
Abstract
Sedimentary strata on Mars often contain a mix of sulfates, iron oxides, chlorides, and phyllosilicates, a mineral assemblage that is unique on Earth to acid brine environments. To help characterize the astrobiological potential of depositional environments with similar minerals present, samples from four naturally occurring acidic salt lakes and adjacent mudflats/sandflats in the vicinity of Norseman, Western Australia, were collected and analyzed. Lipid biomarkers were extracted and quantified, revealing biomarkers from vascular plants alongside trace microbial lipids. The resilience of lipids from dead organic material in these acid saline sediments through the pervasive stages of early diagenesis lends support to the idea that sulfates, in tandem with phyllosilicates and iron oxides, could be a viable target for biomarkers on Mars. To fully understand the astrobiological potential of these depositional environments, additional investigations of organic preservation in ancient acidic saline sedimentary environments are needed.
1. Introduction
Although abiotic synthesis of some organic compounds may occur in hydrothermal settings (McCollom and Seewald, 2007), the vast majority of sedimentary organic matter reflects biological processes that are highly characteristic of the depositional environment. Biomarkers—organic compounds derived from living organisms—are particularly important in reconstructing Earth's microbial history before the Cambrian radiation emergence of readily fossilized body features (Simoneit et al., 1998). Biomarkers have been found in rocks and sediments that are associated with some of the earliest life forms on Earth, and they have been used to characterize both physical environments and primitive biological systems (see examples in Peters et al., 2007). Lipid biomarkers, in particular, are paramount in the identification of ancient bacterial metabolism that is preserved in Archaean and Proterozoic rocks (Summons and Walter, 1990), as well as in modern microbial communities (White, 1983). Microbial lipids that survive in the rock record typically have highly branched or cyclic hydrocarbons, such as isoprenoids and sterols, which are resistant to the early stages of diagenesis, during which time most degradation occurs. Often, the more polar membrane lipids such as fatty acids are rapidly degraded (De Leeuw and Largeau, 1993), although recent research demonstrates that select taphonomic windows exist that allow phospho- and glycolipids to survive initial diagenesis (Wilhelm et al., 2017, 2018).
If similar biomarkers formed and were preserved in the geological record on the surface of Mars, where would we be most likely to find them? The best way to answer this might be to evaluate biomarker preservation in analog mineral environments on Earth. In the vicinity of Norseman, Western Australia are dozens of acid salt lakes hosted by the deeply weathered Archaean rocks of the Yilgarn Craton (see Fig. 1). They range in diameter from m2 to km2, and they are marked by cycles of flooding, evapoconcentration, and desiccation (Benison et al., 2007), as well as by winds. Despite the extreme conditions (pHs as low as 1, salinity as high as 32% total dissolved solids, high fluxes of solar radiation, water stress during desiccation, high metal concentrations, and large shifts in diurnal temperature), these sites are highly habitable, and they are home to a diverse bacterial-, archaeal-, algal-, and fungal-rich microbiome (Mormile et al., 2009; Johnson et al., 2015; Zaikova et al., 2018; Aerts et al., 2019).

Sampling sites at ephemeral acid saline lakes in southern Western Australia.
The acid brine lakes and associated mudflats/sandflats have been interpreted as an excellent analogue for martian sedimentary deposits for three reasons (Benison and LaClair, 2003; Benison and Bowen, 2006). First, there are similar sedimentary textures and sedimentary structures, including laminations, wave ripple cross-bedding, and mudcracks in both terrestrial and martian environments, suggesting processes such as flooding, desiccation, and eolian reworking. Second, there are similar diagenetic features, including iron oxide concretions, displacive crystals, and sulfate veins, likely formed from acid saline groundwaters. Third and most importantly, the sedimentary mineral assemblage on Mars is quite similar to that of acid saline lakes in Western Australia.
The mineral assemblage at acid saline lakes and mudflats/sandflats in southern Western Australia is unusual and includes a siliciclastic component, chlorides, sulfates, iron oxides, phyllosilicates, and opaline silica (Benison et al., 2007; Story et al., 2010).
Halite, gypsum (and other sulfates such as epsomite and basanite), hematite, kaolinite, and, less commonly, jarosite, alunite, and silica precipitate directly from acid saline lakes as chemical sediments (Benison et al., 2007). Halite and gypsum also grow from acid saline groundwater centimeters below the surface of mudflats and sandflats adjacent to the lakes, making displacive halite and gypsum, as well as halite and gypsum intergranular cements (Bowen et al., 2013). Hematite, jarosite, alunite, and kaolinite also precipitate directly from acid saline groundwaters as early intergranular cements (Benison et al., 2007: Bowen et al., 2012). Hematite concretions form early in Western Australian acid saline lake and mudflat/sandflat sediments and contain hematite, jarosite, gypsum, halite, and kaolinite (Bowen et al., 2008). The siliciclastic component of the sediments includes quartz and feldspar sand and mud grains that are the products of the weathering of Archaean host rocks (Bowen et al., 2013).
Some of these minerals on Mars, particularly jarosite and alunite, serve as strong indicators of past acidic conditions. Jarosite, a hydrated iron sulfate that forms in acid saline waters (Nordstrom, 1982; Klingelhöfer et al., 2004), was first hypothesized on Mars by Burns (1987), then discovered by the Opportunity rover in 2004 (Klingelhöfer et al., 2004), suggesting that acidic conditions once dominated the landscape at Meridiani Planum. Jarosite has since been detected in other multiple sites on Mars, suggesting that it is a relatively common mineral (Cull et al., 2014; Léveillé et al., 2015). Alunite, a hydrated aluminum sulfate in the same family as jarosite, has been found in the Terra Sirenum region of Mars, within a spatially extensive formation within Cross crater, where in some areas it is closely associated with kaolinite. Alunite also strongly suggests low pH, sulfuric fluids at or near the martian surface (Ehlmann et al., 2016).
Oxidized minerals have long been considered weak targets for the preservation of organic molecules (Klein, 2005). Thermodynamics suggest that organics will quickly react with SO2- and Fe3+ (Sumner, 2004; Stumm and Morgan, 2012). The kinetics of organic carbon oxidation in the presence of iron is believed to be extremely rapid—months to years under most conditions—largely due to the generation of free radicals and H2O2 on the surface of iron-bearing minerals, and the calculations of Stumm and Morgan (1996) suggest that Fe(III)-bearing minerals are not stable at the low electron potentials that are necessary for organic carbon stability (Sumner, 2004; Stumm and Morgan, 2012). Sulfate reduction in the presence of organics, while slower in general, requires the transfer of more electrons and is believed to occur at a faster rate at low pH conditions (Ohmoto and Lasaga, 1982).
Mineral stability fields, however, depend critically on ion pairing in solution. This, in turn, depends on brine compositions and pH. Although significant progress has been made in this area—on how Mg, Ca, and Fe-sulfates might have formed from the evaporation of acidic fluids derived from weathering olivine-bearing basalt (Tosca et al., 2005), on how iron-bearing phyllosilicates might have formed from weathering and hydrothermal alteration of martian basalt (Catalano, 2013), and on how clay-bearing sequences might have formed through downward percolation and neutralization of acidic H2SO4–HCl solutions (Zolotov and Mironenko, 2016)—these compositions are not fully constrained for the early martian surface.
To analyze the astrobiological potential of mixtures of sulfates, iron oxides, and phyllosilicates in a highly acidic and saline terrestrial Mars analogue, this study investigated the types of biomarkers present and the range of biomarker recovery by using gas chromatography-mass spectrometry (GC-MS), which illuminates the chemical structure of biomolecules from characteristic fragment ions in their mass spectra.
2. Materials and Methods
2.1. Sample acquisition
Sediment samples were collected in June 2018 from four acid saline lakes within the vicinity of Norseman in southern Western Australia. The ambient air temperatures were high (up to 34.6°C). Lakes in the region are ephemeral; of those with standing surface water in June 2018, lake diameters were typically less than a few 100 m and no more than a meter deep. Lakes Aerodrome, Gilmore, Gneiss, and Typhoon were chosen because they had been previously studied as part of an initial biomarker study (Johnson, 2008) and had long records of geological and geochemical observation (e.g., Benison and LaClair, 2003; Benison and Bowen, 2006, 2015; Benison et al., 2007, 2008; Bowen and Benison, 2009; Story et al., 2010; Bowen et al., 2012, 2013; Johnson et al., 2015).
Sediment samples were collected from acid saline mudflats/sandflats ∼1 m from the active shoreline of flooded Lake Gneiss and Typhoon Lake. Other samples were collected from the lake facies of desiccated Lake Aerodrome and Lake Gilmore. At these sites, minerals and water chemistry change through time. At the time of sampling, Lake Gneiss was underlain by yellowish-brown mud and a thin halite layer of ∼1 mm in thickness. The nearby Typhoon Lake had a thicker halite layer, ∼3 mm in thickness; Lake Aerodrome, near an unpaved landing strip on the southern part of Cowan Basin, is composed of a bed of bottom-growth gypsum crystals up to 5 cm in length. At Lake Gilmore, the salt crust was much more prevalent, ∼1 cm in thickness.
Sampling in the field and subsequent laboratory procedures were conducted with sterile procedures to avoid contamination. At each sampling site, samples of ∼100-g wet weight were collected with organically clean spatulas and organically clean glass jars (precombusted at 550°C for 8 h) from just beneath the salt crust to a depth of ∼5 cm (Table 1). Samples were kept frozen until the time of analysis. Approximately 10 g of material was subsampled for mineralogical and particulate organic carbon and nitrogen analyses. The other 90 g was reserved for lipid extraction. For the sample sites where lake water was present, a water sample was also collected in a sterile 50-mL falcon tube.
Environmental Characteristics for the Four Mud/Sandflats Sampled in this Study
If present, pH and TDS were also measured on lake water. Previously observed pH data are from the supplementary information in Bowen and Benison (2009).
TDS, total dissolved solids.
2.2. X-ray diffraction
Samples were crushed via mortar and pestle and sieved to <63 μm for powder X-ray diffraction (XRD) analyses on a Bruker D8 Discover diffractometer. The patterns were acquired from 2 to 70° 2θ (Cu Kα radiation, λ = 1.54059 Å), and quantitative mineralogy was obtained by Rietveld refinement methods using the MDI (Materials Data Incorporated) Jade analysis software. Due to the poorly crystalline nature of certain phyllosilicate minerals (e.g., kaolinite, smectite), standard Rietveld refinement methods cannot be used to properly quantify the abundance of such phases. Among the samples analyzed, kaolinite was identified by its characteristic ∼7.2 Å peak and quantified by combining a whole pattern fitting technique with the Rietveld refinement of the crystalline fraction. The scale factor of a pure kaolinite pattern (Clay Mineral Society KGa-2; relative intensity ratio = 2.1) was optimized to yield an abundance relative to the crystalline phases. As will be discussed later in Section 3, the sample diffraction patterns show lower 001:02l kaolinite intensity ratios than expected for randomized patterns, indicative of acid alteration of the kaolinite (Pentrak et al., 2012). To minimize quantification uncertainties due to kaolinite alteration, the kaolinite abundance was estimated from 02l band scattering, a more stable measurement region compared with the 001 basal peak. Clay minerals were characterized by isolating the <2-μm size fraction and performing XRD on oriented mounts.
2.3. Contextual environmental measurements
For lakes where water was present, the water was analyzed for pH on return to the lab with a Mettler Toledo pH meter. Total dissolved solids were measured with VeeGee STX-3 handheld refractometer at 1:10 dilution with deionized water. Particulate organic carbon and nitrogen on all four mudflat/sandflat samples were measured according to standard protocols by using a CHN Elemental Analyzer at Woods Hole Oceanographic Institution Nutrient Analytical Facility, Massachusetts (Table 1).
2.4. Lipid extraction
Sediment samples of ∼90-g wet weight (mixtures of sand, mud, salt crystals, and groundwater) were freeze dried for 36 h. The samples were then crushed to a fine powder by using a solvent-washed (3× methanol, 3× methanol:dichloromethane 1:1, 3× dichloromethane) mortar and pestle. Lipids were extracted by using a modified Bligh and Dyer method (Bligh and Dyer, 1959) alongside an extraction blank to monitor for any signs of contamination. Extractions took place in prewashed 50-mL Teflon tubes that were ultra-cleaned by two sequential 30-min sonications in methanol and dichloromethane. Nineteen milliliters of a 10:5:4 mixture of methanol: chloroform: dichloromethane-washed H2O was added to each dry sample powder (of approximate weight 10 g) within the Teflon tubes. A mixture containing four internal standards—heneicosane (C21), 2-methyl-octadecanoic acid, 1-nonadecanol (C19) and p-terphenyl-d14—was then added to the total lipid extract (TLE).
The sample tubes were then vortexed for 5 min and sonicated for 20 min before centrifugation at 2500 rpm for 10 min. The liquid phase was transferred to a precombusted glass separatory funnel, and the process was repeated three times with the remaining sediment. For the final extraction, 19 mL of a 10:5:4 mixture of methanol: dichloromethane: H2O with 1% trichloroacetic acid was used; then, 30 mL of deionized water was added to the separatory funnel to aid in phase separation. The aqueous phase was re-extracted two more times by using dichloromethane.
To desulfurize samples, acid-activated and solvent-cleaned copper pellets were added to the 60-mL precombusted glass vial containing the TLE before drying it and allowed to sit overnight. The TLE was reduced to less than 50 μL under N2 gas.
2.5. Silica gel column separation
Approximately half of the total lipid extract was then applied to a 10-cm silica gel pipet column for each sample. Columns with each sample, alongside an extraction blank to monitor for signs of contamination, were sequentially eluted with 1.4 column-volumes of hexane and 2 column volumes of 8:2 hexane-dichloromethane to obtain the nonpolar fractions F1 and F2; this was followed by 2 column volumes of dichloromethane for fraction F3, 2 column volumes of 8:2 dichloromethane-ethyl acetate for fraction F4, and 3 column volumes of 7:3 dichloromethane-methanol for fraction F5. Each fraction was then gently evaporated under N2 gas. Because of an issue with clumping of the silica gel, the column fractions were lost for the Lake Gilmore mudflat/sandflat sample (for this reason, only the total lipid extract was analyzed on the GC-MS and only those results are reported in Table 2; for all other samples, results from the better separated fractions are reported in Table 2).
Detections Shown as Percentages of Total Alkanes, Fatty Acids, Alcohols, and Sterols in Each Sample

Darker shading indicates higher abundances.
2.6. Derivatization for GC-MS
Trimethylsilyl (TMS) reagents were used to derivatize (reducing the polarity of) functional groups in polar fractions F3, F4, and F5 before GC-MS analysis. Nonpolar fractions were reconstituted in 20 μL of hexanes. Ten microliters of bis(trimethylsilyl)trifluoroacetamide (BSTFA) in 10 μL of pyridine was added; then, samples were heated at 70°C for 30 min for the derivatization to occur. All polar fractions were run on the GC-MS within 36 h of derivatization.
2.7. GC-MS conditions
An injection volume of 1 μL from each fraction was analyzed on a ThermoFisher Trace 1310 gas chromatograph coupled to a ISQ-LT quadrupole mass-spectrometer (GC-MS). The GC is equipped with a programmed-temperature vaporization (PTV) inlet, a 30-m MXT-5 capillary column with 0.25-mm i.d., and 0.25-m film thickness; helium (purity ≥99.999%) was used as carrier gas. The PTV injector temperature was set at 330°C to prevent any condensation of the compounds analyzed. Injections were administered in split mode with a 5-mL/min split flow and a 1.2-mL/min constant carrier flow rate. The five fractions were analyzed by using the same GC temperature program: initial temperature of 60°C immediately followed by a 10°C/min ramp to 150°C, immediately followed by a second ramp temperature at 4°C/min to 350°C held for 10 min. This temperature program allowed all the compounds targeted to be analyzed while being well separated.
Both the MS transfer line and ion source temperatures were set at 300°C. The MS was set to scan the ions produced from the electron impact ionization source (70 eV) in the range m/z 30 to 600. A solvent delay of 6 min was applied to preserve the MS filament and prevent saturation of the GC column by the solvents associated with the fractions and the BSTFA derivatization reagent.
2.8. Peak identifications
Chromatograms and the mass spectra for different compounds were viewed by using the Xcalibur qual browser software from ThermoFisher. Identifications were based on spectral reduction by AMDIS software followed by NIST Spectral Library software matches when available. Otherwise, spectra were identified by comparison with a laboratory-acquired spectral library and retention times of known molecules.
3. Mineralogy Results
Mineralogical composition derived from XRD analyses of the samples is shown in Fig. 2. Quartz, halite, and kaolinite were present in all samples. Quartz was the dominant mineral in Aerodrome (93.5 wt.%), Typhoon (85.8 wt.%), and Gilmore samples (39.7 wt.%). Halite content varies between 2.4 (Aerodrome) and ∼16 wt.% (Gilmore and Gneiss).

Random powder XRD patterns and quantitative mineralogy for the mudflat/sandflat samples collected at
Aerodrome and Typhoon, both quartz rich samples, contained very low amounts of kaolinite (0.8–1 wt.%) compared with Gilmore and Gneiss (7.7–10.6 wt.%). Kaolinite exhibited a striking decrease in d 001 peak intensity at 12.2° 2Ѳ compared with that expected from kaolinite (Fig. 2). This preferential decrease in d 001 peak intensity compared with d 02l peak intensity (20° 2Ѳ) is a characteristic of acid attack (Pentrák et al., 2012). XRD analyses of oriented mounts of the <2-μm size fraction of each sample (not reported) confirmed the presence of kaolinite in each sample and also revealed the presence of a trace 10 Å phyllosilicate that could be either illite or a mica mineral.
Plagioclase is identified in most samples (1.7–6.6 wt.%) as well as alunite, which is most abundant (49.5 wt.%) in the sample from the most acidic lake Gneiss (pH = 1, Table 1). Gilmore is the only sample containing gypsum with a significant abundance of 29 wt.%. Hematite and goethite were identified as minor phases in Aerodrome and Gneiss with 0.6 and 1.8 wt.%, respectively. It is important to note, however, that mineral suites in the Western Australian acid saline lakes and adjacent mudflats/sandflats are heterogeneous over meter-scales and change in mineral relative abundances in response to flooding, evapoconcentration, desiccation, and host rock composition (Benison et al., 2007; Story et al., 2010; Bowen et al., 2012).
4. Lipid Results
Typical GC-MS chromatograms show the most abundant lipids recovered from the Bligh and Dyer-extracted nonpolar fractions (F1 and F2) and polar (F3, F4, and F5) fractions. Representative chromatograms are presented in Figs. 3–5. Identifications are noted in Table 2. The following internal standards were used: heneicosane (C21), 2-methyl-octadecanoic acid, 1-nonadecanol (C19), and p-terphenyl-d14.
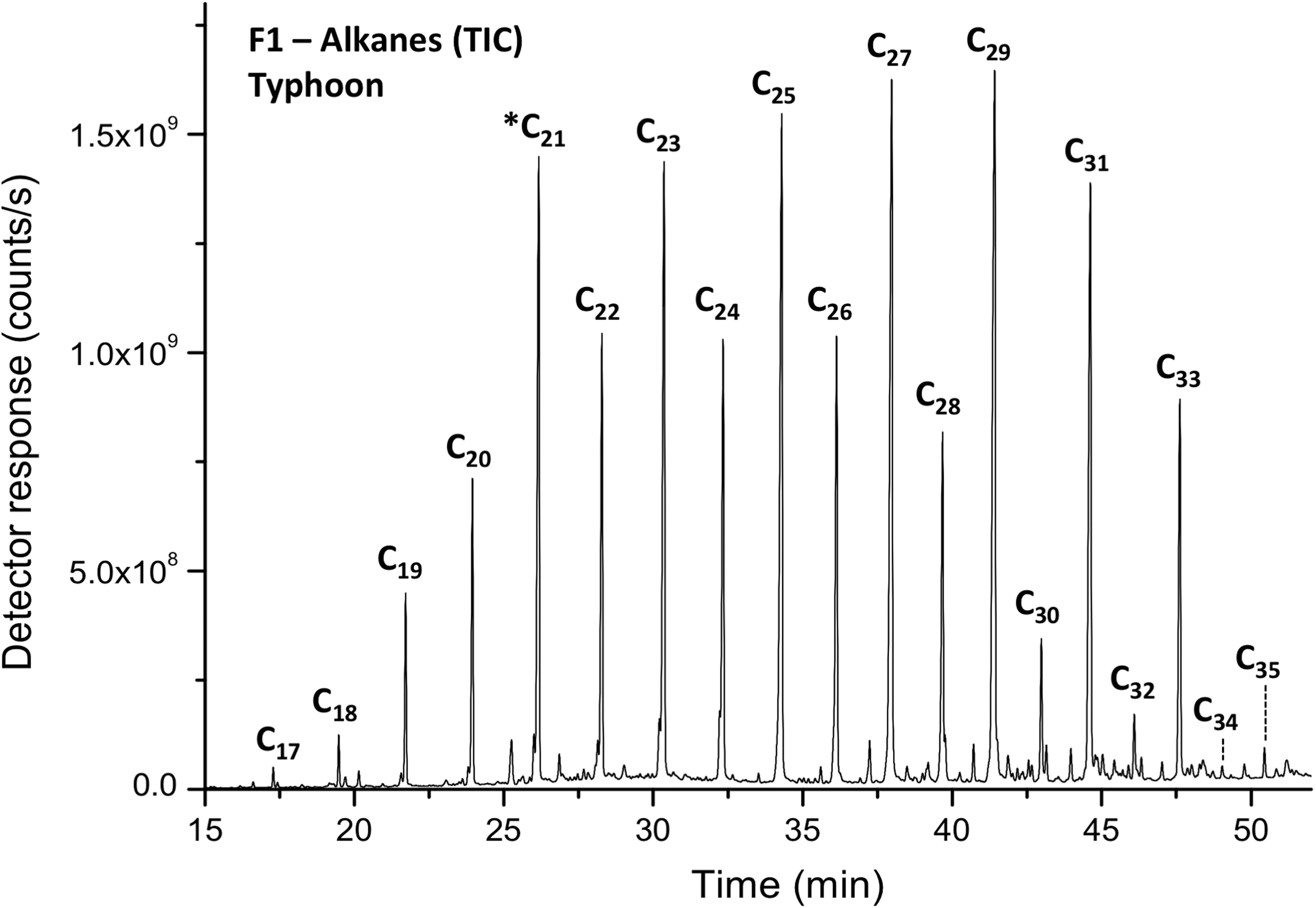
A representative GC-MS chromatogram showing the detection of an odd-over-even carbon number preference in C17–C35 alkanes in Typhoon Lake, a pattern indicative of terriginous plant material (Nonpolar Fraction F1). *Heneicosane (C21) was added as an internal standard. GC-MS, gas chromatography-mass spectrometry; TIC, total ion current.
n-Alkanes were identified in fractions F1 and F2 after Bligh–Dyer extractions and column separation. The n-alkanes range in carbon number from 17 to 35, whereas the most dominant homologues are the C25–C31 n-alkanes; no evidence of cyanobacteria in short chain n-alkanes was seen. There is a strong odd over even carbon number predominance (Fig. 3).
Fatty acids in the polar fractions range in carbon number from 12 to 32 and n-alcohols range from 14 to 32. Even numbered carbon compounds dominate the distributions of fatty acids and n-alcohols (Fig. 4). Only very low levels of branched fatty acids were detected (C15:1, C16:1, and C18:1, all <1% of the total fatty acid distribution). Sterols, ursolic acid derivatives, and betulin were also found in high levels in the polar fractions (Fig. 5). Based on retention times and fragmentation patterns, we also uncovered likely pentacyclic triterpenoids in some of the sampled sites, possibly bacterial hopanes or oleanane-type compounds.

A representative GC-MS chromatogram showing the detection of predominantly even carbon number fatty acids, as well as ursolic acid isomers and betulin, in Typhoon Lake (Polar Fraction F4). The shorter chain (i.e., C16 and C18) fatty acids shown are likely microbial in origin.
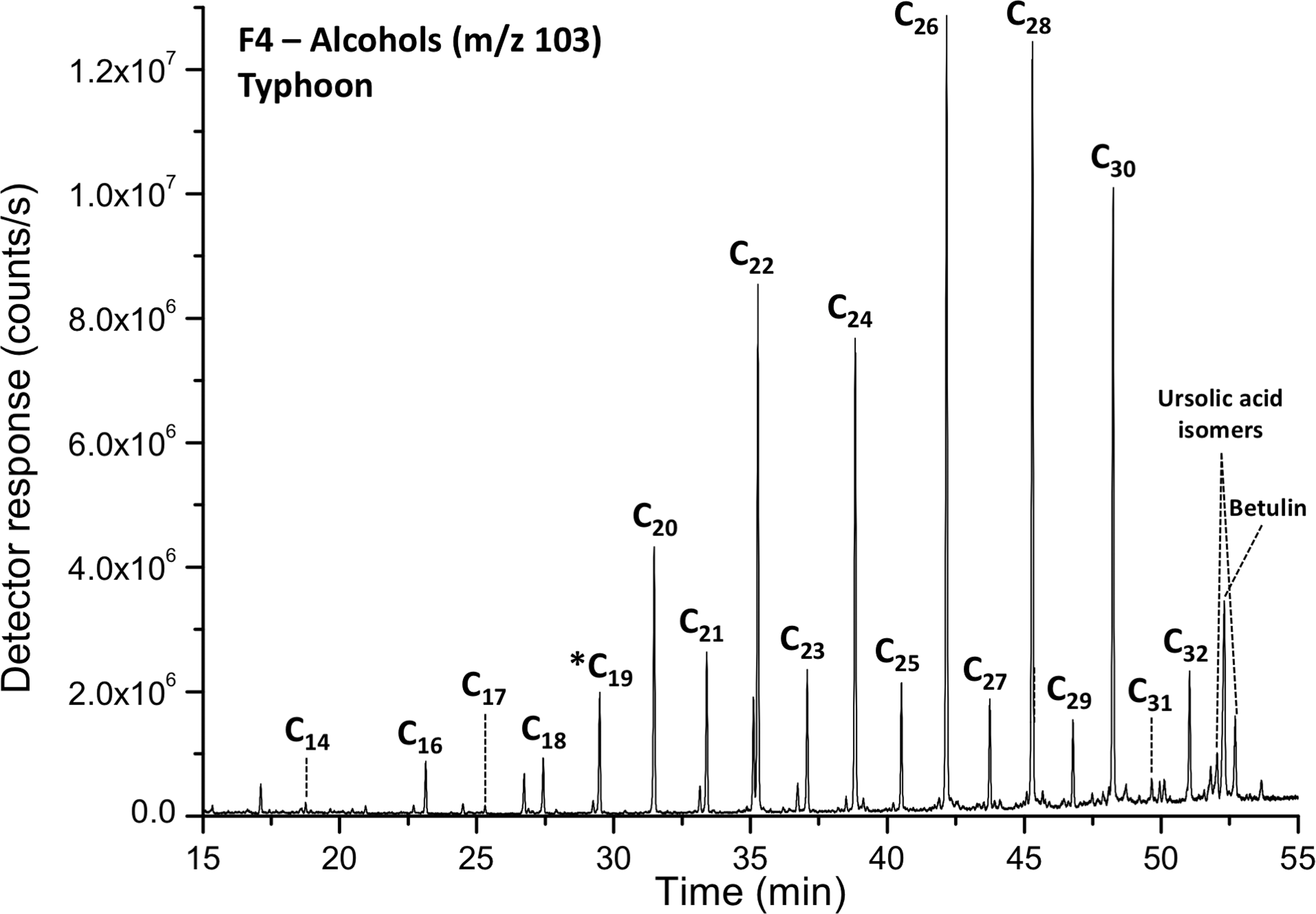
A representative GC-MS chromatogram showing a predominantly even carbon number alcohols, as well as ursolic acid isomers and betulin, in Typhoon Lake (Polar Fraction F4). *1-Nonadecanol (C19) was used as an internal standard.
5. Discussion
On Earth, organics are often sequestered in tandem with phyllosilicate minerals, such as smectites and associated noncrystalline iron and aluminum oxides/hydroxides (Wattel-Koekkoek et al., 2003). Micrometer-size phyllosilicate minerals dominate the organic matter stabilization literature, in large part because these minerals have relatively large surfaces areas and typically carry a substantial charge, allowing them to chemically stabilize organic matter (Keil and Mayer, 2014). These observations suggest that terrain-dominated phyllosilicates are the best targets for lipid biomarker recovery; thus, sites with analogous phyllosilicate deposits on Mars, such as Jezero Crater, have been targeted for future exploration (Ehlmann et al., 2013). The question remains as to whether phyllosilicates found in association with other mineral groups, such as the sulfate and iron oxide-bearing phyllosilicates of the Upper Murray unit at Gale crater (Rampe et al., 2017), would also be worth targeting for sample return.
Our study is among the first to investigate the biomarker potential of mixtures of sulfates, iron oxides, and phyllosilicates in a highly acidic terrestrial Mars analogue. We found a strong odd over even carbon number predominance in long-chain n-alkanes (carbon preference index, or CPI, ranged from 3.6 to 5.7), indicating a terrigenous origin, probably in the primary form of cuticular waxes (Eglinton and Hamilton, 1967). These have also been shown to be carried in eolian dust (Simoneit, 1977; Gagosian et al., 1987).
We also found a predominance of even numbered carbon compounds in the distributions of fatty acids and odd numbered compounds in the n-alcohols. Although even-over-odd carbon chain length preferences occur in short-chain fatty acids derived from microbial cellular processes (Volkman, 2006), even-over-odd long-chain fatty acids (>C20) are typically indicative of waxes and terrigenous plant inputs (Eglinton and Hamilton, 1967; Williams et al., 2019). Although an algal origin of the long-chain fatty acids does remain a possibility, other signs of algae, such as higher levels of unsaturated fatty acids, would be expected (Volkman et al., 1998). The predominance of C29 sterols over C27 and C28 sterols again suggests a land plant source (Huang and Meinscheim, 1979), and ursolic acid derivatives and betulin are indicative of higher plant leaf waxes (Jäger et al., 2007).
The overall assemblage of saturated alkanes, alcohols, fatty acids, and sterols we recovered strongly point to the overprinting of plant debris, likely originating from the low-diversity eucalypt forests that exist on the Yilgarn Craton (Yates et al., 2000), though low abundances of microbial lipids were also detected. We surmise that the amount of plant biomass simply overshadowed the traces of microbial biomass in the sediment, as diverse microbial communities have been observed within lake water and mud/sandflat sediment as part of genomic studies (Mormile et al., 2009; Johnson et al., 2015; Zaikova et al., 2018). It is possible that there is a preservational bias for plant material in this system (perhaps sequestered in such a way that is more conducive to preservation) or simply for larger molecules (plant terpenoids, potential hopanes); however, easily hydrolyzed shorter chain fatty acids (e.g., C16, and C18, likely associated with microbial communities) were also observed. It is interesting to note that although similar trends were found in terms of biomarker recovery as part of the 2005 pilot study (Johnson, 2008), there were noticeably fewer short-chain fatty acids recovered in this 2018 investigation. This may be due to seasonal fluctuation in the active microbial community, as the 2018 samples were collected in the Austral winter, whereas the 2005 samples were collected in the Austral summer when evapoconcentration and desiccation are most likely to occur. Although microbes may be present as part of an active subsurface microbial community, the plant lipids (very long chain alkanes, plant terpenoids, etc.) are certainly from dead organisms—washed into the lake basins at some point in the past. This suggests that structural biomarkers not only associated with extant biology are preserved through at least the early stages of diagenesis in the biosedimentary system represented by these lakes (Table 2).
Interestingly, we observed the highest levels of organic carbon associated with sulfate-bearing samples (Table 1). Substantial quantities of alunite (8.4 wt.%) and gypsum (28.9 wt.%) were found in the crystalline phase of our XRD data in the sediment of Lake Gilmore, which had the highest organic carbon content. The sediment of Lake Gneiss, which had the second highest organic carbon content, also had a significant sulfate-bearing component, with 49.5 wt.% alunite in the crystalline phase of our XRD data. These sulfate-rich lakes had the largest pH range difference among the lakes that we sampled (2.6–5.8 for Lake Gilmore, 1.4–3.7 for Lake Gneiss), suggesting that pH alone may not be a straightforward indicator of preservation. Some level of microbial activity may also be associated with the precipitation of acid salt lake sulfates (Johnson et al., 2015), even though our results strongly suggest that plants are the dominant source of the organics recovered. We found the lowest organic carbon content, as well as the only below-detection levels of nitrogen, in Lake Aerodrome, where the sediments bore no sulfate-bearing minerals and were instead dominated by quartz.
Although there have been a few systematic investigations of the preservation potential of sulfate minerals on Earth, organics—including amino acids and their amine degradation products—have also been detected in Mars analog sulfate mineral matrices, including gypsum from the Anza-Borrego Desert, California, gypsum from the Haughton impact crater, Canada, and gypsum, anhydrite, and jarosite samples from Panoche Valley, California (Aubrey et al., 2006), as well as ferric sulfate-rich streams in St. Oswald's Bay (Tan et al., 2018) and acid saline lake gypsum from northern Chile (Benison and Karmanocky, 2014). Magnesium sulfates have also been shown to protect organics trapped inside the crystal lattice or present in fluid inclusions from oxidation induced by the decomposition of perchlorates, powerful oxidants present on the martian surface (François et al., 2016).
Our results build on other recent findings that suggest that organics could be found in mineralogically diverse sediments on Mars, including those with jarosite group minerals (alunite, jarosite). It is possible that the presence of acid sulfate minerals alongside smectites and other phyllosilicates gave rise to habitable environments where microbial activity was concentrated at redox interfaces, in the past or even today in the planet's subsurface. For example, a recent study of mudstones and sandstones in the Painted Desert of Arizona also revealed far higher concentrations of amino acids, in addition to carboxylic and dicarboxylic acids, alcohols, aliphatics, and other complex molecules, in the presence of clay minerals and jarosite together compared with the presence of clay minerals alone (Noe Dobrea et al., 2016).
The processes affecting the preservation of organics in later stages of diagenesis, however, require additional investigation, and we have only investigated the very earliest stages of diagenesis as part of this study. There are very few examples of these types of depositional systems on Earth, but the ancient acid ephemeral saline lakes of the Permian Opeche Shale of the Williston Basin, North Dakota, provide one such opportunity (Benison et al., 1998; Benison and Goldstein, 2002). Here, the oxidation of sulfide minerals along with other acidification processes, which may have been mediated by a combination of climate, rock-water interactions, and microbes, gave rise to extremely acidic waters (pH <1) in tectonically stable, closed-drainage basins (Benison and Bowen, 2015). These ancient sites precipitated evaporite minerals in an arid climate characterized by abundant, carbonate-poor red siliciclastics, much like lithified strata that have been observed on Mars at Gale crater. The discovery of lipid biomarkers in places such as the Permian Opeche Shale would further bolster the search for remnant organic matter on Mars.
Cosmic rays, UV irradiation, peroxides, and a multitude of nonaqueous chemical oxidations over very long timescales are critical challenges to the recovery of martian biomarkers (Summons et al., 2008), and it is likely that deep subsurface samples from ancient acid saline playa lakes would be better targets for biomarker preservation on Mars than the shallow subsurface samples we analyzed here.
Although hyperaridity is a major challenge to microbial survival on Earth, the hyperaridity of the martian surface, which is 100–1000 times drier than the driest regions on Earth, may facilitate the preservation of lipids generated during environmentally favorable periods (Wilhelm et al., 2017). There is extensive evidence that aqueous conditions were widespread on Mars in its ancient past; so if microbial life did arise on Mars before the desertification of the planet, biomarkers may still be preserved in the geological record.
6. Conclusion
Our investigation of the types of biomarkers recoverable within the naturally occurring acid salt lakes in the vicinity of Norseman, Western Australia—a terrestrial analogue with striking mineralogical and sedimentary similarities to Mars—has revealed the vestiges of several plant biomolecules alongside trace microbial lipids. The resilience of lipids from dead organic material in the saline sediments represented by these lakes lends support to the idea that sulfates, in tandem with phyllosilicates and iron oxides, may be a viable target for preserved biomarkers on Mars, and it motivates investigations of organic preservation in acidic saline pans that have been preserved in our planet's deep geological record.
Footnotes
Acknowledgments
The authors would like to thank Angus Turner for assistance in their initial survey of the area and Roger Summons for insightful advice on the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors would like to acknowledge the support of the William F. Milton Fund and a Georgetown University Main Campus Research Grant.