
Abstract
This work demonstrates the tolerance of lichen Pleurosticta acetabulum under extreme conditions similar to those encountered in extraterrestrial environments. Specifically, the impact of three extreme Mars-like conditions—complete dehydration, extremely low temperature (-196°C/77K), and oxygen depletion—on lichens was investigated. The symbiosis of mycobiont and photobiont partners creates a micro-ecosystem that ensures viability of both symbiotic partners under prolonged desiccation and extremely low temperatures without any cultivation care. Changes in the molecular structure and function of the photosynthetic apparatus, in the level of chlorophylls, polyamines, fatty acids, carbohydrates, ergosterol, efflux of K+, and DNA methylation ensure the ecological integrity of the system and offer resistance of lichens to above-mentioned extreme environmental conditions. For the first time, we also demonstrate that the unprecedented polyextremophilic characteristic of lichens could be linked to biotechnological applications, following exposure to these extreme conditions, such that their ability to produce a high yield of hydrogen was unchanged. All these support that lichens are (a) ideal model systems for a space mission to inhabit other planets, supporting also the aspect that the panspermia theory could be extended to incorporate in the traveling entities not only single organisms but micro-ecosystems like lichens, and (b) ideal model systems for astrobiotechnological applications (hydrogen production), such as in the development of bioregeneration systems for extraterrestrial environments.
1. Introduction
A
Moreover, it has been demonstrated that lichen symbiosis gives additional advantages to both organisms for biotechnological applications. Lichen Pleurosticta acetabulum and a series of other lichen species were able to produce a high amount of molecular hydrogen (H2) under a specific set of conditions (Papazi et al., 2015). Molecular hydrogen is considered one of the most acceptable fuels in the future (Antal et al., 2011). In a closed system, the mycobiont partner consumes oxygen, and the photobiont partner produces hydrogen through photosynthesis. Oxygen consumption enables the activation of the oxygen-sensitive chloroplastic enzyme hydrogenase of the photobiont partner, in order to produce H2 mainly through a dark fermentation pathway (Papazi et al., 2015).
Here, we expand the approach beyond symbiosis by treating lichen as a micro-ecosystem. The ecosystem approach addresses the interactions among participants and their environment as an integrated system through the study of the factors that regulate the pools and fluxes of materials and energy through separate systems (Chapin et al., 2011). The aim of this work is to demonstrate the potential of lichen Pleurosticta acetabulum to survive in extreme environmental conditions, like those on Mars. Specifically, the impact of three extreme Mars-like conditions—absolute water deficit (desiccation), extremely low temperature (-196°C/77K), and oxygen depletion—on lichens is investigated. More attention is paid to the structural maintenance of participants in their dry and regenerated state, while the ecosystem approach is attempted by studying multiple variables for the functional integrity of the separate systems at a higher level of biological complexity. Stress-related changes in their physiology and DNA methylation patterns are evaluated. Moreover, the ability of lichens to produce H2 after their exposure to these extreme conditions is tested. This combined study aims to assess whether functions of lichens as micro-ecosystem ensure their ability to survive exposure to extreme conditions, maintaining in parallel their biotechnological performance. If so, (a) lichens are ideal model systems for a space mission to inhabit other planets, and the theory of panspermia first proposed by Richter (1865) and Arrhenius (1903) could be extended to incorporate in the traveling entities not only single organisms but micro-ecosystems; and (b) lichens are ideal model systems for astrobiotechnological applications, such as the development of bioregeneration systems for extraterrestrial environments.
2. Materials and Methods
2.1. Lichen sampling
Pleurosticta acetabulum thalli were collected from Acer sempervirens and Quercus coccifera substrates of Mountain Idi, Crete, Greece (HTRS07/TM07 X = 578576.83 m, Y = 1900901.19 m). All GPS coordinates follow the Hellenic Terrestrial Reference System 2007 (HTRS07/TM07). Permission to collect the lichen samples was granted by the Hellenic Ministry of Environment Energy and Climate Change, Special Secretariat for Forests (No 108436/956) in accordance with the national law (N3937/2011) on biodiversity conservation. All collections derived from public lands and areas where specific permissions were not required (Article 17), since the wildlife of these areas is not protected by local/regional regulations or national law. Moreover, the field studies did not involve endemic, endangered, or protected species.
Lichen thalli were maintained in a dry state in the lab. According to the regeneration protocol (Papazi et al., 2015), lichen thalli are washed with deionized water for 10 min, followed by wrapping in a wet towel. The wrapped lichens are incubated at room temperature (20–22°C) for 1 h before they are transferred to a cold room (4–6°C) for an additional 16 h, while the quality of regeneration is tested before any other treatment is applied with fluorescence induction measurements of the photosynthetic efficiency (Pirintsos et al., 2011). This is also considered the control state (LR treatment) for all subsequent treatments (see below).
2.2. Tissue treatments
Dehydration stress was achieved by leaving lichen thalli to air-dry at room temperature (20–22°C) until tissue weight could not be reduced further (L treatment). Exposure to extremely low temperature was achieved by incubating dry lichen tissue in liquid nitrogen (-196°C/77K) for 30 min. Then, lichen thalli were returned to room temperature (20–22°C) for 1 h and afterward regenerated (LNR treatment). Additionally to this treatment, regenerated lichen tissue was also exposed in liquid nitrogen (-196°C/77K) for 30 min, and afterward lichen thalli were returned to room temperature (20–22°C) for 1 h (LRN treatment). The LNR_LNR treatment resulted from LNR treatment after the regeneration step: after leaving the thalli to air-dry, exposure to liquid nitrogen followed by regeneration was repeated. The abbreviations of the treatments are presented in Table 1. Following all these tissue treatments (L, LR, LNR, LNR_LNR, LRN), samples were collected for the determination of physiological, biochemical, and molecular variables described below.
2.3. Fluorescence induction measurements
The Handy Plant Efficiency Analyser (Handy PEA, Hansatech Instruments, King's Lynn, Norfolk, UK) was used to estimate the molecular structure and function of photosynthetic apparatus. The JIP method, described by Strasser and Strasser (1995), was used to estimate the maximum yield of photochemistry (Fv/Fm). This method is based on the measurement of a fast fluorescence transient with a 10 μs resolution in a time span of 40 μs to 1 s. Fluorescence was measured at 12-bit resolution and excited by three light-emitting diodes providing a saturated light intensity of 3000 μmol m−2 s−1 of red (650 nm) light. Lichen samples were maintained in the dark for 5 min before the measurements.
2.4. Extraction and quantification of chlorophylls
Chlorophylls were extracted with hot methanol from lyophilized lichen tissue and photometrically estimated according to the method and the equations of Holden (1965).
2.5. HPLC analysis of ergosterol
Ergosterol was extracted from lyophilized lichen tissue according to the method of Dahlman et al. (2002). A high-performance liquid chromatography (HPLC) system was used for the quantification of ergosterol levels. Analysis was performed with an LC-10AD, VP Shimadzu HPLC apparatus equipped with a diode array detector (SPD-M10A VP, Shimadzu) and a narrow-bore column (C-18, 2.1 mm × 200 mm, 5 μm Hypersil, Hewlett Packard). A reference curve was designed with known ergosterol amount in order to estimate unknown ergosterol level in the samples.
2.6. Polarographic measurements of maximum photosynthetic and respiratory rates
A Clark-type electrode system (Hansatech Instruments, King's Lynn, Norfolk, UK) was used to determine the maximal photosynthetic and respiratory rates of lichen thalli. The actinic light (1500 μmol m−2 s−1) was generated with two lamps (ENX360 W/82 V). The infrared part of the applied irradiation was filtered off by inserting a 2% CuSO4 containing cuvette (4 cm path length) into the light beam.
2.7. Potassium efflux estimation
A flame photometer (Sherwood M410) was used for the estimation of potassium efflux from lichen samples, as described by Puckett et al. (1977). Lichen treatments were immersed in 50 mL dH2O for 1 h. A sample of LR lichen after exposure to microwave radiation (600 W) three times for 10 s each time was used as positive control. A sample of dH2O was used for the estimation of potassium efflux in the flame photometer. A reference curve was designed with known potassium amount in order to estimate potassium level in the dH2O samples.
2.8. HPLC analysis of polyamines
Total polyamines were extracted from lyophilized lichen tissue according to the method of Kotzabasis et al. (1993). A HPLC system was used for the quantitative and qualitative estimation of polyamines. The analysis was performed with an LC-10AD, VP Shimadzu HPLC apparatus equipped with a diode array detector (SPD-M10A VP, Shimadzu) and a narrow-bore column (C-18, 2.1 mm × 200 mm, 5 μm Hypersil, Hewlett Packard). The equations described by Kotzabasis et al. (1993) were used to estimate the amount of each polyamine in the lichen samples.
2.9. GC-FID analysis of fatty acids
For fatty acid analysis, lipids were extracted from lyophilized and homogenized dry material of lichen according to Bligh and Dyer's (1959) method and were further prepared for fatty acid analysis according to the procedure of Kates (1972). Fatty acid methyl esters were analyzed by a gas chromatograph (Hewlett Packard, 5890-Series II) equipped with a flame ionization detector (GC-FID). A capillary column (SGE-BPX 70; 50 m length, 0.22 mm diameter, 0.25 μm film thickness) and a split/splitless capillary inlet (19251A) were used. Helium was used as carrier and nitrogen as auxiliary gas. The operating conditions were 320°C injection port, 300°C flame ionization detector, and column temperature was programmed to change between 155°C at the beginning and 220°C at the final step. For peak identification, solutions of reference substances (Supelco 18919-1amp FAME mix C4-C24 and FAMEs purchased from Sigma) were analyzed under the same conditions, and their retention times and chromatograms were compared to those of samples. The contribution of each identified compound was expressed as the percentage of its peak area to the total area of all peaks eluted in each chromatogram.
2.10. Extraction and quantification of carbohydrates
For carbohydrate extraction, 50 mg of whole lichen thalli were submerged in 2 mL dH2O and were exposed to microwave radiation (600 W) three times for 10 s each time, with intermediate time breaks on ice in order to separate soluble from insoluble intracellular carbohydrates. The quantification of carbohydrates was accomplished by the Molisch test (Cammack et al., 2006). Carbohydrates are quantified by measuring the absorption in 565 nm, based on a reference curve which was designed with known concentrations of glucose.
2.11. GC-TCD measurements of H2 and O2
Hydrogen and oxygen measurements were made with a gas chromatograph, using a thermal conductivity detector (GC-TCD) (Shimadzu GC 2010 Plus, Kyoto, Japan). To separate H2 and O2, argon was used as the carrier gas under pressure of 5 bar and at oven temperature of 120°C. The column used was a capillary Vici Metronics MC (Poulsbo, USA) with length 30 m (diameter: 0.53 mm) and film thickness 20 μm. The temperature of the TCD was set at 200°C for the detector and 180°C for the injector. The quantification of hydrogen and oxygen was accomplished by injecting known quantities in the GC-TCD and designing a reference curve.
2.12. Methylation-sensitive amplification polymorphism (MSAP) analysis
For each sample, MSAP analyses were performed by using both EcoRI/HpaII and EcoRI/MspI digests. Digestion of 200 ng aliquots of genomic DNA with either of the two isoschizomers (HpaII, MspI) was carried out in 20 μL containing 1X for all Buffer, 4 U EcoRI (New England Biolabs, Ipswich, MA, USA) and 3 U of either HpaII or MspI enzyme (New England Biolabs, Ipswich, MA, USA) for 3 h at 37°C. Two different adapters, one for the EcoRI sticky end and one for the HpaII/MspI sticky end, were designed to avoid reconstruction of restriction sites. The adapters were ligated to DNA after digestion by adding to each final digestion 5 μL of a mix containing 5 pmol of EcoRI adapter, 50 pmol of HpaII/MspI adapter, 1 mM ATP, 1X of all buffers, and 1 U of T4 DNA ligase (Invitrogen, CA, USA). The ligation was incubated for 3 h at 25°C. The EcoRI adapter consisted of the combination of two primers. EcoRI and HpaII/MspI adapters are presented in Supplementary Table S1 (see Supplementary Data at
2.13. MSAP data collection and processing
Methylation-sensitive amplification polymorphism product mixtures were denatured in formamide at 94°C for 2 min and electrophoretically fractionated on an ABI Prism 3730xl (Applied Biosystems, Carlsbad, CA, USA). In order to reduce the impact of potential size homoplasy, only fragments ranging from 150 to 500 bp in size were counted. Fragment size was determined with Genemapper v4.0 (Applied Biosystems, Carlsbad, CA, USA) employing an internal standard (GS 500 LIZ, Applied Biosystems), and an initial data matrix was produced with fragment presence scored as “1” and absence as “0.”
Scoring initially entailed determining marker presence/absence between the control treatment of lichens (defined as LR) and any of the treatment states [L, LNR, LNR_LNR, LRN, LR/Lcs (LR + 5 days illuminated in a closed system), LNR/Lcs (LNR + 5 days illuminated in a closed system), LNR_LNR/Lcs (LNR_LNR + 5 days illuminated in a closed system), LR/Dcs (LR + 5 days darkness in a closed system), LNR/Dcs (LNR + 5 days darkness in a closed system), LNR_LNR/Dcs (LNR_LNR + 5 days darkness in a closed system)]. Each state was investigated for marker presence/absence with respect to the two isoschizomer profiles and for each individual sample separately. All samples were taken to originate from a “single identical genotype” representing collectively the different genomes of the two lichen symbionts. All amplified fragments were initially scored; nevertheless, only polymorphic ones (present or absent in more than 5% of each sample X treatment combination) were retained for subsequent analysis. Each state was described by two binary digits (“0” or “1”; duplets) corresponding to the two isoschizomer profiles. Quadruplets (a series of four binary digits) were subsequently produced by pooling together duplets from the reference as well as from the treatment state and were assigned to change events between reference and treatment states. Sixteen alternate quadruplets could thus be produced, each classified as either de novo methylation, de-methylation, no change, or other variation, thus producing four types of methylation change events. All data management steps, including event classification and cluster analysis, were performed employing an in-house script.
2.14. Data analysis
Each experiment was repeated three to five times. Standard deviation and means of values across all experiments were calculated and presented in the diagrams. Differences between the treatments have been tested with analysis of variance (ANOVA), followed by pairwise t-tests, using Holm's adjustment for the p-values. Analysis took place in R, version 3.2.2.
Unweighted Pair Group Method with Arithmetic Mean (UPGMA) analysis was performed by using the online version of DendroUPGMA: A Dendrogram Construction Utility (
3. Results
3.1. Extremophilic behavior of lichen under complete dehydration and extremely low temperature
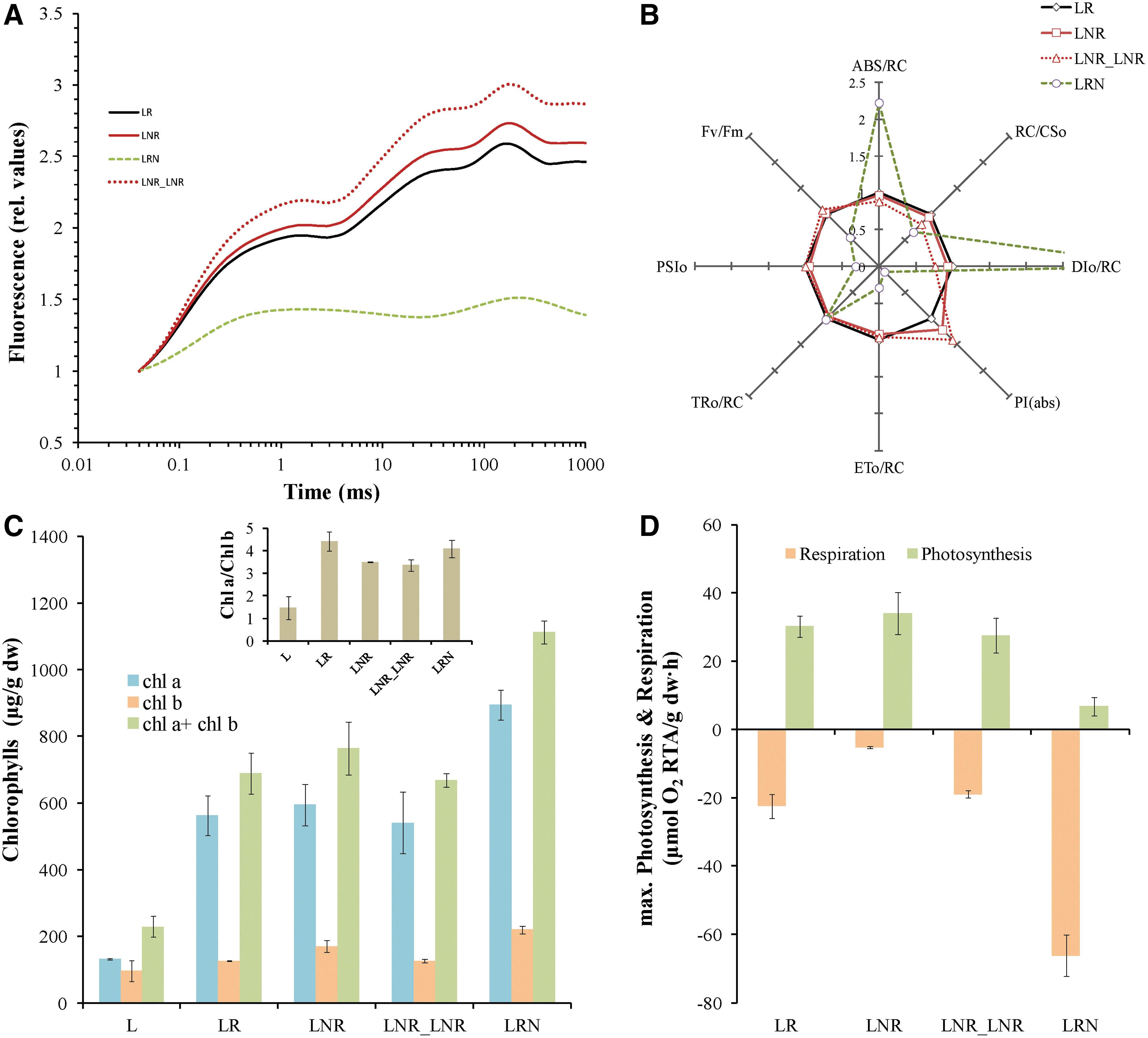
Lichen Pleurosticta acetabulum was studied for its resistance to prolonged drought and extremely low temperatures (-196°C/77K). Both dried and regenerated lichen tissue was incubated in liquid nitrogen (-196°C/77K) for 30 min (LNR and LRN treatments, respectively) and compared with control treatment LR (regenerated lichen). A fourth treatment (LNR_LNR) resulted from LNR treatment after being incubated in liquid nitrogen twice (see Materials and Methods). Fluorescence induction measurements (JIP test) were used to estimate the molecular structure and function of photosynthetic apparatus of the photobiont partner. A sample of dry lichen (L treatment) is excluded from this experiment, as it is metabolically inactive and measurements cannot be made. Fluorescence induction curves (Fig. 1A), as well as a number of parameters reflecting the molecular structure and function of the photosynthetic apparatus, were estimated (Fig. 1B). In LNR and LNR_LNR treatments, all parameters of the JIP test were not differentiated with respect to control (Fig. 1B), indicating that the photobiont partner remains unaffected. The opposite observation is made for the LRN treatment, in which JIP test parameters show that the photosynthetic apparatus is under stress. On the other hand, when regenerated lichen is exposed to -196°C/77K (LRN treatment), the photobiont partner shows clearly a stress response but does not collapse. It is known that stress induces an increase of functional photosynthetic antenna (ABS/RC) and a decrease in the density of active reaction centers (RC/CS). This leads to enhanced non-photochemical energy quenching (DIo/RC) and to decreased photochemical quenching (PSIo) and therefore to a decrease of photosynthetic efficiency expressed as Fv/Fm. In the LRN treatment, the Fv/Fm value decreased to 0.5, whereas the corresponding values in the treatments LR, LNR, and LNR_LNR remain unchanged about 0.7.
(
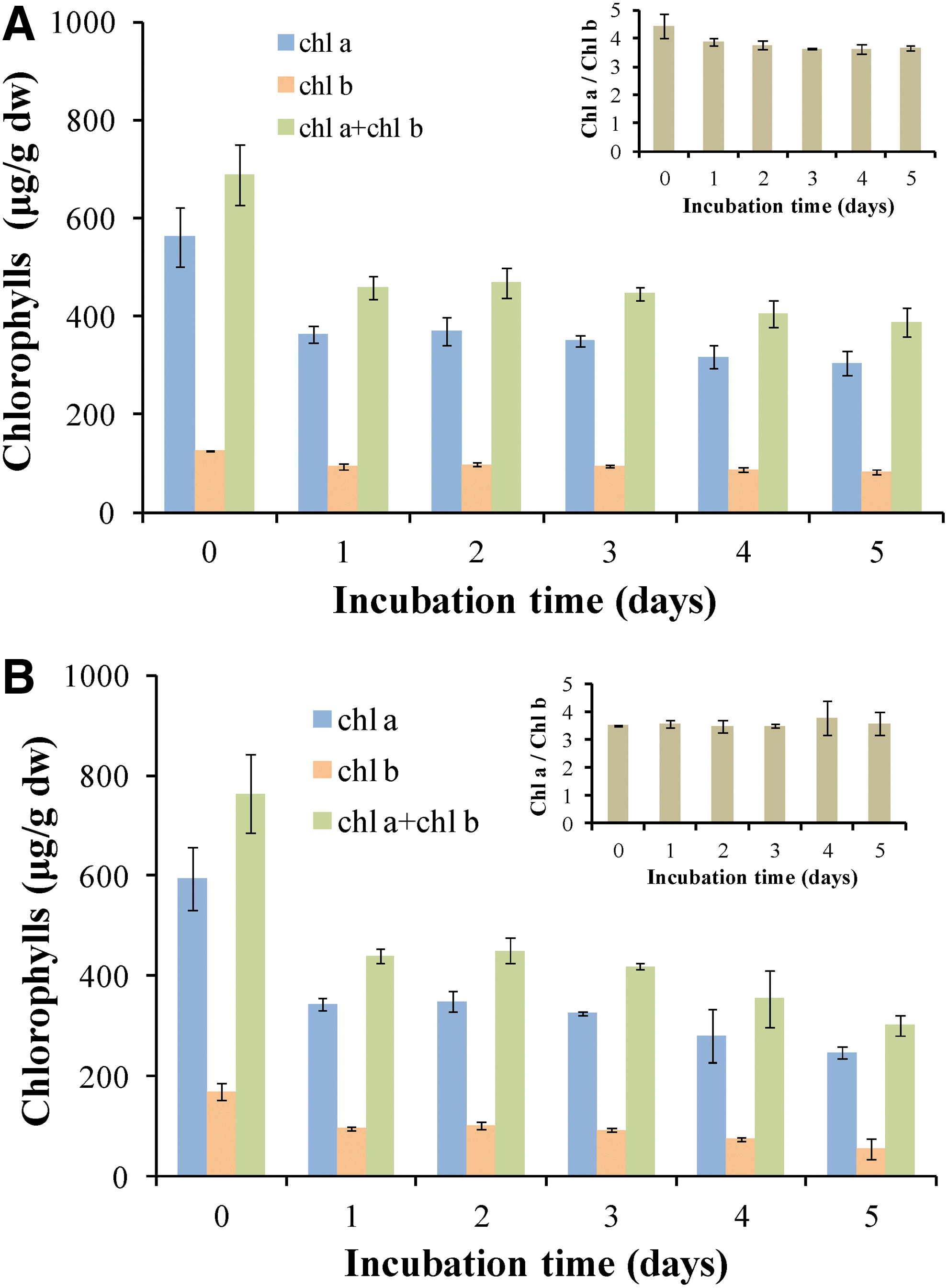
To further confirm these findings, total chlorophylls were extracted and quantified in order to estimate the preservation of photosynthetic apparatus. In this experiment, L treatment is included. In Fig. 1C, the levels of chlorophyll a (Chla) and chlorophyll b (Chlb) and the ratio of chlorophyll a to chlorophyll b (Chla/Chlb) are represented. Total chlorophylls, as well as the ratio Chla/Chlb, increase during the regeneration of lichen tissue and the transition from absolute dehydration status (L treatment) to the LR treatment. LNR and LNR_LNR treatments have the same amount of chlorophylls and the same Chla/Chlb ratio as control (LR treatment), confirming that absolute drought and extremely low temperature stress did not affect the photosynthetic apparatus. In LRN treatment, total chlorophylls are significantly higher than control (Fig. 1C). This could be explained by the increase in antenna size (ABS/RC), as shown in Fig. 1B. This observation demonstrates that the structure and function of photosynthetic apparatus is not completely damaged in LRN treatment and the lichen could be revived following that treatment.
Polarographic measurement of photosynthesis and respiration gives an overall view of the metabolic activity of whole lichen thalli. In this experiment, L treatment is excluded again, as lichen has no metabolic activity in the dry state. The results confirmed that the photosynthetic activity of the photobiont is mostly affected in LRN treatment (Fig. 1D). The rest of the treatments are no different than control treatment. Regarding respiration, LNR treatment has lower and LRN has higher respiratory rate than control. A possible explanation is that LRN treatment is coping with stress and consequently needs more energy to activate defense mechanisms. More energy is obtained by increasing the rate of respiration (Fig. 1D).
The study of the photobiont partner is followed by the study of the mycobiont partner of lichen. This was achieved by the isolation and quantification of ergosterol levels. All four treatments seem to have a similar amount of ergosterol as control (Fig. 2A). The results of this experiment show clearly that the mycobiont partner is also completely tolerant under extreme drought and cold stress in all treatments. The only treatment that exhibited a relatively low amount of ergosterol (about 50% of the control) is the dry state of the lichen tissue (L treatment), but its value reaches the control value immediately after regeneration (Fig. 2A).

(
An additional accurate method to confirm the vitality of lichens in complete dehydrated form and under extremely low temperature is the estimation of potassium efflux. This method is based on the fact that when plasma membranes are disrupted, potassium cations are released in the medium (Puckett et al., 1977). LR treatment after exposure to microwave radiation was used as a positive control (Fig. 2B: LR-MW), as radiation disrupts membranes. LNR_LNR treatment is excluded from this experiment, as in this treatment two separate hydration and dehydration procedures take place that possibly wash the extracellular cations before the end of the treatment and a valid measurement cannot be made. The bar chart in Fig. 2B shows clearly that L and LNR treatments do not differentiate from control (LR). Slightly more potassium cations were measured in LRN treatment (about 7% of the positive control LR-MW), proving that lichen thalli in this treatment are stressed to a degree but are not destroyed.
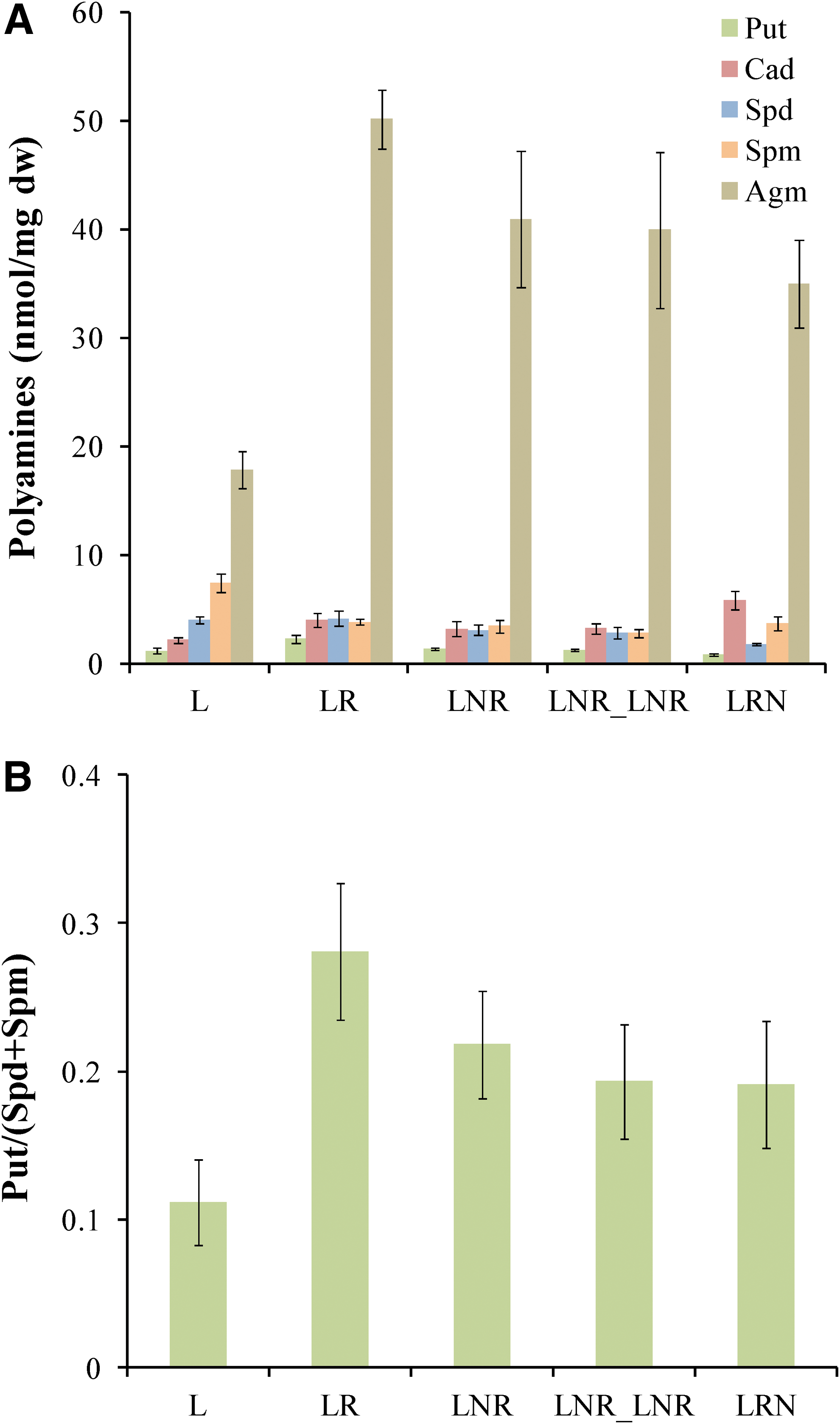
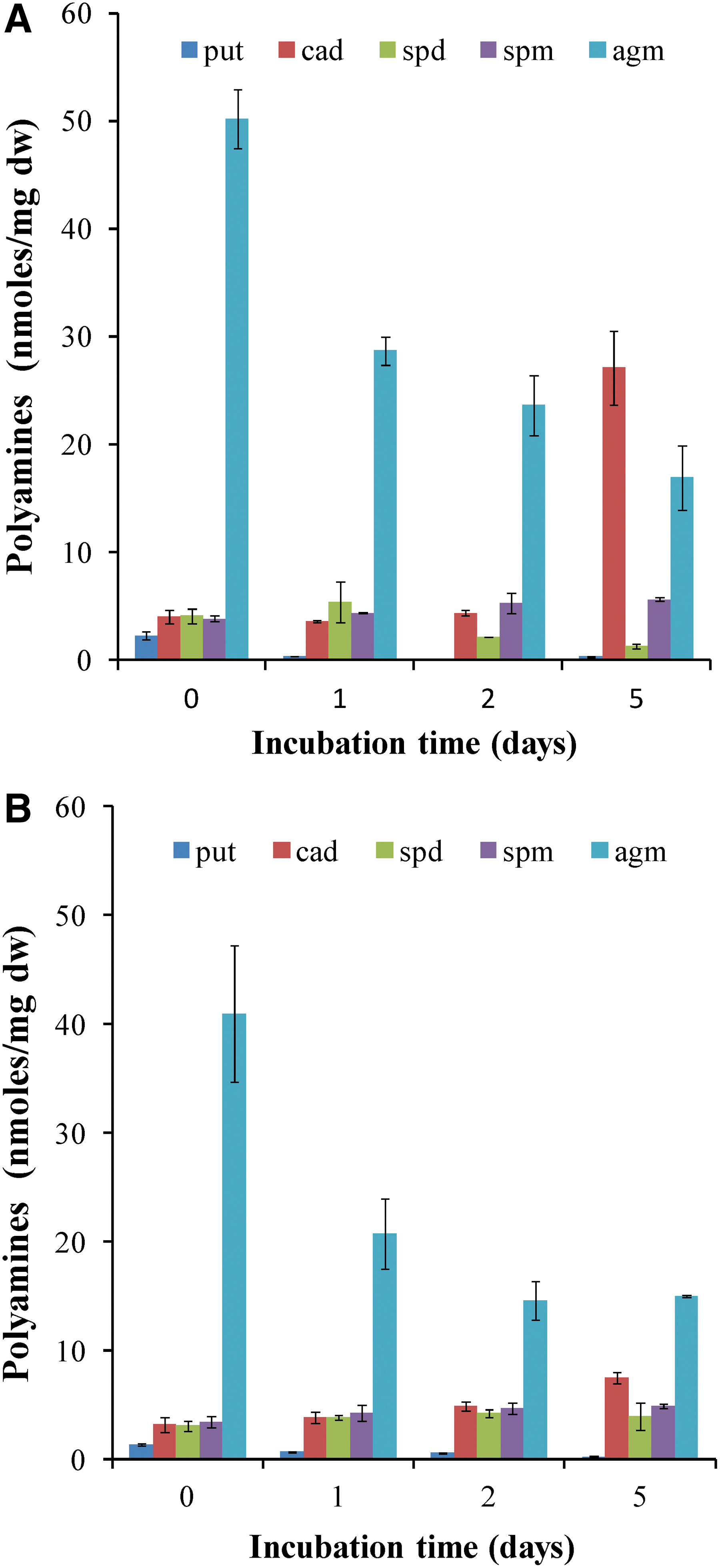
In Fig. 3, it is clearly presented that the cellular amount of all polyamines [agmatine (Agm), cadaverine (Cad), putrescine (Put), spermidine (Spd), and spermine (Spm)], as well as the ratio Put/(Spd+Spm) in the treatments LR, LNR, LNR_LNR, and LRN, remains almost unchanged. It is worth mentioning that only cadaverine (Cad) increases significantly in LRN treatment. This polyamine is correlated with defense mechanisms against stress (Jancewicz et al., 2016). This observation confirms previously presented results (Fig. 1) that LRN is the most stressed of all treatments. The L treatment (lichen in dry state) contains a different polyamine profile with considerably lower Put/(Spd+Spm) ratio compared to the other treatments (Fig. 3B).
(
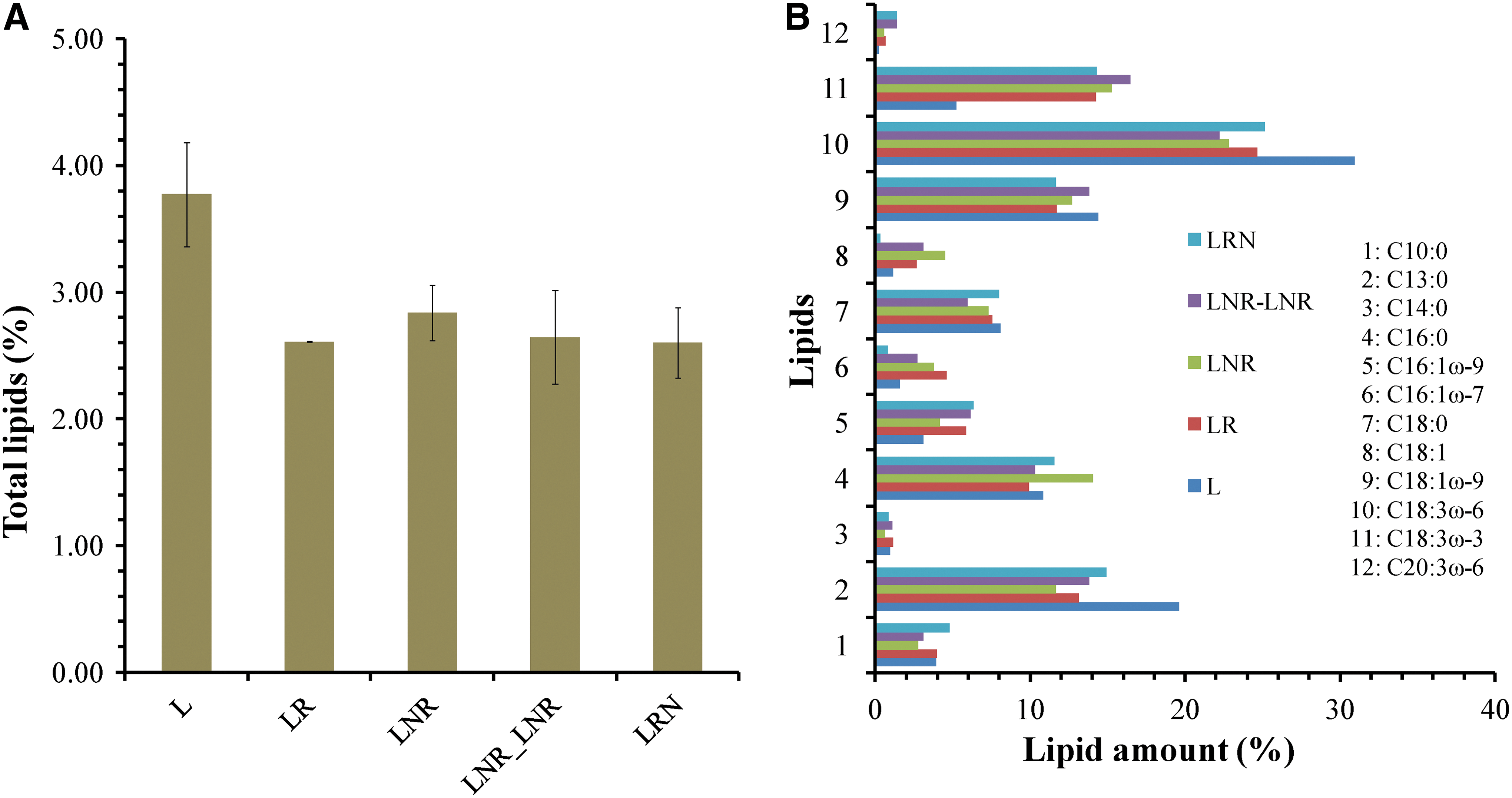
To further investigate the responses of lichens under extremely low temperature and intense water deficit, lipids and intracellular carbohydrates were isolated and quantified. In Fig. 4A, total lipid content is represented. It seems that L treatment has the highest amount of lipids compared to the control (LR), while there is not significant differentiation between the other treatments and the control. It is worth mentioning that 12 fatty acids were identified in all treatments, and they are represented in Fig. 4B. Two of them, tridecanoic acid (C13:0) and 6,9,12-octadecatrienoic acid (C18:3ω-6) are increased in L treatment, resulting in total lipids increase. The same treatment (L) seems to have the most intracellular carbohydrate content of all treatments, as is shown in Fig. 5. The above results demonstrate that changes in lipids and carbohydrates are necessary for lichen in L treatment to adapt to prolonged dehydration. These changes are not observed in the LNR, LNR_LNR, and LRN treatments.
(
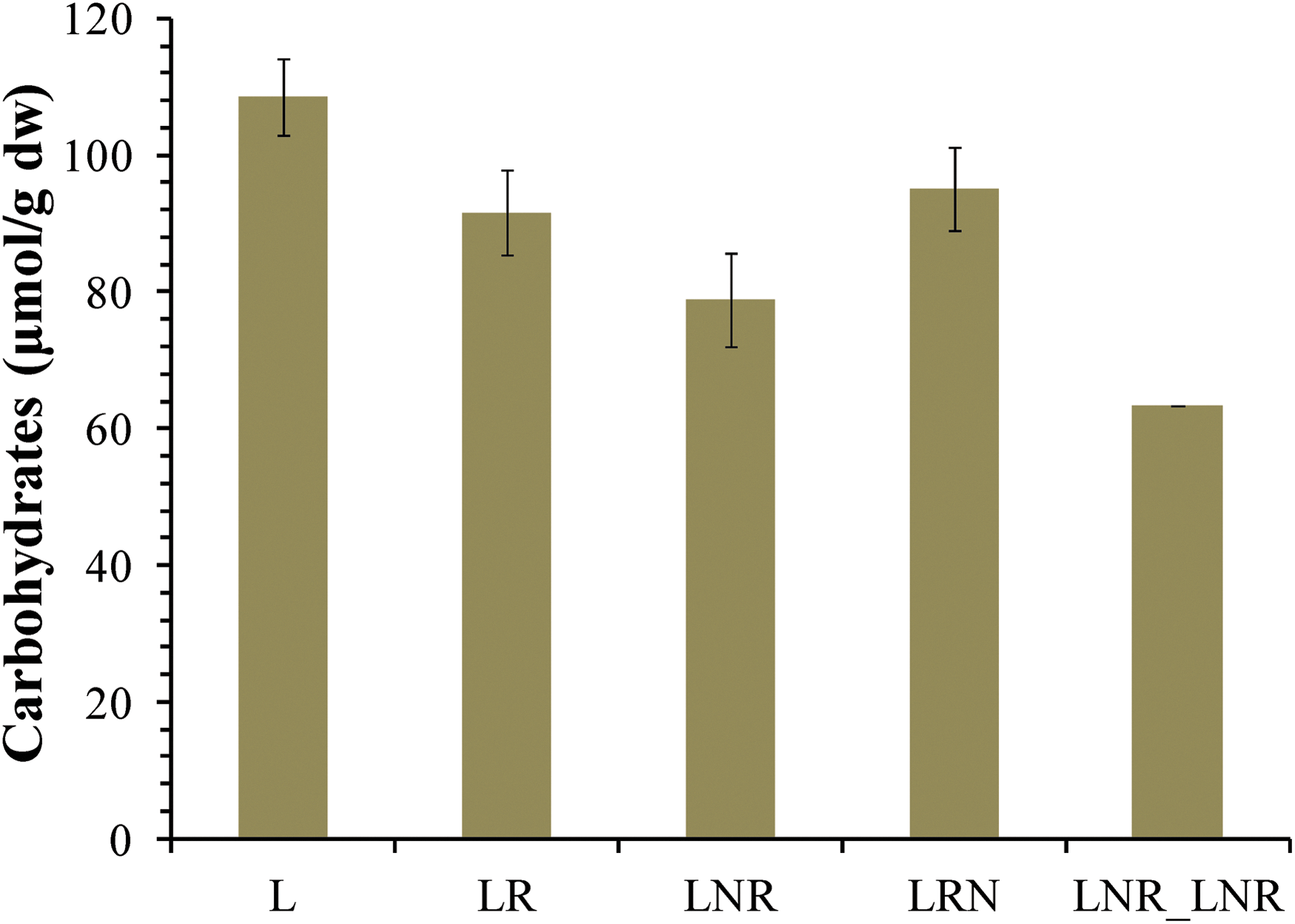
Intracellular carbohydrate content in lichen Pleurosticta acetabulum in the treatments L, LR, LNR, LNR_LNR, LRN.
3.2. Lichen's ability to produce high yield of H2 under anoxic conditions after dehydration and exposure to extremely low temperature
Our previous study proved for the first time that lichens produce a high amount of molecular hydrogen (H2) when incubated in a closed system (Papazi et al., 2015). In this work, lichen Pleurosticta acetabulum was studied for its ability to produce molecular hydrogen (H2) after dehydration and exposure to extremely low temperature (-196°C/77K). LNR and LNR_LNR treatments were tested for their ability to produce H2, as they were exposed to both extreme stress conditions and were the most resistant. These treatments as well as the control treatment LR were incubated in hermetically sealed bottles under light or dark conditions. The kinetics for O2 consumption and H2 production in a time span of 5 days under light conditions are shown in Fig. 6. The H2 production remains unchanged in LNR and LNR_LNR treatments compared to control (LR). In all cases, O2 is completely consumed by day 2, and hypoxic conditions are subsequently established. H2 is then produced until day 5. Similar results were observed when the experiment was repeated under dark conditions (data not shown). These results indicate that intense dehydration and extremely low temperature do not affect H2 production by lichens. Also, it seems that lichens are functional after growing for 5 days under oxygen-depleted conditions.
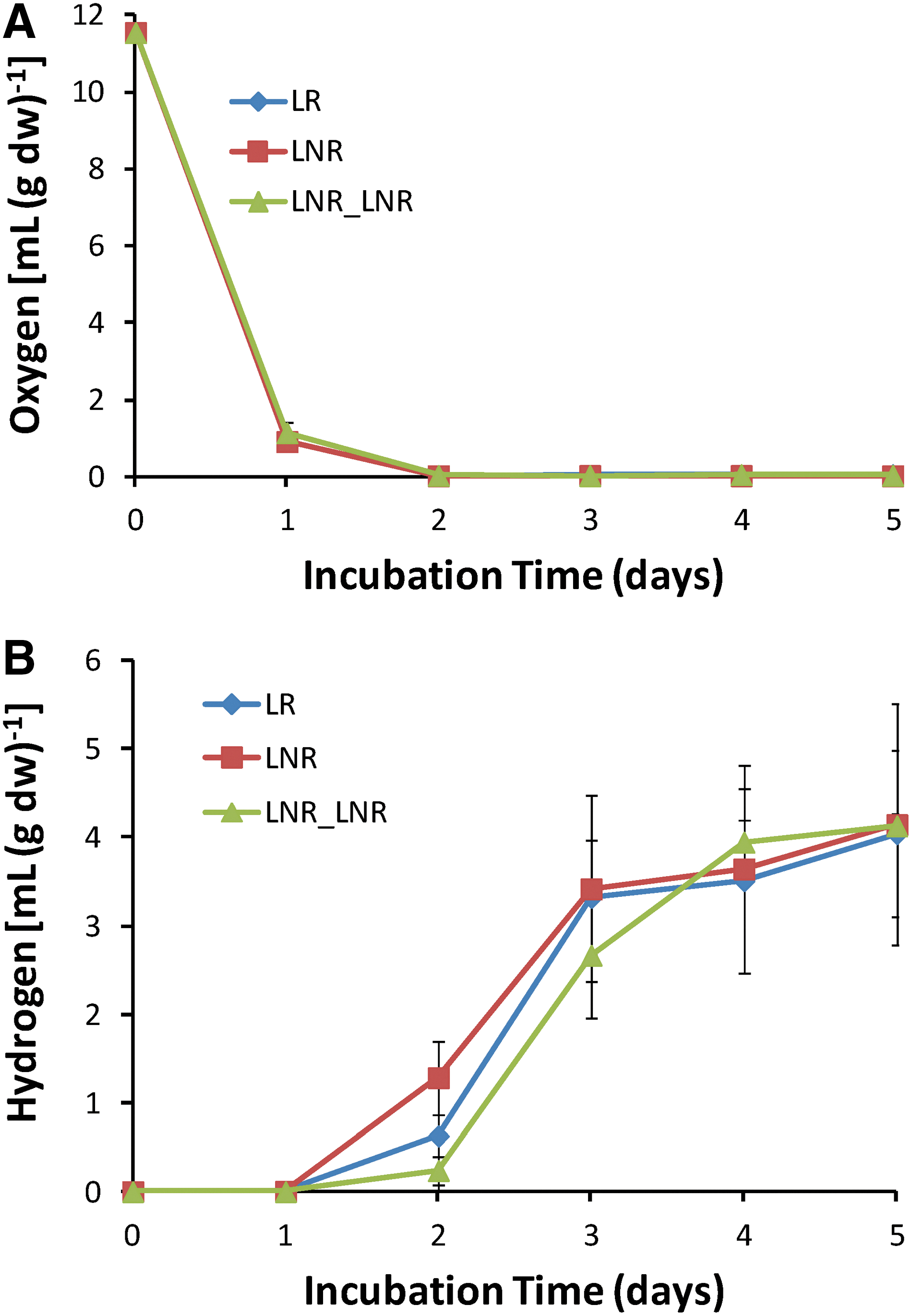
(
To further investigate the resistance of lichen in a third extreme condition, hypoxia, the amount of chlorophyll during the 5-day incubation was measured in order to acquire information about the photobiont partner. In Fig. 7, the amount of chlorophyll a and b decreases in LR and LNR treatment after 1 day of incubation in closed bottles, but it remains stable until day 5 without any change in the Chla/Chlb ratio (Fig. 7). This indicates that oxygen-depleted conditions do not destroy the photosynthetic apparatus and the photobiont partner has the ability to produce hydrogen.
(
The resistance of mycobiont partner in oxygen depletion is estimated by the level of ergosterol. Ergosterol biosynthesis requires oxygen (Toshumi et al., 1978), and this explains the decrease of ergosterol level (about 10%) on the first day of incubation, in all tested treatments (Fig. 8). The fact that on day 5 the amount of ergosterol is the same as on day 1 confirms that the mycobiont partner (in addition to the photobiont partner discussed earlier) is not considerably affected by low oxygen conditions.
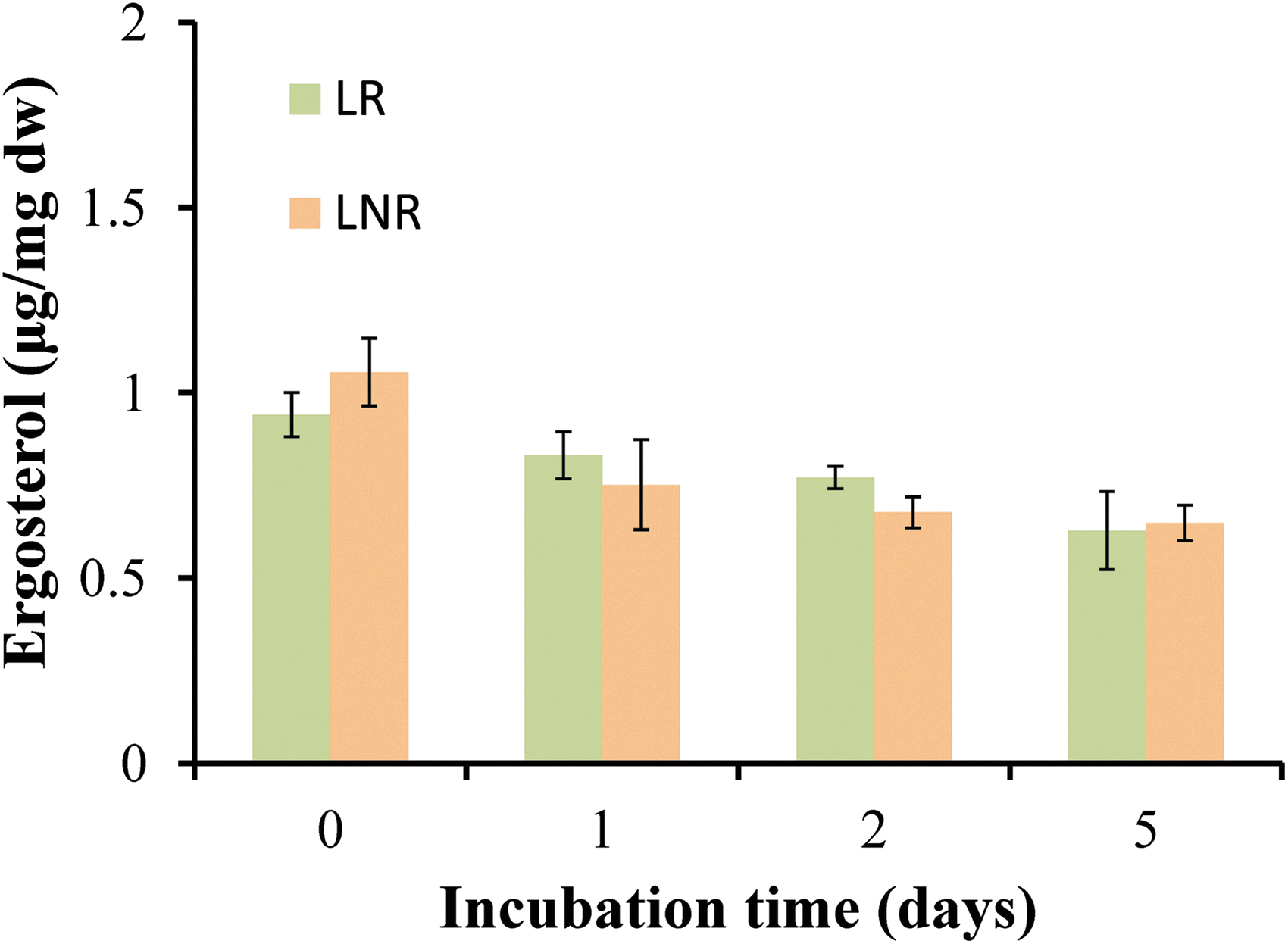
Ergosterol levels in lichen Pleurosticta acetabulum in LR and LNR treatments during 5 days incubation in sealed bottles and light conditions.
In order to estimate the adaptation in the tested condition, total polyamines from LR and LNR treatment during 5 days of incubation in sealed bottles and light conditions were isolated and quantified. In Fig. 9, a gradual decrease in polyamine Agm during the incubation time in both treatments is observed. Additionally, the polyamine Cad is increased on the fifth day of incubation, mostly in LR treatment (Fig. 9).
(
Changes in the type of fatty acids are linked with mechanisms that lichen uses to adapt in stress conditions (MacGillivray and Helleur, 2001). Especially under hypoxic conditions, lipid biosynthesis pathway is crossed with hydrogen production pathway through dark fermentation (Papazi et al., 2015). In Fig. 10, total lipid content is almost on the same level as in LR as well in LNR treatment during the incubation time of 5 days. Comparing the fatty acid composition under hypoxic conditions on day 1 and day 5, there is an increase of C16:0 (hexadecanoid acid) and C16:1ω-9 (9- hexadecenoic acid) with parallel decrease of the amount of C10:0 (decanoic acid) and C13:0 (tridecanoid acid) in LR treatment (Fig. 11). In LNR treatment, respectively, the fatty acids C13:0 (tridecanoid acid), C14:0 (tetradecanoid acid), and C18:1 (E-9-octadecenoic acid) disappeared, whereas the fatty acid C10:0 (decanoic acid) strongly increased (Fig. 11).

Total lipid content in lichen Pleurosticta acetabulum in LR and LNR treatments during 5 days incubation in sealed bottles and light conditions.

The 12 fatty acids identified in lichen Pleurosticta acetabulum in LR and LNR treatments in a closed system in light conditions on day 5 compared with day 0.
3.3. DNA methylation pattern changes
In order to assess the effect of the extreme stress conditions on the DNA methylation pattern, the MSAP molecular marker technique was employed. Given the relatively small size of the genomes of the two symbiotic organisms comprising the lichen, it was decided to employ a small number of additional nucleotides (i.e., those added to the core of the primers; see Materials and Methods above) during the selective amplification step of the MSAP procedure. Specifically, for the HpaII/MspI end, the same primer was employed for both the pre-selective and the selective amplification steps bearing only one additional nucleotide (Supplementary Table S1). Using the same number of differentiating nucleotides, in both the pre-selective and the selective amplification steps, is not found frequently within the AFLP family of protocols. Eventually, this approach was proven appropriate, since it allowed for the production of 567 MSAP markers, in total, from all primer X enzyme combinations (data not shown). This number compares very favorably with published values with other species having larger genomes (Marconi et al., 2013; Sáez-Laguna et al., 2014) and points to the reliability of the MSAP methodology applied to the lichen for this first time. On the other hand, it needs to be emphasized that, under the MSAP technique, the two symbiont genomes cannot be separated.
Consequently, produced marker profiles represent an amalgamation of two separate ones ideally originating from each individual genome. Percentages of methylation change events are shown in tabular format in Fig. 12A, while in Fig. 12B a cluster dendrogram is shown which depicts gradation of epigenetic differentiation among treatments. It can be seen that the least effect on the lichen DNA methylation status is induced by the L treatment, since it is the only one exhibiting 69.09% “no change” events with respect to the control treatment LR. All other treatments [LNR, LNR_LNR, LRN, LR/Lcs (LR + 5 days illuminated in a closed system), LNR/Lcs (LNR + 5 days illuminated in a closed system), LNR_LNR/Lcs (LNR_LNR + 5 days illuminated in a closed system), LR/Dcs (LR + 5 days darkness in a closed system), LNR/Dcs (LNR + 5 days darkness in a closed system), LNR_LNR/Dcs (LNR_LNR + 5 days darkness in a closed system)] indicate much higher changes (Fig. 12A). By examining the UPGMA dendrogram (Fig. 12B), it can be seen that indeed the L treatment is the most distantiated from all the rest since it exhibits the least differentiation from the control, while all the rest exhibit an increased differentiation vis-à-vis the control.

(
4. Discussion
Extremophiles are organisms that thrive in extreme environments (Rothschild and Mancinelli, 2001). The discovery of extreme environments and the organisms that inhabit them has made more plausible the search for life outside Earth and even the transport of life from one planet to another, which is called panspermia (Madigan and Marrs, 1997). A number of studies include lichens in the group of extremophiles, as they are tolerant in extreme environments (Beckett et al., 2008; Kranner et al., 2008; Dyakov et al., 2015). For that reason, they were chosen as model organisms for space missions (Sancho et al., 2007; de Vera et al., 2010; Raggio et al., 2011).
Here, beyond considering lichens only as organisms, we also study lichens as mini-ecosystems, and we link this concept with panspermia. The concept of “lichen as an ecosystem” has its roots in the 1970s, when J.F. Farrar postulated that “lichens can be considered as ecosystems with two trophic levels: algal producers and fungal consumers,” considering also carbon, nitrogen flows, and net productivity issues (Farrar, 1976; Wrzosek et al., 2017). This concept has been neglected for long, but its current acceptance has gradually expanded (Tehler and Wedin, 2008; Wrzosek et al., 2017).
This work explores some resistance aspects of Pleurosticta acetabulum under three extreme Mars-like conditions: absolute dehydration, extremely low temperature (-196°C/77K), and hypoxia. The measured responses, some of which are attributed to each separate component, appear an ecological integrity that contributes to the resistance of lichen. In these conditions, the thallus remains structurally and functionally intact, and it is able to produce high amounts of molecular hydrogen following incubation in a closed system. These results document the resistance of lichen and point to possible biotechnological applications in extreme environments, making lichen the ideal micro-ecosystem for space missions and for astrobiotechnological applications. In addition, the results support that the panspermia theory could be extended, incorporating the clarification that in the traveling entities not only single organisms but micro-ecosystems could participate. The term ecosystem is defined as a community of organisms and their physical environment interacting as an ecological unit (Capra and Luisi, 2014). This aspect is also strengthened by the lithopanspermia concept (Melosh, 1988, 2003), which postulates that life can be transferred from one planet to the other by rock. Rock is part of the physical environment of saxicolous and endolithic lichen micro-ecosystems, which in addition to protection may act as a reservoir of minerals. The Lithopanspermia space experiment, which included for the first time the lichen Aspicilia fruticulosa (Raggio et al., 2011), offers empirical evidence to this argument.
4.1. Lichen's resistance under extreme conditions that simulate space conditions
In the present work, the lichen Pleurosticta acetabulum is presented as a polyextremophilic organism that tolerates extremely low temperature (-196°C/77K) and complete dehydration. Previous studies demonstrated the resistance of the whole lichen thalli under simulated Mars-like conditions (de Vera et al., 2010). Water absence and extremely low temperature are two conditions that characterize the environment on Mars (Hess et al., 1977).
The present contribution invades into the resistance of lichen with respect to both partners under these two extreme conditions. Appropriate experiments were carried out either for the photobiont or the mycobiont or for both of them. Fluorescence induction measurements (JIP test parameters) and chlorophyll levels were used to estimate the molecular structure and function of photosynthetic apparatus of the photobiont partner (Fig. 1A, 1B). The Fv/Fm values that express the photosynthetic efficiency show that exposure of the lichen to -197°C/77K (liquid nitrogen) had a detrimental effect on the photosynthetic activity only for the LRN treatment but absolutely no effect on treatments LNR and LNR_LNR, as compared with control LR treatment (Fig. 1A, 1B). It seems that transition to dry state contributes to the resistance of the lichen photobiont to extremely low temperatures. Chlorophyll level (Fig. 1C), as well as the polarographic measurements of photosynthesis (Fig. 1D), confirms the above observation for LNR and LNR_LNR treatments. Photosynthesis could be considered a marker expressing the functional integrity of the photobiont. The chlorophyll amount is significantly lower in L treatment, because photosynthesis is inactive under absolute dehydration conditions, and it is higher in LRN treatment. The latter implies that photosynthetic apparatus maintain its structure despite stress and chlorophyll level is high due to an increase in functional antenna size per reaction center (ABS/RC), as shown in Fig. 1B. Maximum net photosynthesis is decreased at LRN treatment (Fig. 1D), compared to control treatment LR, indicating that, although photosynthetic apparatus is intact, photosynthesis is suppressed. This is in agreement with the responses of microalgae to different abiotic stresses as previously described (Sfakianaki et al., 2006; Demetriou et al., 2007; Sfichi-Duke et al., 2008).
Ergosterol originates exclusively from the mycobiont partner, and its levels correlate with metabolic activity of the lichen (Dahlman et al., 2002). In the present work, the amount of ergosterol appears unchanged for all treatments with respect to control treatment LR (Fig. 2A), indicating the resistance of the mycobiont in the above-mentioned extreme conditions. Polarographic measurement of respiration, which is performed mainly by the mycobiont, shows that respiratory levels are significantly higher for the LRN treatment compared to control (Fig. 1D). This could be explained by the fact that LRN treatment inflicts a greater extent of stress and, consequently, respiration is increased in order to produce more energy that is necessary for lichen to cope with the stress. In the same figure, it is interesting to observe that LNR_LNR respiratory levels are higher compared to LNR treatment and similar to the corresponding one of the control treatment LR. It seems that in LNR_LNR treatment the lichen is able to better adapt its metabolism after being exposed to dehydration and extremely low temperature, twice raising the prospect of a preconditioning response.
Another experiment that confirmed the above observation is the measurement of potassium cation efflux. According to Puckett et al. (1977), K+ is released when cells are disrupted under stress conditions. In this work, L and LNR treatments do not differ from control (Fig. 2B), confirming lichen resistance to dehydration and extremely low temperature. LRN treatment has slightly higher efflux of K+, indicating that only some cells were affected and that lichen, under this treatment, could recover. So far it is well established that lichen's mycobiont and photobiont partners are tolerant to a great extent to dehydration and extremely low temperatures at least when dehydration is preceded by extremely low temperature (LNR and LNR_LNR treatments).
Previous reports refer to polyamines as molecules with an important role in stress tolerance (Pirintsos et al., 2004, 2009). Polyamines are involved in the signaling cascade for the activation of stress-related mechanisms (Cramer et al., 2011). In fact, a high ratio of Put/(Spd+Spm) shows that the organism is tolerant to stress, while a low ratio of Put/(Spd+Spm) shows the opposite. In this study, polyamine levels were quantified for all treatments, and the Put/(Spd+Spm) ratio was determined. This ratio is significantly lower in LNR, LNR_LNR, and LRN treatments, compared to control (Fig. 3). This observation is in line with previous studies, which report the decrease of Put/(Spd+Spm) ratio when the organism is under stress (Navakoudis et al., 2003; Demetriou et al., 2007; Ioannidis and Kotzabasis, 2007; Sfichi-Duke et al., 2008; Ioannidis et al., 2012; Ioannidis and Kotzabasis, 2014). In fact, LRN treatment has the lowest Put/(Spd+Spm) ratio, in accordance with the fact that it appears to be the most stressed one. The decrease of polyamine levels is linked with the signaling cascade that will eventually activate mechanisms to protect lichen thalli from extreme stress conditions. Here, it is clear that these mechanisms have been successfully activated, at least for LNR and LNR_LNR, as they do not exhibit any sign of stress.
These mechanisms further include changes in the level of lipids and carbohydrates. It is known that the fluidity of membranes in general decreases with decreasing temperature. The ability to withstand temperatures below freezing relies on two strategies: protection of the cells from ice formation by freezing avoidance and, if ice forms, protection from damage during thawing (Clarke et al., 2003). Lipids have a protective role under abiotic stress conditions (Gigon et al., 2004). They maintain the integrity of plasma membrane by increasing the ratio of unsaturated fatty acids to saturated fatty acids. In this study, the total amount of lipids (per dry weight) increases when lichen is in a dry state (L treatment), compared to control LR (Fig. 4). The same pattern is also observed for carbohydrates, which are considered as osmolytes regulating cell osmosis under dehydration and extremely low temperature (Hájek et al., 2009a, 2009b). Lichen in L treatment has the highest amount of intracellular carbohydrates of all treatments (Fig. 5). Intracellular carbohydrates are accumulated in order to maintain cell structure under complete dehydration (Shao et al., 2006). The rest of the treatments (LNR and LNR_LNR, LRN) do not differ significantly from control (LR).
4.2. Dehydration and extremely low temperature do not affect the ability of lichens to produce high yield of H2
So far, it is clear that both the photobiont and the mycobiont partner of lichen survive the combination of prolonged desiccation and extremely low temperature (-196°C/77K) (treatments LNR and LNR_LNR). Recent study from our laboratory proved for the first time that lichens produce a high amount of molecular hydrogen (Papazi et al., 2015). In a closed system, the mycobiont partner consumes oxygen through respiration after 48 h, establishing hypoxic conditions, and the photobiont partner through a hydrogenase produces hydrogen using the dark fermentation pathway (Papazi et al., 2015). In order to ascertain that the metabolic activity of lichen in LNR and LNR_LNR treatments remains intact, their ability to produce molecular hydrogen was examined. It was found that the lichen, under both treatments, is able to produce approximately the same amount of molecular hydrogen as control treatment LR (Fig. 6), irrespective of light or dark conditions, which is in agreement with our previous results (Papazi et al., 2015). In that earlier work, we had found that lichens, through the dark fermentation pathway, produce equally high amounts of hydrogen under dark as well as under light conditions. It is interesting that lichen is active even after oxygen-depleted conditions have been established after 48 h. This indicates that regenerated lichen tissue may be tolerant to another stress factor characteristic of space: absence of oxygen.
To further investigate this hypothesis, LR and LNR treatments were incubated for 5 days in a closed system while oxygen-depleted conditions were established on the second day (Fig. 6). There is a slight decrease in the amounts of chlorophylls on day 1, but after this, chlorophyll, remains about the same for both treatments (LR and LNR) and all days (Fig. 7), indicating that the structure and function of the photosynthetic apparatus is not considerably affected by the absence of oxygen. The inactivation of PSII is necessary for the production of molecular hydrogen, but the photosynthetic apparatus remains intact and functional until Day 5. The same observation is made for ergosterol levels of the mycobiont partner (Fig. 8). There is a small decrease in ergosterol level on Day 1 most probably due to the absence of oxygen, which is necessary for its biosynthesis (Toshumi et al., 1978). Nevertheless, ergosterol levels remain relatively stable until Day 5. That means that the mycobiont partner is also metabolically active during the incubation under oxygen depletion. In conclusion, both partners of lichens seem to be tolerant under desiccation, extremely low temperatures, and oxygen deprivation for at least 5 days.
Polyamines were identified and quantified in LR and LNR treatments in order to assess the level of stress. It is remarkable that cadaverine (Cad), a polyamine that is related to stress (Jancewicz et al., 2016), is increased only on day 5 (Fig. 9). It is possible that Cad is accumulated in order to activate stress-related mechanisms against anoxic conditions (Hunter et al., 1983; Crawford et al., 1994; Yan et al., 1996; Chirkova et al., 1998; Blokhina et al., 1999). Total lipids remain on the same level until day 5 for all treatments (Fig. 10). It is important that in LNR treatments, although total lipids are almost unchanged, the fatty acids C13:0 (tridecanoid acid), C14:0 (tetradecanoid acid), and C18:1 (E-9-octadecenoic acid) disappeared, whereas the fatty acid C10:0 (decanoic acid) strongly increased. It is obvious that changes in specific fatty acids are connected with the ability of lichen to adapt in three extreme conditions: complete dehydration, extremely low temperature, and hypoxia.
In plants, abiotic stress alters the expression of stress-related genes, some of which are linked to histone modifications and nucleosome density (Kim et al., 2008, 2012; To and Kim, 2014). In addition, it has been reported that drought stress (Kim et al., 2015) and cold stress (Chinnusamy et al., 2007; Matsui et al., 2008) can induce additional epigenetic modifications, such as DNA methylation, but there is no evidence up to now about epigenetic modifications in nonmedical lichens. In order to assess whether the polyextremophilic behavior of the lichen, documented during the initial stages of the present study, is also related to DNA methylation pattern changes, MSAP analyses were conducted (Fig. 12). To the best of our knowledge, this is the first time that the MSAP methodology has been applied to lichens, and the results support the view that epigenetic mechanisms are included in the group of mechanisms that allow lichens to cope with extreme conditions.
5. Conclusion
Our findings indicate that the lichen, as a micro-ecosystem, is tolerant to three extreme conditions [complete dehydration, extremely low temperature (-196°C/77K), and hypoxia] while specific mechanisms are activated allowing lichen to cope with these stress conditions through an ecological integrity. Dehydrated thallus (L treatment) is able to cope successfully with temperatures that reach -196°C/77K (LNR and LNR_LNR treatment). Not only that, but it is also able to produce molecular hydrogen under anoxic conditions in light and darkness. The molecular hydrogen could then be used as fuel to meet energy demands in space with high efficiency and without pollution burden. Thus, this work also demonstrates for the first time that this unprecedented polyextremophilic characteristic of lichens could be linked with biotechnological applications under extreme environmental conditions. These facts further support (a) the idea of lichen as an ideal micro-ecosystem for potential astrobiological applications in space missions, as well as the argument that the theory of panspermia could be extended to incorporate the clarification that in the traveling entities not only single organisms but also micro-ecosystems like lichens could participate and (b) the importance of lichens as model systems for astrobiotechnological applications (hydrogen production), such as in the development of bioregeneration systems for extraterrestrial environments.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Associate Editor: Petra Rettberg
Associate Editor: Petra Rettberg
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.