
Abstract
The search for life beyond Earth involves investigation into the responses of model organisms to the deleterious effects of space. In the frame of the BIOlogy and Mars EXperiment, as part of the European Space Agency (ESA) space mission EXPOSE-R2 in low Earth orbit (LEO), dried colonies of the Antarctic cryptoendolithic black fungus Cryomyces antarcticus CCFEE 515 were grown on martian and lunar analog regolith pellets, and exposed for 16 months to LEO space and simulated Mars-like conditions on the International Space Station. The results demonstrate that C. antarcticus was able to tolerate the combined stress of different extraterrestrial substrates, space, and simulated Mars-like conditions in terms of survival, DNA, and ultrastructural stability. Results offer insights into the habitability of Mars for future exploration missions on Mars. Implications for the detection of biosignatures in extraterrestrial conditions and planetary protection are discussed.
1. Introduction
Habitability, defined as the “ability of an environment to support the activity of at least one known organism” (Cockell et al., 2016), is currently a key concept in Astrobiology. The investigation into the possibility of life beyond Earth, and its persistence, implies a thorough definition of those conditions life requires.
To accomplish such investigations, the responses of microorganisms, such as bacterial cells and spores, cyanobacteria, fungi and lichens, to selected space factors (microgravity, galactic cosmic radiation, solar UV radiation, and space vacuum) have been the subject of study over the course of the past two decades in both laboratory simulations and space experiments (Billi et al., 2000; de Vera et al., 2002, 2004; Demets et al., 2005; de la Torre Noetzel et al., 2007; de la Torre et al., 2010; Sancho et al., 2007, 2008; Onofri et al., 2008, 2012, 2015; Olsson-Francis et al., 2009; Horneck and Baumstark-Khan, 2012; Baqué et al., 2013; Meeßen et al., 2015).
Some extreme environments on Earth represent good extraterrestrial analogs, and associated microorganisms in these environments are perfect models for the study of how life can augment its ability to persist and resist (Fairén et al., 2010). Antarctica offers a number of very suitable extraterrestrial analogs to be investigated. Antarctic water tracks, the evolution of which has been investigated in satellite images collected in the Goldman Glacier basin of Taylor Valley, are considered terrestrial analogs to recurrent slope lineae on Mars, which have been inferred to be a possible result of the flow of liquid, likely a saline brine (Levy, 2012). The >400 subglacial lakes that exist across the Antarctic continent (Siegert et al., 2016) are perfect analogs as well for extraterrestrial environments of icy moons such as Europa and Enceladus (Cockell et al., 2011), in particular the largest known subglacial lake, Lake Vostok (Bulat et al., 2004; Lavire et al., 2006).
Mars is certainly the prime target for the search for life beyond Earth. Even if conditions at the surface of Mars are inhospitable for terrestrial life due to cold temperatures, a lack of standing water, and harmful ionizing radiation (Horneck et al., 2010), subsurface refugia could extend the window of habitability, and pockets of habitable conditions could potentially still harbor extant life and their biosignatures.
Our understanding of the evolution of Mars is now sufficient to assign specific terrestrial environments to early or present conditions of the Red Planet. The ice-free areas of Continental Antarctica, such as the McMurdo Dry Valleys, represent the best terrestrial analogs for Mars environments due to their hyperarid, cold, and oligotrophic conditions; associated strong UV irradiation; and geological characteristics (Wynn-Williams and Edwards, 2000). Microbes that live or even thrive in this region lend themselves well to investigation into the possibility of extinct and extant life on Mars; among these, the cryptoendolithic black meristematic fungus Cryomyces antarcticus is a remarkable example. Due to its bewildering ability to resist a number of different stresses, including extremes of temperature; desiccation; high doses of UV-B and UV-C, both sparsely and densely ionizing radiation (Onofri et al., 2008; Selbmann et al., 2011; Zakharova et al. 2014; Pacelli et al., 2017b, 2017c), this organism has been repeatedly selected for space experiments (Onofri et al., 2012, 2015; Pacelli et al., 2017b).
One such experiment is the BIOlogy and Mars EXperiment (BIOMEX), an investigation involving exposure to space conditions that began on October 22, 2014, and ended on February 3, 2016. This experiment involved 16 months of space exposition overall on the facility EXPOSE-R2 payload (Rabbow et al., 2015, about the previous mission EXPOSE-R) on the Russian Zveszda module of the International Space Station (ISS). Objectives for this experiment included (i) to test the ability of biomolecules and cell components to preserve their stability under space and Mars-like conditions, to be used as biosignatures; (ii) to investigate the survival capability of microorganisms under space and simulated Mars-like conditions in space while grown on martian and lunar analog regoliths with the intent to verify any interference or possible toxic effects on microbes (de Vera et al., 2012) and to prepare for the ExoMars Rover mission (Vago et al., 2017).
This study focuses on the analyses of space and Mars simulations in space where samples of the fungus C. antarcticus CCFEE 515 were exposed in the framework of the BIOMEX Experiment and were grown on martian and lunar regoliths. Our results offer insights into the astrobiological relevance of future space exploration missions to Mars and the search for putative traces of past or present life.
2. Materials and Methods
2.1. Organisms and cultivation conditions
The microcolonial black yeast-like fungus C. antarcticus CCFEE 515 was isolated by R. Ocampo-Friedmann from sandstone. Rocks were collected at Linnaeus Terrace in McMurdo Antarctic Dry Valleys (Southern Victoria Land) by H. Vishniac, in the Antarctic expedition 1980–1981 (Selbmann et al., 2005).
Substrata were prepared by mixing malt extract agar (MEA) in Petri dishes with (i) Antarctic sandstone, which is the original substrate (OS); (ii) lunar analogs (L), which mainly consist of anorthosite; and (iii) Sulfatic Mars Regolith Simulant (S-MRS) and Phyllosilicatic Mars Regolith Simulant (P-MRS) to mimic two distinct martian soil compositions that represent soil from two different martian epochs (Böttger et al., 2012). The mineralogical composition of the substrates is reported in Table 1. The fungus was grown on the different substrata for 3 months at 15°C; dishes (12 mm diameter) were cut to fit within the 14 mm diameter wells of the exposure carrier (Fig. 1), and then dehydrated under sterile cabin at room temperature overnight. Twelve samples were prepared overall, four to be accommodated in the upper tray, four in the lower tray of the exposure facility, and four to be maintained in the laboratory in dried condition as laboratory controls. To maintain their integrity in the exposure facility EXPOSE-R2, samples were prepared according to the optimized protocol as reported in the work of Pacelli et al. (2017b).

EXPOSE-R2 facility during exposure outside the ISS. ISS, International Space Station.
Mineralogical Composition of Sulfatic and Phyllosilicatic Mars Regolith Simulants (Present Mars and Early Mars, Respectively) and Lunar Analog Anorthosite (Lunar) in Weight%
P-MRS, Phyllosilicatic Mars Regolith Simulant; S-MRS, Sulfatic Mars Regolith Simulant.
Modified from Böttger et al. (2012).
Additional information on the Expose-R2 payload used in BIOMEX and samples used in the experiments is described in deeper detail in the work of De Vera et al. (2012).
2.2. Space exposure conditions during the EXPOSE-R2 mission
Exposure conditions during the BIOMEX experiment are reported in Table 2. Two sets of samples were integrated into two of the three trays of the EXPOSE-R2 facility: in tray 1 for LEO (low Earth orbit) space exposure and in tray 2 for simulated Mars exposure. They were accommodated in stacks of two sample carriers that allowed a completely identical exposure of the samples in one compartment to space conditions, but only the top samples to solar UV radiation. The samples in the lower carriers were completely shaded as in-flight dark samples. Samples were covered with magnesium fluoride MgF2 (tray 1) or quartz (tray 2) filters and windows, attenuating the solar UV radiation with wavelength >120 nm (tray 1) and a Mars-like UV spectrum with wavelength >200 nm (tray 2) to 0.1% of the total fluence.
Exposure Conditions During the BIOlogy and Mars EXperiment
The EXPOSE-R2 was launched from Baikonur to the ISS on board the Progress cargo spacecraft 56P on July 23, 2014. Outside exposure began on August 18, 2014, and ended on February 3, 2016, after 531 days. For those samples that were evacuated to space, the experiment ran for 509 days, 62 days of which they were in the dark for outgassing, which was followed by 469 days of exposure to UV radiation (the valve was closed on January 11, 2016, to capture the vacuum inside the tray, approximately a month before they were brought back into the ISS and UV irradiation ended). Total mission duration until return to ground on June 18, 2016, with 45S, was 696 days.
In tray 1, the exposure of the samples to LEO vacuum ranged from 1.33 × 10−3 to 1.33 × 10−4 Pa (Rabbow et al., 2015). Tray 2 was already filled with a Mars-like gas mixture composed of 95.55% CO2, 2.70% N2, 1.60% Ar, 0.15% O2, ∼370 ppm H2O (Praxair Deutschland GmbH) at a pressure of 980 Pa in the Planetary and Space Simulation facilities at DLR Cologne on July 2, 2014, leading to a total exposure of the samples to Mars gas and pressure for 722 days.
The lowest temperature measured during the mission of EXPOSE-R2 was −20.9°C, which was limited by a heating system. The highest temperature was 57.98°C. The lowest temperature of the samples investigated was −19.7°C. The highest temperature was 47.2°C. These differences were due to the position of the carriers that contained these samples in the EXPOSE-R2 hardware. The final total mission UV-fluence values calculated by RedShift Design and Engineering BVBA, Belgium, with regard to the different sizes and heights of the samples that led to different fields of view, and the values for the biologically active wavelength ranging from 200 to 400 nm at sample site are not yet available. Preliminary determination of the fluence was 439 and 437 kJm−2 which takes into account the individual attenuation by the 0.1% UV filters for the space tray 1 and the Mars tray 2, respectively.
The total mission cosmic ionizing radiation dose experienced by EXPOSE-R2 was determined with passive detectors and reached values of up to 1 Gy (Berger et al., unpublished, results presented during the EXPOSE-R2 Postflight Review, ESTEC, December 2016).
In parallel to the space mission, laboratory control samples were preserved dried at ambient conditions (room temperature, 1 atm) in the laboratory at DLR in Cologne, Germany.
2.3. Survival assessment
2.3.1. Cultivation test
After space exposure, the exposed, in-flight dark, and laboratory control samples were rehydrated for 3 days in 1 mL of physiological solution (NaCl 0.9%). The colonies were diluted to a final concentration of 50,000 cells/mL, and 0.1 mL of suspension was spread on those Petri dishes supplemented with MEA (five replicates). Plates were incubated at 15°C for 3 months, and then colony-forming units (CFUs) were counted.
2.3.2. Cell membrane integrity
Propidium MonoAzide (PMA; Biotium, Hayward, CA) was added to samples at a final concentration of 200 μM, and to treated colonies and laboratory controls after 72 h rehydration (1 mL NaCl 0.9%). PMA penetrates only damaged membrane cells and cross-links to DNA after light exposure, and thereby prevents polymerase chain reaction (PCR). DNA extraction and purification were followed by quantitative PCR (qPCR), to quantify the number of fungal internal transcribed spacer (ITS) ribosomal DNA fragments (281 bp) present in both PMA-treated and nontreated samples, according to the work of Onofri et al. (2012). Before qPCR, DNA was quantified using a QUBIT system and diluted at the final concentration of 2 ng. Tests were performed in triplicate.
2.3.3. Metabolic activity
For MTT (2-(4,5-dimethyl-2-thiazolyl)-3,5-diphenyl-2H-tetrazolium bromide) assay, a MTT solution in PBS was added to a cell suspension in microplates at a final concentration of 0.5 mg/mL. After incubation at room temperature for 24 and 48 h, 100 μL of dimethyl sulfoxide was added. The absorbance was read at 550 and 650 nm, and the absorbance at 650 nm was subtracted from the other. The results show the 48 h readings, since these were the best reading values obtained. MTT results were normalized by the number of cells per well, and these values were again normalized with the laboratory controls.
2.4. Statistical analyses
Means and standard deviations were calculated. Values obtained for each analog (OS, L, S-MRS, and P-MRS) were normalized to each laboratory control, respectively. Statistical analyses were performed by one-way analysis of variance (ANOVA) and pairwise multiple comparison procedures (Tukey's test), which were carried out by using the statistical software SigmaStat 2.0 (Jandel).
2.5. DNA integrity
The integrity of DNA template was assessed by amplifying different DNA gene lengths of fungal colonies after treatments. DNA was extracted from dried colonies with the Nucleospin Plant kit (Macherey-Nagel, Düren, Germany), following the protocol optimized for fungi (Onofri et al., 2012). Before amplification, DNA was quantified by using a QUBIT system and diluted at the same concentration (2 ng/μL).
ITS and Large SubUnit-coding Sequences (LSU) amplification was performed with BioMix (BioLine GmbH, Luckenwalde, Germany), and adding 5 pmol/ng of each primer and 20 ng of template DNA at a final volume of 25 μL. Random amplification of polymorphic DNA (RAPD) was performed with BioMix and adding 5 pmol of the primer and 1 ng of template DNA at a final volume of 25 μL. The primer used for RAPD was GGA7 (GGA GGAGGAGGAGGAGGA GGA) (Kong et al., 2000). The amplification was carried out with MyCycler Thermal Cycler (Bio-Rad Laboratories GmbH, Munich, Germany) equipped with a heated lid.
ITS, LSU, and RAPD amplification conditions were as those reported by Pacelli et al. (2017b). Band intensities were measured and compared by using ImageJ software (Schneider et al., 2012).
2.6. Sequencing and alignment
Sequencing reactions were carried out by MACROGEN, Pathfinder in Genomics Research, 1001 World Meridian Center, Seoul, Korea (
2.7. Ultrastructural damage
After rehydration, laboratory control and in-flight colonies were prepared for transmission electron microscopy (TEM), according to the following protocol (Pacelli et al., 2017b): samples were treated with 5% glutaraldehyde/cacodylate sucrose buffer 0.1 M (pH 7.2) for 12 h at 4°C, washed three times in the same buffer for 1 h each at 4°C, and fixed with 1% OsO4 + 0.15% ruthenium red in 0.1 M cacodylate buffer (pH 7.2) for 3 h at 4°C. Samples were washed in distilled water (two times for 30 min at 4°C), treated with 1% uranyl acetate in distilled water for 1 h at 4°C, and washed in distilled water (two times, 30 min at 4°C). Samples were dehydrated in ethanol solutions: 30%, 50%, 70% (15 min each at room temperature), and 100% EtOH (1 h at room temperature). They were then critical point dried and infiltrated in ethanol 100%: LR White resin (Agar Scientific) (2:1 for 3 h; 1:1 for 3 h, 1:2 overnight), in rotator, at 4°C and in pure resin for 1 day and overnight. As a final step, the samples were included in pure resin in gelatinous capsule for 2 days at 48–52°C. Sections obtained using the Reichert Ultracut ultramicrotome were stained with uranyl acetate and lead citrate; the image acquisition was performed at the Center for High Instruments, Electron Microscopy Section of University of Tuscia (Viterbo, Italy).
3. Results
3.1. Survival: Cultivation and PMA tests
The capability to undergo cell division and form colonies, after 72 h of rehydration, was investigated.
All the results obtained were normalized against the respective laboratory controls. Also, to give a real picture of the differences among analogs, and in comparison with fresh cultures, raw data are reported in supplementary material (Supplementary Table S1; See Supplementary Data at
Cryomyces antarcticus retained a significant colony-forming ability after 16 months of exposure to real space and simulated Mars-like conditions on the ISS, showing a similar trend of survival, as CFU counts, in all the substrates tested (Fig. 2).
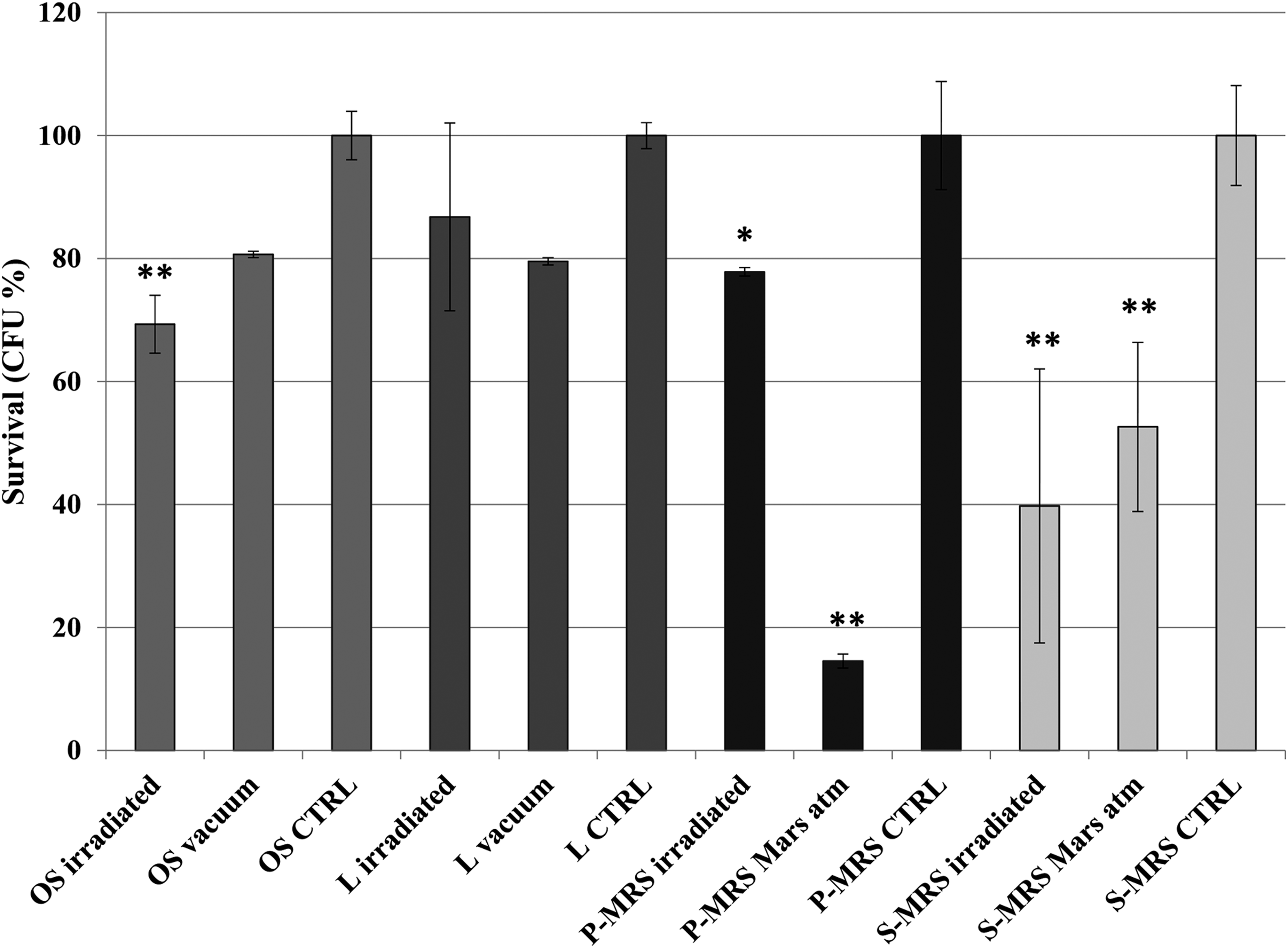
Cultivation test after space exposure: percentage of CFUs of Cryomyces antarcticus on different substrates and conditions as reported in Table 2, relative to laboratory controls on the same substrate. Significant differences are calculated by Tukey's test with *p < 0.05 and **p < 0.001. CFU, colony-forming unit.
Notably, a considerable degree of survival was still maintained; 70% (±4.7), 86% (±15), 78% (±1), and 40% (±22) of survivors were recorded for fully irradiated colonies grown on OS, lunar analog (in the space vacuum), P-MRS, and S-MRS (in space simulated Mars atmosphere), respectively. The radiation plus vacuum represents the most important stress for the fungus cultivated on the OS (i.e., Antarctic sandstone) compared with the vacuum in the in-flight dark samples; a decrease of 20% of the vitality was measured in OS vacuum in the in-flight dark samples compared with the laboratory control, and an additional decrease of 10% was recorded for fully irradiated samples. Similar trends were obtained for S-MRS samples, but differences between fully irradiated and in-flight dark samples in Mars-simulated atmosphere are not statistically significant. In P-MRS samples (early Mars analogs), Mars-simulated atmosphere in the dark seems more effective on survival than in full solar exposure (>120 nm). Differently, for samples grown on lunar analogs, no statistically significant differences were observed between vacuum in the in-flight dark (L vacuum) and fully irradiated samples (L irradiated), the latter not statistically different when compared with the laboratory control. Data discussed above, normalized against laboratory controls, show the same trend for the raw data (Supplementary Table S1).
Viability, in terms of cell membrane integrity, was investigated through PMA treatment followed by qPCR (Fig. 3). The results broadly confirm what was obtained from the cultivation experiment. In OS (Antarctic sandstone) samples, the full irradiation treatments (>120 nm), including vacuum, resulted in 60% (±4) of undamaged cell membranes (presumably alive), while effectiveness of vacuum in the in-flight dark samples was not statistically significantly different. A similar situation was observed for samples cultivated on lunar analogs; yet, a lower percentage of cells with an intact membrane was recorded (34% ± 2). In P-MRS (early Mars analog) Mars-simulated atmosphere in the dark seems more damaging than Mars-simulated atmosphere plus radiation (>200 nm), according to the cultivation tests. In S-MRS (present Mars analogs), the exposure to radiation (>200 nm) and simulated Mars-like atmosphere showed 17% (±5) of undamaged cells, also confirming the cultivation tests. A similar high damage was observed in in-flight dark samples exposed to Mars-simulated atmosphere compared with the laboratory control.

3.2. Metabolic activity analyses
The MTT assay was performed to assess cell metabolic activity recovery after space exposure. The most representative results were obtained after 48 h and are shown in Figure 4. The MTT activities decreased accordingly to the treatment from the vacuum/Mars conditions to the full irradiation for colonies grown on OS and S-MRS; while L and P-MRS samples showed a more marked reduction of the metabolic activity in the in-flight dark samples, with a higher decrease on the latter (below 20%). Overall, the fungus recovered a good level of metabolic activity, even when fully irradiated in all substrates: 45% (±2), 70% (±1), 35% (±2), and 33% (±1) of activity were recorded for OS, L, P-MRS, and S-MRS, respectively (Fig. 3). These results are in accordance with the survival tests reported above.

Assessment of metabolic activity of C. antarcticus by XTT (2,3-bis(2-methoxy-4-nitro-5-sulfophenyl)-5-[(phenylamino)carbonyl]-2H-tetrazolium hydroxide) assay, after space exposure on different substrates and conditions as reported in Table 2. The statistical analyses were performed as shown in Figure 2.
3.3. DNA damage
The presence of possible genomic DNA damage was assessed by testing its efficiency as PCR template after space exposure. All the amplifications were successful for both the ITS and LSU regions (Figs. 5 and 6). In particular, for the 600 bp gene length, no intensity reduction of PCR bands was recorded for all samples. The integrity of DNA was confirmed by amplicon sequencing; the sequences from samples under different treatments gave invariably perfectly preserved electropherograms, and no mutations, insertions, or deletions were visible in the alignment, even in the fully irradiated samples (Fig. 5B).
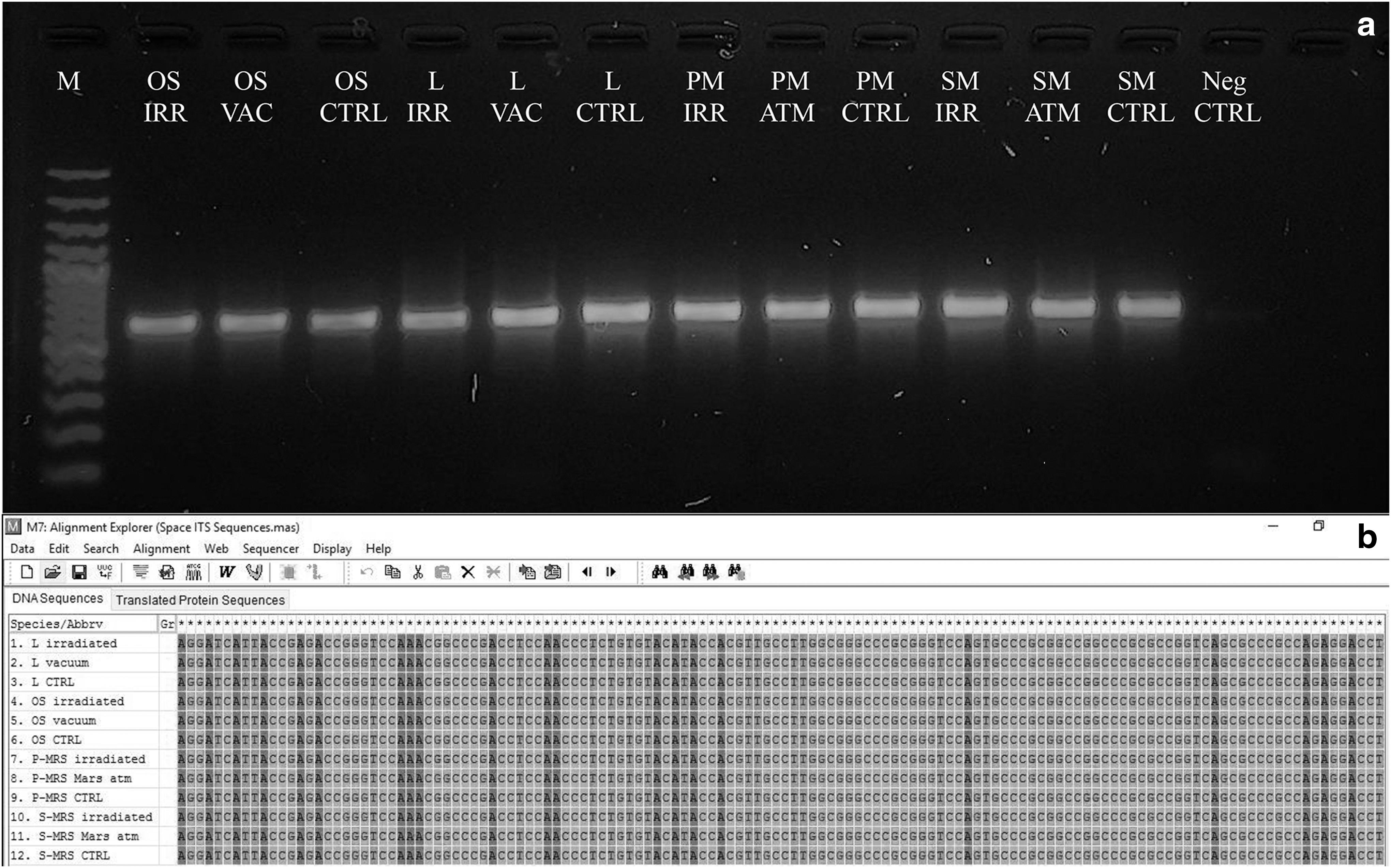
Assessment of the DNA damage on C. antarcticus after space exposure on different substrates and conditions as reported in Table 2;
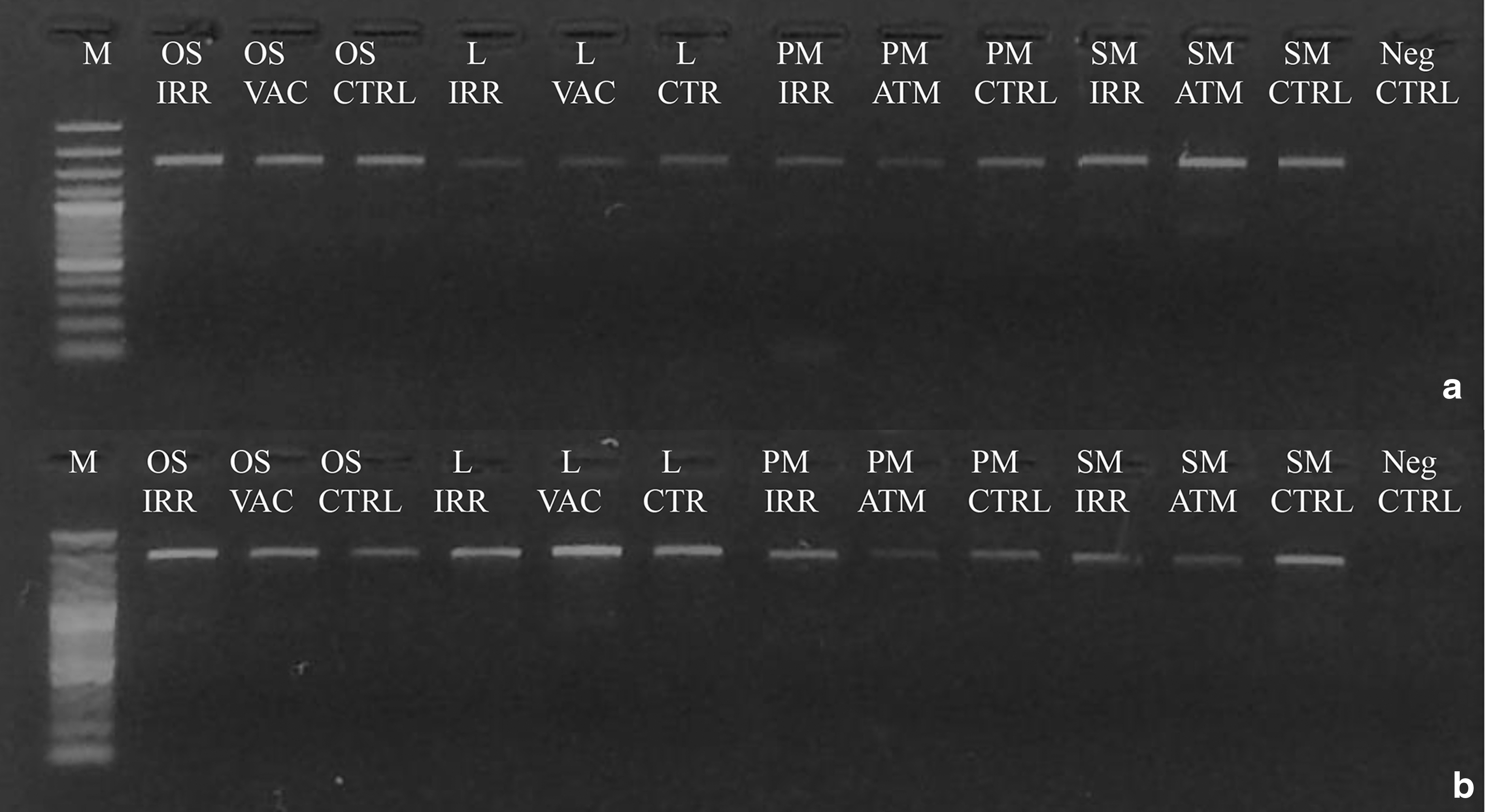
Assessment of the DNA damage on C. antarcticus after space exposure on different substrates and conditions as reported in Table 2; single gene PCR
For the 1600 and 2000 bp gene length, a decreased intensity of amplicons was visible in some of the treated samples, compared with the laboratory control; however, no disappearance of amplicon bands was reported for all treatments (Figs. 6A, B), indicating a good preservation of the DNA template. The RAPD profiles were preserved in all the conditions tested (Fig. 7), demonstrating an overall good preservation of the whole fungal genomic DNA.
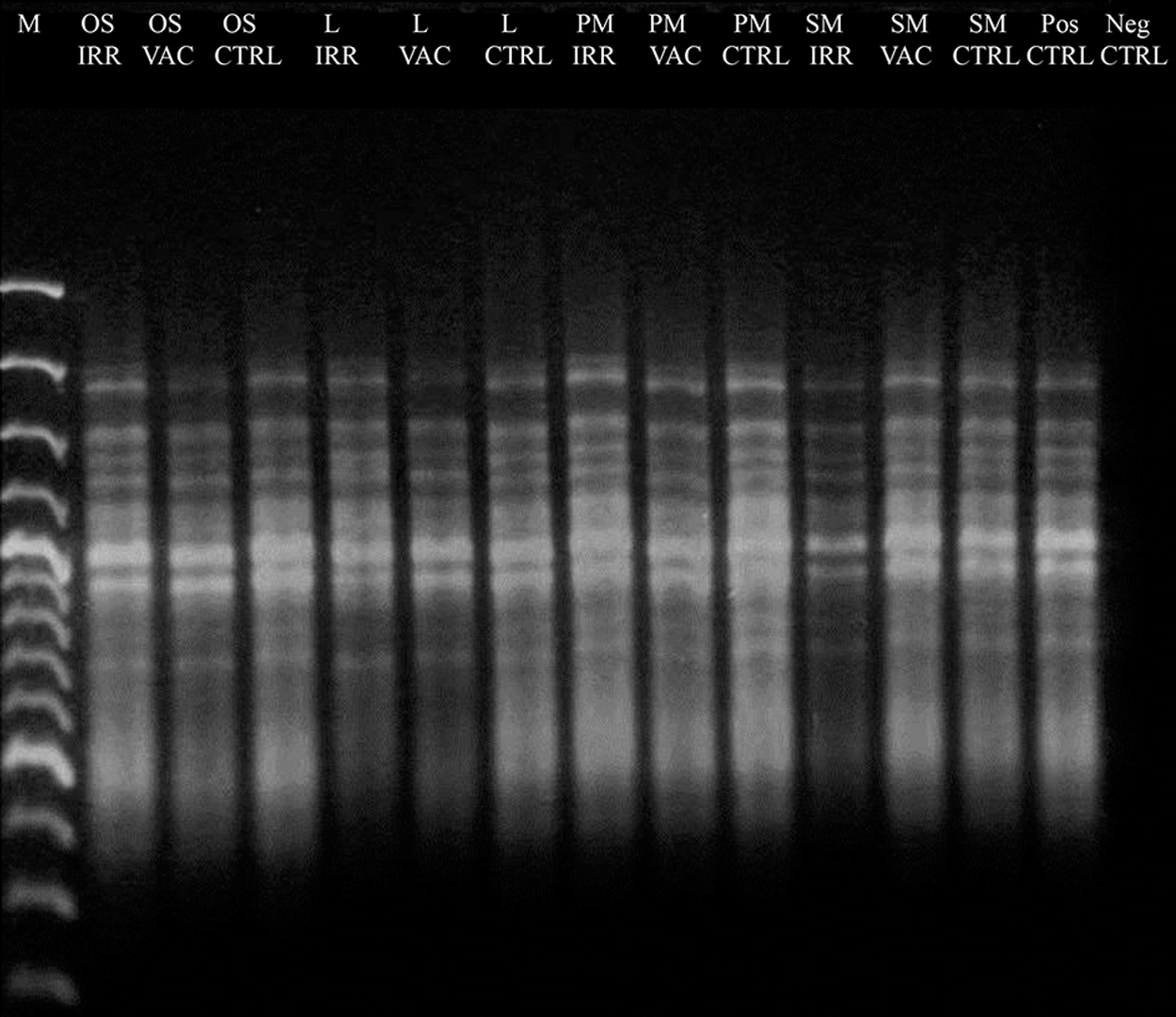
Assessment of the DNA damage on C. antarcticus after space exposure on different substrates and conditions as reported in Table 2 by total DNA fingerprinting (RAPD). No alterations are visible in space samples compared with the laboratory controls. RAPD, random amplification of polymorphic DNA.
3.4. Transmission electron microscopy
Well-preserved ultrastructural features were observed after all of the experimental conditions performed on the ISS and in the laboratory control samples (Fig. 8). Compared with fresh cells (data not shown), TEM observation on treated samples and laboratory controls revealed the presence of evident plasmolysis of the cells due to dehydration. Cells from cultures on OS (i.e., Antarctic sandstone) showed high preservation of ultrastructural features of nuclei, membranes, and organelles in space conditions, in both irradiated and in-flight dark samples, and in laboratory controls (Fig. 8a–c, white arrows); the same has been observed in lunar analogs (Fig. 8d–f). Some damage to membranes and organelles can be noticed in S-MRS (present Mars analog) and P-MRS (early Mars analog) samples exposed to Mars-like radiation (>200 nm) and Mars-like simulated atmosphere (Fig. 8g, j, black arrows), but ultrastructure was preserved in in-flight dark and laboratory control samples (Fig. 8h, i, k, and l).
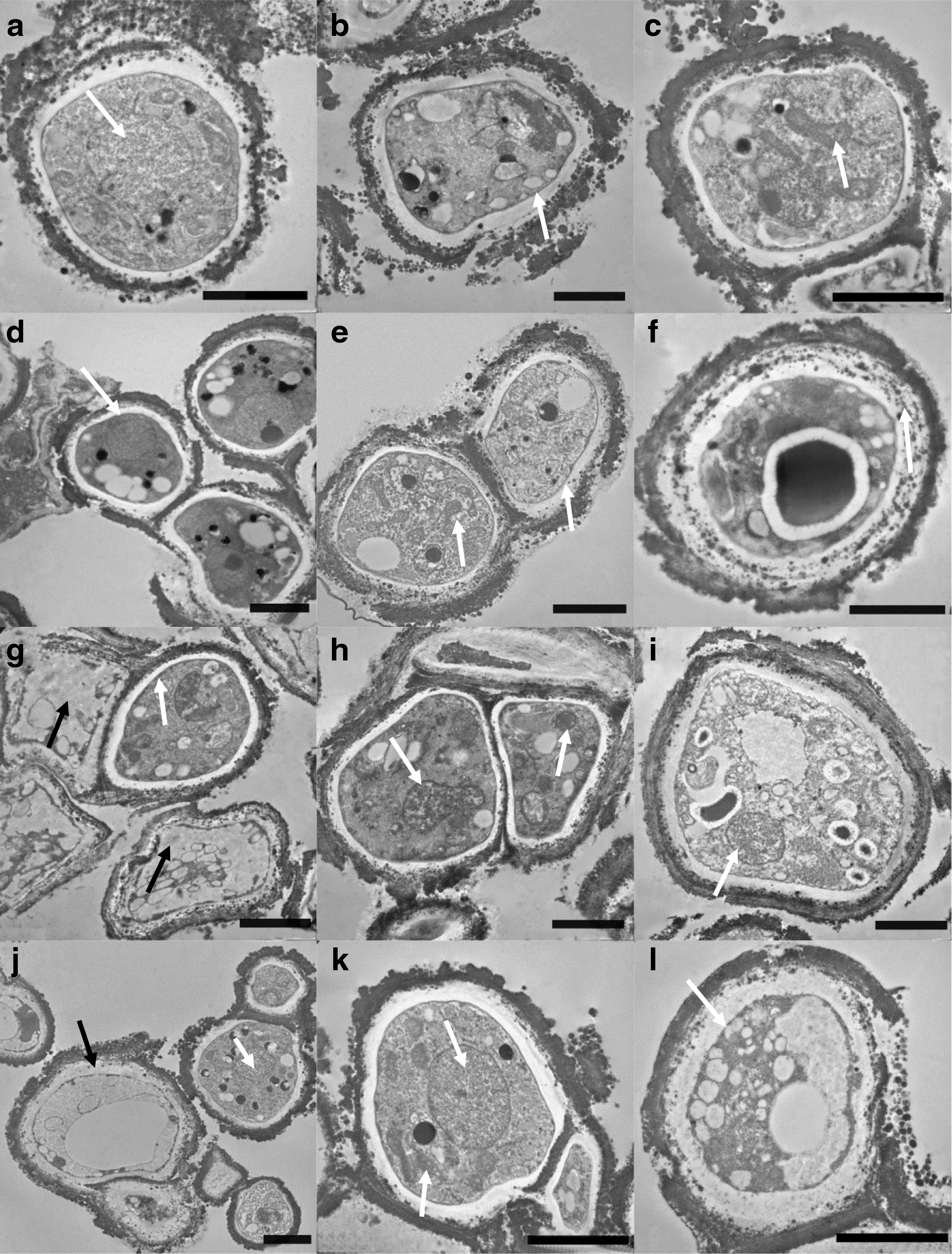
TEM micrographs of cells of C. antarcticus grown on original substrate
4. Discussion
Cryomyces antarcticus is one of the best eukaryotic astrobiological model organisms to date, given its capability to resist, when dehydrated, space vacuum, radiation, and simulated Mars-like conditions in space (Onofri et al., 2012, 2015); moreover, C. antarcticus proved to be metabolically active during simulated Mars-like conditions in ground-based experiments (Zakharova et al., 2014). In the frame of the BIOMEX experiment, the possible interference or toxic effect of extraterrestrial substrata on eukaryotic and prokaryotic astrobiological models was investigated, while exposed to real space and Mars-simulated conditions in space.
The survival and resistance of C. antarcticus, grown on Antarctic sandstone, on lunar and Mars analogs, exposed to space and Mars-like conditions, the latter was simulated in space, on the ISS, was investigated as follows: (i) colony-forming ability; (ii) cell membrane damage; (iii) metabolic activity; (iv) DNA integrity; and (v) ultrastructural damage.
The survival, expressed as CFUs obtained culturing cells of C. antarcticus once back from space, was very high, ranging from 40% to 80% in fully irradiated samples, investigated in both space and Mars-like conditions (Fig. 2). Survival of in-flight dark samples in vacuum and in Mars atmosphere was also elevated, but the latter was apparently more harmful.
Cell membrane integrity of C. antarcticus space samples observed in this study was not as good as reported in previous space experiments where it never fell below 60% (Onofri et al., 2012, 2015), but it is of particular interest since the fungus was grown on lunar and Mars analogs (Fig. 3). Results obtained from the cultivation method are confirmed also for in-flight dark P-MRS samples where the effect of Mars atmosphere seems more harmful than Mars-simulated radiations (>200 nm) (Fig. 3).
All the treated samples recovered metabolic activity of up to 70% with regard to L samples (Fig. 4). The general trend was congruent with the survival obtained from cultivation test; in-flight dark P-MRS samples, in fact, showed the lowest results in both survival and PMA tests; that is, 17% of metabolic activity recovery only.
The PCRs were successful in all the tested conditions, indicating a good integrity of the template both as single gene amplification and as whole-genome fingerprinting (Figs. 5 –7). PCR-stop assays are a powerful tool for evaluating DNA damage and are based on the idea that lesions block the DNA polymerase progression (Kumar et al., 2004; Trombert et al., 2007), and long amplicons have a higher likelihood to encounter DNA damage than short PCR amplicons (Rudi et al., 2010). In addition, genomic PCR fingerprinting, which employs short primers of arbitrary sequences to amplify random portions of the sample DNA by PCR (Williams et al., 1990), reveals DNA lesions by yielding altered PCR profiles with earlier disappearance of higher MW bands (Atienzar et al., 2002; Atienzar and Jha, 2006).
These approaches have been set in dried C. antarcticus colonies exposed to UV irradiation (Selbmann et al., 2011), and then successfully applied to both real space and ionizing radiation and alpha particle-exposed samples of the same fungus in the frame of LIFE and STARLIFE experiments, respectively (Onofri et al., pers. comm.; Pacelli et al., 2017a, 2017c).
TEM observations revealed the ultrastructural damage. Even if some injured cells were undoubtedly present in the irradiated samples (Supplementary Fig. S1), mainly in P-MRS (early Mars analog, Fig. 8g) and S-MRS (present Mars analog, Fig. 8j) samples, intact cells were also observed and were more frequent in OS (i.e., Antarctic sandstone, Fig. 8a–c) and lunar analog (Fig. 8d–f), where survival and metabolic activity were higher. Overall, TEM observations were congruent with the survival results.
All tests performed revealed that P-MRS samples exposed to Mars atmosphere were more affected than irradiated P-MRS samples in terms of survival, membrane, and DNA integrity and metabolic activity. These results are difficult to explain and could be attributed to a technical problem during the flight.
These results clearly indicate that extraterrestrial soils did not hamper survival of our test organism; contrary to expectations, it has been recently reported that the oxidation–reduction potential of Red Planet soils is within the range expected for habitable soils (Quinn et al., 2011), and data obtained from this work are in accordance.
In conclusion, this work corroborates our understanding of the potential for C. antarticus not only to survive extraterrestrial conditions over longer time scales of applied space exposure but also to cope with extraterrestrial regolith analogs. This evidence has a number of implications on (i) the possibility of life transfer among planets, as suggested by the Lithopanspermia theory (Arrhenius, 1903, 1908; Mastrapa et al., 2001; Stöffler et al., 2007), (ii) the habitability of present Mars, and (iii) planetary protection issues. Knowing how terrestrial organisms can augment their survival potential informs with regard to laboratory control and the prevention of biological contamination of target planetary bodies during space exploration missions and the search for life beyond Earth. This is a key prerequisite to the preservation of the pristine nature of planets within the Solar System and facilitates the avoidance of misleading investigation results.
Isenbarger et al. (2008) and Callahan et al. (2011) indicated that DNA would be a beneficial tool for life detection on Earth and on other planets. The results obtained from this study with regard to the potential to detect DNA that is surrounded by a protective cell wall, even after injury due to intense space radiation, corroborate what has been observed in previous ground-based and space experiments (Onofri et al., 2012, 2015; Pacelli et al., 2017b, 2017c). This leads to the conclusion that the DNA molecule may serve as a viable biosignature if shielded, for instance, by a proper cell wall. Accordingly, it follows that, in the event DNA is detected by future missions to Mars or the icy moons of the outer Solar System, such a discovery may indicate the existence of extant life. Therefore DNA should be taken into account in the development of biosignature databases mainly as a potential signature of present life.
Footnotes
Acknowledgments
We thank the European Space Agency (ESA) for supporting the ISS experiment and the Russian cosmonauts who took care of the samples during the exposure. We also thank the Italian Space Agency (ASI) for cofunding the research (BIOMEX MCF Experiment on ISS for tracking biosignatures on martian and lunar rock analogs and E-GEM GeoMicrobiology for Space Exploration grants), the Italian National Program of Antarctic Researches (PNRA), and Italian National Antarctic Museum “Felice Ippolito” for funding the collection of Antarctic fungi CCFEE.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.