
Abstract
One of the most important laboratory animal species is the nematode Caenorhabditis elegans, which has been used in a range of research fields such as neurobiology, body development, and molecular biology. The scientific progress obtained by employing C. elegans as a model in these areas has encouraged its use in new fields. One of the new potential applications concerns the biological responses to hyperacceleration stress (g-force), but only a few studies have evaluated the response of multicellular organisms to extreme hypergravity conditions at the order of magnitude 105 x g, which is the theorized force experienced by rocks ejected from Mars (or similar planets). Therefore, we subjected the nematode C. elegans to 400,000 x g (equivalent to that force) and evaluated viability, general morphology, and behavior of C. elegans after exposure to this stress. The metabolic activity of this nematode in response to the gravitational spectrum of 50–400,000 x g was also evaluated by means of the MTT assay. Surprisingly, we found that this organism showed no decrease in viability, no changes in behavior and development, and no drastic metabolic depression after hyperacceleration. Thus, we demonstrated for the first time that this multicellular research model can withstand extremely high g-forces, which prompts the use of C. elegans as a new model for extreme hypergravity. Key Words: Caenorhabditis elegans—Hypergravity—Ultracentrifugation—Acceleration—Panspermia—Astrobiology. Astrobiology 18, 825–833.
1. Introduction
T
Among the most recent applications of C. elegans as a model organism is the study of anhydrobiosis, that is, the tolerance to extreme desiccation regimes (Erkut et al., 2011, 2013). In addition, C. elegans displays tolerance to other types of stress such as low temperatures (Ujisawa et al., 2014) and interesting responses to radiation (Onodera et al., 2010; Tang et al., 2016). Life's resilience to abiotic stresses such as extremes of temperature, radiation, and diverse gravitational conditions is of great interest within the discipline of astrobiology, that is, the study of life in the Universe (Allen, 2001). In this context, research efforts on the physiological effects caused by variations in gravitational forces have also employed C. elegans as a model (Johnson and Nelson, 1991; Honda et al., 2012, 2014; Gao et al., 2015).
More specifically, some studies were successful in cultivating C. elegans in space. Nelson et al. (1994a, 1994b), for example, exposed this nematode to space environment for a period of time equivalent to two C. elegans life cycles. Surprisingly, the worms analyzed after this period did not present gross behavioral changes and reproduced normally (Nelson et al., 1994a, 1994b).
Szewczyk et al. (2005) reported that C. elegans could withstand re-entry into Earth's atmosphere without special protection, as evidenced by worms collected from the Space shuttle Columbia, which disintegrated in 2003 during its return. Live C. elegans worms, maintained on “C. elegans maintenance medium” (CeMM) and in “nematode growth medium” (NGM), were found in five recovered canisters. Only animals grown on CeMM maintained their reproductive capabilities, whereas individuals grown on NGM were arrested in dauer larvae or L1 stage. However, when worms present in each medium were recultivated, they grew normally. In addition, although altered phenotypes were observed in some individuals, they were not heritable. Owing to several variables that interfered in the experiment after the Columbia explosion, it was not possible to evaluate the physiological responses of C. elegans to the space environment (Szewczyk et al., 2005).
Despite the ability of C. elegans to maintain viability and reproductive capacity after spaceflight (Honda et al., 2012, 2014; Qiao et al., 2013; Higashibata et al., 2006), Hartman et al. (2001) observed that there was an increased number of mutations in worms sent to space due to exposure to cosmic radiation (Hartman et al., 2001). Moreover, the exposure of this organism to hypogravity conditions, for example, might be used to understand changes in perception of orientation that occur in astronauts in outer space (Clark et al., 2015).
Furthermore, C. elegans has also been used as a model to understand the effects of hypergravity on genetic and cellular physiological processes (Sasagawa et al., 2003; Kim et al., 2007; Kalichamy et al., 2016; Saldanha et al., 2016). However, the values of hypergravity tested to date do not exceed 1.6 × 103 x g (Johnson and Nelson, 1991), which is much lower than the hypergravity conditions to which a hypothetical organism would be exposed during ballistic and lithopanspermia scenarios. The ballistic panspermia, also known as “interplanetary panspermia,” argues that rocks expelled from a planet's surface would serve as transfer vehicles for spreading life between planets within a solar system. The lithopanspermia idea deals with the possibility of life's communication between two celestial bodies via rock fragments (Burchell, 2004). In this context, by means of mathematical calculations, Mastrapa, et al. (2001; based on Melosh, 1993) estimated that the hyperacceleration conditions to which an organism would be exposed in these scenarios would be on the order of 105 x g. Thanks to the studies of Ernest Mach and Albert Einstein, we know that acceleration and gravity are equivalent forces (Halliday et al., 1993; Russomano et al., 2007). Thereunto, studies focused on understanding the response of life to hypergravity can take direct advantage of ultracentrifugation to simulate such conditions.
During the first decades of the 20th century, high-performance ultracentrifuges were developed, making the exposure of diverse organisms to accelerations up to 105 x g possible. Accordingly, recent studies have demonstrated that the bacteria Pseudomonas denitrificans and Escherichia coli, and the algae Chlamydomonas sp., can withstand remarkable conditions of 403,627 and 670,000 x g, respectively (Deguchi et al., 2011; Gao et al., 2013). These studies have opened a new avenue of investigation: life's resilience to extreme hyperacceleration regimes. These reports focused on the analyses of unicellular organisms, and unfortunately there is only a limited number of studies that have evaluated the response of other organisms to extreme conditions of hypergravity. Thus, in this study, we evaluated the effects of exposing C. elegans to 400,000 x g on its viability, behavior, development, and metabolism, thus contributing to elucidate the response of multicellular organisms to extreme hyperacceleration regimes.
2. Materials and Methods
2.1. Nematode maintenance
C. elegans, kindly provided by Prof. Alan Tunnacliffe (University of Cambridge, United Kingdom), was maintained in the dark, at 20°C, on NGM agar plates and fed with a layer of E. coli (OP50 strain). Mixed populations, composed of all life cycle stages, were used in viability and behavioral analyses, whereas synchronized L2 larvae populations were used in developmental and metabolic analyses.
2.2. Viability assay
Initially, NGM agar plates were rinsed with M9 buffer to dislodge and collect worms, which were subsequently washed three times with M9 buffer to remove any excess of E. coli bacteria. Worms were then subjected to ultracentrifugation as described hereunder. Thereafter, we performed a survival assay using a modified version of a protocol, which has been used for isolated cells (Krause et al., 1984) and has already been successfully used for another nematode species (Souza et al., 2017). In brief, the supernatant was removed and erythrosin B dye was added (0.4% w/v in M9 buffer). After 1 h, samples were washed three times with M9 buffer to remove any excess of dye. Dead worms were stained in pink, whereas live worms remained unstained.
2.3. Synchronization
Mixed populations of C. elegans (i.e., containing eggs, all larval stages, and adult individuals) were subjected to synchronization through bleaching. In brief, mixed populations were exposed to a bleaching solution (NaOH 1%; NaClO 30%) for 10 min, which disintegrates worms and leaves only eggs. Subsequently, three washes were performed with distilled water intercalated with centrifugations at 2000 x g for 30 s. The remaining eggs were deposited on agar NGM plates containing OP50 bacteria and were maintained in the dark at 20°C. After 14 h, plates were rinsed with M9 buffer to dislodge and collect L2 larvae, which were used in the developmental and MTT experiments.
2.4. Experimental groups
The experiments were carried out and encompassed three groups: negative control 1 (NC1), negative control 2 (NC2), and experimental group (EG). NC1 was composed of worms that were not subjected to centrifugation and were maintained at 20°C for 1 h. NC2 was composed of noncentrifuged worms maintained at 4°C for 1 h. EG was composed of worms centrifuged at 400,000 x g for 1 h at 4°C. In the MTT experiment, EG included other hyperacceleration conditions (i.e., 50, 100, 1000, 10,000, or 100,000 x g) for 1 h at 4°C.
2.5. Hyperacceleration assays
Conditions of high g-force (hypergravity) were achieved by centrifugation at 4°C for 1 h. Experiments were performed on Eppendorf centrifuge models: 5417R (50 and 1000 x g), 5415R (100 x g), and 5403 (10,000 x g), all from the Department of Biology, FFCLRP, University of São Paulo (USP). Centrifugation at 100,000 and 400,000 x g was performed in the Optima MAX-XP high-performance ultracentrifuge—Beckman Coulter (Department of Biochemistry, FMRP, USP). After placing samples into the ultracentrifuge, hypercentrifugations (≥100,000 x g) started only when ideal conditions were reached (1.34 Pascal inside the centrifuge chamber).
The time needed to reach full speed (top g-force) is 0.5 s for 50 x g, 1 s for 100 x g, 3 s for 1000 x g, 10 s for 10,000 x g, 120 s for 100,000 x g, and 300 s for 400,000 x g.
2.6. Morphological and behavioral analyses
Morphology and behavior of worms centrifuged at 400,000 x g at 4°C for 1 h were analyzed within 60 min after the experiment. Images and videos were generated with a DCM800 digital camera coupled to a Labomed TCM400 inverted light microscope, through ScopePhoto software. The parameters evaluated in the morphological analyses were based on visible features under light microscopy such as the general size, structure, and integrity of the eggs and worms (vulva, pharynx, and cuticle). Behavioral analysis evaluated the following parameters: (1) spatial locomotion pattern, (2) lethargy or difficulty in locomotion, (3) presence of twitching (i.e., uncontrolled muscle contraction), and (4) presence of rolling (circular movement around the body axis). Three biological replicates were performed, each one consisting of three technical replicates (each one comprising all three groups: NC1, NC2, and EG). N = 600 worms/technical replicate/group. Statistical analyses (one-way analysis of variance [ANOVA]) were performed comparing only the groups within the same time point.
2.7. Development analysis
C. elegans L2 larvae were centrifuged at 400,000 x g at 4°C for 1 h, and subsequently 30 worms (of each group: NC1, NC2, or EG) were deposited on NGM agar plates containing OP50 bacteria. The plates were maintained at 20°C for 10 days in the dark. After this period, viability (live worms/total number of worms) and population growth (total number of worms after 10 days x 100/number of worms initially plated) were determined. Three biological replicates were performed, each one consisting of three technical replicates (each one comprising all three groups: NC1, NC2, and EG). N = 30 worms/technical replicate/group. Statistical analyses (one-way ANOVA) were performed comparing only the groups within the same time point.
2.8. Measurement of metabolic activity
Initially, 150 larvae (L2) were separated in 100 μL of M9 buffer. Subsequently, 100 μL of MTT solution (10 mg/mL) were added in each tube. After that, they were centrifuged at 50, 100, 1000, 10,000, 100,000, or 400,000 x g at 4°C for 1 h. Metabolic activity was measured by using the MTT assay according to the work of James and Davey (2007) with some modifications. After each experiment, worms were immediately immersed in liquid N2 to interrupt the MTT metabolism. Subsequently, the solutions containing worms were centrifuged at 500 x g and the supernatant was discarded. Then, 200 μL of dimethyl sulfoxide (DMSO) was added to each tube, and after 1 h, 100 μL of the solution was collected and transferred to a 96-well plate for analysis with the Epoch Biotek spectrophotometer and the Gen5 2.06 program. Results were obtained by means of an absorbance reading at 575 nm (James and Davey, 2007). As internal control, only blank tests with the DMSO were analyzed. The comparison between the various g-forces was carried out by a normalization that considered the longer periods of acceleration and deceleration involved in exposures to 100,000 and 400,000 x g. Each group (NC1, NC2, or EG) for each hyperacceleration regime (50–400,000 x g) was composed of six technical replicates. Each individual technical replicate encompassed 150 larvae. A total of 7200 worms were included in the entire experiment. Statistical analyses (one-way ANOVA) were performed by comparing only the groups within the same time point.
2.9. Statistical analysis
Except for MTT assay (six technical replicates), all experiments were performed in biological triplicates (each one consisting of technical triplicates), and data are presented as mean values and standard deviations. Statistical analyses were performed using “one-way ANOVA” (with Student Newman post hoc). Statistical differences were considered significant when p ≤ 0.05. Identical letters indicate those groups that are not statistically different. Distinct letters indicate those groups that are statistically different.
In some cases, one group may be statistically not different from only one specific group, within a larger set of groups indicated with a different letter. In these specific cases, those groups were indicated with hashtag (#), meaning they are not statistically different.
3. Results
Interestingly, C. elegans' viability did not decrease after exposure to 400,000 x g, that is, the values remained practically the same as those observed in the two controls (NC1 and NC2) (Fig. 1). Moreover, population growth that was measured 10 days after ultracentrifugation also did not reveal any decreases in reproductive potential or viability (Fig. 2). Likewise, morphological and behavioral analyses did not identify any structural or behavioral changes in C. elegans exposed to 400,000 x g (Fig. 3). MTT analysis indicated a significant (p < 0.05) decrease in the metabolism between NC1 and the EG (50–400,000 x g). A significant (p < 0.05) decrease in metabolism was also observed between NC2 group and those exposed to 50, 1000, 10,000, and 400,000 x g. Together, these data suggest a reduction in metabolic rate due to centrifugation; however, this effect is not proportional to the hyperacceleration regime, instead it is stabilized (Fig. 4).
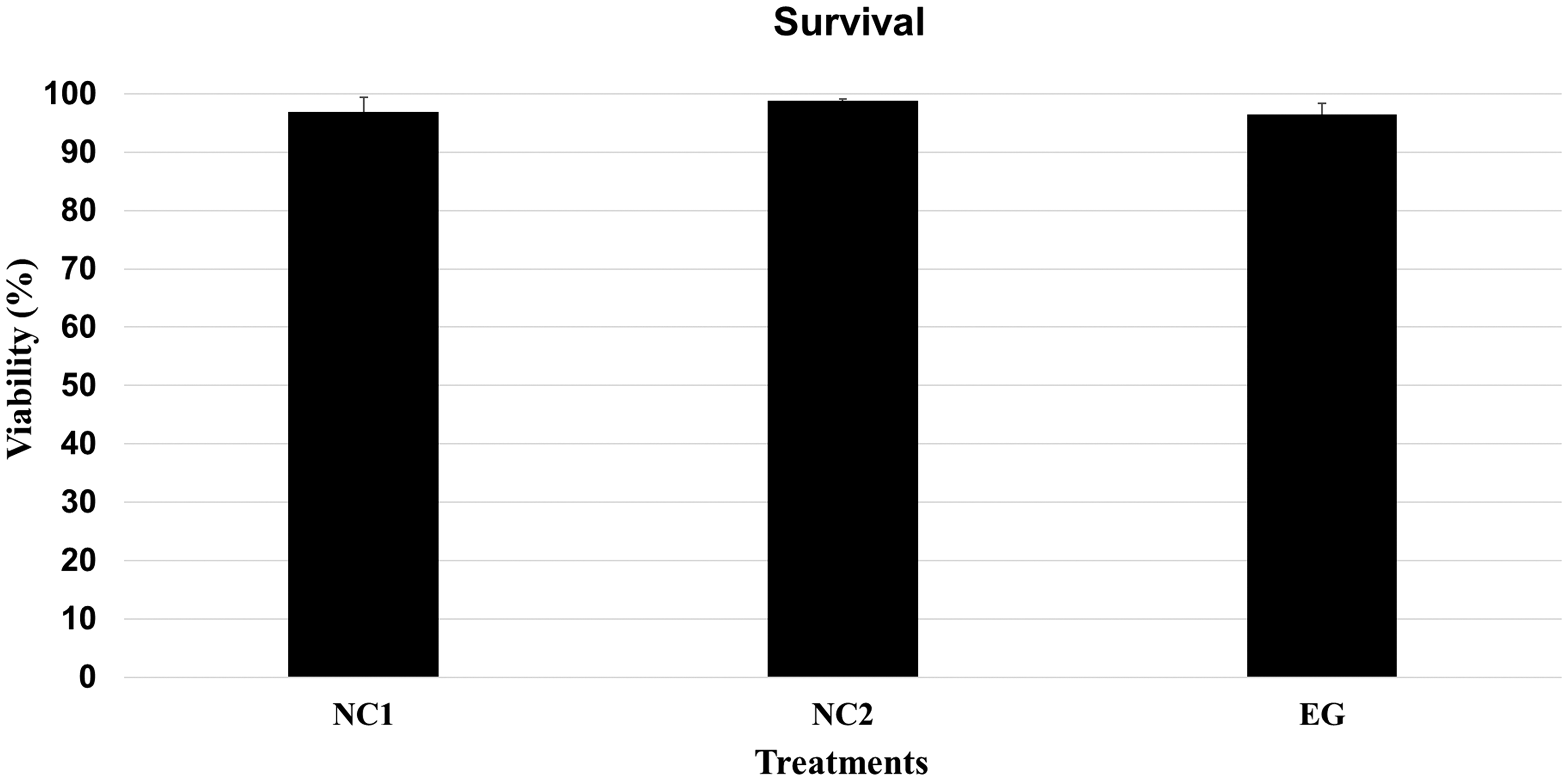
Survival percentage (viability) of Caenorhabditis elegans after exposure to 400,000 x g for 1 h. NC1: noncentrifuged worms in M9 solution at room temperature (20°C) for 1 h. NC2: noncentrifuged worms in M9 solution at 4°C for 1 h. EG: centrifuged worms in M9 solution at 4°C (400,000 x g for 1 h). The experiments were performed in technical and biological triplicates. Number of worms (mixed population) per experiment: 600. Different letters indicate statistically significant differences (p < 0.05). EG, experimental group; NC1, negative control 1; NC2, negative control 1.
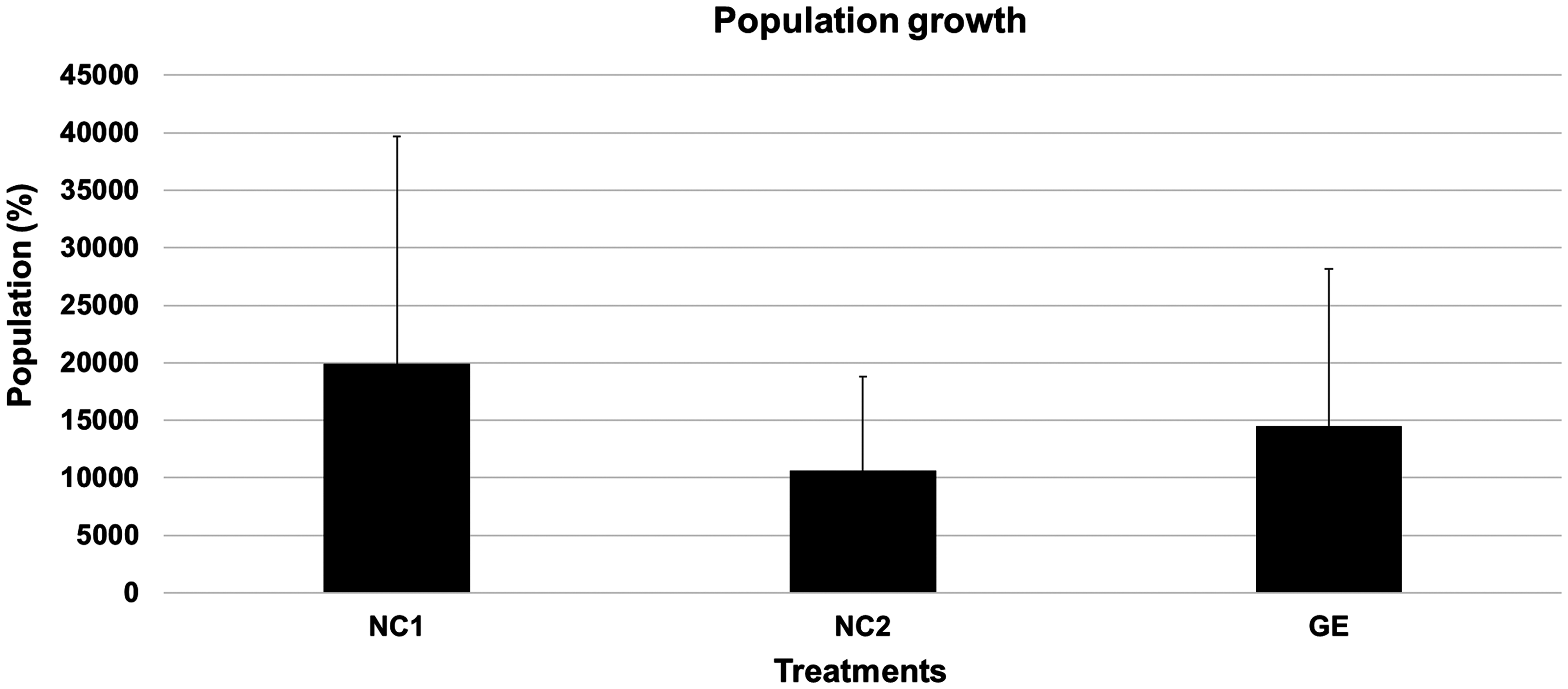
Population growth of C. elegans measured 10 days after exposure to the different experimental conditions. NC1: noncentrifuged worms in M9 solution at room temperature (20°C). NC2: noncentrifuged worms in M9 solution at 4°C for 1 h. EG: centrifuged worms in M9 solution at 4°C (400,000 x g for 1 h). The experiments were performed in technical and biological triplicates. Number of worms (input L2 larvae) per experiment: 30. No statistical differences were observed (p < 0.05).

Evaluation of C. elegans metabolic rate under different hyperacceleration regimes. NC1: noncentrifuged worms in M9 solution at room temperature (20°C) for 1 h. NC2: noncentrifuged worms in M9 solution at 4°C for 1 h. EG: centrifuged worms in M9 solution at 4°C (50–400,000 x g for 1 h). The experiments were performed in technical hexaplicates. Number of worms (L2 larvae) per replicate: 150. Different letters indicate statistically significant differences (p < 0.05).
4. Discussion
Currently, ultracentrifugation has been widely used to evaluate and characterize the physical, metabolic, and genetic responses of several organisms to g-forces (hypergravity regimes), generally in the range of 101–102 g ( Casey et al., 2012; Tamaoki et al., 2014; Kim et al., 2016).
Comparatively, few studies have investigated the effects >104 x g. For example, Vasanthan et al. (2017) performed a seminal analysis on the resistance of tardigrades to hypergravity. They showed that >80% of a Hypsibius dujardini population tested were able to withstand up to 16,060 x g for 1 min; however, only 50% were still alive 11 days postcentrifugation. Although they observed a decrease in survival (<20%) and in egg production (up to ∼70% after 30 days) in the gravitational spectrum between 3421 x g and 16,060 x g, no major morphological alterations were noted (Vasanthan et al., 2017). In our analyses, C. elegans maintained a viability of >90% not only in controls but also in the EG, with no decrease in viability due to exposure to hypergravity. Moreover, as observed in H. dujardini, no morphological changes were identified in C. elegans.
More importantly, since H. dujardini is able to enter into anhydrobiosis—a condition of suspended animation brought about by desiccation such that tolerance to a number of physical stresses is attained (reviewed in Tunnacliffe and Lapinski, 2003)—this organism's potential as a model in astrobiology research is undeniable. Therefore, both H. dujardini and C. elegans represent potential models for further studies evaluating the effects of hypergravity on multicellular organisms.
Remarkably, Beams and King (1936) exposed eggs of the nematode Ascaris suum to a condition similar to that adopted in the current work (400,000 x g for 1 h). These authors assessed cell divisions of the centrifuged eggs as a parameter with which to evaluate viability after the ultraacceleration. Interestingly, they observed that after 48 h, 90% of the eggs had cells that divided at least once (Beams and King, 1936). According to the authors, most of the organisms are expected to die when subjected to stress of such magnitude due to physical and structural damage that does not affect Ascaris eggs owing to mechanical protection provided by the shell. Therefore, survival of nematode larvae and adults to this magnitude of stress was not considered plausible.
We have recently shown that the anhydrobiotic nematode Panagrolaimus superbus is able to withstand 400,000 x g while in the extremely stable state of anhydrobiosis. Unexpectedly, the control group that comprised fully hydrated living worms was also tolerant to the test conditions (Souza et al., 2017). These two findings (Beams and King, 1936; Souza et al., 2017) prompted us to evaluate whether C. elegans is also tolerant to such stress.
The first and most important analysis performed in this study was the determination of the viability percentage of C. elegans subjected to 400,000 x g (Fig. 1). Its notorious tolerance to this degree of physical stress allowed subsequent morphological (possible structural damages) and behavioral analysis. C. elegans may exhibit several behavioral alterations that are induced in response to stressors. In cases of large agglomeration, absence of food, or exposure to high temperatures, for example, C. elegans can enter into an alternative state of development known as dauer larva (Golden and Riddle, 1982; Ailion and Thomas, 2000). In this larval stage, C. elegans' metabolism changes and it becomes lethargic (Hu, 2006).
In addition, C. elegans may present other phenotypic changes such as rolling and twitching. Feeding, reproductive activity, and oviposition are other parameters that were evaluated during behavioral analyses and can be altered in mutants (Moerman et al., 1982) or by changing assay conditions (Hart, 2006). Surprisingly, no obvious or dramatic changes in the normal behavioral pattern were observed after centrifugation at 400,000 x g. Centrifugations at 3000 x g for 1 min are frequent during various laboratory procedures involving C. elegans. Any researcher working with this species may have already observed that worms are very active soon after such routine procedures, something intuitively unexpected since the same g-force would be lethal to humans. Thus, it is noteworthy that these worms presented apparently normal agitation (swimming in M9 buffer) moments after their removal from the ultracentrifuge (400,000 x g), indicating that this ultraacceleration is innocuous to them. The absence of any changes in viability, morphology, and behavior verified after 400,000 x g centrifugation indicates that this nematode can withstand extremely high g-force values. However, the possible existence of internal structural damage caused by ultracentrifugation would not necessarily be detectable after the experimental procedure.
Therefore, the results of the developmental analysis were especially important to corroborate the morphological data (Fig. 3). The absence of changes in the population growth 10 days after the exposure to hypergravity indicates that ultracentrifugation at 400,000 x g does not affect a worm's viability and does not compromise its correct development and reproductive potential (Fig. 3). Taken together, these data reveal that C. elegans is not only tolerant to this stress but may have the capability as well to tolerate much greater g-forces (500,000 x g or beyond).
In addition to these analyses, evaluation of MTT conversion in C. elegans provided an assessment of the metabolic status within different hyperacceleration conditions (50–400,000 x g) (Fig. 4). Changes in metabolism may occur in response to various factors and are observed in virtually every animal phylum (Guppy and Withers, 1999). In the parasitic nematode Anguina tritici, for example, the metabolic rate may reach 0.06% of the normal metabolism when it is subjected to extreme desiccation (Guppy and Withers, 1999; apud Bhatt and Rohde, 1970). Interestingly, the data presented in this work show that C. elegans can tolerate 400,000 x g without going into such a deep state of metabolic depression. Moreover, our data reveal something new: even after subjecting the worms to a g-force five orders of magnitude higher than that naturally experienced on Earth (1 x g), they remain alive and metabolically unchanged concerning the 50 x g group.
C. elegans is an important species in biology and has recently been shown to be an interesting model with which to study several different conditions such as epilepsy (Williams et al., 2004), sleep-like states (Raizen et al., 2008), tolerance to extreme desiccation (Erkut et al., 2011, 2013), and photoperception (Ward et al., 2008). The data obtained by our research group reveal for the first time that this worldwide-known research model has one more intriguing feature in its biological repertoire: tolerance to extreme hyperacceleration. Our analyses indicated that C. elegans flexibly supports forces at least up to 400,000 x g for 1 h. Perhaps more impressive is the fact that the metabolism is not dramatically affected by such a physical stress.
The exact process by which life began on Earth is unknown. An alternative hypothesis, denominated panspermia (Burchell, 2004; Świeżyński, 2016), considers the possibility that “seeds of life” had seeded Earth, thus shifting the focus from the “the origin of life” and giving greater attention to experiments that demonstrate the plausibility of the “transfer of forms of life between celestial bodies.” A number of studies focused on the transfer of life align with the hypothesis that the planet Earth was initially colonized by life from Mars (McKay et al., 1996). The lithopanspermia concept considers the possibility of the transport of microorganism between planets by means of rock fragments (Nicholson et al., 2000, 2005; Horneck et al., 2008; Fajardo-Cavazos et al., 2007; Nicholson, 2009). In this context, volcanic activity is considered one of the natural phenomena that occur on several planets and could eject mass containing life-forms toward space, both in lithopanspermia and interplanetary panspermia scenarios. Interestingly, there is evidence that >53,000 meteorites have impacted Earth, indicating an intense exchange of rock material between planets (Vasanthan et al., 2017). Among these meteorites, >100 have a proven martian origin (Meyer and Righter, 2017). Several analyses have estimated that the escape velocity of Mars is 5.03 km/s (Fajardo-Cavazos et al., 2009), and diverse ballistic experiments have been performed to simulate such conditions (reviewed and discussed in Roten et al., 1998). Burchell et al. (2001) fired projectiles loaded with Rhodococcus sp. at a speed of 4.5 ± 0.6 km/s. The projectiles were shot against a variety of materials (rock, glass, and metal). In four impacts of bacteria on rocks, it was possible to identify the formation of colonies, which were not observed when the substrate was glass and metal (Burchell et al., 2001).
Other microorganisms such as Bacillus pumilus, Bacillus subtilis, and Deinococcus radiodurans have also been shown to be effective models with which to demonstrate resilience of life to hypergravity associated with ejection scenarios (Mileikowsky et al., 2000; Mastrapa et al., 2001; Benardini et al., 2003). Moreover, analyses have shown that the freezing of rocky fragments containing microorganisms would not be a hindrance to the interplanetary communication of life (Burchell et al., 2003). Recently, Burchell et al. (2014) demonstrated that even frozen fossilized diatoms can be preserved in scenarios of extreme shock induced by hypervelocity impacts, indicating for the first time that it would be possible to transfer fossils from Earth to the Moon by means of rocky bodies ejection and impact. In lithopanspermia scenarios, the organism must be resistant not only to hypergravity associated with ejection but also to space travel and re-entry through another planet's atmosphere (Mastrapa et al., 2001). In the case of the nematode C. elegans, some experiments have already demonstrated that it has the capability to survive under conditions of spatial hypogravity (Nelson et al., 1994a, 1994b; Hartman et al., 2001; Ishioka and Honda, 2012) and to withstand re-entry into Earth's atmosphere (Szewczyk et al., 2005). However, none of the studies analyzing the exposure of C. elegans to hypergravity evaluated conditions compatible with an ejection scenario toward space, which characterizes the originality of our analyses.
Interestingly, Mastrapa et al. (2001) calculated the g-force that an organism present in a volcanic fragment from Mars would have to endure during its acceleration toward space. Using computational simulations, she concluded that the maximum ejection acceleration predicted for a rock fragment ejected from Mars would be 387,000 x g. Surprisingly, our results demonstrate that C. elegans can withstand conditions of greater force than that estimated by Mastrapa et al. (2001) without any impairments on viability, morphology, behavior, or metabolism, which encourages further study to decipher the genetic and physiological mechanisms involved in the resilience of this nematode to extreme hypergravity.
Two differences must be highlighted between Mastrapa's study and the present work. First, the acceleration time (0.5 ms) and jerk (i.e., the rate of change of acceleration; 6 × 109 m/s3) theorized for an expelled rock are very different from those reported here using ultracentrifuges (300 s and 1.3 × 104 m/s3, respectively). Second, once rocks achieve top speed, the g-force is no longer present, which is quite different from our experiments that subjected the nematode under continuous g-force for 1 h. Therefore, although the experimental jerk is 4.6 × 105 lower that the theoretical jerk, the experimental time of exposure to the hyperacceleration is 7.2 × 106 longer than the calculated time.
These differences are relevant and may lead to different outcomes; nevertheless, we believe our study represents an interesting experimental set up to be compared and analyzed along with Mastrapa's calculations.
Since C. elegans is an important animal model used worldwide, the biology of which is very well characterized (genome is sequenced, complete neuronal connectome is known, and life cycle is short and defined), its potential for study in the field of hypergravity is enormous.
5. Conclusions
Considering the data presented in this work, we conclude that C. elegans can be used as a new model for studies on the effects of hypergravity at different levels (molecular, morphological, behavioral, and populational). Future efforts to explore the maximum tolerance of C. elegans and other species to hypergravity may be done once ultracentrifuges with the capability of reaching astonishing g-forces are in continuous improvement (e.g., 1,000,000,000 x g) (Gray, 1951). Furthermore, our study helps to discern the limits of the gravitational spectrum compatible with life, and assess the hypothesis that life may exist on other planets.
Footnotes
Acknowledgments
The authors thank Prof. Elza Tiemi Sakamoto Hojo and Dr. Ana Paula de Lima Montaldi for helping with MTT analyses. We are also thankful to Prof. Roy Edward Larson (FMRP/USP) and Gabriel Sarti Lopes for granting us access to the ultracentrifuge. T.A.J.S. is recipient of studentship from CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brazil).
Authors' Contributions
T.A.J.S. and T.C.P. have participated in the conception and design of this work and the analysis of the data, as well as the writing of the article and taking public responsibility for its content.
Author Disclosure Statement
T.A.J.S. and T.C.P. certify that no parties have a direct financial interest in the results of this article. T.A.J.S. is recipient of studentships from CAPES.