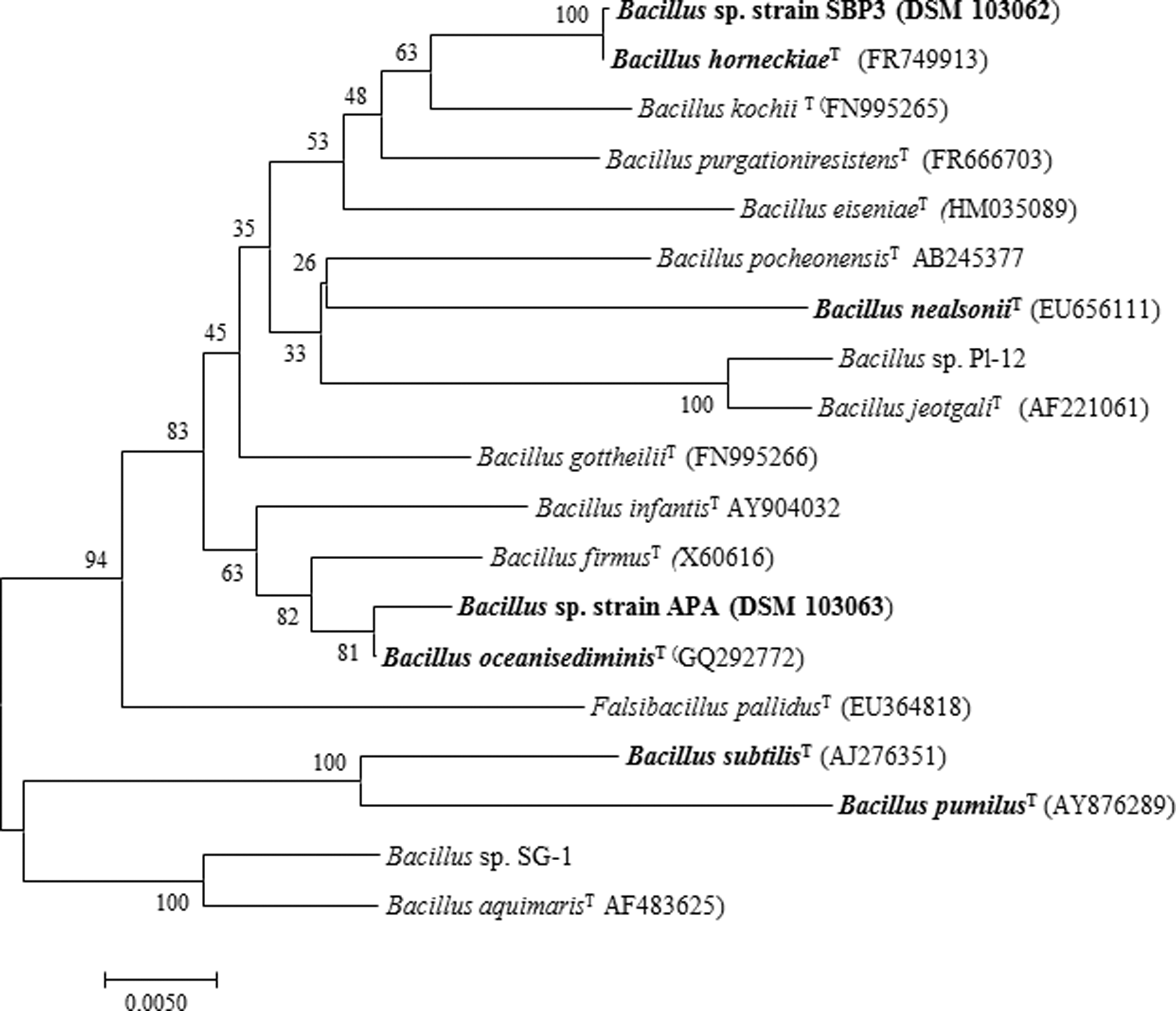
Phylogenetic tree based on 16S rRNA gene sequence analyses, showing the close relationships among Bacillus sp. strains SBP3 and APA and related type strains. Scale bar = 0.005 substitutions per nucleotide position. For comparison and as references of previously dominant hydrothermal sediments, plumes or vent-isolated Bacillus sp. species/clusters, two species/clusters, that is, (1) Bacillus sp. Pl-12 and (2) Bacillus sp. SG-1 have been added (for details on Bacillus sp. Pl-12 and Bacillus sp. SG-1 phylogeny, see Dick et al., 2006).
Values are percentages of total fatty acids.
n.d. = not determined.
3.2. Spore resistance to UVC radiation, low-pressure argon plasma, X-rays, UHV, H2O2, and dry and wet heat
To study the impact of spaceflight-relevant environmental and artificial conditions on spore viability, dormant spores of different B. subtilis strains, either in suspension or as an air-dried monolayer, were tested. After each treatment, inactivation kinetics of each strain were determined in response to the respective treatment. Survival curves were obtained and the resulting best-fit curves were used to calculate LD90 values (i.e., fluence, dose, or time reducing survival to 10% of the population). A summary of the individual spore inactivation curves and a statistical comparison of their respective spore inactivation constants are given in the Supplementary Data (Supplementary Figs. S1–S14 and Supplementary Table S1). The respective LD90 values or CFUs (in %; in the case of the spore resistance to space-like vacuum) are summarized in Table 3 and were used for statistical comparison (Table 3). As illustrated from the values, spores of B. pumilus SAFR-032 exhibited the highest degree of spore resistance compared with all other species tested, which is in agreement with previous studies (Link et al., 2004; Kempf et al., 2005; Horneck et al., 2012). In addition, the other spacecraft-isolated strain, B. nealsonii, showed a high spore resistance to radiation, chemical, and vacuum treatment. Spores of strain SBP3 showed significantly similar tolerance to low-pressure argon plasma, H2O2 (10%), and dry heat (120°C) with B. nealsonii and to wet heat (100°C) with B. pumilus (Table 3). Moreover, when compared with the planetary protection-relevant spores of its closest relative strain B. horneckiae, spores of SBP3 exhibited higher resistance to all treatments except to H2O2, in which similar resistance was displayed. A comparison of SBP3 spores with the space-biological model system B. subtilis revealed that SBP3 spores were significantly more resistant to low-pressure argon plasma, H2O2, and dry and wet heat. Interestingly, SBP3 spores were more resistant to UVC radiation than B. horneckiae but not as resistant as B. subtilis. APA spores were more resistant than B. subtilis in the wet heat treatment exclusively. Spores of APA and its closest relative strain, B. oceanisediminis, showed only slight significant differences in tolerance of UVC, LPP, H2O2, vacuum, and wet heat, but had similar spore resistance to X-rays and dry heat. However, APA was identified as the most sensitive strain to UVC, LPP, H2O2, and vacuum (Table 3).
Data (spore resistance: LD90 values or survival) are expressed as averages and standard deviations (n = 3). Asterisks (*) indicate survival values that were significantly different (p ≤ 0.05) from the survival value for spores of highly resistant B. pumilus SAFR-032.
R (reduced) or I (increased) survival values that were significantly different (p ≤ 0.05) from the survival value for spores of the respective type strain (i.e., B. horneckiae [Bho] vs. Bacillus sp. SBP3 (DSM 103063) [SBP3]; and B. oceanisediminis [Boc] vs. Bacillus sp. APA (DSM 103062) [APA]; for details see Fig. 3).
A detailed summary and overview of the individual spore inactivation curves and a statistical comparison of the respective spore inactivation constants are given in the Supplementary Material section (Supplementary Figs. S1–S14 and Supplementary Table S1).
Bacillus sp. strains: B. subtilis 168 [Bsu], B. pumilus SAFR-032 [Bpu], B. nealsonii [Bne], B. horneckiae [Bho], Bacillus sp. SBP3 (DSM 103063) [SBP3], B. oceanisediminis [Boc], and Bacillus sp. APA (DSM 103062) [APA].
Treatment: UV = 254 nm UVC radiation, LPP = low-pressure plasma (argon gas flow rate: 100 sccm, pressure of 10 Pa, power of 500 Watt), X-rays (150 keV/20 mA), H2O2 = 10% hydrogen peroxide, UHV = ultrahigh vacuum (10−7 Pa), dry heat = 120°C heat exposure, wet heat = 100°C heat exposure. Spores were either treated as air-dried spore monolayers (air) or as spores in suspensions (liquid).
4. Discussion
The increasing knowledge of microbial physiology in extreme habitats has led numerous scientists to consider the possibility of finding life in various planetary bodies within the Solar System (Nicholson et al., 2000, 2009; Horneck et al., 2010; reviewed in Fajardo-Cavazos et al., 2016). However, robotic probes, including their components and assembly facilities, which may be used to detect biosignatures in extraterrestrial environments, are in need of new cleaning and sterilization procedures to avoid contamination with terrestrial organisms and subsequent false-positive detection (reviewed in Link et al., 2004; Crawford, 2005; Kempf et al., 2005; Nicholson et al., 2009; Khodadad et al., 2017). To expand our knowledge of survival under extreme conditions, this work investigated and compared the resistance of Bacillus spores, isolated from hydrothermal and spacecraft associated environments, toward differing environmental stressors and decontamination treatments.
Our results indicate that the hydrothermal strains of SBP3 and APA were more thermotolerant than their closest relative, B. horneckiae and B. oceanisediminis, respectively. These results were perhaps expected, as these strains inhabit and evolved toward survival in hot, acidic, and salty hydrothermal vents (Caccamo et al., 2000; Maugeri et al., 2009, 2010; Gugliandolo et al., 2012). This is reflected in the measured fatty acid content of the two strains (Diomandé et al., 2015). Specifically, membrane lipids of the strain SBP3 may aptly be designated as thermophilic lipids by displaying the highest iso-C15 (63%) and the lowest anteiso-C15 (nearly 9%) fatty acids content (Table 2). Thermophilic strains able to grow between 45°C and 70°C have iso-C15 content (30–50%) higher than that of anteiso-C15 (lower than 10%). Previous research states that because thermophilic and psychrophilic strains do not appear to regulate the branched chain C15 fatty acid content, the iso-C15 fatty acid content tends to dominate in thermophilic, and the anteiso-C15 fatty acid in psychrophilic bacterial species (Koga, 2012). The fatty acid content of the SBP3 strain is, therefore, highly concordant with the high temperature (130°C) vent environment from which it was isolated, suggesting potential membrane adaptations to the temperature extreme (Koga, 2012). The adaptive mechanisms of these strains could, thus, give novel insights into bacterial abilities to survive in conditions considered prohibitive for life, as well as the conditions where extraterrestrial life forms may be found in the future (Mesbah and Wiegel, 2008).
In agreement with previous reports, B. pumilus SAFR-032 and B. nealsonii, both frequently found as spacecraft assembly facilities isolates, showed the highest degree of spore resistance to spaceflight-relevant environmental stressors (UVC radiation, X-rays, and space-like vacuum) (Link et al., 2004; Checinska et al., 2012; Horneck et al., 2012; Tirumalai et al., 2013). However, in some cases, SBP3 spores displayed similar trends in survival to these highly resistant strains. For example, strain SBP3 showed the same degree of spore resistance to wet heat (100°C) treatment to both B. pumilus SAFR-032 and B. nealsonii (Table 3). Indeed, both thermotolerant spores of SBP3 and APA showed high resistance to wet heat, most likely due to their relatively low core water content and to the intrinsic thermostability of their macromolecules to inhabit the high temperature vents. It has been reported that thermophiles invariably have more wet heat resistant spores than those of mesophiles (reviewed in Nicholson et al., 2000). Contrary to what may be expected at elevated temperatures in dry conditions, spore inactivation by wet heat treatment does not occur through DNA damage (e.g., depurination) (Coleman and Setlow, 2009). Rather, evidence suggests that wet heat treatment targets spore proteins through denaturation processes (Belliveau et al., 1992; reviewed in Setlow, 2016). As previously reported, spores of B. subtilis are well protected against DNA damage caused by wet heat treatment due to the saturation of the spore DNA with small acid soluble proteins (SASPs) and the high levels of divalent mineral ions in spores (mainly Ca2+ and Mg2+ and to a lesser extent Mn2+) (reviewed in Nicholson et al., 2000). In general, the higher the levels of mineral ions associated with the spore core, the greater the wet heat resistance of the spores (Melly et al., 2002; reviewed in Setlow, 2016). Continued investigation into these mechanisms could offer greater understanding of spore resistance to thermal extremes.
Across all treatments, strains SBP3 and APA shared comparable spore resistance with their closely related type strains as well as with space-relevant Bacillus species. Spores of the SBP3 strain demonstrated their ability to survive multistressing conditions at a higher degree, in many cases, than those of the current space-biological model system B. subtilis: SBP3 spore resistance to LPP, H2O2, and wet heat was comparable with or exceeding that of B. subtilis under all simulated extraterrestrial stressors. Because of similar spore resistance patterns, we may suppose that similar mechanisms of resistance could be involved. B. subtilis spore resistance to chemical, heat, and LPP exposure is known to be due to the structure of the coat layers, the impermeability of the spore core, and the low water content of the spore (reviewed in Setlow, 2016) rather than by the presence of enzymes involved in the reactive oxygen species detoxification, such as catalases and superoxide dismutase (Moeller et al., 2014). For spores of B. pumilus, two additional catalases have been isolated and characterized that confer an increased H2O2 resistance (Gioia et al., 2007; Checinska et al., 2012; Tirumalai et al., 2013). It should be kept in mind that multiple mechanisms may be involved in spore resistance to specific agents, since the different responses to UV and ionizing radiation exposure could be due to (i) the content of α/β-type SASPs, core water content, level of mineralization, or number of chromosomes and to (ii) the efficiency of the DNA repair processes during spore outgrowth (for details, see Setlow, 2016).
Here, the two isolates from the hydrothermal vents could represent novel bacterial models for the investigation of spore responses to space environmental stressors and sterilization treatments (Friedline et al., 2015; Mandic-Mulec et al., 2016). Spore formers and thermophiles are among the select bacterial groups able to survive adverse environmental extremes, presenting implications for planetary protection initiatives (Gosh et al., 2010). Because of its resistance to wet heat, radiative stressors, and oxidative stressors suggested from this study, SBP3 could be a superior bacterial model organism for future investigations into the potential for contamination of an ocean world, which could likely harbor these environmental stressors. Such environments, in particular, may play increasing roles in future space exploration and the search for life on ocean worlds (Rummel and Race, 2016; Lunine, 2017). Understanding the resistance patterns of isolates living in deep hydrothermal vents helps to uncover the genetics and physiology required for extreme tolerance. This study encourages future research, especially regarding the mechanisms involved in spore resistance to extreme conditions (natural and laboratory relevant), which is currently largely confined to B. subtilis spore mechanisms but almost unknown for spores of other Bacillus species, including SBP3 and APA (as suggested by Fajardo-Cavazos et al., 2016; Mandic-Mulec et al., 2016).
Footnotes
Acknowledgments
The authors are very grateful to Andrea Schröder, Marina Raguse, and Katja Nagler for their excellent support during spore preparation and inactivation experiments. We express our gratitude to Wayne L. Nicholson for cordially providing the Bacillus pumilus (SAFR-032) strain. We also thank the two anonymous reviewers for their valuable comments. This work was supported by grant from the German Academic Exchange Service (DAAD) to V.Z. (fellowship N.91590859), and, in part, by grants from the German Aerospace Center (DLR-FuE-Projekt ISS LIFE, Programm RF-FuW, Teilprogramm 475) to F.M.F. and R.M. and Deutsche Forschungsgemeinschaft/German Research Foundation (DFG) provided funding to R.M. under grant number MO 2023/2-1, and to M.F. and K.S. under grant number AW 7/3-1.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.