
Abstract
As discoveries of multiple planets in the habitable zone of their parent star mount, developing analytical techniques to quantify extrasolar intra-system panspermia will become increasingly important. Here, we provide user-friendly prescriptions that describe the asteroid impact characteristics which would be necessary to transport life both inwards and outwards within these systems within a single framework. Our focus is on projectile generation and delivery, and our expressions are algebraic, eliminating the need for the solution of differential equations. We derive a probability distribution function for life-bearing debris to reach a planetary orbit and describe the survival of microorganisms during planetary ejection, their journey through interplanetary space, and atmospheric entry.
1. Introduction
A
Despite these advances, the applicability of these processes to extrasolar planetary systems is still in question, partly because in those systems we lack the detailed knowledge of our own planetary system. Nevertheless, efforts to characterize panspermia between different extrasolar systems, or between the Solar System and extrasolar systems, have contributed to our understanding (Adams and Spergel, 2005; Valtonen et al., 2009; Wesson, 2010; Belbruno et al., 2012; Lin and Loeb, 2015; Gale and Wandel, 2017). However, panspermia among extrasolar planets within the same system has received a relatively little but increasing amount of attention (Heller and Armstrong, 2014; Steffen and Li, 2016; Krijt et al., 2017; Lingam and Loeb, 2017a). A potential reason for this relative dearth of studies is the lack of observational evidence of multiple planets in the habitable zone of the same star.
This situation has now changed with the groundbreaking discovery of multiple potentially habitable planets in the TRAPPIST-1 system (Gillon et al., 2017). TRAPPIST-1 is an M dwarf with a mass of 0.08 M ⊙ that harbors seven observed transiting planets all with masses similar to the Earth and three (planets e, f and g) which are securely in the star's habitable zone (although all seven may be, with effective temperatures ranging from about 150 K to 400 K). Because all seven planets are seen transiting from the Earth, their orbits are nearly coplanar. The system is compact (all planets could fit well within Mercury's orbit) and is likely to be resonantly interacting in a long chain (Luger et al., 2017). We do not yet know if an equivalent of the Late Heavy Bombardment event (believed to have occurred in the Solar system, even if less intense than originally thought; Bottke and Norman, 2017) has or will occur in that system, nor what types of potential impactors lurk beyond the most distant planet (h) and outside of our field of view. Despite the uncertainties, investigation of panspermia from within this system has already been undertaken (Krijt et al., 2017; Lingam and Loeb, 2017a) and might be prompted further by additional observations, which are currently underway.
Here, we study and derive several aspects of lithopanspermia in more general closely packed multiplanet systems, with a focus on analytics and dynamical delivery, but also addressing microorganism survival at each stage. Numerous Solar System studies have taught us that N-body simulations are both computationally expensive and dependent on a large number of parameters (Dones et al., 1999) which are unknown in exoplanetary systems. Computational times for most known extrasolar planetary systems would be worse because of their compact nature. Therefore, we adopt a purely analytical approach, one which could be applied to extrasolar systems with multiple habitable planets. The characterization of such systems is expected to increase steadily over the next decade, culminating with the PLATO mission (Rauer et al., 2014), which will measure habitable planets out to about 1 AU.
Throughout the paper, our subscript convention for physical quantities will be: i for the impactor, no subscript for the source planet, a single prime for the fragmented debris ejected from the source planet, and a double prime for the target planet. In Section 2, we establish our setup and describe how a life-bearing rock could be transferred between one planet and the orbit of another planet; Appendix A provides most of the intermediate equations required for this section, and Appendix B contains an extension with a fictitious template compact system which can be used for quick estimates. Section 3 then details the likelihood of that rock actually impacting a target planet. Section 4 constrains the characteristics of the ejecta that would both satisfy the dynamics and have the capability to harbor life. In Section 5, we consider the biological prospects of life surviving all aspects of lithopanspermia. We conclude in Section 6.
2. Orbit Transfers
2.1. Setup
Consider a pair of planets such that one, the “source,” contains a life-bearing organism, whereas the other, the “target,” initially does not. An impactor crashes into the source, producing a spray of life-bearing ejecta. By assuming that the ejecta is “kicked” impulsively, we estimate its direction and speed such that it would reach the orbit of the target. By impulsively, we refer to the timescale of the kick being much smaller than the orbital period of the source. In multiplanet systems within the detectability threshold of transit photometry surveys, planet orbital periods are on the order of days, whereas impact kick timescales would typically be on the order of minutes.
The underlying formalism we use was established in Jackson and Wyatt (2012) and expanded upon in Jackson et al. (2014), and similar to that in Appendix A of Le Feuvre and Wieczorek (2008). We briefly repeat here the geometrical setup in Jackson et al. (2014): Denote the kick speed as
For perspective, the circular speed of the TRAPPIST-1 planets are, from planet b to h moving outward, {79.9, 68.2, 57.5, 50.2, 43.7, 39.6, 33.9} km/s. The ratio
At impact, the source is assumed to lie at the pericenter of its orbit such that its argument of pericenter, longitude of ascending node, and inclination I are all zero.
1
The source is assumed to move counterclockwise, and the kick direction is defined by two variables: θ and
2.2. Ejecta orbit characteristics
The orbit of the ejecta, whose elements are denoted by primes, is related to the unprimed quantities (which refer to the source planet), through Equations A1–A8 (Jackson et al., 2014).
Because known compact multiplanet systems are dynamically “cold”—exhibiting low eccentricities—henceforth we make the approximation that all planets are on circular orbits. Doing so greatly simplifies the analysis. For example, the upper limits for the planetary eccentricities in the TRAPPIST-1 system are all under 0.085. Further, because for any compact multiplanet system with habitable plants we cannot yet know if panspermia has occurred, or will occur, at a particular time, we assume, without loss of generality, that the true anomaly f = 0. We hence constrain only when ejecta intersects the orbit of the target and use Equations A7 and A8 for this purpose.
2.3. Coplanarity restrictions
First however, consider that in order for ejecta to hit the target, the ejecta must coincide with the target in all three spatial dimensions. This intersection is most easily achieved if the orbital planes of the source, ejecta and target lie close to one another as we can then be guaranteed of coincidence in one of the three dimensions. Hence, in this section, we quantify how coplanar the orbits of the ejecta, source and target must be to achieve spatial coincidence in the vertical direction (direction of the angular momentum vector of the source) throughout the debris orbit.
This condition is mathematically equivalent to q′ sin I′ ⪅ R or Q′ sin I′ ⪅ R, when assuming that R denotes planet radius, q′ denotes orbital pericenter, Q′ denotes orbital apocentre, and the reference plane is the one which connects a coplanar source and target. Note that unless the longitudes of ascending node can be measured, the mutual inclinations of all source-target pairs in a particular system will remain unknown. 2 The approximations in these relations result from effects not considered here such as gravitational focusing and atmospheric drag.
If the kick direction is perpendicular to the source's angular momentum direction (θ = π/2), then the ejecta orbit will be coplanar with the reference orbit (I′ = 0). If, however, the kick direction deviates from these values, then the result is less obvious and is dependent on ϕ (and e when not assumed to be zero).
Figure 1 illustrates the resulting dependence of I′ on ϕ for two different values of θ, one where θ deviates from coplanarity by 10−3 degrees (left panel) and the other where the deviation is 0.5 degrees (right panel).
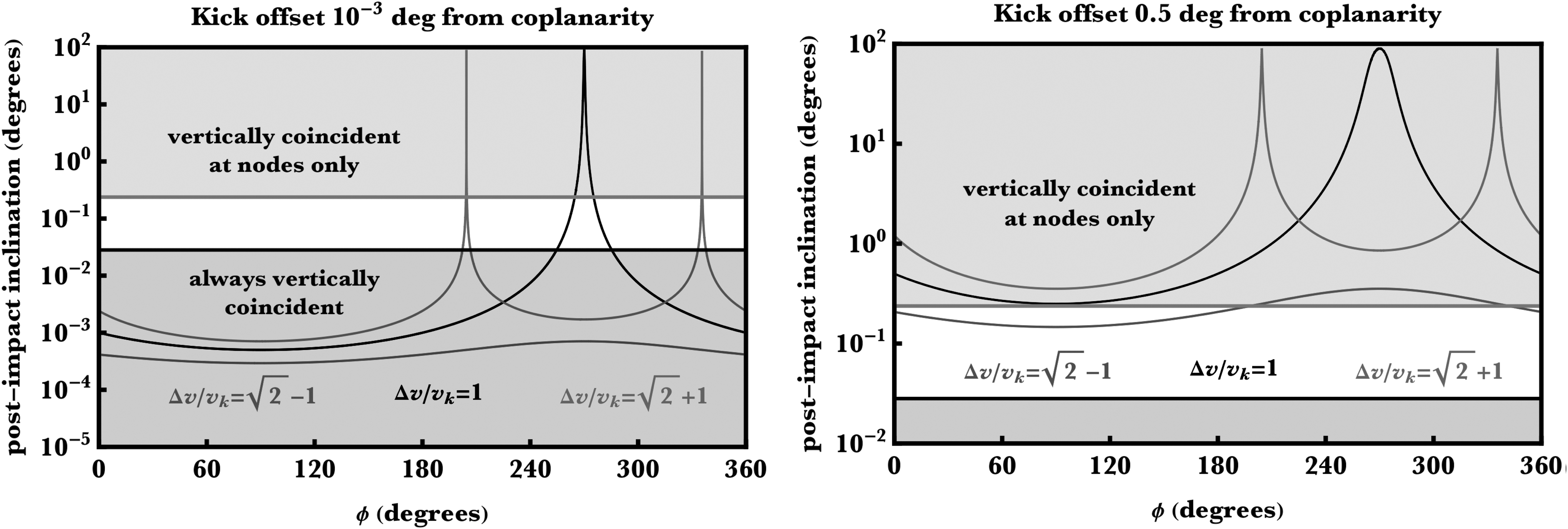
How the kick direction affects the inclination of the ejecta orbit. By assuming coplanarity amongst all TRAPPIST-1 planets, we plot, for three different values of the ratio of kick velocity to circular Keplerian velocity
To provide some context, we superimpose some results from the TRAPPIST-1 system on this figure. The gray region corresponds to where in the TRAPPIST-1 system I′ ≤ arcsin (min (a/R)) = 0.029° and the red region to where I′ ≥ arcsin (max (a/R)) = 0.24°. For this red region, the ejecta and target would be vertically coincident only near the nodes of their orbits, whereas for the gray region, they would be vertically coincident throughout their orbits (increasing the chances of impact). The purple curve, corresponding to
2.4. Other spatial restrictions
Having linked the angles of impact with the inclinations, we now turn to the other two spatial dimensions. The subsequent analysis greatly benefits from three simplifications, which are sufficient for this study. The first two are our continued assumption of circular and coplanar orbits. The third is that we consider the source and target planets in pairs, ignoring the influence of any other planets including those whose orbits lie in-between the pairs. This last assumption degrades in accuracy as the distance between the planets increases, because the debris will take longer to traverse this distance, and hence be diverted to a greater extent by extra bodies. However, these effects are sensitively dependent on the number, masses and locations of other planets in the system.
2.4.1. Intersecting orbits
In order for a collision to occur, the debris must be ejected onto an orbit which intersects the orbit of another planet. We can place bounds on this geometry by considering the pericenter and apocenter. Hence, we begin by expressing these quantities as a function of the impacted planet's semimajor axis, the velocity ratio and ϕ (Eqs. A9–A11). We wish to find the range of velocity kicks for which these pericenters or apocenters can be achieved. Hence, inverting the equations yields, for inward motion toward the orbital pericenter,
where
Recall that a and q′ refer, respectively, to the source planet's semimajor axis and the ejecta's orbital pericenter, and ϕ is the angle between the projection of the kick direction onto the source's orbital plane and the star-source pericenter line.
For panspermia outward from the star, no piecewise function is necessary, as in the following equation the denominator is always positive and the term in square brackets is always negative.
where
Equations 1–4 provide the necessary and sufficient conditions for kick speeds to propel ejecta into the orbit of another planet.
2.4.2. Application to TRAPPIST-1
To demonstrate how these equations can be applied to a real system, Figure 2 illustrates the application of Equation 1 to the TRAPPIST-1 system. The figure displays inward panspermia from planet h, as well as what minimum kick speeds are necessary to thrust the ejecta just into the orbits of planets g, f, e, d, c and b. Note that this value is highly dependent on ϕ and exceeds the system escape speed for many values of ϕ. Further, the dependence on ϕ is non-monotonic.
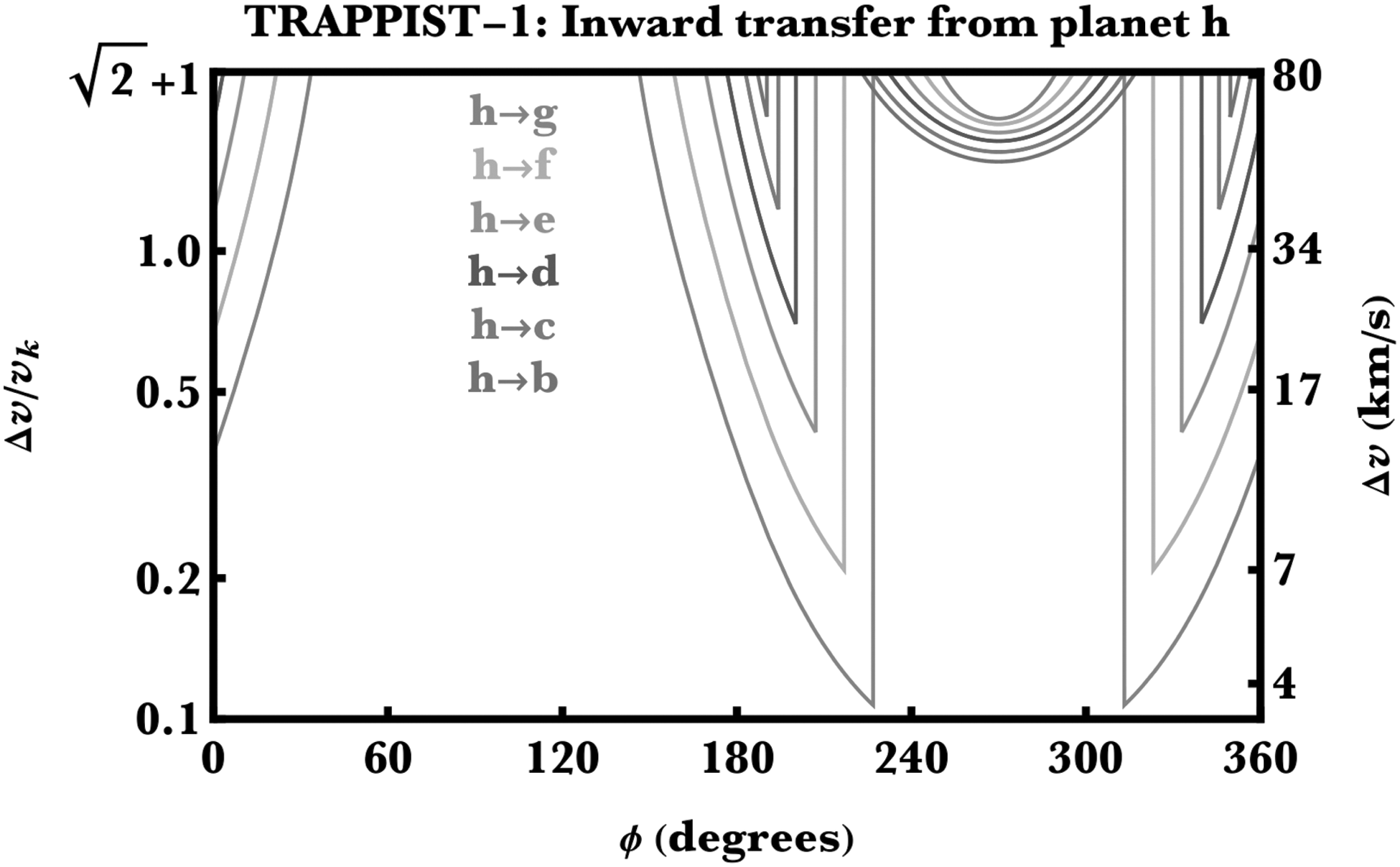
Inward panspermia in the TRAPPIST-1 system from planet h. Shown is the minimum speed kick required to place ejecta from planet h into the orbit of another planet, as a function of kick direction (ϕ). For many values of ϕ, the minimum kick speed exceeds the system escape speed (top axis of plot).
When we instead consider outward panspermia to planet h, the functional form changes. Application of Equation 3 yields Figure 3. Here, for all values of ϕ, the minimum kick speed required to propel the ejecta to another planet's orbit is smaller than the system escape speed. The smallest kick would be from planet h's nearest neighbor (planet g), whereas the greatest kick is required from the planet furthest away (planet b).

Outward panspermia in the TRAPPIST-1 system to planet h. Shown is the minimum speed kick required to place ejecta from a planet into the orbit of planet h, as a function of kick direction (ϕ). In all cases, the minimum kick speed is less than the system escape speed (top axis of plot).
Both Figures 2 and 3 do not take into account the escape speed from the planets themselves. The escape speeds of the TRAPPIST-1 planets are, from planet b to h moving outward, {9.90, 12.79, 8.15, 9.19, 9.02, 12.2, 9.35} km/s. The relative velocity of the ejecta after escape from the planet is likely to be comparable to the escape velocity of the planet.
2.4.3. When keeping ϕ fixed
If instead one has reason to assume a particular fixed value of ϕ, then Equations 1–4 may be considered as functions of
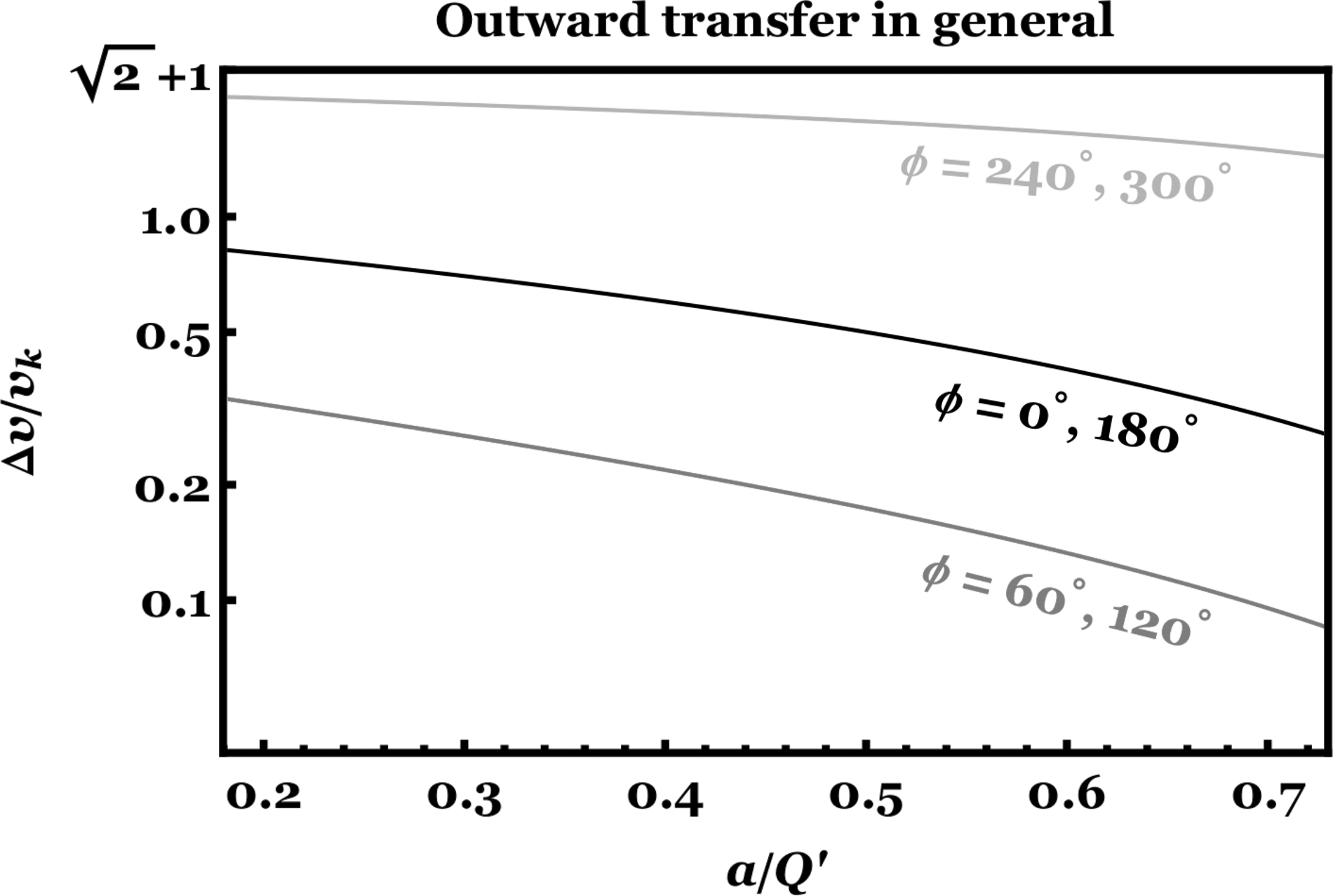
Outward panspermia as a function of (a/Q′) when ϕ is kept fixed. The x-axis includes the entire relevant range in the TRAPPIST-1 system; the upper end also roughly corresponds to outward panspermia from Earth's orbit to Mars' (assuming that they are on circular, coplanar orbits).
2.4.4. Curve extrema
Returning to Figures 2 and 3, to find the minimum value of these curves as a function of ϕ, we consider all possible extrema of Equation 1, which are found in Equation A12. For outward panspermia,
and
These equations explain why the system escape speed corresponding to
Alternatively, for inward panspermia, there are multiple real solutions:
The absolute minimum could arise from either Equations 8 or 9. Equation 8 is the absolute minimum when
2.4.5. Probability distributions
Given these constraints, we can now construct probability distribution functions for debris reaching the target planet's orbit. We obtain a probability P of a given ratio
Now we provide an example using these probability functions. We utilize the TRAPPIST-1 system both to be consistent with the previous applications and because fortuitously, planets b and h are sufficiently well-separated to allow us to sample a special case where
Figures 5 and 6 are the result. Note first that for inward panspermia, P never reaches unity, unlike for outward panspermia: the geometry responsible for these relations is folded into the curves. Second, for both directions, transferring to neighboring planets is easier than for those further away. Third, the kinks in Figure 5 ultimately result from the piecewise nature of the velocity ratio in Equation 1. The kink in the bottommost curve occurs at P = 0 because the separation of planets h and planet b is wide enough to cross the critical threshold mentioned in the last paragraph.
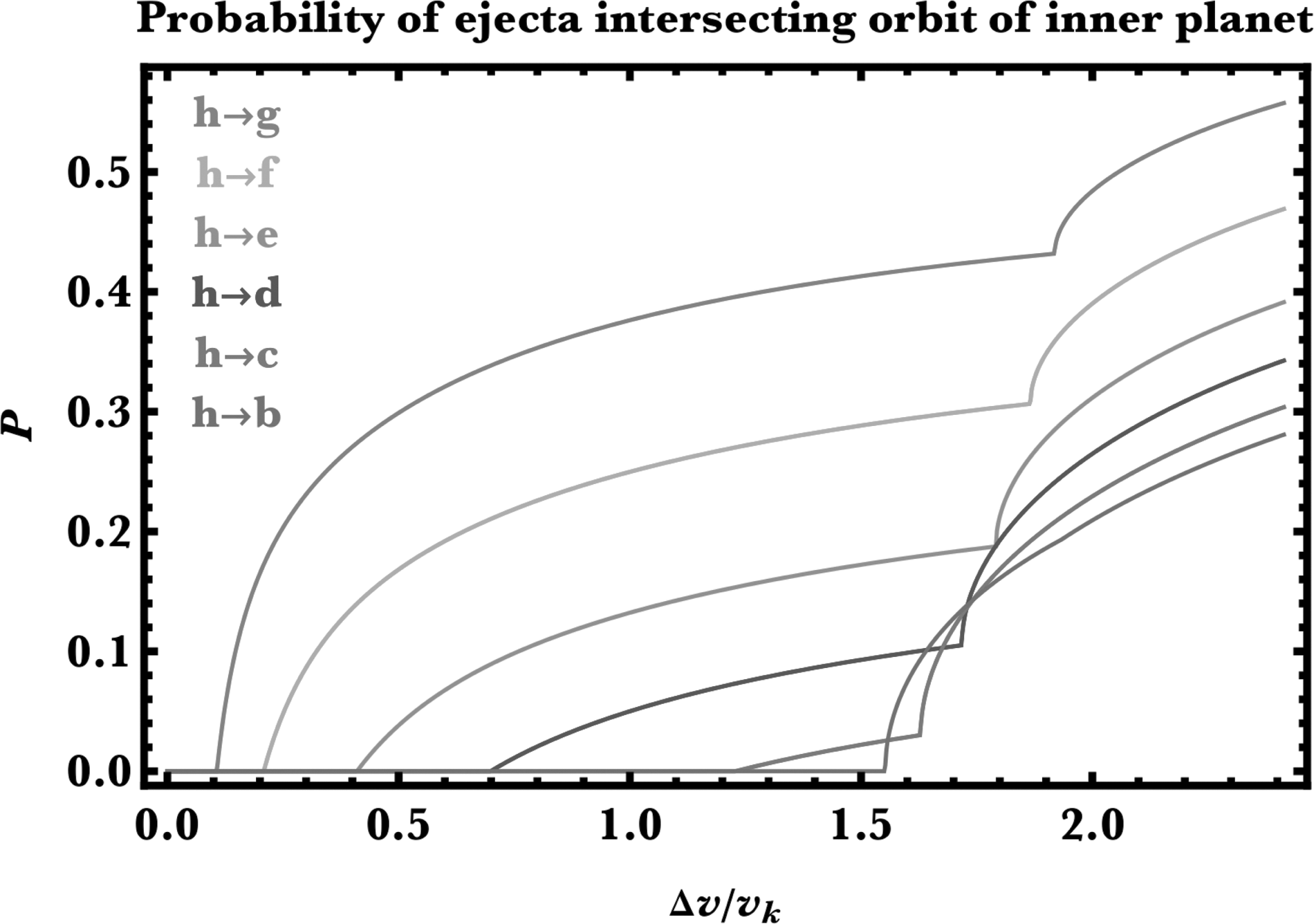
Application of the general algorithm to yield transfer probabilities (Eqs. A21–A24) to the TRAPPIST-1 system. Shown is inward panspermia in to planet h. The only variables needed to generate this plot were the semimajor axes of the planets.
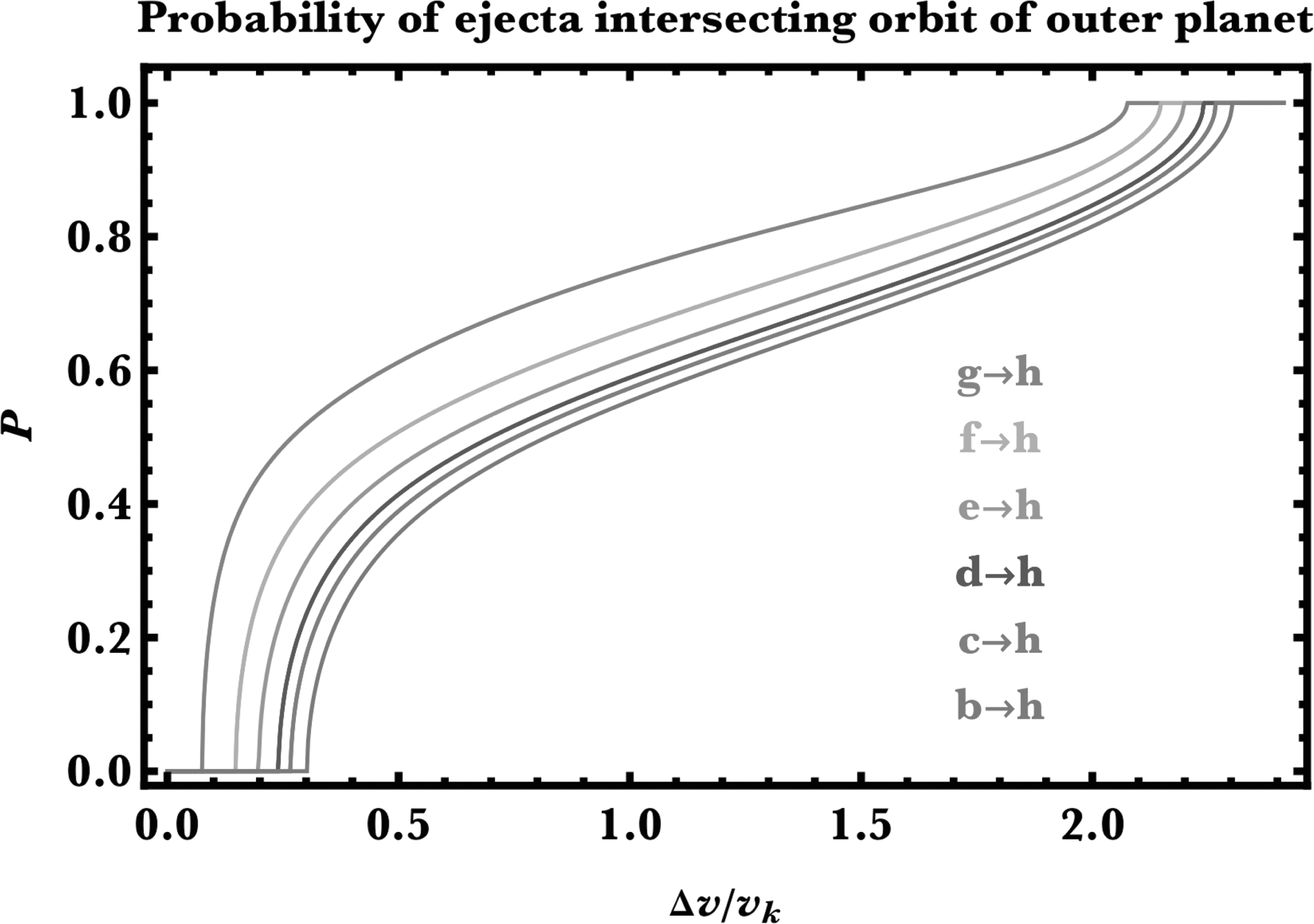
Similar to Figure 5, but for outward transfer of debris to the orbit of planet h (Eqs. A18–A20). For high-enough kick velocities, ejecta will always have the capability of colliding with an outer planet.
3. Impacting the Target in One Pass
In the last section, we placed bounds on orbital properties of ejecta that could hit the target planet. To better quantify the probability of actually impacting the target, we now consider the location of the debris in space, rather than just its orbit.
Assuming that the ejecta orbit intersects or almost intersects with the orbit of the target, and that these orbits remain fixed, then expressions exist for the probability of collision in one pass. These expressions, pioneered by Öpik (1951) and Wetherill (1967), have led to substantial and wide-ranging applications. A recent updated and simplified series of derivations was provided by JeongAhn and Malhotra (2017). They found in their equations 29 and 37 the probability of collision per revolution of the ejecta,
We now consider these dependencies in more detail. Because we assume that the target is on a circular orbit, the gravitational acceleration at the collision point is GM
star
/a″
2
(recall that the target's orbital parameters and mass are denoted with double primes). The collision radius is the sum of the radii of the ejecta and target multiplied by the gravitational focusing factor 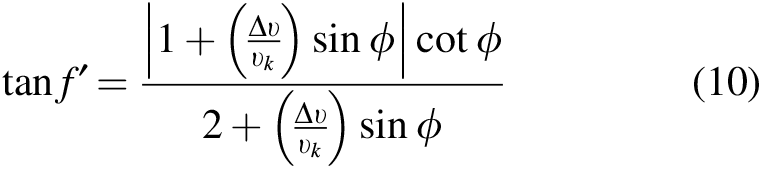
which leads to Figure 7.

Relating f′ to ϕ for different kick speeds.
Adding these various components above together leads to the following expression for the collision probability per time in our formalism. Assuming that the target and ejecta are much less massive than the parent star allows us to concisely write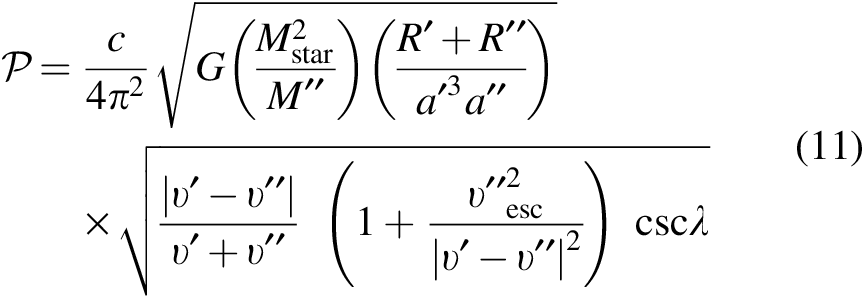
where c is a constant equal to either
The time taken for the debris to travel from the source to the target planet—a measure which influences the survival rates of microbes—may be approximated by the inverse of Equation 11. This transit time is hence strongly influenced by
4. Ejecta Characteristics
Alternatively, if we assume that all of the ejecta represents a single boulder or pebble of mass m′, then we can speculate on the characteristics of this piece of ejecta, and whether it could be life-bearing. In this section, we neglect more complex possibilities, such as indirect supplementary ejection from small source planets. Source planets the size of Ceres, for example, have low-enough escape velocities and surface pressures to be susceptible to glaciopanspermia (Houtkooper, 2011; Lingam and Loeb, 2017b). We also neglect the portion of the atmosphere—even for large source planets—that will invariably be ejected along with the debris (Berera, 2017). The debris can act as an agent to transfer the source planet's atmospheric constituents to the target. If the transfer of material is extensive enough, as during perhaps a period of heavy bombardment (de Niem et al., 2012; Bottke and Norman, 2017), then the atmosphere of the target planet may become less or more susceptible toward hosting life on its surface.
4.1. Escaping the atmosphere
First, consider that the radius and density of this piece of ejecta are related to m′ through m′ = (4π/3)ρ′R′
3
. This radius must be larger than the critical radius needed to maintain escape velocity from the source. If the source planet contains an atmosphere with surface pressure p and gravitational acceleration g, then the radius R′ of a piece of ejecta which could escape an atmosphere is (Artemieva and Ivanov, 2004)
where g = GM/R
2
such that any ejector could escape from an atmosphere-less source as long as
We plot Equation 13 in Figure 8 assuming that the source planet is an Earth-analog. The plot illustrates that atmospheric drag has the strongest effect when the post-impact speed is close to the escape speed, and flattens out as
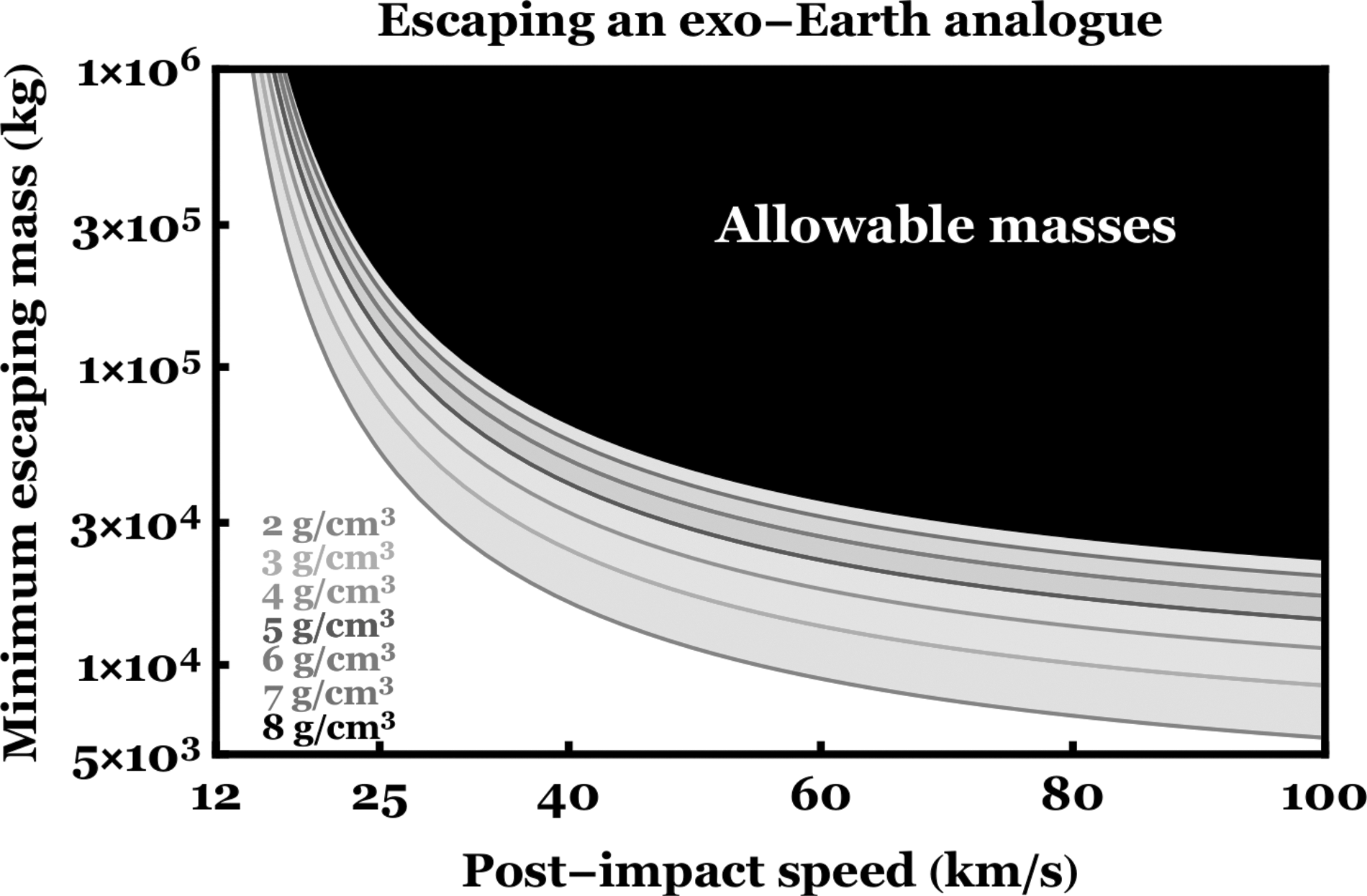
The minimum mass of an impact fragment that could be ejected from an Earth-analogue. The surface pressure (P), surface gravity (g) and density of the Earth (ρ) were assumed. Each curve represents a different impact fragment density (ρ′). The x-axis represents the post-impact speed
4.2. Largest fragment
Recall that m′ represents a fragment of the impact. Now we consider how to compute the largest possible size of an impact fragment, with radius
We just use (Mileikowsky et al., 2000)
where R
i and
We may relate the pre-impact and post-impact speed from Equation 14 by assuming that the energy of the impact is deposited at a depth which is comparable to R
i. Then (Melosh, 1984, 1988)
where d > R
i is the distance from the ejection point to the center of energy deposition.
3
Consequently, the ejecta speed cannot be larger than the impactor speed. Otherwise, the speed ratio is largely determined by the detailed physics of the impact. As we do not pursue such detail here, we suffice to leave Δv/v
i as a free parameter ranging from 0 to 1. Then, we can express Equation 14 for basalt (adopting ρ = 3 g/cm
3
) as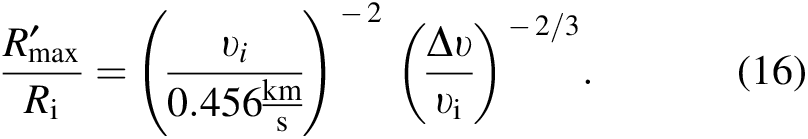
We plot Equation 16 in Figure 9. The plot illustrates that, in general, fragment radii are no more than about 5% of the radius of their progenitor.
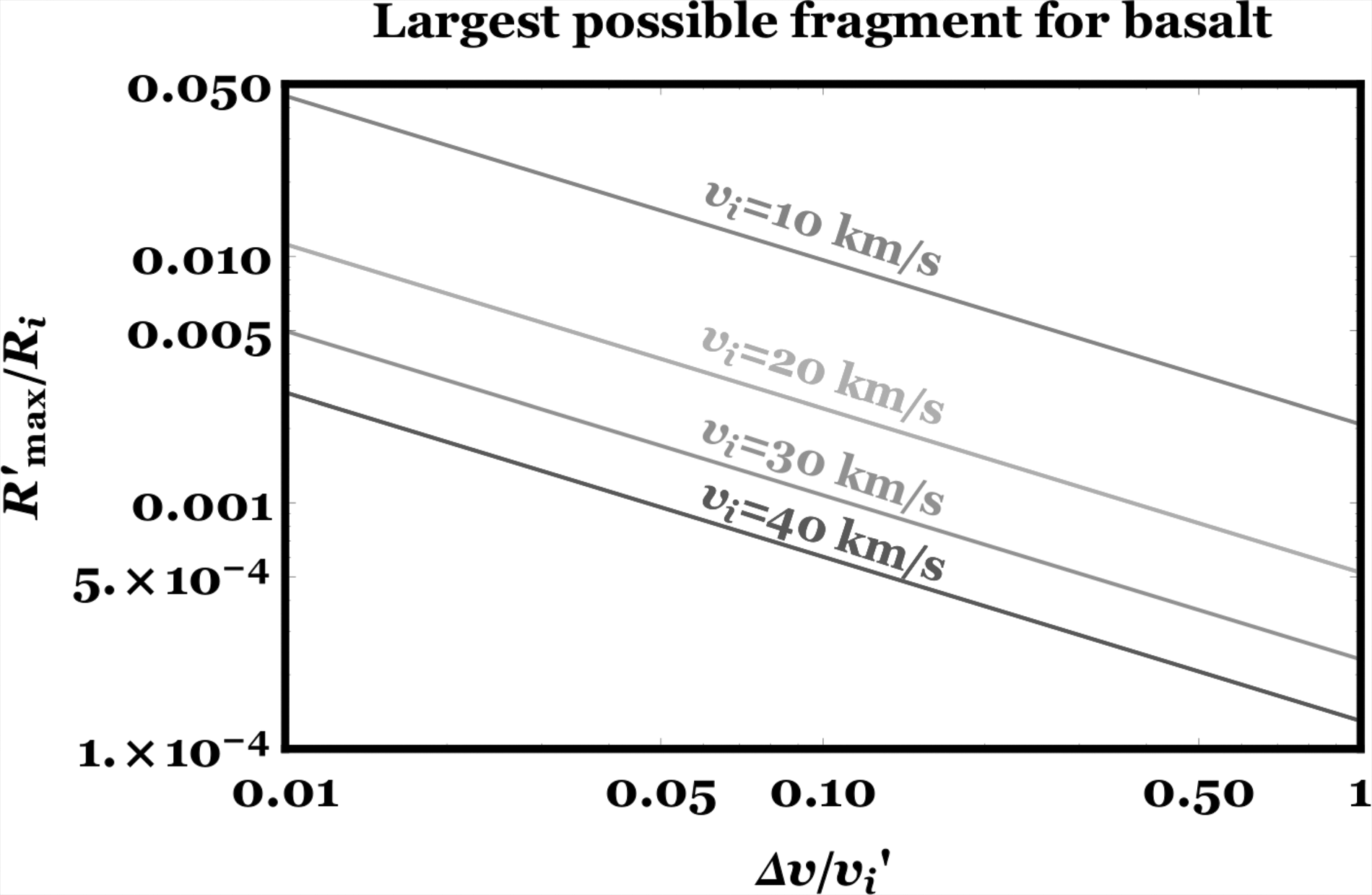
The maximum impact fragment size in terms of the initial radius of the impactor, for different incoming impactor speeds. The tension at fracture is assumed to be 0.1 × 109 Pa, and the density and Weibulls modulus are consistent with basalt.
4.3. Liberating material
Now we may consider the minimum impactor size that could liberate material. If
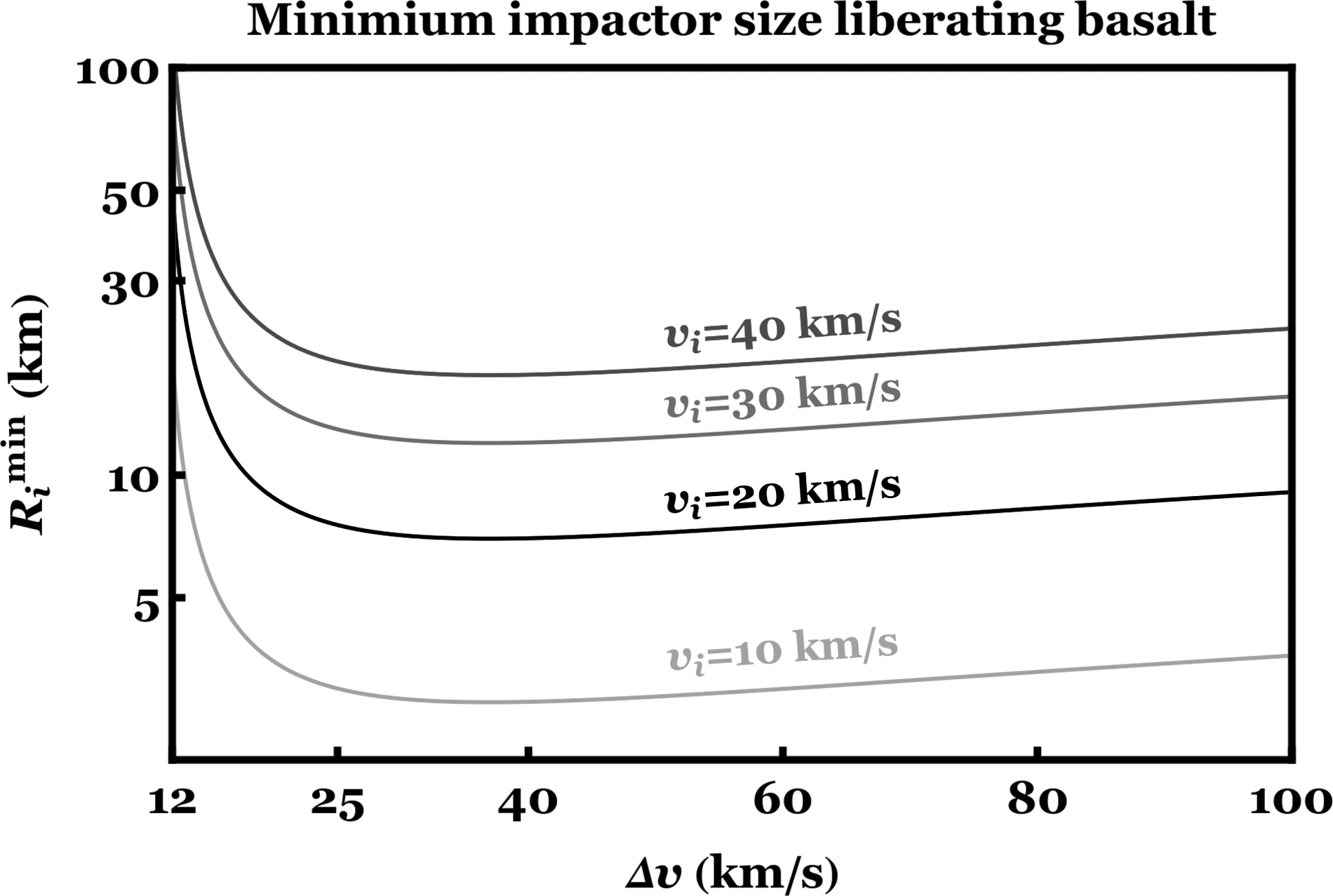
The minimum impactor size that could liberate material from the planet's gravitational well. The impactor and planet have densities of 3 g/cm 3 , which is consistent with basalt. The figure illustrates that panspermia could occur typically only if the impactor size is at least on the order of tens of km in size.
4.4. Destroying the source
Finally, if the impactor is too energetic, then it could sterilize, eviscerate or break apart the source planet. Any of these processes would inhibit prospects for panspermia. Here, we do not analyze the consequences in detail, but merely provide bounds for the impactor in the most extreme case of breakup.
We can place an upper bound on the maximum size of the impactor by considering the maximum specific energy E max imparted to a source for which the source will remain intact. The conditions for catastrophic disruption have an extensive associated literature and can be characterized through a variety of metrics. For example, see Benz and Asphaug (1999) and the hundreds of more recent papers which cite that one.
We adopt the explicit formalism in Section 5 of Movshovitz et al. (2016). They show that the source planet would break apart if the following condition is met:
where
with M
i being the mass of the impactor, and θ the impact angle, such that a head-on collision corresponds to θ = 0. The numerical range given in Equation 20 applies for
We can quantify Equations 17–20 in a simplistic but comprehensive manner by reducing the number of degrees of freedom in the equation. Assume that the collision is head-on and the impactor and source planet are made of the same substance (or more technically have equal densities). Then we can reduce the condition to a function of two ratios as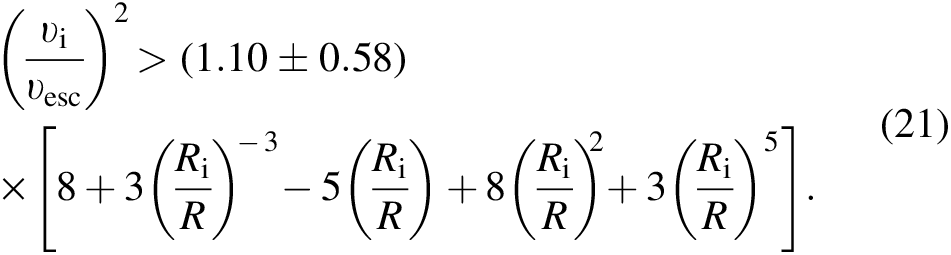
Figure 11 illustrates the phase space of Equation 21. The plot demonstrates that for common definitions of asteroid and planet, an asteroid could never destroy a planet. However, the result of a Mars-sized object colliding with an Earth-sized object is less clear.
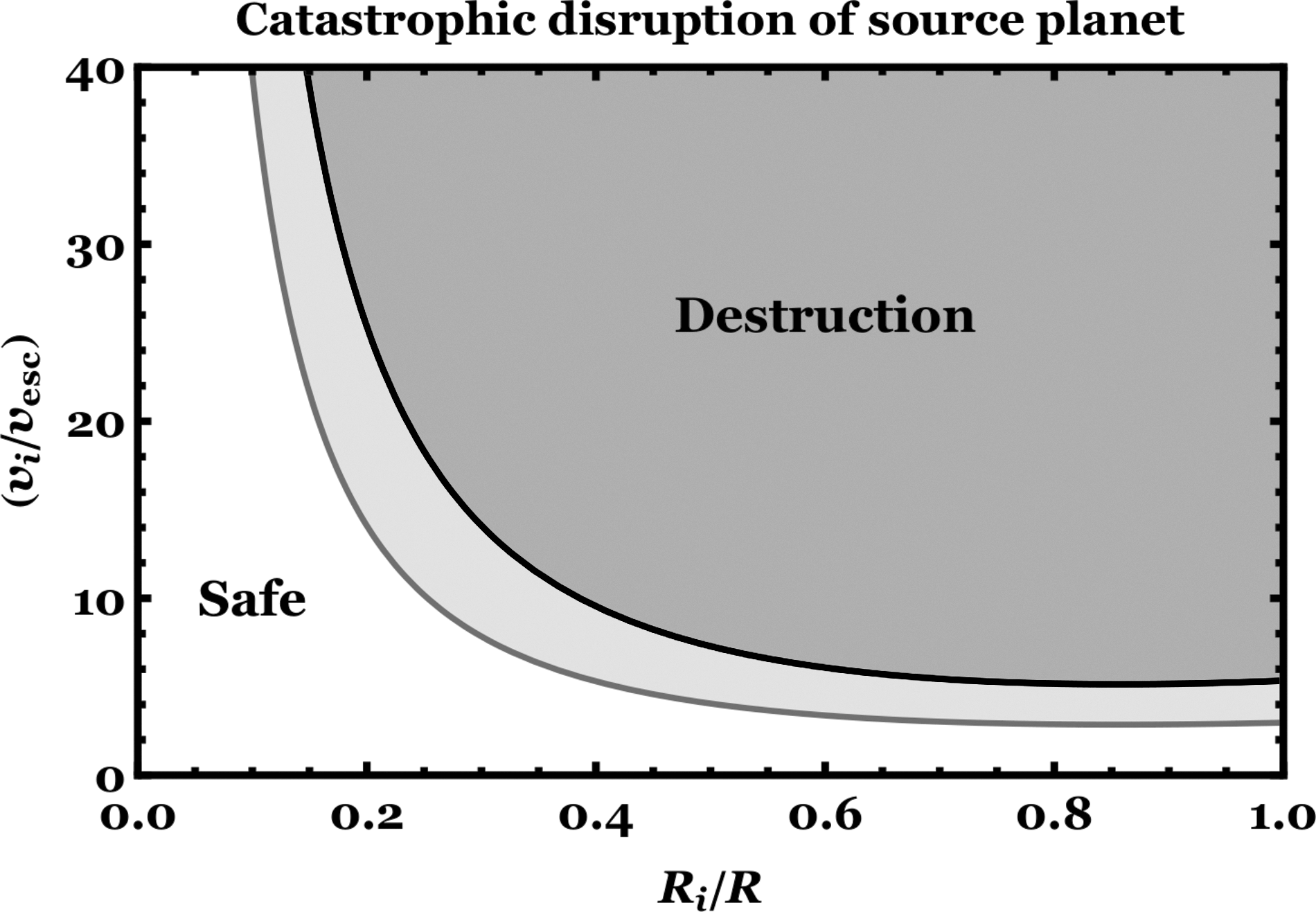
The speed and radius of an impactor that would catastrophically destroy the source planet. The collision is assumed to be head-on, and the impactor and source planet are assumed to be made of the same material. The two curves represent the bounds of the model prediction from the numerical coefficient of Equation 21. If the impactor radius is less than about 10% of the source planet's, then the required impact speed for a destructive collision would be unrealistically high.
4.5. Temperature at impact
We now have some idea of the size of potential ejectors. Returning our attention to panspermia, we must also consider temperature and how it will affect the ability of life to survive such an impact. Figure 4 of Weiss and Head (2016) illustrates averaged ejecta temperatures as a function of both crater diameter and surface heat fluxes on Mars. They find a range of 215–600 K for crater diameters between 0 and 150 km in roughly linear relationships for surface heat fluxes of 20–100 mW/m 2 . Therefore, the temperatures generated are a strong function of crater diameter.
Despite this wide range in temperature, simulations have shown that a significant proportion of the ejecta will not exceed 100°C due to the existence of a “spall” zone (Melosh, 1984). This zone comprises much of the surface layer undergoing the impact and refers to a region in which the shock wave from the impactor is effectively cancelled via superposition with its reflected counterpart. These relatively low-temperature fragments would offer more favorable conditions to any lifeforms residing upon them. The following section will discuss specific forms of life that have shown great promise when tested against the panspermia hypothesis, from initial ejection (Section 5.1) through interplanetary travel (Section 5.2) to eventual atmospheric entry upon reaching the target planet (Section 5.3).
5. Survival of Microorganisms
Over the past few decades, it has become possible to simulate the three stages of panspermia: (1) initial ejection from the impacted planet, (2) the subsequent journey through interplanetary space, and finally (3) impact with the target planet. Each step provides a new set of challenges to the survival of life. This section aims to supply a brief account of how certain microorganisms have fared when placed in environments that are reminiscent of the panspermia process and places these constraints into an extrasolar planetary context. A more comprehensive review of the near-Earth and Solar System contexts can be found in Horneck et al. (2010).
5.1. Planetary ejection
A precondition for ejecta to be produced is the existence of impactors. Not every exoplanet host star contains compact asteroid belts (Martin and Livio, 2013). However, evidence from white dwarf planetary systems indicates that between one quarter and one half of Milky Way white dwarfs host asteroid belts or Kuiper belt analogs (Koester et al., 2014) that are dynamically excited by a number of mechanisms involving planets (Veras, 2016), the same fraction of main sequence stars thought to host planets throughout the Galaxy (Cassan et al., 2012). 4 Further, the transfer of either biomolecules (pseudo-panspermia; Lingam and Loeb, 2017a) or dead organisms (necropanspermia; Wesson, 2010) imposes less stringent requirements than the transfer of living organisms.
In compact extrasolar systems, we might expect billions of rocks to be transferred between their planets, as it has been estimated that in the much more expansive Solar System hundreds of millions of rocks could have already been ejected from the spall zones of martian impacts and made their way to Earth (Mileikowsky et al., 2000). Such a healthy estimate has provided motivation for a number of investigators to conduct simulations which attempt to investigate the survivability of an impact large enough to eject material from Mars. One study, by Stöffler et al. (2007), applied pressures of 5–50 GPa to micron-thin layers of different microorganisms. This pressure range, applied via high explosives, is thought to be typical of martian impact ejection (Artemieva and Ivanov, 2004). Spores of Bacillus subtilis exhibited survival fractions (N/N 0 where N is the number of surviving cells and N 0 is the original number of viable cells) of 10−4 under a pressure of 42 GPa.
Bacterial spores are resilient casings that contain identical genetic information to their corresponding microorganism. Thought to be due to their low water content, the cores of bacterial spores have been found to exhibit notably low enzyme activity. This property is believed to contribute to the resilience of the spores, alongside the fact that the bacterial DNA is mixed with acid-soluble proteins, which aid in enzymatic reactivation (Setlow, 1995). Besides the application of high explosives, the planetary ejection stage has also been simulated by firing projectiles at layers of spores, with similar survival rates to that outlined above (Burchell et al., 2004; Fajardo-Cavazos et al., 2009).
5.2. Journey through interplanetary space
Following successful ejection from the impacted planet, the life must then endure the trip to the target planet. Although not as violent as impact-driven ejection, this stage of interplanetary panspermia has been shown to be equally as deadly for a number of microorganisms, owing to the deleterious conditions present in the space environment. The space between planets is vast and empty; vacuum pressures can drop as low as 10−14 Pa, inducing severe desiccation. Exposure to both stellar and galactic cosmic radiation can also be highly damaging, with stellar UV posing the biggest threat. Temperature extremes during interplanetary transit have the potential to rival those of the ejection phase, depending on the orientation of the fragment's orbit around its host star. Together, these form a lethal concoction for many microorganisms. Some, however, have exhibited impressive resilience against the harsh conditions of space.
The seven planets of TRAPPIST-1 orbit an ultra-cool M dwarf star. Due to the star's relatively low temperature, its habitable zone lies very close in (at hundredths of an astronomical unit). As such, it is likely to play host to a radiation environment that is much more damaging than the one surrounding the Earth. Investigations into the X-ray/EUV irradiation of the planets were undertaken by Wheatley et al. (2017). They deduced that TRAPPIST-1 has an X-ray luminosity similar to that of the quiet Sun; it has been hypothesized that such a high flux at these wavelengths could have stripped the planets of their atmospheres (Dong et al., 2017; Roettenbacher and Kane, 2017), raising severe doubts regarding their habitability. The planetary atmospheres are also expected to alter frequently due to persistent flaring events (Vida et al., 2017), which can penetrate magnetospheres (Garraffo et al., 2017) and also affect the surface-based biospheres (Carone et al., 2018) in conjunction with geochemistry (Barr et al., 2017). It would appear, therefore, that the TRAPPIST-1 planets are unlikely to support atmospheres that would enable the harboring of life.
For more Solar System-like exoplanetary systems, we can consider the numerous exposure missions that have taken place in low Earth orbit (LEO), namely at altitudes less than 2000 km. It is important to note that the results of LEO experiments provide a mere estimate of the survivability of panspermia; such orbits are relatively close in proximity to the Earth and therefore fail to accurately mirror the conditions expected during an interplanetary transit. For instance, the minimum vacuum pressure in LEO is ∼10−6–10−7 Pa, several orders of magnitude higher than would be experienced for the majority of a planet–planet trip. Furthermore, the magnetic field of the Earth would shield the lifeform from much of the harmful cosmic radiation that would be plentiful in other regions of the Solar System. Nevertheless, LEO missions have contributed vastly to our understanding of the survival limits of copious microorganisms, placing constraints on the plausibility of panspermia as a concept.
5.2.1. Long duration exposure facility
Still holding the record for the longest exposure to LEO, NASA's Long Duration Exposure Facility subjected spores of B. subtilis to a combination of the space vacuum, solar UV and multiple components of galactic cosmic radiation (Horneck et al., 1994). In accordance with several other astrobiological experiments, solar UV was found to cause most damage due to its tendency to target and break DNA strands within the spores. Prospects for survival were greatly improved with adequate shielding from the UV in place; around 70% of B. subtilis spores were able to survive 6 years of exposure to the LEO vacuum when mixed with the sugar glucose.
5.2.2. EURECA
Similar conclusions were drawn from the results of the EURECA mission, which reported a complete loss of viability of Deinococcus radiodurans cells following 9 months of exposure to LEO (Dose et al., 1995). Up to 12 DNA double strand breaks were observed per chromosome in samples exposed to the solar UV, although shielded cells also showed complete inactivation. These findings were surprising, as D. radiodurans is known to be incredibly resistant to desiccation and radiation; the bacterium can withstand radiation doses of 5000 Gy (around 1000 times a typically lethal dose for humans) with no loss of viability (Moseley and Mattingly, 1971).
A more recent study has found that D. radiodurans cells can survive many more dessication-induced DNA double strand breaks than those observed in the EURECA mission, due to the fact that the genome is able to reassemble before each successive cycle of cell division (Cox and Battista, 2005). As such, the survival rate observed following exposure to LEO conditions could have been higher than that inferred from the experimental results. Regardless, it is clear that the combination of stellar UV radiation and space vacuum form a deadly cocktail, survivable only for the most resilient of lifeforms known to inhabit our Earth. Indeed, vacuum-induced dehydration has been found to alter DNA photochemistry in such a way as to enhance the UV sensitivity of B. subtilis spores 10-fold in comparison to irradiation at atmospheric pressure (Horneck, 1998; Nicholson, 2000).
5.2.3. Biopan
Owing to these early findings, a general consensus has emerged that adequate shielding from the harmful environment of interplanetary space must be in place for microorganisms, such as bacterial spores, lichens and tardigrades, to stand a chance of surviving panspermia.
5.2.3.1. Shielding
Mileikowsky et al. (2000) provide a thorough investigation of shielding for the case of Earth and Mars, and Cockell and Knowland (1999) provide a thorough summary of shielding mechanisms from UV radiation. The effects of shielding were explored as part of the series of experiments that took place using the Biopan facilities aboard various Foton satellites (Horneck et al., 2001). A survival fraction of 10–6 was obtained when B. subtilis spores were exposed to the full LEO environment, while much higher fractions of 0.5–0.97 were determined for shielded samples. Clay shielding was found to be ineffective when placed in the form of a “shadowing” layer; much more protection was received if the spores were mixed in with the clay, or ground meteorite powder. Importantly, the samples consisted of multilayers of spores; the outer layers would have encountered the full extent of LEO conditions, inactivating quickly and potentially forming a protective “crust,” offering added protection to the innermost layers of spores. It is also thought that endolithic microorganisms, residing in microcracks present within rocks, likely exist in the form of biofilms embedded within a complex matrix of sugar molecules (Costerton et al., 1987). This configuration would provide additional protection against the space vacuum. As such, lifeforms mixed within a layer of rock or clay are likely to receive much greater shielding from both the stellar UV and the space vacuum.
5.2.3.2. Lichens
During the Biopan 5 mission, thalli of the lichens Xanthoria elegans and Rhizocarpon geographicum were exposed to the space vacuum and selected wavebands of the solar UV for 14.6 days (Sancho et al., 2007). A lichen comprises a stable symbiotic interaction between fungi and/or cyanobacteria. Lichens can be endolithic, growing between the grains inside rock, and are commonly found in mountainous regions. They have been found to survive complete water loss throughout periods of severe desiccation (Kranner et al., 2008) and withstand higher than average levels of UV radiation. Following the exposure to LEO, 83% of X. elegans cells were found to have intact membranes, while a similarly high survival rate of 71% was determined for R. geographicum.
Furthermore, full photosynthetic recovery was observed, even for samples exposed to over 99% of the solar light. The lichens contain certain pigments that provide screening from the UV, heightening protection during exposure, such as parietin phenolic acids (Solhaug et al., 2003). Similar UV-screening properties were exhibited by cells of the halophilic cyanobacterium Synechoccus following 2 weeks of exposure to LEO as part of the Biopan 1 series of experiments (Mancinelli et al., 1998). Interestingly, X. elegans has also been tested in simulations of the planetary ejection stage of panspermia. The lichen fared similarly to B. subtilis spores, with survival rates dropping by only four orders of magnitude upon the application of 50 GPa pressure (Horneck et al., 2008).
5.2.3.3. Tardigrades
Biopan 6, on the other hand, provided the first testing ground for tardigrades in space (Jönsson et al., 2008). Tardigrades have been identified as one of the most resilient animals on Earth, so are a natural choice for testing in LEO. They have been found to survive extreme temperatures and pressures for significant periods of time (Hengherr et al., 2009; Horikawa et al., 2009), and show incredible resistance to radiation, surviving doses of up to 5000 Gy (Hashimoto et al., 2016). A recent study by Sloan et al. (2017) deduced that tardigrades are likely to survive any mass extinction event with an astrophysical cause, such as a nearby supernova, gamma-ray burst or large asteroid impact.
In a similar way to bacterial spores, tardigrades can undergo a process known as cryptobiosis, whereby metabolic processes shut down in a reversible fashion during times of extreme stress. One particular form of cryptobiosis, known as anhydrobiosis, is of particular relevance to our discussion of survival in space. In this process, a tardigrade will contract and lose the vast majority of its water content, enabling cell stabilizers like trehalose to be formed and metabolism to, in the most extreme cases, be temporarily halted altogether (Welnicz et al., 2011). Samples of the tardigrade species Milnesium tardigradum and Richtersius coronifer survived exposure to the LEO vacuum very well. Combined exposure to both the space vacuum and solar/galactic radiation resulted in reduced, yet still finite, survival for both species tested. Tardigrades therefore have joined bacterial spores and lichens in the list of lifeforms that have survived exposure to the full LEO environment.
5.2.4. EXPOSE
The most recent results obtained from exposure missions in LEO are those of the European Space Agency's EXPOSE facilities, mounted aboard various modules of the International Space Station. Conducted upon EXPOSE-E, the LIFE experiment subjected a variety of eukaryotic organisms to long-term exposure (1.5 years) for the first time (Onofri et al., 2012). Most notably, X. elegans once again achieved full photosynthetic recovery, provided the samples were shielded from UV irradiation. The AMINO experiment, which took place aboard the EXPOSE-R facility, exposed organic molecules to LEO both in their natural state and embedded in meteorite powder (Bertrand et al., 2015). Chosen for the key roles they play in the formation of macromolecules considered essential for life, the amino acids glycine, alanine and aspartic acid showed minimal deterioration following exposure, with 72% of glycine remaining unaffected in unshielded form. Samples of the prokaryote Halorubrum chaoviator, a halophilic archaeon, were exposed to LEO as part of the OSMO experiment (Mancinelli, 2015). If shielded from the solar UV, the archaea exhibited 90% survival rates.
5.3. Atmospheric entry
From the many exposure experiments that have taken place in LEO, it is clear that the deleterious conditions in space can have a devastating effect on many microorganisms. However, it is also clear that a number of lifeforms possess the necessary resilience to survive in such hazardous environments, especially when adequate shielding is in place. We now turn our attention to the final stage of material transfer: atmospheric entry upon reaching the target planet.
Because entry speeds range from 12 to 20 km/s for typical asteroids, the overall process can occur in the space of a few tens of seconds (Nicholson, 2009). Frictional heating over this rapid timescale leads to the formation of a fusion crust on the surface of the meteorite. This crust ensures that the heating fails to penetrate further than the first few millimetres of material, allowing the interior to maintain a relatively constant temperature throughout. Provided the target planet possesses an atmosphere, the eventual impact with the surface will occur at terminal velocity (50 m/s for Earth), a far tamer value than what is involved in the planetary ejection phase.
Thus far, the best attempts to assess the ability of microorganisms to survive meteoric entry have been those of the STONE missions, conducted upon the recovery module heat shields of the same Foton satellites used to host the Biopan 5 and 6 facilities (Parnell et al., 2008; Foucher et al., 2010). The entry speed was measured to be 7.7 km/s, falling short of the expected speeds for asteroids provided above. Nevertheless, none of the microorganisms tested showed any signs of viability following retrieval, most notably B. subtilis. For one of the samples, the fusion crust was found to be around 5 cm deep, possibly due to cracks in the surface of the shield. It would seem, therefore, that further experimentation is required to make any sort of conclusion regarding the survivability of the entry stage of panspermia.
6. Conclusions
The strong prospects for future discoveries of habitable multiplanet systems prompted us to analyze several aspects of panspermia and derive new results. Here, we have applied an impulse formalism from Jackson et al. (2014) to generate orbital constraints on life-bearing ejecta travelling between planets in the coplanar circular case (Eqs. 1 and 3). Resulting analytic probability distributions depend only on the semimajor axes of the source and target planets (Eqs. A18–A24) and can be readily applied to compact multiplanet systems. We have also repackaged and consolidated physical relations that are associated with ejecta to fit within one framework (minimum radius and mass to escape atmosphere: Equations 12 and 13; largest impact fragment: Equations 14 and 16; minimum impactor size to liberate material: Section 4.3; speed and impactor radius to destroy source: Eq. 21). We finally included biological constraints from impact, interplanetary travel and atmospheric entry (Section 5). We hope that our results will represent useful tools to analyze future discoveries of compact multiplanet habitable systems.
Footnotes
Acknowledgments
We thank both referees for particuarly helpful and specific comments on the manuscript, resulting in an improved document. D.V. gratefully acknowledges the support of the STFC via an Ernest Rutherford Fellowship (grant ST/P003850/1) and has received funding from the European Research Council under the European Union's Seventh Framework Programme (FP/2007-2013)/ERC Grant Agreement no. 320964 (WDTracer). D.J.A. is supported by STFC through consolidated grant ST/P000495/1, and JAB is supported through STFC grant ST/R505195/1. A.P.J. acknowledges support from NASA grant NNX16AI31G.
Author Disclosure Statement
No competing financial interests exist.