
Abstract
Icy environments may have been common on early Earth due to the faint young sun. Previous studies have proposed that the formation of large icy bodies in the early ocean could concentrate the building blocks of life in eutectic fluids and, therefore, facilitate the polymerization of monomers. This hypothesis is based on the untested assumption that organic molecules are virtually incompatible in ice Ih (hexagonal ice). In this study, we conducted freezing experiments to explore the partitioning behavior of selected amino acids (AAs; glycine,
1. Introduction
L
Apart from Earth, icy environments have been shown to be pervasive in habitable planetary bodies. For example, Mars is thought to have had abundant surface waters in the Noachian and early Hesperian periods (Andrews-Hanna and Lewis, 2011; Grotzinger et al., 2014). But compared with early Earth, early Mars surface temperatures were relatively cool because of the planet's greater distance from the faint young sun and its considerably smaller mass in comparison with Earth (Feulner, 2012). Although high concentrations of greenhouse gases that could have sustained a warm and wet early Mars surface have been proposed (Pollack et al., 1987; Ramirez et al., 2014), subsequent modeling simulations and geological observations have invalidated such high levels of greenhouse gases (Batalha et al., 2015; Bristow et al., 2017). Therefore, early Mars likely had widespread icy environments as well. Icy environments are also widely distributed in other astrobiologically interesting planetary bodies in the solar system and beyond, such as Europa or Enceladus.
Below the icy environments at the surface of planetary bodies, subsurface liquid water could potentially act as a cradle of life, where organics could form abiotically during water–rock interactions in anoxic/reducing environments (Sleep et al., 2011; Holm et al., 2015). The behavior of organic molecules during the freezing of planetary waters, in particular those that constitute the building blocks of life, could play an important role in the origin of life. For example, it has been suggested that the formation of large icy bodies in the early Earth ocean could have concentrated the building blocks of life from the diluted “primordial soup” of eutectic fluids (formed at equilibrium with ice when aqueous solutions are cooled below the freezing point of pure water but above the eutectic point, where crystallization is completed at constant composition) and, therefore, facilitated the polymerization of monomers (Sanchez et al., 1966; Bada et al., 1994; Trinks et al., 2005; Menor-Salván and Marín-Yaseli, 2012). The critical assumption behind the concentration hypothesis is that organics are highly incompatible with ice Ih (hexagonal ice; the predominant form of ice in the environment) during the freezing of aqueous solutions. This assumption has been shown to be true for most inorganic compounds, such as Na+, Cl−, NO3 −, and SO4 2− (Gross and Svec, 1997), although a few exceptions exist, including F−, NH4 + (Gross and Svec, 1997), and some gases (Diehl et al., 1998; Santachiara et al., 1998; Hoog et al., 2007). In contrast to the vast number of experimental studies of inorganic species, no detailed experiment has been conducted, to date, with the intent to explore the partitioning behavior of organic compounds, especially the building blocks of life, during the equilibrium freezing of aqueous solutions. Therefore, it is of interest to specifically investigate the partitioning behaviors of small monomers such as amino acids (AAs) between ice and aqueous solutions.
In addition to concentrating organics in the early oceans, the partitioning of organic molecules into ice has significance in the extraterrestrial transport of organics to the early Earth. Chemical analyses of meteorites and comets have indicated various kinds of organic molecules that have been abiotically synthesized, such as AAs, nucleobases, and sugars (Ehrenfreund and Charnley, 2000; Burton et al., 2012; Altwegg et al., 2016). Previous studies have proposed several mechanisms for the abiotic synthesis of organic molecules in space. For example, experimental simulations have suggested that monomers such as AAs, sugars, and nucleotide bases could be synthesized through the ultraviolet photolysis of the surface of ice (Bernstein et al., 2002; Muñoz Caro et al., 2002; Meinert et al., 2016). Assuming that these organics are highly incompatible in ice, they would have attached to icy meteorites by way of surface adsorption. In such a case, the organics would have been very vulnerable to lethal UV radiation and high temperatures during transport, which would have severely limited their survivability (Basiuk et al., 1999; Ehrenfreund et al., 2001). The surface adsorption concept contradicts the fact that many AAs found in icy meteorites on Earth have survived their voyage through space. One critical, but untested, assumption behind this problem is again the high incompatibility between organics and ice Ih, and that organics would mainly attach onto the surface of ice.
In this study, we conducted freezing experiments of glycine,
2. Materials and Methods
2.1. Sample preparation
Solutions of the AAs, namely, glycine,
Solubility of the amino acids at 0°C (Lundblad and Macdonald, 2010).
van der Waals volume (Golemis and Adams, 2005).
pH at the isoelectric point (Haynes et al., 2016).
A small droplet (1.5 μL) of solution was loaded onto a thin silicate glass. To prevent evaporation during the experiment, we designed an enclosed sandwich structure with a small copper ring in the middle and another thin silicate glass on the top. Vacuum grease was rubbed onto the rim of the copper ring to seal the reactor.
2.2. Setup and methods for in situ partitioning measurements
Experiments were performed in a Linkam cooling/heating stage. To prevent evaporation, the sample was installed on a precooled platform at 5°C. The temperature was monitored and controlled by the TMS94 instrument (Linkam) with a resolution of 0.1°C. Cooling was achieved by a controlled flow of liquid nitrogen and heating by a resistance coil.
Raman spectra were measured by a Jobin Yvon HORIBA Raman LabRam HR800 spectrometer equipped with a Mitutoyo™ long-working distance 50× objective and an Ar+ laser at 514.532 nm, 100 mW. Again, to prevent the evaporation of water or laser-induced melting of ice during the experiment, we adjusted the parameters of Raman acquisitions to minimize laser exposure of the sample and used grating of 600 grooves/mm, light filter with optical density as 0.3, and acquisition time as 1 × 30 s or 1 × 15 s.
During the experiment, the state of the droplet was continuously monitored by optical microscopy integrated with Raman spectroscopy. In the beginning, the system was quickly cooled to −30°C with a rate of −10°C/min to freeze the whole droplet. After the quench freezing, temperature was increased to about −4°C with a rate of +5°C/min to melt the droplet. When the whole droplet was at the liquidus, two to four small ice crystals were selected, and the crystal seeds were grown by decreasing temperature. We set −0.1°C/min as the cooling rate and retained one temperature until no apparent growth of crystals was observed. In this way, we wanted to make sure the ice crystals were in equilibrium with the eutectic fluid. At a given temperature, Raman spectra of ice crystal and nearby coexisting fluid were measured without changing the settings of acquisition including the focus of the laser, so that the intensities of characteristic peaks in ice and fluid could be directly used to calculate the partition coefficient. We continued the measurements until most of the droplet was crystallized from the optical microscopy. In the meantime, precipitation of crystals other than ice was always monitored during the experiment.
Raman spectra were processed with PeakFit software; a Voigt function was fitted to the Raman peaks of interest. We selected pairs of Raman spectra based on the quality of ice spectra in each measurement (see the following section). The partition coefficient (Kp) of AA between fluid and ice at a given temperature is defined by
where CAA refers to the concentration of AA and IAA represents the intensity of characteristic peak of AA in ice (IAA,ice) or aqueous solution (IAA,fluid).
2.3. Selection of the representative spectra
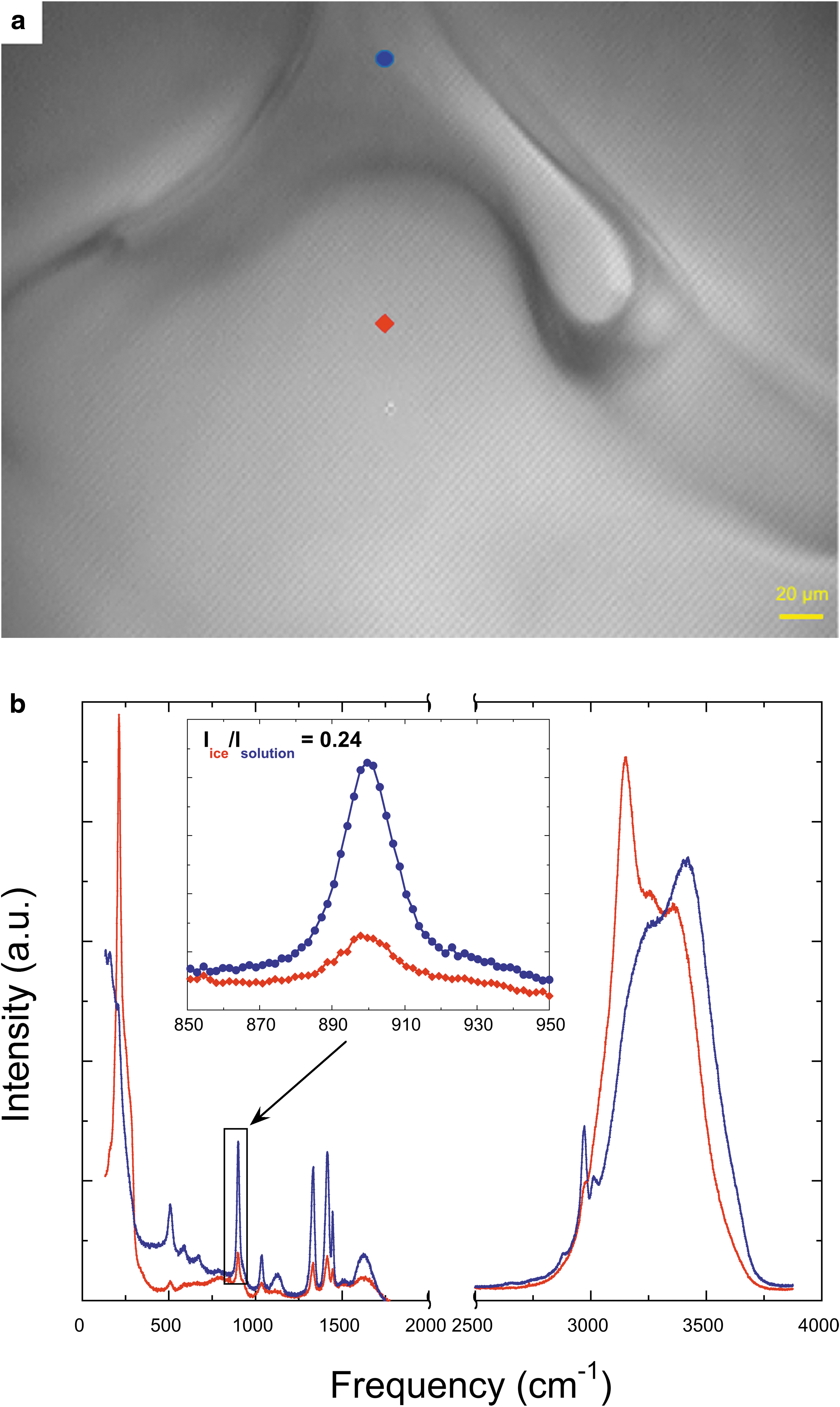
As for data analysis, we carefully selected representative Raman spectra based on the quality of ice spectra to calculate the partition coefficient of the AAs. The major criterion is to have high intensity of the OH-stretching peak at 3150 cm−1 (I3150) relative to that of the OH-stretching peak at 3360 cm−1 (I3360). I3360/I3150 lower than 0.9 was assigned as the criterion to indicate the presence of only ice Ih (Đuričković et al., 2011) (see ice spectra in Supplementary Fig. S2). Additional evidence comes from the strong translational mode at 212 cm−1 of ice spectra (Liu and Ojamäe, 2016). In contrast, there is no Raman signal in the low-frequency region of aqueous solutions. The OH-bending band at 1630 cm−1 is comparatively not very sensitive to the phase change between ice Ih and fluid as is also revealed by the spectra of ice and aqueous solution, both exhibiting an obvious signal of OH-bending band (Fig. 1b).
3. Results
3.1. Freezing behavior of the AA solutions
The freezing behaviors of glycine,
The freezing behavior of 0.05 mol/L phenylalanine and 0.55 mol/L NaCl solution was different. The droplet started showing considerable melting at −2.0°C, indicating a higher melting point than the already mentioned frozen droplet of glycine, alanine, and proline solutions. This difference is explained by considering the much lower concentration of phenylalanine that was used in the experiment as opposed to those of other AAs because of the low solubility of phenylalanine (Supplementary Fig. S1). Similar to the already mentioned solutions, we grew selected ice seeds by decreasing temperature slowly until the temperature reached −6.0°C and the droplet was almost fully frozen.
3.2. Partitioning behavior of AAs between the ice Ih and aqueous fluid
To calculate the partition coefficient of AAs between ice Ih and aqueous fluid, we carefully measured the Raman spectra of ice and coexisting solution during the freezing of the eutectic systems. Figure 1a shows one example of the measurements on the edge of an ice crystal and in the coexisting glycine solution at −6.0°C. The Raman spectra of ice (in red) and solution (in blue) shown in Figure 1b were measured between 130 and 3875 cm−1, but between 1900 and 2500 cm−1, there is no relevant signal. The spectra display the characteristic Raman modes of AAs together with the bands of liquid water or ice Ih. The Raman peaks of glycine are similar in ice and fluid, but with different intensity. There are several characteristic peaks of glycine in the spectra of both water and ice Ih, among which the most intense peaks are at 507, 900, 1035, 1331, 1415, 1445, 2970, and 3011 cm−1 (Fig. 1b). In the ice Ih, we recognized the intense translational model at 212 cm−1 (Liu and Ojamäe, 2016). The librational modes between 600 and 1100 cm−1 are less intense (Liu and Ojamäe, 2016) and masked by the intense peaks of glycine in this study. The bending modes between 1530 and 1700 cm−1 are clearly visible, and the OH-stretching band between 2800 and 3700 cm−1 displays three peaks at 3150, 3255, and 3360 cm−1 that arise mostly from complex strong and weak intermolecular interactions (Li and Skinner, 2010). In the OH-stretching region, the Raman spectrum of liquid water exhibits similar features, which arise from different structural environments controlled by the local hydrogen bonding structure. As a matter of fact, the OH-stretching region in ice is very sensitive to any small contribution of the fluid phase that would increase the intensity of the blue side of the band (Supplementary Data).
Among the Raman peaks of glycine, we chose the symmetric C-C stretching vibration band of the molecule at 900 cm−1 and the CH2 wagging band at 1331 cm−1 (Shigeto and Dlott, 2007) to calculate the partition coefficient of glycine because these bands have the strongest intensity and minimum convolution with others. Since the Raman spectra of the water and ice were taken with exactly the same settings, including the laser power, beam focus, and acquisition time, the ratio of intensities of the glycine peaks at 900 or 1331 cm−1 in ice and solution is equal to the relative concentration of glycine between the ice and solution, which is defined as the partition coefficient of glycine (Kp,glycine). Figure 1b shows a specific example with a Kp,glycine value of 0.24 based on the characteristic Raman peak of glycine at 900 cm−1 in the ice and solution. We obtained similar values of the partition coefficient based on the characteristic peak of glycine at 1331 cm−1 within uncertainties (Supplementary Table S1). Therefore, the selection of the characteristic peak does not have any apparent influence on the calculated partition coefficient.
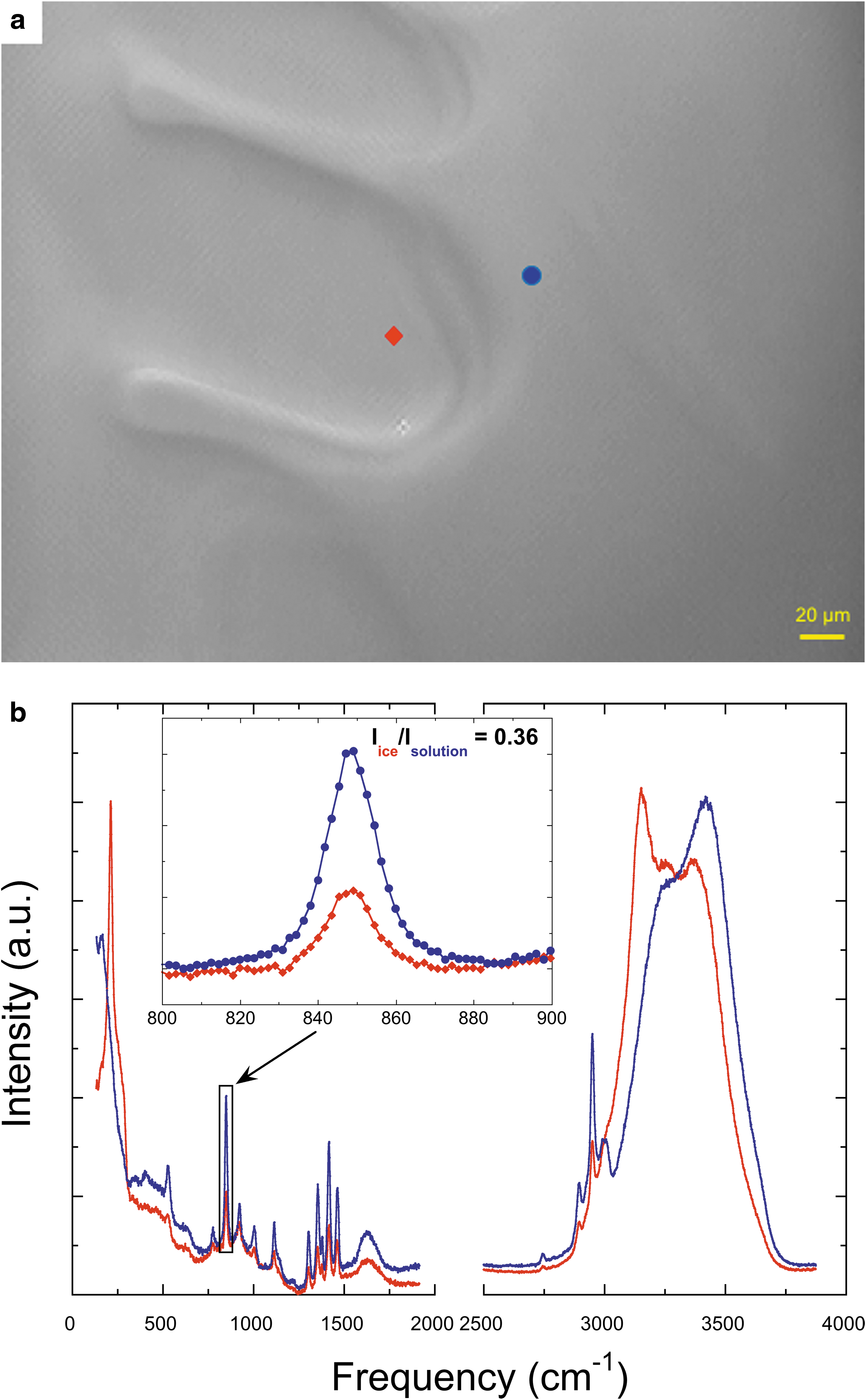
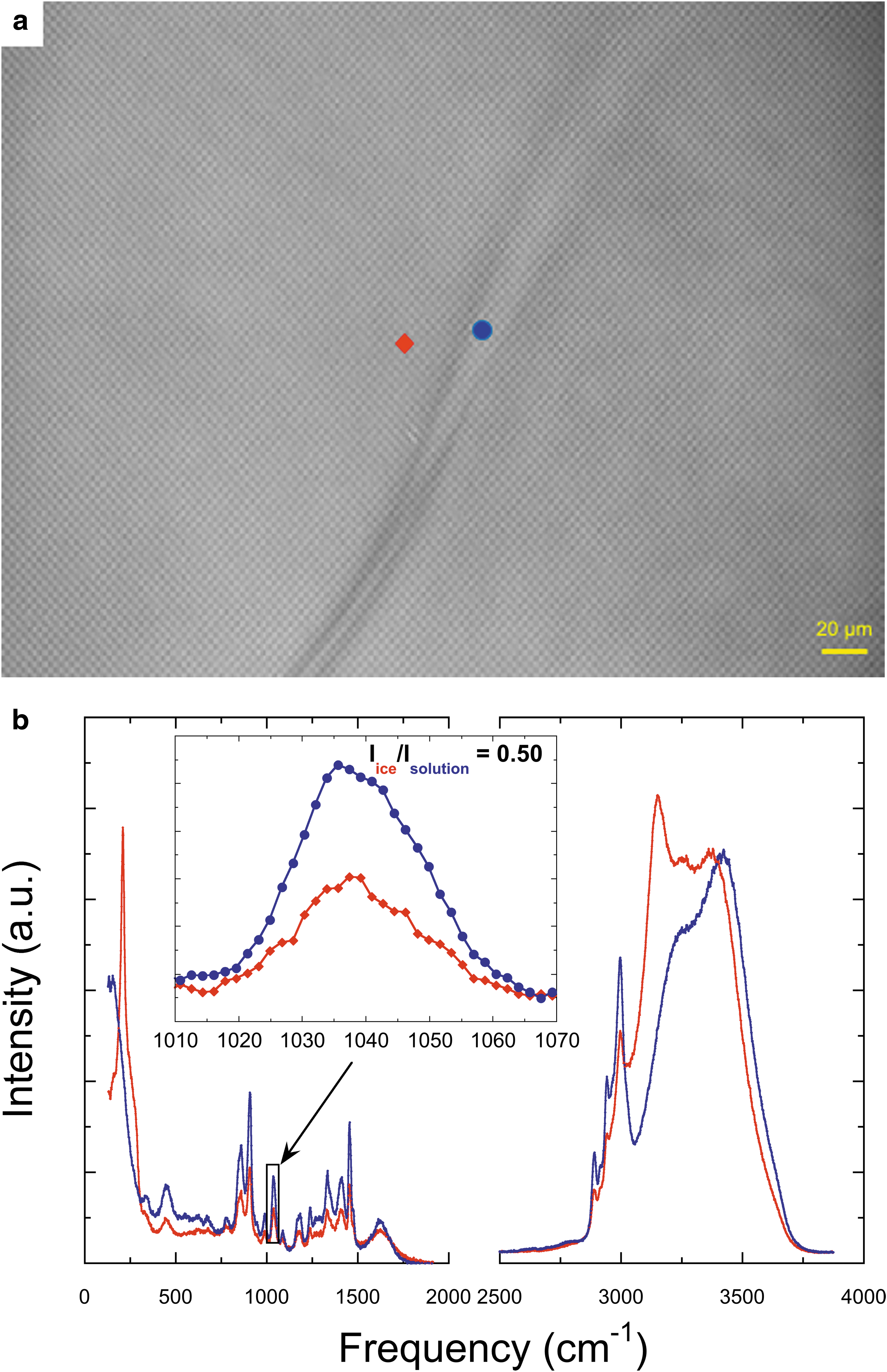
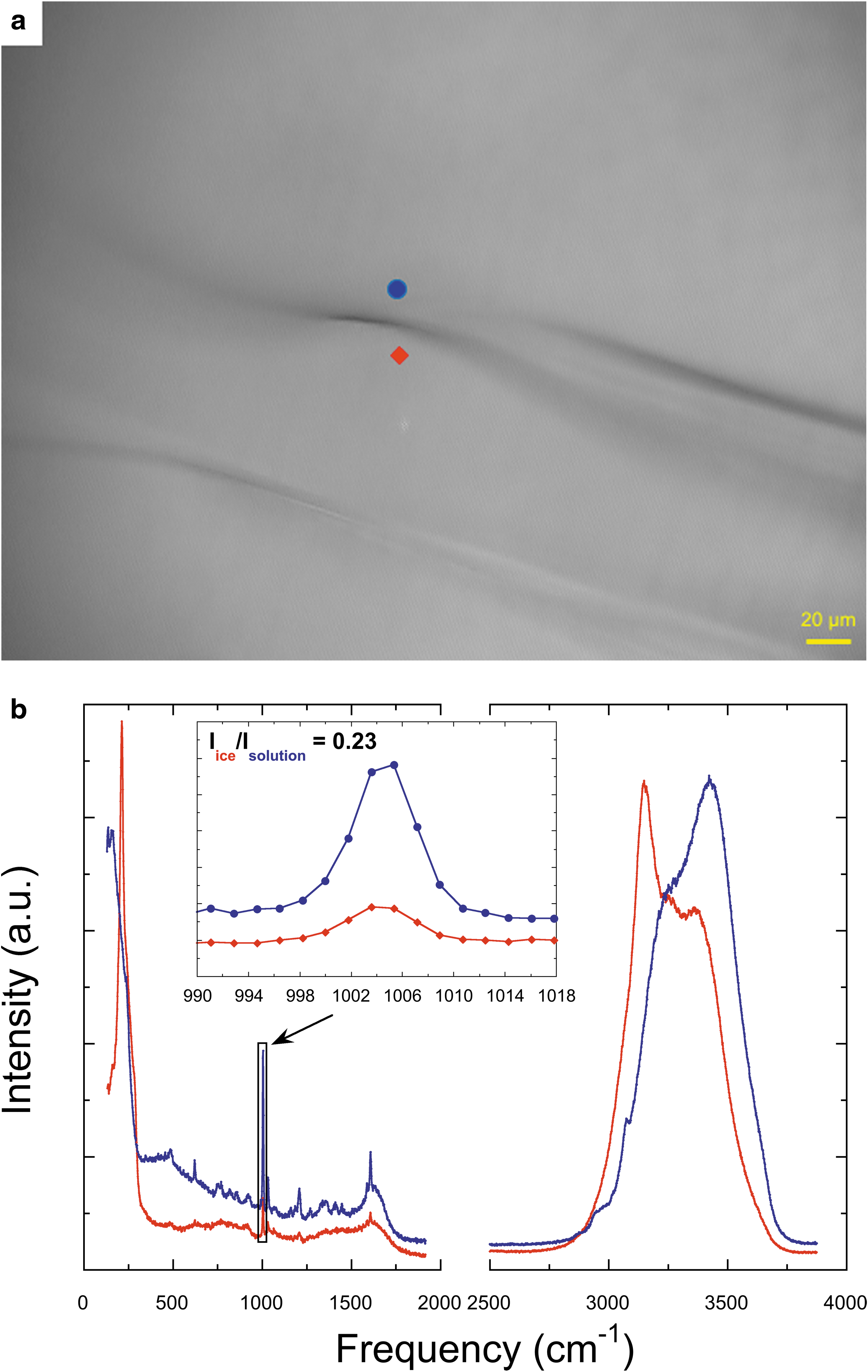
Similarly, we carefully measured the Raman spectra of alanine, proline, and phenylalanine in ice and solutions (Figs. 2 –4). As already mentioned, we selected the strongest and most isolated peaks of alanine, proline, or phenylalanine and calculated the relative intensity of the selected peak in the spectra of ice and AA solution to get the partition coefficient. Specifically, the Raman peaks of alanine at 848 and 1417 cm−1 were chosen as the representative characteristic peaks, and the calculated partition coefficient of alanine based on the selected spectra is 0.36 (Fig. 2b). The Raman peak has been assigned to C-N and C-C stretching vibrations at 848 cm−1 and CH3 deformation at 1417 cm−1 (Kumar et al., 2006). As for proline, we selected the peaks at 989 and 1038 cm−1, which represent CH2 wagging vibration (Herlinger and Long, 1970), and the calculated partition coefficient based on the selected spectra is 0.50 (Fig. 3b). For phenylalanine, we chose the Raman peak at 1004 and 1032 cm−1, which is due to the deformation mode of the benzene ring (Zhu et al., 2011), and the partition coefficient based on the selected spectra is 0.23 (Fig. 4b). The results for the calculated partition coefficients are summarized in Table 1 (details see Supplementary Table S1).
Owing to the small range of temperature between the liquidus and solidus of the solutions, that is, between −2°C and −6°C, we did not detect any dependence of partition coefficient on the experimental temperature (Supplementary Table S1). Therefore, we can directly compare the average values of the partition coefficients of the different AAs (Table 1). Among the AAs investigated in this study, alanine and proline have significantly larger partition coefficients than those of glycine and phenylalanine. This result is unexpected as intuitively one would anticipate that smaller AAs are easier to be incorporated into ice. Our results imply that the partitioning behavior of AAs is not controlled by the size of the AAs.
Most importantly, considering that the partition coefficients are within one order of magnitude <1, our experimental results suggest little incompatibility of AAs in ice Ih, that is, considerable amounts of glycine, alanine, proline, or phenylalanine could be incorporated into the ice Ih during equilibrium freezing of the solutions. In addition, we used high concentrations of glycine, alanine, and phenylalanine close to their solubility limits in the freezing experiment (Table 1) for analytical purposes. Solubilities of the AAs investigated in this experiment have been demonstrated to decrease with lowering temperatures (Supplementary Fig. S1). If AAs were significantly incompatible in ice during the freezing of their solutions, the concentrations of AAs in the eutectic fluids would be elevated during freezing and rapidly exceed their solubility limits. However, we never observed any precipitation of AA crystals, neither by optical microscopy, nor by the Raman spectra during the experiment, further supporting the minimal incompatibility of AAs in ice Ih during equilibrium freezing.
4. Discussion
Unlike inorganic salts, it is obvious that all the AAs investigated in this study showed little incompatibility in the ice Ih during the freezing of the eutectic systems (Table 1 and Supplementary Table S1). The partition coefficients of most inorganic salts have been reported to be around 10−3 except for NH4 + and F−, which showed relatively higher solubility in ice Ih with partition coefficients between 10−2 and 10−1 (Gross and Svec, 1997; Petrenko and Whitworth, 1999). In contrast, the partition coefficients of AAs, as measured in this study, are as high as 0.2 to 0.5 (Table 1), one to two orders of magnitude higher than most inorganic salts. Our results imply that, during the equilibrium freezing of the AA solutions, considerable amounts of AAs could be incorporated into the ice Ih. Clearly, the behavior of AAs is quite different from inorganic ions, invalidating the previous assumption that organics were highly incompatible in ice Ih during the freezing of water solutions (Bada et al., 1994; Menor-Salván and Marín-Yaseli, 2012).
Few studies have been conducted to investigate where impurities could exist in the ice Ih structure. It has been proposed that small impurity atoms could substitute for oxygen or hydrogen atoms in the ice Ih lattice, for example, a fluorine atom replacing oxygen (Petrenko and Whitworth, 1999), but this should not hold true for AAs because of their complex structure with many atoms. Alternatively, the impurities could possibly occupy interstitial sites as in the case of K+ (Petrenko and Whitworth, 1999). However, considering the much larger sizes of the AA molecules (Table 1) compared with those of inorganic atoms and water molecules, it is doubtful how AAs could occupy interstitial sites in the ice Ih lattice.
Another possibility could be the formation of clathrate hydrates of AAs like those of hydrocarbons. However, it has previously been proposed that biological molecules including AAs would inhibit the formation of clathrate hydrates since these hydrophilic molecules have many hydrogen-bonding functional groups that could interfere with the host structure (Jeffrey and Saenger, 1994). This viewpoint has been recently questioned by observations that have shown the formation and stability of clathrate hydrates with some hydrophilic guest molecules (Buch et al., 2009; Alavi et al., 2010; Shin et al., 2013). With regard to the AAs, recent experiments definitely confirmed the inhibiting effects of glycine,
The inhibiting effects of AAs to form hydrates have been attributed to the abnormal incorporation of these AAs into crystalline water by replacing water molecules with the order of inhibiting effects such as
Overall, we rule out the possibility of clathrate hydrates in our study. It is more likely that AAs are mostly incorporated into the ice structure, that is, AAs replace the water molecules in the ice Ih structure. The formation of hydrogen bonding between the functional groups of AAs and water molecules might favor the incorporation of AAs in the ice Ih structure. For example, the amine and carboxylate groups of all AAs could form hydrogen bonds with the water molecules, which promote the replacement and favor a high partition coefficient. In addition, the hydrophilic nature of some AAs like glycine might also facilitate incorporation in the solid water crystal structure (Sa et al., 2014). The crystal chemistry of the AAs in the ice lattice would require X-ray diffraction measurements or quantum chemical or molecular dynamics simulations that are beyond the scope of the present contribution.
5. Implications for Astrobiological Studies
In cool environments on early Earth, freezing of aqueous solutions has been suggested as a potential way to concentrate monomers and, therefore, facilitate polymerization (Bada et al., 1994; Menor-Salván and Marín-Yaseli, 2012). This hypothesis is based on the assumption that organic compounds are highly incompatible with ice Ih like most inorganic ions. However, our experiments showed that considerable amounts of the AAs could be incorporated into ice Ih during the equilibrium freezing of aqueous solutions, with partition coefficients between 0.2 and 0.5. This observation indicates that freezing of early surface waters, if it occurred on the primitive Earth or it occurs on other planetary bodies, is definitely not an efficient way to concentrate AAs or likely other kinds of monomers in the remaining solution of a eutectic system, all of which presents a challenge to the cogency of the previously proposed concentrating process. Moreover, regarding the Raman spectra collected during the experiment, there were no characteristic peaks of peptides formed during the experiment possibly due to the fact that considerable amounts of AAs had entered the ice Ih or because of kinetic hindrance of polymerization at low temperatures.
Icy environments are very common in the solar system and beyond, such as those of icy moons and other water-rich planetary bodies. Organic molecules, particularly AAs, have been shown to be abiotically synthesized through the UV photolysis of ice surfaces (Bernstein et al., 2002; Muñoz Caro et al., 2002), long duration freezing of inorganic solutions (Levy et al., 2000; Miyakawa et al., 2002a, 2002b; Menor-Salván and Marín-Yaseli, 2012), or water–rock interactions (Sleep et al., 2011; Holm et al., 2015). Extraterrestrial transport of organics by meteorites and comets to the primitive Earth has been suggested as one of the major sources of building blocks for the origin of life (Chyba et al., 1990; Chyba and Sagan, 1992; Goldman et al., 2010; Martins et al., 2013). Recently, it was reported that comets had contributed >20% of Earth's atmosphere (Marty et al., 2017). Considering that comets are usually organic-rich, which includes AAs (Altwegg et al., 2016), substantial amounts of organic carbon might have been transported by comets to early Earth. However, the survival of organics in planetary bodies during extraterrestrial delivery is thought to be heavily threatened by UV radiation and high temperature (Basiuk et al., 1999; Ehrenfreund et al., 2001) because previous studies have assumed that AAs are highly incompatible in ice and can only attach on the surface. Thus, the amount and stability of AAs transported by icy meteorites would be severely limited if such a scenario were entirely valid.
Here, we have shown that AAs could be efficiently incorporated into ice Ih during the freezing of aqueous solutions. Under these circumstances, the capacity for ice meteorites to carry AAs could be much higher than previous estimates. Moreover, the incorporation of AAs into ice could significantly improve the survivability of AAs against lethal UV radiation (Pernet et al., 2013; Maté et al., 2015) in space or impact heating (Pierazzo and Chyba, 1999). Collectively, these factors could indicate a greater efficiency of extraterrestrial delivery of AAs to the primitive Earth, perhaps by up to three orders of magnitude compared with previous estimates. Consequently, extraterrestrial delivery in ice might have contributed substantially to the availability of those building blocks required for the origin of life.
Apart from the primitive Earth, our findings in this study have important implications to the origin of life in other water-rich planetary bodies as well. For example, Kimura and Kitadai (2015) calculated the Gibbs free energy of polymerization reaction of organic monomers including glycine and alanine and suggested that the cold environments of icy moons (80–120 K) thermodynamically favor the polymerization of monomers. On one hand, if considerable amounts of AAs could be incorporated into ice as shown in this study, it would limit the availability of AAs as the reactant in the eutectic fluids, and AAs in ice would be far less prone to polymerize than those in fluids. On the other hand, an exchange of chemicals, including AAs, between different layers of icy moons, was previously thought to be inhibited by the icy crust (between surface and subsurface ocean) and/or mantle (between subsurface ocean and silicate core). High-pressure polymorphs of ice such as ice III, V, and VI are thought to constitute the deep solid ice crust and mantle of icy moons (Prieto-Ballesteros et al., 2015). If the partitioning behavior of AAs in Ih is also true for the high-pressure ices, a steady transport of AAs through diffusion in ice might exist, which would allow the transport of AAs from the surface and to a deep silicate core that could fuel a subsurface ocean. In addition, as we mentioned previously, the incorporation of AAs into ice could protect these organic compounds from UV radiation and impact heating, which would increase the opportunity for possible ice-phase chemistry. AAs in the eutectic systems, especially ice structure, might have a low rate of racemization (Bada and McDonald, 1995) and, therefore, provide support for the search for extraterrestrial life by analyzing chirality information of AAs in icy comets.
This study represents a preliminary and essential step in understanding the partitioning behaviors of small organic molecules into ice during the equilibrium freezing of aqueous environments. The findings reported here apply to AAs in ice Ih under the constrained circumstances investigated in this study. The concentration effects by freezing processes might still occur with other building blocks of early life due to a variety of partitioning behaviors, examples of which can be seen in the works of Levy et al. (2000) and Miyakawa et al. (2002a, 2002b).
Footnotes
Acknowledgments
This work is financially supported by The French National Research Agency through the PREBIOM (Primitive Earth - Biomolecules Interacting with Hydrothermal Oceanic Minerals) project no. ANR-15-CE31-0010. J.H. acknowledges the postdoctoral fellowship from LABEX Lyon Institute of Origins (ANR-10-LABX-0066) of the Université de Lyon within the program “Investissements d'Avenir” (ANR-11-IDEX-0007) of the French government operated by the National Research Agency (ANR). The Raman facility at the Ens de Lyon (France) is supported by the Institut National des Sciences de l'Univers (INSU). We also thank H. Cardon (Ens de Lyon) for his help with the Raman analysis and D.A. Sverjensky and R.M. Hazen for the suggestions. Helpful comments from two anonymous reviewers and Dr. S.L. Cady are also acknowledged.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.