
Abstract
Aerobic respiration—the reduction of molecular oxygen (O2) coupled to the oxidation of reduced compounds such as organic carbon, ferrous iron, reduced sulfur compounds, or molecular hydrogen while conserving energy to drive cellular processes—is the most widespread and bioenergetically favorable metabolism on Earth today. Aerobic respiration is essential for the development of complex multicellular life; thus the presence of abundant O2 is an important metric for planetary habitability. O2 on Earth is supplied by oxygenic photosynthesis, but it is becoming more widely understood that abiotic processes may supply meaningful amounts of O2 on other worlds. The modern atmosphere and rock record of Mars suggest a history of relatively high O2 as a result of photochemical processes, potentially overlapping with the range of O2 concentrations used by biology. Europa may have accumulated high O2 concentrations in its subsurface ocean due to the radiolysis of water ice at its surface. Recent modeling efforts suggest that coexisting water and O2 may be common on exoplanets, with confirmation from measurements of exoplanet atmospheres potentially coming soon. In all these cases, O2 accumulates through abiotic processes—independent of water-oxidizing photosynthesis. We hypothesize that abiogenic O2 may enhance the habitability of some planetary environments, allowing highly energetic aerobic respiration and potentially even the development of complex multicellular life which depends on it, without the need to first evolve oxygenic photosynthesis. This hypothesis is testable with further exploration and life-detection efforts on O2-rich worlds such as Mars and Europa, and comparison to O2-poor worlds such as Enceladus. This hypothesis further suggests a new dimension to planetary habitability: “Follow the Oxygen,” in which environments with opportunities for energy-rich metabolisms such as aerobic respiration are preferentially targeted for investigation and life detection.
1. Introduction
Molecular oxygen (O2) on Earth is produced almost exclusively by oxygenic photosynthesis, which is an evolutionary singularity that evolved only once in Earth's history through the evolution of a complex electron transport chain involving coupling of two independent photosystems (Blankenship and Hartman, 1998; Falkowski, 2011; Fischer et al., 2016a). This invention is thought to be essential for the eventual evolution of complex life on Earth by providing O2 for aerobic respiration (Nursall, 1959; Berkner and Marshall, 1965; Raff and Raff, 1970; Catling et al., 2005). That it evolved only once, relatively late in Earth's history, suggests that it is a rare innovation emerging from a complex and contingent evolutionary process—the sort of evolutionary development that may not occur in the same way, if at all, if the tape of life were replayed (Gould, 1990). The evolution of complex multicellularity (i.e., displaying not only cell adhesion but also intercellular communication and commonly tissue differentiation; Knoll, 2011), on the other hand, has occurred independently several times on Earth (e.g., in animals, plants, red algae, and several times within the fungi; Knoll, 2011) but always in aerobic organisms! Simpler forms of multicellularity occur in protists and bacteria (e.g., Fairclough et al., 2010; Claessen et al., 2014) and have even been artificially evolved over short timescales in the laboratory (Ratcliff et al., 2012). The evolution of multicellularity may therefore be “easier,” though it has only occurred on Earth after the rise of O2. The apparent dependence of multicellularity on O2 is likely related to the high energetic yield of this metabolism; O2 reduction is much more thermodynamically favorable and yields significantly more ATP per electron than alternative acceptors such as nitrate and sulfate (Fig. 1), providing a surplus of energy to support not only basic cell metabolism and growth but also more complex behaviors such as motility and cell differentiation (Catling et al., 2005; Koch and Britton, 2008). This suggests that the high energetic yield of aerobic respiration is a necessary precondition for multicellularity and that the belated accumulation of O2 on Earth due to the relatively late evolution of oxygenic photosynthesis delayed the rise of complex, multicellular life.
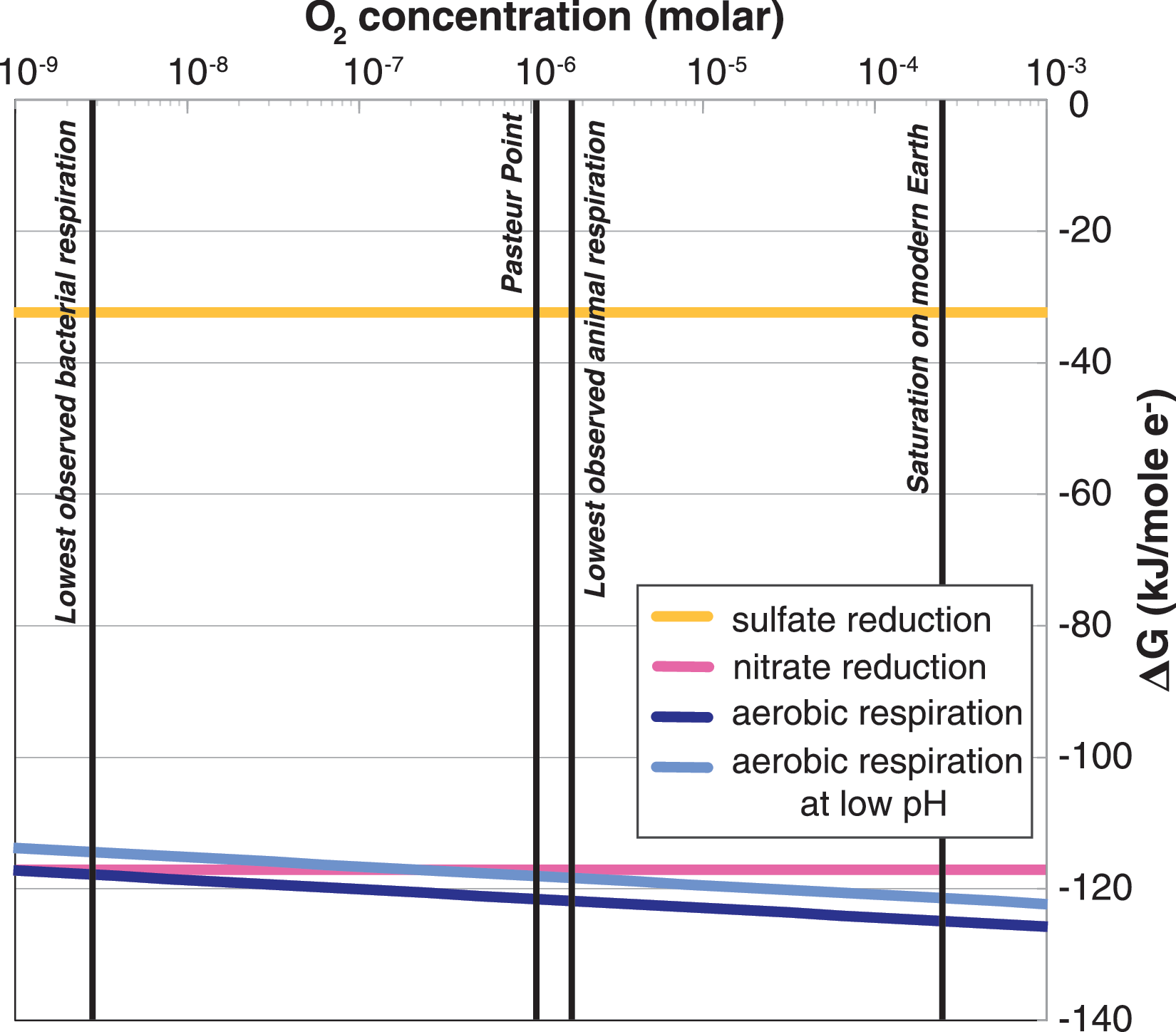
Thermodynamic favorability of microbial metabolisms as Gibbs free energy of reaction across a range of O2 concentrations. Even at very low O2 concentrations, aerobic respiration is still highly favorable (ΔG << 0). Also plotted are lines denoting relevant O2 concentrations, including known limits of respiration for bacteria (Stolper et al., 2010; Zakem and Follows, 2017) and animals (Mills et al., 2014), as well as saturation under surface atmospheric conditions on modern Earth and Mars. Even the thin modern martian atmosphere contains sufficient O2 to support respiration by bacteria such as E. coli and the diverse microbial communities of marine oxygen minimum zones (see Stamenković et al., 2018).
Although some trace abiotic production of oxidized compounds such as O2 and hydrogen peroxide likely occurred on early Earth due to processes such as photolysis of ice (Liang et al., 2006) or water in contact with pyrite (Borda et al., 2001), the geological record shows that biologically meaningful O2 concentrations did not accumulate until ∼2.3 billion years ago, likely due to the small magnitude of abiotic O2 fluxes and the overall reducing state of early Earth (Fischer et al., 2016a). As a result, early life on Earth existed independent of O2 (Sleep and Bird, 2007), though small amounts of abiotic hydrogen peroxide could have spurred the evolution of mechanisms to protect against reactive oxygen species (e.g., Kirschvink and Kopp, 2008). In contrast, abiotic O2 production may be more significant on other planets, so oxygenic photosynthesis may not be necessary to supply O2 for aerobic respiration. For example, on small worlds like Mars, hydrogen escape is more significant, so sufficient O2 may accumulate to support aerobic life even in the absence of oxygenic photosynthesis (though absolute fluxes may still be low relative to biogenic O2 on Earth today) (Stamenković et al., 2018).
In this work, we examined the possibility of aerobic environments on other worlds within the framework of what is known about the history of O2 on Earth and the role of O2 in biology. The history and modern distribution of O2 on Mars and Europa were used as test cases to begin examining the opportunities for aerobic respiration on other planetary bodies where the history of O2 is not contingent on the evolution of oxygenic photosynthesis. This provided a basis for considering the more general case of exoplanets, which under a range of circumstances may support oxygenated atmospheres that may be detectable with upcoming missions.
Drawing on this framework, we hypothesize that on worlds where O2 accumulates through abiotic processes and is therefore not dependent on the complicated evolution of oxygenic photosynthesis, multicellularity could evolve more quickly and easily than it did on Earth. This hypothesis appears to be viable given recent improvements in our understanding of the O2 requirements of life on Earth, as well as the distribution and abundance of oxygen-rich environments on other worlds as the result of abiotic processes. This hypothesis has relevance for the occurrence of life in the Universe more broadly but is more immediately testable within our own solar system in the context of Mars and icy moons like Europa where abiotic O2 and redox gradients may support life in the absence of photosynthesis.
2. O2 on Earth
The history of O2 on Earth is closely tied to photosynthesis. Atmospheric O2 concentrations are constrained to below 10−5.7 atm before the Great Oxygenation Event (GOE) around 2.35 Ga, as recorded by a range of proxies (Holland, 1984; Rye and Holland, 1998; Farquhar et al., 2000; Bekker et al., 2004; Guo et al., 2009; Maynard, 2010; Johnson et al., 2013, 2014; Fischer et al., 2014; Luo et al., 2016). During the GOE, O2 accumulated in the atmosphere to 1–10% of present atmospheric levels (PAL) (∼10−3 to 10−2 atm). While the abundance and regulation of subsequent atmospheric O2 concentrations are contested (e.g., Johnston et al., 2009; Laakso and Schrag, 2014; Cole et al., 2016; Zhang et al., 2016; Daines et al., 2017), it is clear that O2 has remained a significant component of the fluid Earth since the GOE (Lyons et al., 2014). The oxygenation of Earth's atmosphere was enabled by oxygenic photosynthesis developed within Cyanobacteria (e.g., Falkowski, 2011; Shih, 2015) and may have occurred rapidly following the evolution of this metabolism (Ward et al., 2016; Fischer et al., 2016a; Shih et al., 2017; Soo et al., 2017). The presence of O2 in Earth's atmosphere is therefore tightly coupled to the history of a major and singular biological innovation: oxygenic photosynthesis. The availability of O2 ultimately paved the way for the evolution of aerobic respiration (Hemp et al., 2012), endosymbiosis and radiation of the eukaryotes (Katz, 2012), and complex multicellularity (Catling et al., 2005; Knoll, 2011).
While thermodynamics predicts that aerobic respiration should be viable down to vanishingly low concentrations of O2 (Fig. 1), it has remained contentious whether life could make use of these lowest concentrations, or if there was a threshold O2 concentration below which respiration is not physiologically viable. Underlining this discordance, the Pasteur point was typically considered to mark the minimum O2 concentration for aerobic respiration, below which organisms transition to fermentation (or other anaerobic metabolisms) to maintain energetic balance. The Pasteur point corresponds to an O2 concentration of about 1% of PAL dissolved in seawater, or about 3 μM. This had also been thought to be the minimum O2 concentration for aerobic respiration (e.g., Berkner and Marshall, 1965), but recent studies have expanded the known O2 range of respiration downward (Morris and Schmidt, 2013). For example, experiments have demonstrated aerobic respiration by Escherichia coli at vastly lower O2 concentrations, closer to 0.001% PAL (Stolper et al., 2010). This range is consistent with more recent theoretical considerations, which recovered an ecological limit to aerobic respiration of 0.1–10 nM O2 (Zakem and Follows, 2017). This greatly extends the potential range of environmental O2 concentrations supportive of aerobic organisms, as confirmed by observations of obligately aerobic organisms in environments such as oxygen minimum zones (Ulloa et al., 2012; Sperling et al., 2013b), freshwater sediment (Ettwig et al., 2010), and wastewater sludge (Yamada et al., 2006; Ward et al., 2015). This suggests that more environments could be suitable for aerobic life than previously recognized. The topic of the expanded range of environments habitable for aerobes has primarily been discussed in the context of Earth history (e.g., Fischer, 2016; Zhang et al., 2016) and low O2 environments on modern Earth (e.g., Ulloa et al., 2012), but it also has broad relevance to a range of planetary environments, including those on Mars, Europa, and elsewhere.
Aerobic respiration has played a critical role in the development of complex life. Aerobic respiration was a major factor in the development of eukaryotes (Katz, 2012) and ultimately animals (Knoll, 2011), as large, complex organisms have inherently high energetic demands that can be met by aerobic respiration but not by anaerobic metabolisms (Catling et al., 2005). Historically, it has been assumed that animals required high O2 concentrations to evolve due to inherently high O2 demands of multicellularity (e.g., Nursall, 1959). These critical O2 concentrations have never been rigorously quantified, though they have been estimated based largely on diffusion limitations (e.g., Runnegar, 1991). Recent efforts to understand the minimum O2 concentrations necessary to support sponges and other basal animals have demonstrated survival and growth under surprisingly low O2 concentrations, down to about 1% PAL (Sperling et al., 2013a; Mills et al., 2014). While these values are still much higher than the minimum O2 concentrations for bacterial respiration (Stolper et al., 2010; Zakem and Follows, 2017), they greatly extend the spatial and temporal range of environments habitable for animals (e.g., oxygen minimum zones and Proterozoic oceans, e.g., Sperling et al., 2013b; Fischer, 2016; Zhang et al., 2016) and are an important new constraint in thinking about O2 as a requirement and biosignature of complex life on other planets.
3. O2 on Mars
The history of O2 on Mars provides a curious contrast to that on Earth (Fig. 2). While terrestrial O2 is tied to the biological process of oxygenic photosynthesis, Mars appears to have a history of O2 despite no apparent biological source. It is widely appreciated that the surface of Mars is, and perhaps has been since its early history, “oxidizing” (e.g., Hunten, 1979; Zent and McKay, 1994; Christensen et al., 2001; Goetz et al., 2005; McLennan et al., 2005; Tosca et al., 2005; Bibring et al., 2006; Zahnle et al., 2008; Hecht et al., 2009; Hurowitz et al., 2010, 2017; Leshin et al., 2013; Farley et al., 2016). Though O2 is the fourth most abundant species in the atmosphere, Mars currently has low relative and absolute atmospheric concentrations of O2—a mixing ratio of ∼1.45 × 10−3 (Mahaffy et al., 2013). These levels of O2 are higher than the upper levels of O2 allowed by geochemical proxy data on Archean Earth (Farquhar et al., 2000; Johnson et al., 2014). However, a range of proxies suggest that O2 concentrations may have been higher in Mars' past, perhaps when the atmosphere was thicker (e.g., Brain and Jakosky, 1998; Manga et al., 2012; Kite et al., 2014). This notion is further supported by the presence of enriched manganese deposits (Lanza et al., 2014, 2016; Arvidson et al., 2016), multiple oxygen isotope ratio data collected from shallow, low-temperature carbonates preserved in the martian meteorite ALH84001 (Farquhar et al., 1998; Halevy et al., 2011; Shaheen et al., 2015), and the presence of high concentrations of oxidized salts on the surface of Mars thought to be derived from reactions involving ozone and other O2-derived species (e.g., Zahnle et al., 2008; Catling et al., 2010).
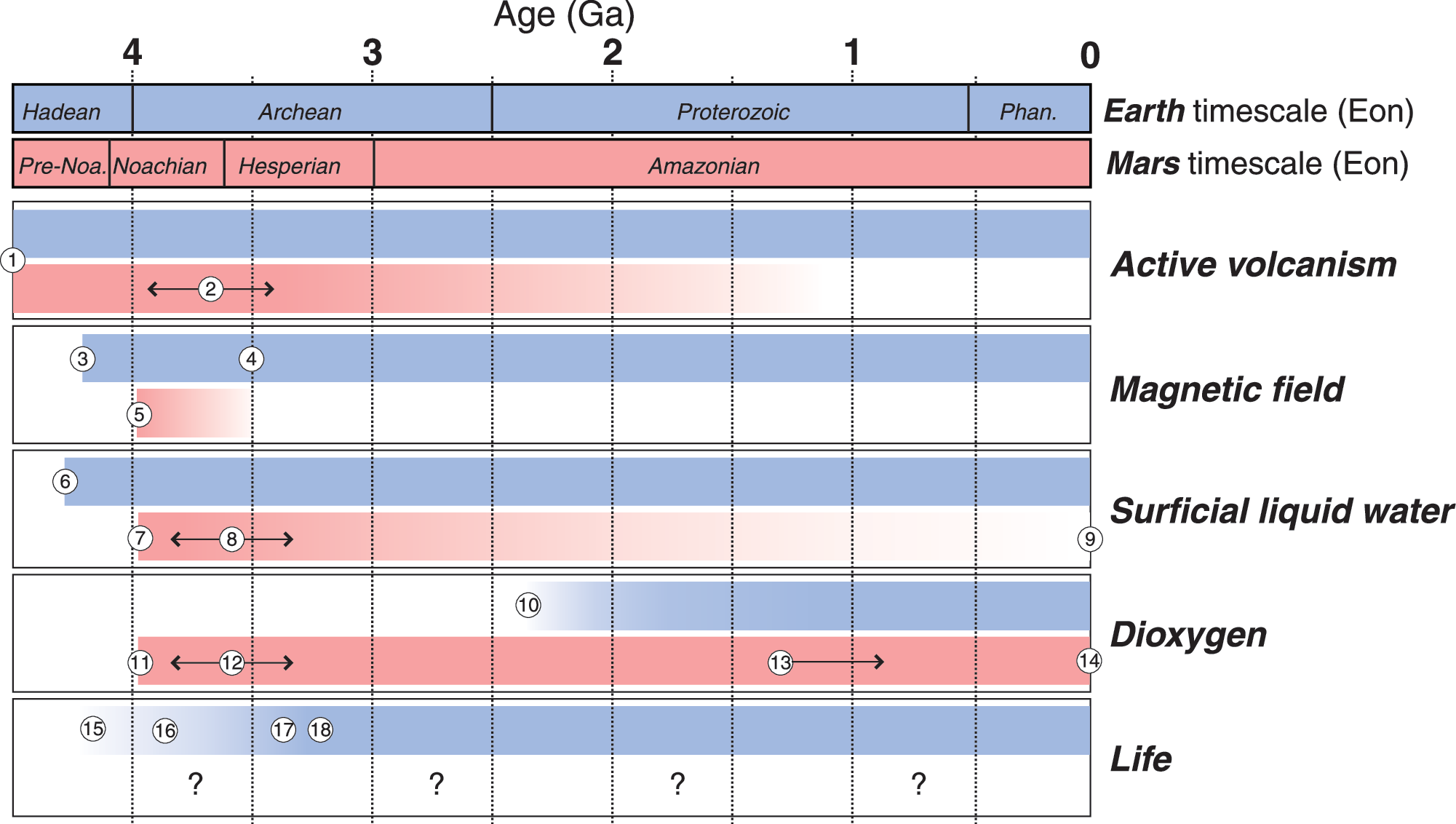
Comparative timelines of Earth and Mars. While the surfaces of Earth and Mars are now quite different, their early histories were likely more similar, with early Mars having once had a magnetic field, surficial water, frequent volcanism, and a denser atmosphere. Mars, however, appears to have had at least periodic significant O2 and ozone early in its history, in contrast to Earth. Numbers reflect key observations and geological phenomena. This timeline largely focuses on qualitative observations of particular relevance for the origin and evolution of life. For more comprehensive timelines of other aspects of the history of Mars, see Ehlmann et al. (2011), Michalski et al. (2013), or Michalski et al. (2018). Active volcanism assumed due to the thermal status of both planets early in their history.
The ancient atmosphere of Mars is thought to have been much thicker than it is today, perhaps on the order of 0.5–2 bar (Kite et al., 2014; Jakosky et al., 2017; Kurokawa et al., 2018). While it is unclear whether martian O2 concentrations scaled linearly with atmospheric pressure through time, under the naïve assumption that the mixing ratio of O2 were similar in this thick early atmosphere to that of modern Mars, saturation of O2 would be on the order of ∼4 μM—sufficient not only for aerobic bacteria but in fact at concentrations above the Pasteur point and in line with the O2 requirements of basal animals like sponges (Sperling et al., 2013a; Mills et al., 2014) (Fig. 1).
Mounting geological evidence suggests abundant O2 deeper in Mars' history. In particular, the observation of enriched manganese oxide minerals by Curiosity points to strongly oxidizing conditions in the presence of liquid water some time in the past (Lanza et al., 2014, 2016). While manganese is an abundant component of the crust of both Earth and Mars (Turekian and Wedepohl, 1961; Taylor and McLennan, 2009), it requires strongly oxidizing conditions to be oxidized and concentrated in sedimentary rocks—and even then, this process is sluggish and hence suggests a prolonged process (Morgan, 2005; Luther, 2010). As a result, manganese-rich sedimentary rocks do not appear on Earth until after the evolution of photosynthesis around the rise of oxygen (Maynard, 2010; Johnson et al., 2013).
Additionally, it is possible that O2-rich fluids could exist on Mars even under the low modern atmospheric abundance, without invoking higher atmospheric O2 concentrations in Mars' past. We have previously shown that even on modern-day Mars the ability of low-temperature brines to contain large concentrations of O2 may result in fluids with sufficiently high dissolved O2 to drive oxidative weathering and to support aerobic respiration by microbes and even basal animals like sponges (Stamenković et al., 2018). These O2-rich brines could serve as localized oxygen reservoirs for any aerobic organisms that exist on Mars wherever they occur, on the surface or in the subsurface.
4. O2 on Europa
Europa is uniquely positioned in the outer Solar System as a promising candidate for providing an environment habitable to aerobic life with the intriguing caveat that any europan biosphere would likely not be supported by photosynthesis but instead by lithotrophic metabolisms fueled by redox chemistry between oxidants produced at the surface and reductants sourced from water/rock interactions at the base of the ocean (Chyba and Hand, 2001).
Europa is thought to host a subsurface ocean with substantial amounts of O2 sourced from ice radiolysis (Hand et al., 2007, 2009). It is generally accepted that Europa possesses an extensive subsurface ocean potentially containing 2–3 times the volume of water on Earth (Anderson et al., 1998; Carr et al., 1998; Kivelson et al., 2000; Stevenson, 2000). This ocean is predicted to be salty, based on magnetic field measurements (Khurana et al., 1998; Kivelson et al., 2000; Hand and Chyba, 2007), potentially dominated by magnesium sulfate (Zolotov and Shock, 2001). The base of Europa's ocean is predicted to be in contact with the rocky core, where water/rock interactions can provide a source of hydrogen and other reduced compounds through serpentinization-like reactions or high-temperature hydrothermalism (Anderson et al., 1998; Vance et al., 2007). These observations suggest a geologically active world with sources of both reduced and oxidized compounds that could potentially support a biosphere in the ice-covered ocean (McCollom, 1999; Russell et al., 2017).
Detection of a tenuous O2 atmosphere of ∼10−11 bar was first made from Hubble Space Telescope observations (Hall et al., 1995), with the dominant source of O2 and other oxidants in the Europa system being bombardment of water and CO2 ice on the surface by energetic electrons and ions, such as iogenic sulfur, driven by Jupiter's magnetic field (Johnson et al., 2004). As these radiolytically sourced oxidants can be trapped in ice (Spencer and Calvin, 2002) and potentially delivered into Europa's ocean (e.g., Kattenhorn and Prockter, 2014), this radiolytic source could ultimately serve as a substantial source of abiotic O2 to any biosphere that may exist on Europa. The potential for biologically relevant O2 accumulation in the subsurface ocean has been described previously (e.g., Chyba, 2000; Hand et al., 2007). While the rate of O2 production at the surface of Europa can be reasonably well estimated (e.g., Chyba and Phillips, 2001), the rate of resurfacing and of admixture of O2 into the subsurface ocean is not known but will ultimately determine the flux of O2. Estimates of resurfacing rates have been made based on crater counts and assuming subduction-like recycling of the ice shell into the interior, suggesting a resurfacing time of <90 million years (Zahnle et al., 2003; Kattenhorn and Prockter, 2014). Direct cycling of O2 into the ocean on this timescale could yield a delivery flux of O2 >109 mol/year (Hand et al., 2007).
Depending on the rate at which Europa's ice shell is mixed with the ocean, as well as the rates at which this O2 is consumed, it may accumulate to concentrations higher than those in Earth's oceanic oxygen minimum zones, in which multicellular life-forms like polychaete worms exist (Hand et al., 2007, 2009). Interestingly, in cases of vigorous, rapid cycling of Europa's ice shell with its ocean, O2 concentrations could potentially exceed Earth's oceanic O2 concentration (Hand et al., 2009). O2 concentrations in these environments might therefore be sufficiently high to not impose any limitations on aerobic respiration, though any net global productivity would instead likely be due to limited fluxes of reducing power from water/rock reactions at the base of the subsurface ocean (McCollom, 1999; Ward et al., 2017). Improved understanding of the O2 requirements and metabolic efficiencies of aerobic chemolithotrophic metabolisms could allow improved predictions of gross primary production, net primary production, and standing biomass, which could potentially be supported on icy moons like Europa.
5. O2 on Rocky Exoplanets
So far, the only exoplanet atmospheres in which O2 has been detected are of nonhabitable hot Jupiters like HD 189733b or HD 209458b (e.g., Ben-Jaffel and Ballester, 2013). Hot Jupiters are giant gaseous planets orbiting their host stars on very close orbits—resulting in hot equilibrium temperatures and puffy atmospheres, where large scale heights aid in characterizing atmospheric species like O2. Thus far, there have been no detections of O2 on potentially habitable cooler rocky exoplanets, where atmospheric scale heights are much smaller and hence spectroscopic identification of any gas is much more difficult.
From an early exoplanet perspective, O2 has been considered the holy grail of biosignatures in the atmospheres of rocky exoplanets—indicating the presence of photosynthesis (e.g., Owen, 1980). However, various potential abiotic pathways have been discovered since that could possibly generate variable quantities of O2 on rocky exoplanets (for a review, see Meadows et al., 2018). The primary causes for possible abiotic O2 buildup on exoplanets are either (1) CO2 photolysis and a reduced recombination of CO and O in high FUV/NUV environments common around M dwarfs (Tian et al., 2014; Gao et al., 2015; Harman et al., 2015); (2) water photolysis in atmospheres that are poor in noncondensing gases (such as N2 and Ar) making cold-trap mechanisms that prevent water from being dissociated on Earth ineffective around any star type (Wordsworth and Pierrehumbert, 2014); and (3) an extreme runaway greenhouse phase for M dwarfs during their extended pre-main sequence phase (Luger and Barnes, 2015).
M dwarfs are the most frequent stellar type in the galaxy and good candidates for abiotic oxygen formation due to their larger FUV/NUV ratios (see Tian et al., 2014; Gao et al., 2015; Harman et al., 2015) and an extended pre-main sequence phase (Luger and Barnes, 2015). This makes the recently discovered ultracool M dwarf TRAPPIST-1 system (Gillon et al., 2017) of seven roughly Earth-sized or smaller planets with low planet densities and a nondetection of hydrogen-dominated atmospheres (both findings together suggest the possible existence of water; De Wit et al., 2018) an ideal opportunity to potentially observe abiogenic O2-rich atmospheres.
The abiotic O2-forming scenarios mentioned above all need either atmospheres initially enriched in water vapor to be photodissociated (Wordsworth and Pierrehumbert, 2014; Luger and Barnes, 2015) or sufficient CO2 and small atmospheric hydrogen fractions to effectively maintain the photodissociation of CO2 and the formation of large amounts of O2 (Tian et al., 2014; Gao et al., 2015; Harman et al., 2015).
Moreover, O2 accumulation is only effective if there are insufficient sinks for O2 and O2 escape rates are small. Nonbiological sinks could be reducing volcanic gases, rocks, and lava on a geodynamically active planet. Hence if we detect exoplanet atmospheres with coexisting liquid water and O2, then these would make intriguing candidates for extensive characterization and attempts at the detection of (aerobic) life—under the condition that we simultaneously model the geodynamic activity and availability of reducing sinks for such worlds assuming large uncertainties in interior composition and structure (see Stamenković and Seager, 2016, Stamenković et al., 2016, and Bourrier et al., 2017, where such an approach is developed and illustrated).
Potential geophysical O2 sinks are nonperiodic and nonseasonal; hence seasonal atmospheric variability in O2 content (beyond any seasonal variability, such as by CO2 freezing on cooler planets like Mars) could be indicative of aerobic respiration and would be key in distinguishing dead from aerobic exoplanets.
6. Potentially Viable Metabolisms Fueled by Abiotic O2 Sources
Molecular oxygen is, thermodynamically, an incredibly favorable electron acceptor for a range of metabolisms—not just organoheterotrophy (i.e., organisms that consume fixed organic matter as a carbon and energy source) but also aerobic lithoautotrophy (i.e., organisms that oxidize inorganic compounds as an energy source to drive carbon fixation). This latter set of metabolisms allows aerobic respiration to fuel primary production—allowing O2-fueled biospheres to develop in the absence of photosynthesis. To understand the potential for biospheres fueled by O2, it is important to constrain the range of metabolisms that O2 could fuel in different planetary contexts and the amount of productivity these metabolisms could support.
The mounting evidence for liquid water throughout Mars' past as well as potentially on the modern near-surface (DiBiase et al., 2013; McEwen et al., 2014; Grotzinger et al., 2015; Martin-Torres et al., 2015; Ojha et al., 2015), combined with the availability of free O2, suggests a history of martian environments that are not only potentially habitable but also present a variety of thermodynamically favorable metabolic couples. For instance, the initial putative discovery of methane on Mars (Formisano et al., 2004; Mumma et al., 2009; Webster et al., 2015) along with metabolically useful O2 suggests the opportunity for aerobic methanotrophy, the metabolic oxidation of methane to CO2 using O2 while conserving energy, a process driven by diverse bacteria on Earth (e.g., Hanson and Hanson, 1996). This metabolism requires O2 for the activation of methane and typically couples this process to aerobic respiration (Hanson and Hanson, 1996). Recently, however, it has been discovered that the terminal electron accepting process for this metabolism is more versatile than previously realized, with the potential in some bacteria to couple aerobic methanotrophy to nitrate reduction when O2 concentrations are low (e.g., Kits et al., 2015; Skennerton et al., 2015). The recent discovery of nitrate in martian sediment (Stern et al., 2015) therefore additionally supports the potential viability of this metabolism in some martian environments.
Other potentially viable metabolisms on the martian surface include the aerobic oxidation of ferrous iron. Evidence of iron oxidation by O2 has been observed in Gale Crater (Hurowitz et al., 2017), indicating that this process has been thermodynamically feasible at times in Mars' history, potentially providing a niche for aerobic iron-oxidizing organisms. Because the oxidation of reduced compounds by O2 is often a competition between the spontaneous abiotic reaction and microbes performing this process for energy conservation, microbes have adapted to microaerobic conditions where the abiotic rate is sufficiently sluggish. Iron-oxidizing bacteria typically grow under microaerobic conditions below 50 μM O2 and have been demonstrated to grow at O2 concentrations as low as 3 μM (Chan et al., 2016). The relatively low biomass yield of aerobic iron oxidation relative to phototrophy (Ward et al., 2017) may make life supported by this metabolism more difficult to detect. However, the metabolic by-product of iron oxidation is solid ferric iron oxides; iron-oxidizing bacteria must shed ferric minerals, typically in diagnostic, preservable morphologies such as sheaths and stalks (e.g., Chan et al., 2011). If similar microbes were present on Mars, iron biominerals could be observable biosignatures (Banfield et al., 2001).
Extensive water/rock reactions at the base of Europa's ocean have been predicted, potentially supporting ecosystems analogous to hydrothermal vents on Earth by providing a flux of reduced compounds, such as molecular hydrogen, ferrous iron, and/or sulfide (e.g., Zolotov and Shock, 2004). The interaction of reduced compounds from water/rock interactions at the interface of Europa's ocean and rocky core with O2 sourced from the icy shell could support aerobic lithoautotrophic metabolisms, such as hydrogen oxidation, sulfur oxidation, or ferrous iron oxidation.
Molecular oxygen is furthermore essential for the activation of otherwise inaccessible electron donors. Compounds such as methane and ammonia must first be activated utilizing O2 via a monooxygenase enzyme in a non-energy-conserving step before they can subsequently be further oxidized to conserve energy (Hanson and Hanson, 1996). While these processes have an absolute requirement for O2, they can make use of exceedingly low O2 concentrations. Half saturation constants for ammonia oxidation are on the order of ∼300 nM O2 (Bristow et al., 2016), and both methane and ammonia oxidation have been observed in environments with much less than 1 μM O2 (Hatzenpichler, 2012; Kits et al., 2015; Bristow et al., 2016). Moreover, intracellular O2 production via nitric oxide dismutation has been suggested for Methylomirabilis oxyfera, an aerobic methanotrophic bacterium, allowing aerobic methanotrophy without detectable free O2 (Ettwig et al., 2010), highlighting the vanishingly low O2 requirements of these metabolisms.
7. Potential Challenges Regarding Abiotic O2 and the Evolution of Complex Life
A potential caveat to abiotic O2 improving the odds of evolution of complex life is the impact of oxidizing conditions on the origin of life. While conditions necessary and sufficient for the origin of life are not yet known, many proposed environments for the origin of life, such as hydrothermal vents, are characterized by reducing conditions (e.g., Baross and Hoffman, 1985; Martin et al., 2008). It is therefore possible that oxidizing conditions as a result of abiotic O2 as proposed here could inhibit the origin of life, or at least restrict the environments in which it could occur to locally reduced conditions such as adjacent to hydrothermal vents or in redox stratified lakes, such as the one that once occurred in Gale Crater, Mars (Hurowitz et al., 2017). Abiotic O2 could therefore be an evolutionary Catch-22: it provides multiple fast paths for the evolution of complex multicellularity but could challenge the origins of requisite earlier and simpler biology.
Despite the energetic benefits of aerobic respiration, O2 and related oxidants are in many ways toxic to biology. Life on Earth has therefore evolved multiple strategies to tolerate oxidative stress. These adaptations were likely absent on early Earth, before the evolution of oxygenic photosynthesis, so the introduction of O2 to previously anoxic biology may have relied on simpler innate detoxification and protection mechanisms until complex protein-based systems evolved (Fischer et al., 2016b). The radiation of Cyanobacteria and the extinction of other lineages associated with the introduction of toxic O2 may also have led to an evolutionary bottleneck alongside the origin of oxygenic photosynthesis (Sleep, 2018). Today, the abundance of O2 has resulted in the presence of genes related to oxidative stress in essentially all organisms, including strict anaerobes (e.g., Baughn and Malamy, 2004; Hemp et al., 2015; Pace et al., 2015; Ward et al., 2018). On a planet where life first appeared in an oxidizing environment, tolerance or avoidance of oxidative stress would be an essential early step in the origin and expansion of life.
If life did originate on a planet with substantial abiotic O2 levels, the production flux of O2 may prove limiting. The O2 in Mars' atmosphere is likely produced at low rates, and the O2 on Europa may be on the order of 109 to 1010 moles O2/year (Hand et al., 2007)—values that are many orders of magnitude lower than biogenic O2 production on Earth (Field et al., 1998). Unchecked aerobic respiration would therefore lead to rapid depletion of the standing pools of oxidants. On Earth, O2 production and consumption are tightly coupled, partially due to geochemical feedbacks but primarily by aerobic respiration (Ward et al., 2016). Biological feedbacks may therefore maintain O2 abundances on other worlds albeit at respiration rates proportional to O2 production rates. These relatively low rates of abiotic O2 consumption may limit the energetic yield, and therefore carbon fixation flux, of any life that depends on aerobic lithoautotrophy for primary production. The rates, efficiency, and yield of lithotrophic metabolisms relative to phototrophy are typically low (e.g., Ward et al., 2017, 2019). On Earth, the productivity of the biosphere before the evolution of photosynthesis was very low, potentially limited to less than 0.01% of the rates of modern primary productivity, based on the availability of electron donor compounds and the efficiency of their use by lithotrophic organisms such as methanogens (Ward et al., 2019). Similar limitations may have significant implications for the potential size and productivity of any europan biosphere. As a result, the potential for the origin and survival of a substantial biosphere is contested (e.g., Gaidos et al., 1999). These limitations would imply biological productivity much lower than that seen on Earth today, potentially at rates too low to be readily observable, though concentration of life at strong redox gradients or energetic hotspots (e.g., the bottom of Europa's ice shell; Russell et al., 2017) may help alleviate this difficulty. It is also conceivable that abiotic O2 could jump-start the evolution of life (Fig. 3), allowing an initial boost in productivity and development until the abiotic O2 pool has been depleted—the further evolution of life would then either be restrained by abiotic O2 production rates or could be maintained if this initial period of evolutionary innovation provided an opportunity for the evolution of oxygenic photosynthesis. Another possibility is that low background rates of biological productivity could be interspersed with short-lived bursts of activity driven by geological events such as seismic release of H2 accumulated in the subsurface (e.g., Sherwood Lollar et al., 2007; Sleep and Zoback, 2007).
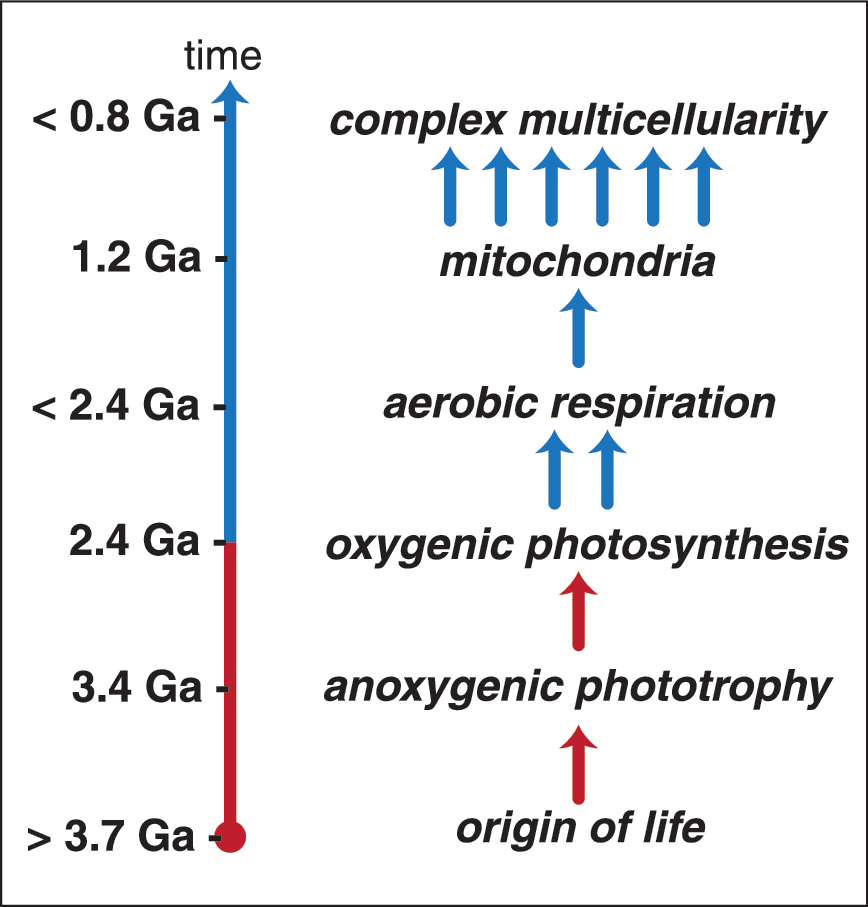
Evolutionary contingency and the origin of complex life. Several steps had to occur for complex life to evolve on Earth. Beginning with the origin of life, evolution of anoxygenic phototrophy, coupling in series of two reaction centers to perform oxygenic photosynthesis, evolution of aerobic respiration, evolution of eukaryotes and endosymbiosis of the mitochondrion, and finally the evolution of complex multicellularity. Of these, some steps have occurred independently multiple times, while others are evolutionary singularities and therefore may represent “rare, hard steps” whose evolutionary difficulty may delay or prevent the development of complex life. On a world with a significant abiotic O2 flux, life may be able to skip the hard step of evolving oxygenic photosynthesis (shown in red) and jump-start the evolution of complexity (shown in blue).
8. Conclusions
The presence and behavior of O2 provides an interesting perspective on habitability, but the history of O2 on Earth is, fundamentally, a biological one. O2 in our atmosphere is produced in staggering quantities—around ∼8.75 × 1016 moles of O2 per year (Field et al., 1998)—as a result of oxygenic photosynthesis by members of Cyanobacteria (i.e., Oxyphotobacteria) and their descendants (i.e., chloroplasts in algae and plants). This O2 is produced rapidly but is kept in balance by being nearly perfectly recycled by aerobic respiration (Ward et al., 2016). This state of affairs has continued for the past 2.3 billion years and has supported the radiation of many microbial groups, the origin and diversification of eukaryotes, and, eventually, complex multicellular organisms like plants, animals, and fungi. Before the rise of oxygenic photosynthesis, Earth was essentially anoxic and supported only microbial life with very limited productivity. The development of complex life therefore required multiple preconditions, some of which appear to have been very challenging for biology to evolve. Among these, oxygenic photosynthesis may be one of the hardest steps (Fischer et al., 2016a). This metabolism is biochemically complicated, required a series of evolutionary contingencies to diverge and recombine two photosystems, and has evolved only a single time in Earth's history. In comparison, comparative biological data show that aerobic respiration has multiple independent origins (heme copper oxidases, alternative oxidases, and bd oxidases), and multicellularity has even more (e.g., animals, plants, multiple fungi, brown algae, and red algae). The relatively late evolution of oxygenic photosynthesis may therefore have delayed the origin of complex life on Earth for a billion years or more.
It seems feasible that O2-rich environments could be produced abiotically on other planets and moons more efficiently than they were on early Earth—from Mars and Europa to exoplanets orbiting M dwarfs, as in the TRAPPIST-1 system. Therefore, aerobic respiration—and the energetic, complex organisms that depend on it—may be capable of evolving more readily. The presence of O2 in a planet's atmosphere, oceans, or local surface or subsurface brines may therefore be an important indicator of habitability (though clearly not an indicator of photosynthesis), as abiotic oxygen may provide a path to jump-starting the emergence of complex life. We therefore see value in a strategy of “Follow the Oxygen,” in which environments with opportunities for energetic metabolisms, such as aerobic respiration, are preferentially targeted for investigation and life detection. If life has appeared in these environments, it may be more likely to have developed more quickly into more complex forms than on Earth-like worlds where oxygenic photosynthesis was a necessary precondition for highly energetic organisms.
While targeting life-detection strategies will become more important as exoplanet discovery continues to accelerate and opportunities for detailed characterization remain limited, our hypothesized role for abiotic O2 in jump-starting biological evolution has test cases within our own solar system: Europa and Mars. Europa presents evidence for redox opportunities for life and a generally habitable environment all coexisting in an ocean where photosynthesis is impossible but O2 may accumulate abiotically. Determining whether life exists on Europa—and if it has developed complexity beyond that present on pre-oxygenic photosynthesis Earth (e.g., eukaryogenesis, multicellularity)—will therefore be a valuable test of our hypothesis, particularly in contrast to Enceladus, which appears to have many ingredients for habitability but lacks mechanisms to accumulate oxidants like O2 (Parkinson et al., 2008). Meanwhile, Mars may provide localized environments habitable to aerobic life (likely in the subsurface), where atmospheric O2 fluxes overlap with geological fluxes of reducing compounds such as hydrogen or methane produced by water-rock reactions such as serpentinization and Fischer-Tropsch-type reactions.
Footnotes
Acknowledgments
This work was performed in part at the Jet Propulsion Laboratory, California Institute of Technology, under contract to NASA. © 2018, California Institute of Technology.
The authors would like to acknowledge funding support from the Simons Collaboration on the Origin of Life (V.S. and W.W.F.), a NASA NESSF fellowship to L.M.W. (NNX16AP39H), the Agouron institute (L.M.W. and W.W.F.), and a NASA Exobiology grant to W.W.F. (NNX16AJ57G).
The authors would like to thank Norm Sleep and two anonymous reviewers for helpful comments that have improved the manuscript.
Associate Editor: Victoria Meadows