
Abstract
Assessing biosignature preservation potential (BPP) in ancient habitable environments on Mars is a top NASA priority. We address this goal through the study of Miocene-Pliocene evaporites of the Verde Formation (central Arizona). We assessed the effects of diagenesis on BPP, integrating outcrop-scale observations with six lab analyses: thin-section petrography, X-ray diffraction, Raman spectroscopy, total organic carbon (TOC), electron probe microanalysis (EPMA), and visible to near-infrared (VNIR) reflectance spectroscopy. We recognized five facies and their diagenetic pathways. Two facies included mudstones which contain clusters of displacive growth gypsum (DGG). Early DGG was altered during diagenesis by dissolution forming crystal cavities and later underwent recrystallization, cation substitution, and sulfate dehydration. Another facies was identified by lenticular beds dominated by halite and late diagenetic thenardite (Na2SO4). These pods are overlain by a sequence of interbedded gray and red mudstones which record cyclic oxidation and Fe-oxide cementation. During the Pleistocene, a lacustrine environment developed, accompanied by magnesite cementation of playa mudstones. TOC analyses were used as a proxy for inferring the BPP in each facies. The highest BPP was associated with both red and gray mudstone facies. This study provides a taphonomic framework for playa environments on Earth that record the impacts of diagenesis on BPP, with potential applications to Mars sample return (MSR) missions.
1. Introduction
1.1. Relevance to NASA's priorities
A
1.2. Biosignature preservation potential in evaporite environments
Evaporite deposits are soluble salts that precipitate from brines as a result of solar-driven evaporation (Warren, 1999). They can be deposited in a variety of environments and forms. During bottom-nucleated growth, evaporite crystals nucleate on the bottom of brine pools and grow upward into a water column (Warren, 1999). In displacive growth, crystal clusters (“rosettes”) precipitate from brine-saturated pore waters and nucleate within host sediments (e.g., mud or sand; Warren, 1999; Fig. 2D, this study). Both types may capture and preserve surrounding sediments, including organic matter (OM), within and/or between crystals during growth.
Complex postdepositional diagenetic processes can enhance, or degrade, the BPP within evaporites, whether on Earth or Mars. The subdiscipline of paleobiology that studies the processes that control the capture, preservation, alteration, and retention of biosignatures is known as taphonomy (Efremov, 1940; Behrensmeyer and Kidwell, 1985; Allison and Briggs, 1991; Allison and Pye, 1994). Many factors affect taphonomic processes in evaporites, such as sediment phase solubility, porosity, and permeability, but they are poorly understood. For example, the high BPP of sulfate evaporites was only recently recognized when Schopf et al. (2012) presented the first account of widespread biosignature and microfossil preservation in bottom-nucleated primary gypsum.
In evaporitic environments, several factors can enhance biosignature preservation, even though it occurs under a narrow range of conditions that exist after deposition (Allison and Briggs, 1991; Allison and Pye, 1994). Burial of OM in fine-grained, clay-rich sediments enhances preservation, particularly as porosity and permeability are progressively reduced following burial, compaction, and cementation. Early cementation of fine-grained sediments is important for opening a taphonomic window for OM stabilization and preservation (Farmer, 1995; Farmer and Des Marais, 1999; Westall and Cavalazzi, 2011). Rapid sedimentation of organic-rich clays can create a closed chemical environment that stabilizes OM, promoting preservation (Allison and Pye, 1994). Preservation is also promoted when expansible clays, especially smectites (Kennedy et al., 2002), bind organic molecules within interlayer sites and/or on charged mineral surfaces (Farmer and Des Marais, 1999, and references therein). In playa environments, shorter exposure to oxidation in a hypersaline water column and contact with density-stratified brines can increase bottom-water anoxia and promote preservation (Powell, 1986; Warren, 1986; Allison and Briggs, 1991; Farmer and Des Marais, 1999; Westall and Cavalazzi, 2011).
On the other hand, stratification may decrease BPP by prolonging OM exposure to oxidation before it reaches an anoxic bottom (Katz et al., 1987). Other unfavorable conditions for biosignature preservation in playa settings include rapid deposition of sulfates, which can dilute the concentration of organics relative to inorganically formed minerals in surrounding mudstones (Warren, 1986; Katz et al., 1987). High concentrations of sulfates may also promote oxidation and degrade labile organics before their deposition (Katz et al., 1987).
Once buried, only a fraction of the OM produced by life survives diagenesis and enters the sedimentary record. An estimated 80% of what remains is kerogen (Allison and Briggs, 1991), a complex, macromolecular assemblage of degraded, thermally mature, amorphous, insoluble aromatic carbon. In this study, we focus on the preservation of OM as kerogen. The preservation of kerogen, or any potential biosignature, depends on the nature and degree of postburial alteration during diagenesis. Postburial diagenetic processes include compaction, cementation, dissolution, and replacement through time (Cady, 2001; Westall, 2008). Additionally, recrystallization may decrease preservation by causing loss of primary microfabrics that protect buried OM against oxidation (Farmer, 1999; Allwood et al., 2009).
1.3. Evaporites on Mars
Among the most promising targets in the search for a martian fossil record are evaporite minerals that have been repeatedly confirmed by both orbital and in situ observations. Surface missions have confirmed the presence of Ca sulfates on Mars with varying hydration states (Nachon et al., 2014; Vaniman et al., 2014; Rapin et al., 2016) in shallow playa-like settings (Glotch and Rogers, 2007), and as veins cutting sedimentary rocks (Squyres et al., 2012; Grotzinger et al., 2014; Rapin et al., 2016). In situ analyses have shown potential microtextural evidence of “hopper crystals” of halite (Squyres et al., 2006) and dissolution cavities of displacive growth sulfates (McLennan et al., 2005) as well as other soluble minerals (Herkenhoff et al., 2004; Clark et al., 2005).
The Mars Science Laboratory rover is currently exploring fluvio-lacustrine deposits within the Peace Vallis alluvial fan system in Gale Crater, which may have sustained ancient habitable environments with a high BPP (Grotzinger et al., 2014). Past habitable conditions are also suggested by the presence of smectite clays and sulfate-rich sediments in the Sheepbed mudstone (Grotzinger et al., 2014; Vaniman et al., 2014). Ongoing exploration of Mt. Sharp's layered sulfate sequences may provide further insight into ancient habitable environments in Gale Crater as the Mars Science Laboratory mission continues.
1.4. Motivations for this study
On Mars, understanding general taphonomic processes within early Mars-like environments is considered a critical step in refining the search strategy for biosignatures and selecting samples for Mars sample return (MSR). Two upcoming missions, NASA's Mars 2020 and European Space Agency's ExoMars rovers, will usher in a new era in Mars exploration: the primary mission of both rovers will be to search for potential biosignatures. Additionally, Mars 2020 will cache samples based on their BPP for later MSR to Earth, where detailed assessment of their biogenicity will be carried out in terrestrial labs. MSR presumes a rover payload that will (1) target deposits recognized to record past habitable environments, (2) identify samples with a high BPP, and (3) prioritize the best samples for in situ analysis and/or caching for return to Earth.
Here, we aim to better understand the diagenetic factors that may impact BPP over geological time frames. This may help refine a biosignature preservation model for a Mars-relevant environment on Earth—a playa evaporite quarry in the Verde Formation (central Arizona)—in order to better inform sample selection for MSR. The goal of this study is to address two questions: (1) What postdepositional and diagenetic processes affected playa evaporites within the Verde Formation? (2) How did those processes affect BPP within each evaporite facies studied?
Previous work examined large-scale, basin-wide diagenetic processes within Verde Formation (Wadell, 1972; Donchin, 1983). However, this is the first work to present an integrated view of the diagenetic history of the Verde Formation playa evaporites and how that history affected BPP.
1.5. Geological context of the study site
This study focuses on a 50 m playa evaporite sequence exposed at the Old Indian Salt Mine quarry (see Fig. 1) within the Verde Formation. The quarry is located along the southern edge of the Verde Basin near Camp Verde, Arizona, within the Transition Zone that separates the Great Basin and the Colorado Plateau Structural Province (Nations et al., 1981; DeWitt et al., 2008). The age of the formation is poorly constrained as late Miocene to early Pliocene.
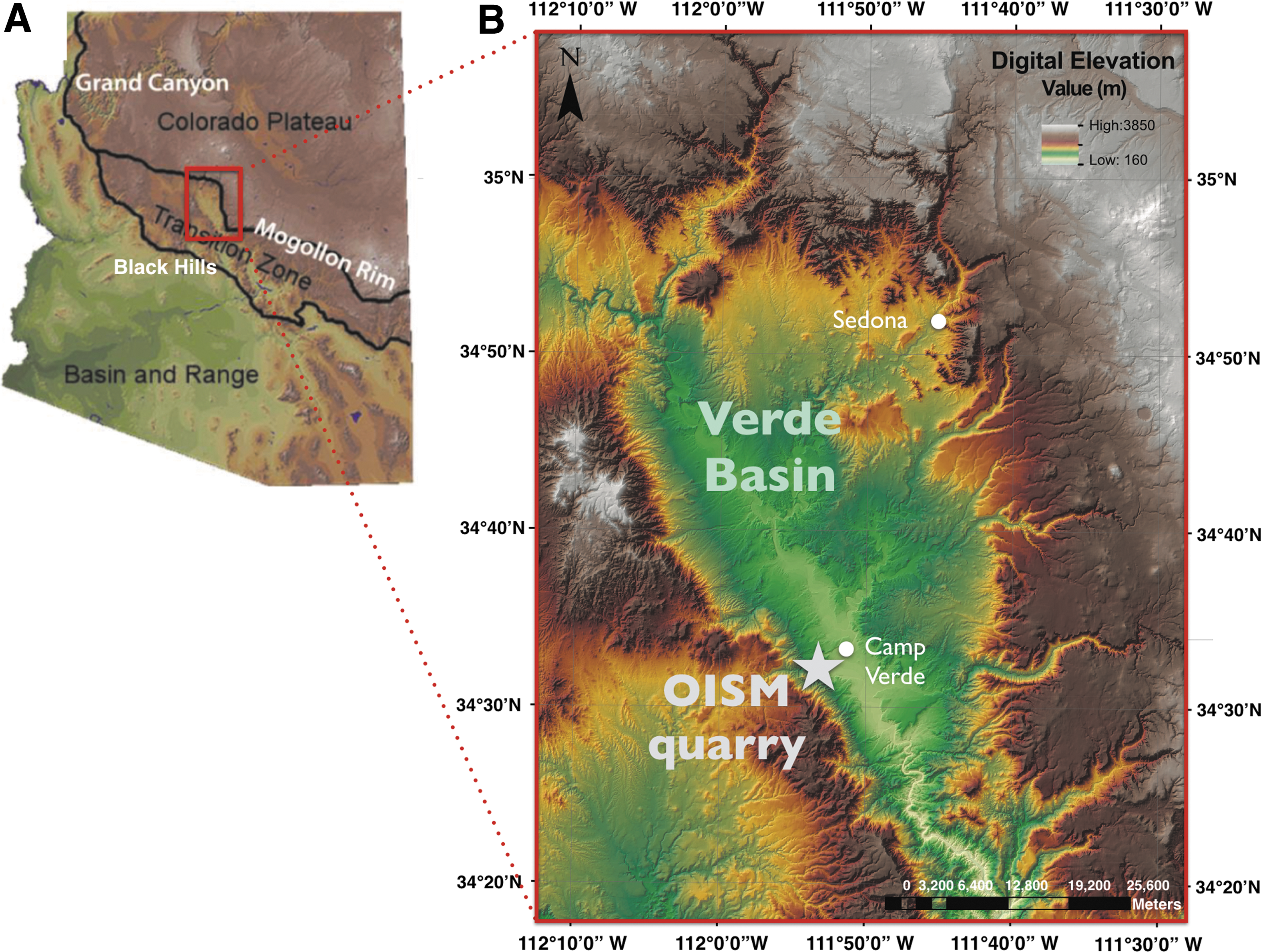
Topographic map of the Verde Basin and OISM quarry within the Transition Zone of Central Arizona. (
Basinwide, the Verde Formation sediments were deposited in three main depositional environments: (1) lacustrine, (2) marginal-lacustrine, and (3) alluvial fan (DeWitt et al., 2008). Following evaporite deposition, a perennial lake developed, depositing basin-wide lacustrine carbonate sediments (Wadell, 1972). Volcanic damming of the Verde River, along with climatic and tectonic events, was accompanied by cyclical changes in lake level (Donchin, 1983). Figure 1 illustrates the regional location of the study site, as well as the specific location of the Old Indian Salt Mine (OISM) quarry.
The Verde Formation playa evaporites are dominated by mudstones rich in magnesite (MgCO3), along with sulfates such as gypsum (CaSO4·2H2O) and thenardite (Na2SO4), and halite (NaCl). Based on this mineral assemblage, a starting composition of Mg–Na–Ca–SO4–Cl brines may be inferred. Section 4.1 details this brine evolution model, reconstructing the mineral evolution and diagenetic processes in OISM playa evaporites that are based on it.
2. Methods
After field analysis and sample collection, six analytical methods were used to analyze samples, including (1) microscopy to interpret mineralogy and place it within a microtextural context, to describe diagenetic processes, and to reconstruct a paragenesis (time-ordered sequence of environmental processes); (2) X-ray powder diffraction (XRPD), to obtain bulk mineralogy; (3) Raman spectroscopy, to identify mineral phases and kerogen; (4) electron probe microanalysis (EPMA), to identify elemental compositions of kerogen and minerals; (5) visible to near-infrared (VNIR) reflectance spectroscopy, to identify Fe-bearing phases; and (6) total organic carbon (TOC) analyses, to obtain bulk percentages of preserved organic carbon for each facies as a proxy for BPP. Each technique is detailed below.
2.1. Field sampling
The main quarry is located at 34°32.757'–34°32.701'N and 111°52.541'–111°52.620'W. Figure 2 shows the OISM outcrop investigated, and the sampling locations and facies. Fieldwork was carried out from 2014 to 2016. Table 1 provides a summary of collection details.
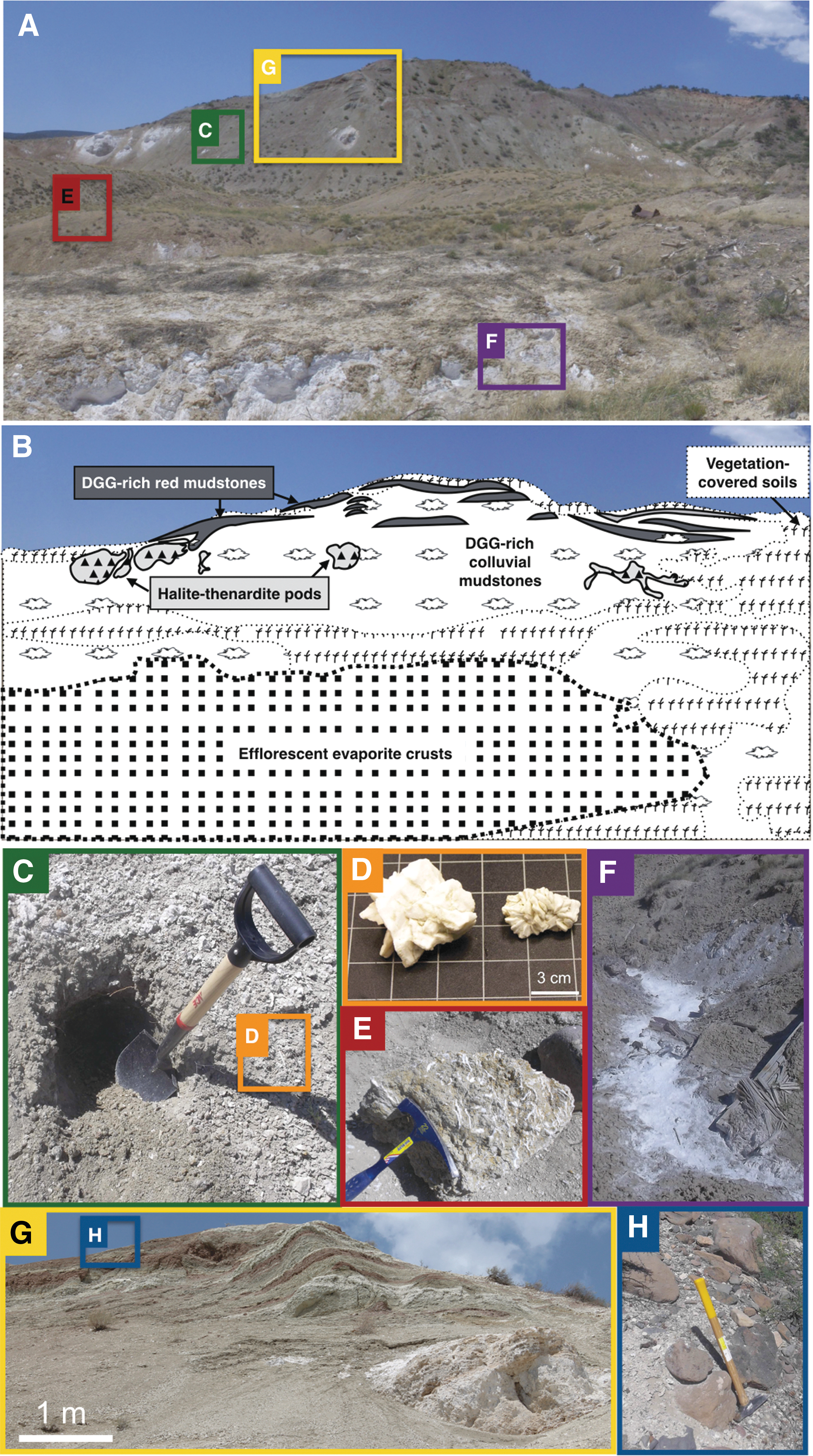
Sample locations within the OISM outcrop. (
The column on the far right denotes where representative field images are located in Fig. 2. NP = not pictured.
2.2. Petrographic microscopy
For spatially integrated data sets using microscopy and Raman spectroscopy analyses, petrographic thin sections were prepared from representative samples of each facies. Standard thin sections were prepared by Spectrum Petrographics from cut slabs or strew mounts in nonfluorescent epoxy.
Thin sections were analyzed with a Nikon Eclipse E600 POL polarizing microscope in plane, cross-polarized, and reflected light. Mineral components, microtextures, and cross-cutting spatial relationships were documented.
Initial identification of minerals and kerogen involved examining mineral and kerogen targets in transmitted and reflected light. Raman spectroscopy was used to interrogate grains of interest for identifications. Sticker markers were adhered around representative target kerogen and mineral grains during microscopy analysis for relocation of target regions for Raman spot analyses.
2.3. Raman spectroscopy
A continuous-wave Raman spectroscopy system was used to obtain point spectra on thin sections. This system uses a 532 nm (visible) Coherent Sapphire SF laser source (Princeton Instruments), a liquid nitrogen-cooled CCD detector, an Acton spectrograph with a 1200 lines/mm grating and an acquisition window of 70–2000 cm−1.
A built-in transmitted light microscope was used for locating targeted kerogen and mineral grains of interest (see above). The laser spot size was ≲5 μm in diameter. Spectra were acquired over a range of integration times (1–30 s), power settings (750 μW to 6 mW), and depths below the surface of thin sections (1–20 μm, manually focused) to maximize signal to noise ratio.
Raman shift positions were calibrated by using cyclohexane. No background correction was applied. Mineral identifications were obtained with CrystalSleuth and RRUFF databases (Downs, 2006; Lafuente et al., 2015).
2.4. X-ray powder diffraction
X-ray powder diffraction (XRPD) analyses were obtained from bulk, powdered samples to provide an independent assessment of mineralogy. (Organics cannot be identified with XRPD.) Bulk samples were ground and sieved to <150 μm powders. A Siemens D5000 X-ray diffractometer was used to identify minerals. Sample powders were scanned from 5° to 82° at 40 kV and 30 mA and by using a CuKα X-ray source. XRD data processing and identification were performed with JADE software V9.
2.5. Electron probe microanalysis
For EPMA, we used standard, doubly polished 1-inch rounds (Spectrum Petrographics, Inc.). Sections were cleaned with compressed air and coated with 0.6 nm of Ir. Elemental analyses were obtained with a JEOL JXA-8530F electron probe microanalyzer. Operating conditions used a 15 keV accelerating voltage, 20 nA beam current, 5 μm beam diameter, 5–10 μm step size, and 50 ms dwell times. X-ray intensities were measured by energy-dispersive spectrometry (EDS). Because C-rich epoxy can infiltrate porous mudstones during thin section preparation and mimic a C-rich kerogen signal, Cl was used as an anti-indicator of kerogen. (Cl is found in epoxy but not in kerogen.)
2.6. Visible to near-infrared reflectance spectroscopy
To identify iron-bearing phases from the Fe-oxide-cemented mudstones, we used VNIR (0.35–2.5 μm) reflectance spectroscopy on hand samples and powders (<150 μm). Spectra were acquired by using an Analytical Spectral Devices (ASD Inc.) FieldSpec 3 spectroradiometer at room temperature and ambient atmospheric conditions. The Muglight option, intended to obtain reflectance and absorbance measurements of granular materials, was used for powders. A goniometer with mounted fiber optic cable and external light source was used for hand samples. Samples analyzed were ∼1 cm in surface area. The Muglight had an internal light source fixed at a 23° incidence angle. The goniometer was set to an incidence angle of 30°. A Spectralon 99% diffuse reflectance standard was used as a white reference. Spectra were processed with ASD ViewSpec software.
2.7. Total organic carbon analysis
Samples from each facies were powdered and sieved to <150 μm. To remove inorganic carbon in carbonate, silver capsule in situ acidification was used, and samples were treated with 1 M HCl. Dissolution continued until no effervescence was observed. Samples were dried in a desiccator between acidification rounds.
Total organic carbon analyses were performed at Arizona State University with a Thermo Delta Plus Advantage isotope ratio mass spectrometer, interfaced with a Costech elemental analyzer. In-house glycine standards as well as the NIST2710 standard were used for calibration and isotopic data normalization. To promote complete combustion, silver capsules were wrapped in tin capsules and combusted at 1020°C.
For each facies, five replicate samples were analyzed. Some replicates showed evidence of evaporite formation on capsule exteriors during acidification. Compromised samples were discarded due to presumed loss of sample mass, resulting in fewer than five replicates analyzed in some facies.
Total organic carbon and carbon isotopes were measured on the same samples to confirm that TOC measurements were derived from organic carbon rather than inorganic sources (which would indicate incomplete carbonate removal).
Uncertainties for both TOC and isotopic measurements, Δ, were determined by using
where RSDinstr is the relative standard deviation between measurements for a glycine standard (instrumental uncertainty) and σ rep is the standard deviation between replicate measurements of a facies.
3. Results
Results for the five facies sampled are described in Sections 3.1–3.4. Table 2 is a summary of mineral phases for each facies. Results from organic analyses are presented in Section 3.5.
Abbreviations are as follows: M = petrographic microscopy; R = Raman spectroscopy; X = XRPD; E = EPMA; and V = VNIR. Lowercase letters indicate weak features.
Limonite was also possible.
Detrital feldspars entrapped within mudstone components included alkali, Na-, K-, and Ca-rich solid solution endmembers.
Indicates a cation-substituted intermediate phase in the same facies.
Nearby postmine minerals (Vance and Condie, 1987), such as kieserite group sulfates, rhomboclase, hexahydrite sulfates, or lausenite (Lausen, 1928), were the best matches for otherwise ambiguous detections by Raman and XRD.
The presence of this uncommon sulfide may be attributed to (1) the presence of sodium chloride and an elevated salinity of ancient brines in the lake and/or (2) regional hydrothermal processes related to the sulfide ore body in nearby Jerome, AZ (Vance and Condie, 1987).
3.1. Gray mudstone facies
Old Indian Salt Mine playa deposits are dominated by gray, clay-rich, calcareous mudstones. Mudstones are slope-forming and structureless, lacking fine layering. The homogeneous nature of the gray mudstones is likely due to haloturbation (mechanical rotation of primary textural features in sediments by cyclic dissolution and reprecipitation of salts or the displacive growth of sulfates within sediments; Warren, 1999).
The OISM calcareous mudstones are predominantly cemented by microcrystalline carbonate (Fig. 3A). While calcite is present, XRPD, EPMA, and Raman confirmed magnesite to be the dominant carbonate in mudstones; this is consistent with results from prior mapping of the evaporite unit (Wadell, 1972).

Gray mudstone facies. (
Mudstone thin sections often contained irregular, angular, fine sand-sized clasts (Fig. 3B), along with detrital grains including micas, feldspars, and quartz (Fig. 3C–3E). Some of these grains, along with selected hematite-rich clasts, displayed orange coatings (Fig. 3B, 3F, 3G) interpreted to be Fe-oxide weathering rinds. Raman analyses failed to identify the composition of rinds due to fluorescence. (The only Fe-bearing phase confirmed by Raman was hematite; Table 2 and Fig. 3G.) XRPD also failed to identify Fe oxides, suggesting that they are poorly ordered and/or below the instrument's detection limit of 5%. VNIR analyses of bulk mudstones (both gray and red facies) confirmed the presence of Fe oxides, including hematite, goethite, and lepidocrocite (Fig. 4) but not their spatial distributions.
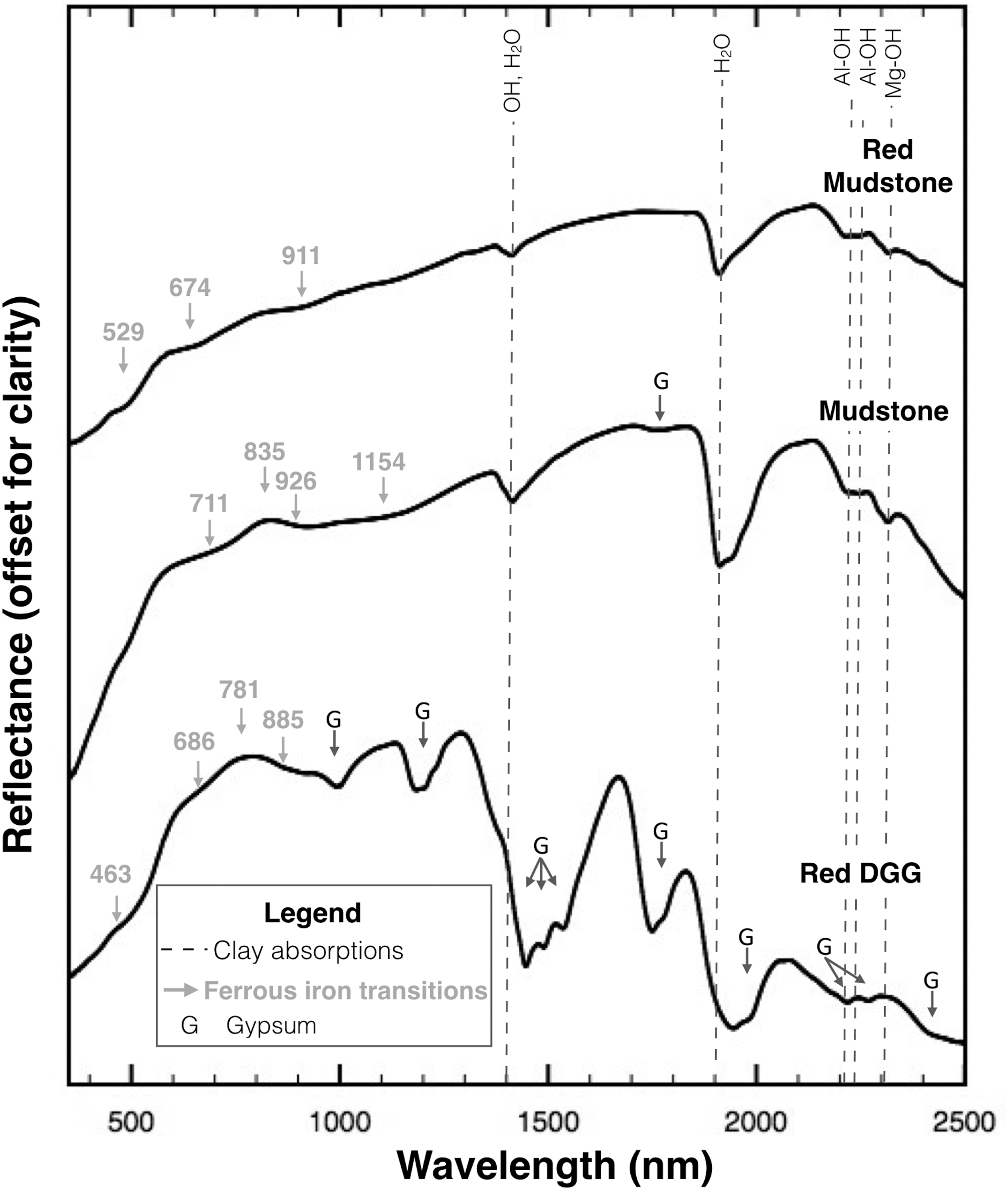
VNIR reflectance spectra. Spectra are shown for powders from a red mudstone sample, #04152015.19, a gray mudstone, #04232014.5, and a hand sample of a red DGG, #04152015.17. Gray arrows indicate the absorption edge, shoulder, reflectivity maximum, or band minimum positions of Fe electronic transition features attributed to mixtures of hematite, goethite, lepidocrocite, or similar Fe-oxide phases (Morris et al., 1985; Cornell and Schwertmann, 2003). Dashed lines indicate montmorillonite and saponite clays at 1410 nm (OH, H2O), 1920 nm (H2O), 2210 nm (Al-OH), 2240 nm (Al-OH), and 2310 nm (Mg-OH; Clark et al., 1990; Bishop et al., 2008). The weak 1154 nm feature is attributed to ferrous iron in saponite. G indicates the positions of gypsum features (Clark et al., 1990). Spectra are stacked and offset for clarity.
Phyllosilicates, particularly smectites, were also abundant in the mudstones. Using XRPD and VNIR analyses, we identified montmorillonite and Fe- and Mg-saponite as major mudstone components (Fig. 4; Table 2), consistent with previous reports of montmorillinoid clays in Verde Formation mudstones by Wadell (1972). Detrital micas (Fig. 3C) were present as minor components.
3.2. Fe-rich mudstone facies
The red mudstone facies is interbedded with the gray mudstone facies at the top of the quarry (Fig. 2G). Red and gray mudstones share similar characteristics. For example, VNIR spectroscopy revealed similar Fe phases present in both gray and red mudstone facies (Table 2).
However, four differences between the mudstone units justify separating them into separate facies. The most obvious difference is that red mudstones exhibit pervasive Fe-oxide cement (Fig. 5A), including hematite and goethite (Fig. 4). Secondly, sand-sized detrital grains are more abundant and coarser-grained in red mudstones, as revealed by EPMA (Fig. 5B, 5C), especially in certain Fe-rich phases like ilmenite (Table 2). Thirdly, sand-sized clasts contained fewer alteration rims in red mudstones, compared to gray mudstone facies. Fourth, EPMA revealed the presence of embayed Fe-oxide grains in red mudstone samples (Fig. 5D).

Red mudstone facies. (
3.3. Displacive growth gypsum facies within gray and red mudstones
Displacive growth gypsum (DGG) rosettes were collected from both gray and red mudstone facies, at several locations (Table 1). DGG crystals were abundant on quarry slopes, where they accumulated by mass-wasting processes and channeling of surface runoff (Fig. 2). All DGG samples analyzed were gypsum-dominated (Table 2; Figs. 4 and 6A). Displacive gypsum growth often incorporated the host sediment (mudstone along with associated OM) into spaces between individual crystals (Fig. 6B). Single crystals entrapped within mudstones commonly displayed lensoidal forms (Fig. 6C, 6D).

DGG facies. (
Patchy coarsening of gypsum was sometimes observed (Fig. 6B), inferred to result from recrystallization. Another recrystallization texture was observed within mudstone cavities (crystal molds): gypsum that infilled precursor cavities was often finer-grained and bladed (Fig. 6E, 6F). Raman spectroscopy analyses on gypsum crystals revealed Na-Ca-Sr sulfates (Fig. 7A, 7B), consistent with Sr-cation substitution (and in a few cases, total Sr replacement to celestine, or SrSO4). Raman spectroscopy analyses also revealed the presence of anhydrite (CaSO4) and bassanite (CaSO4·0.5[H2O]) intermediates (Fig. 7C). This is consistent with previous reports of anhydrite at the study locality (Phillips, 1987).
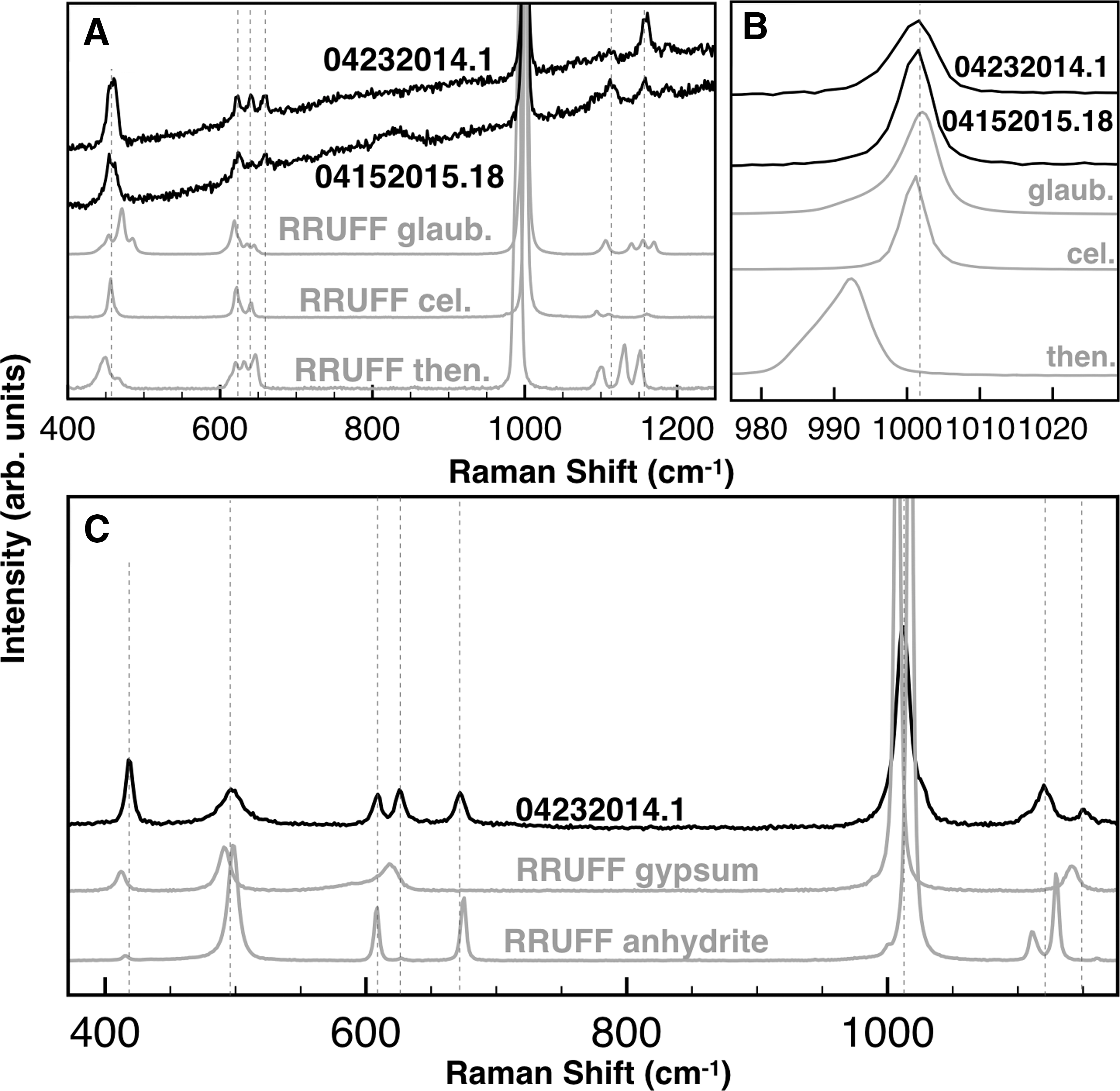
Raman spectroscopy in the red and gray DGG facies samples. (
Comparative analyses of DGG collected from gray and red mudstones revealed them to be similar (Table 2) with one exception. DGG samples from red mudstone units had reddish Fe-oxide coatings acquired from the host mudstones, while those from gray mudstones did not.
3.4. Halite-thenardite pod facies
Another evaporite facies sampled is represented by lenticular beds seen in multiple locations on quarry walls where they crop out as lens-shaped pods within the gray mudstone unit (Fig. 2). Halite is the dominant phase in the pods, indicated by thin section observations of grain textures consisting of white, interlocking cubic crystals (Fig. 8A–8D). These were confirmed to be halite with EPMA (Fig. 8E–8G). EPMA also eliminated concerns that epoxy (which is Cl- and C-rich but Na-poor) had been misidentified as halite (Fig. 8E–8G). Similar to gypsum, halite crystals entrapped mudstone sediments during crystal growth (Fig. 8A).

Halite-thenardite pod facies. (
Thenardite was the second major phase present in pods. Corroded cores of halite were seen within thenardite crystals (Fig. 8C, 8D).
Minor gypsum was also present in pods, interbedded with halite-thenardite and mudstone (Fig. 9A, 9B). In outcrop observations, gypsum crystals were oriented subvertically. The subvertical orientation of gypsum crystals was consistent with bottom-nucleated growth in shallow surface ponds (Fig. 9A, 9B).
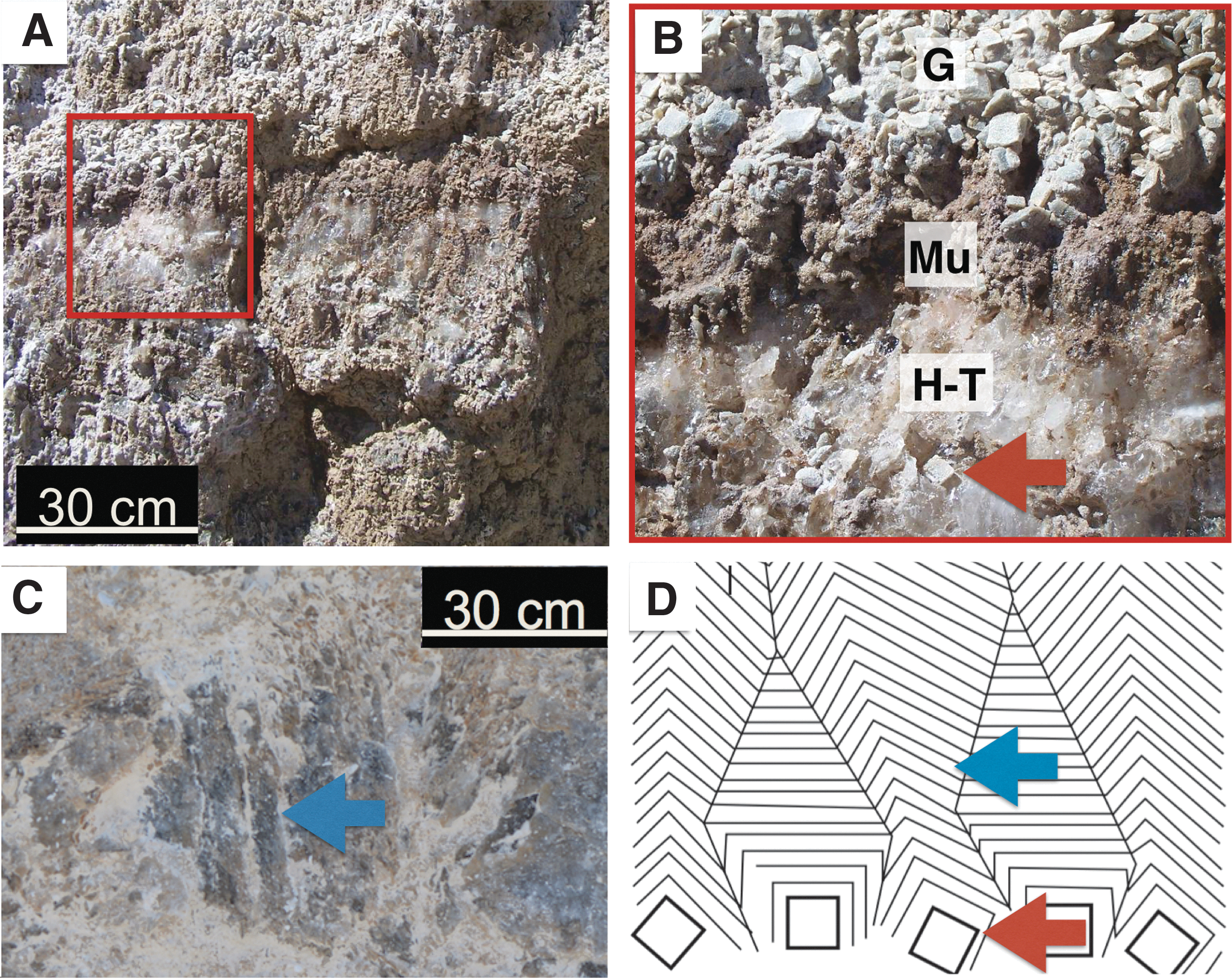
Pod outcrop images. (
Previous literature for the OISM identified glauberite as the dominant phase replacing gypsum rosettes (Twenter and Metzger, 1963; Wadell, 1972; Anthony et al., 1982). However, our analyses only identified glauberite in pod samples as a minor phase (Table 2), present as small crystals entrapped within larger thenardite crystals. This suggests that glauberite is not a common pseudomorph after gypsum in the OISM sequences. Other minor phases detected in pods were similar to those seen in DGG facies (see Table 2).
3.5. Organic analyses
Although the Verde Formation is generally unfossiliferous (Wadell, 1972), particulate kerogen grains (10–100 μm) were abundant in our samples (Fig. 10A). Kerogen was preserved exclusively within mudstone facies (which were common constituents of the other facies present via entrapment). EPMA confirmed that regions suspected to be kerogen were consistent with a kerogen composition (i.e., C-rich and O-poor; Fig. 10). These grains were Cl-poor, indicating that epoxy had not been misidentified as kerogen (Fig. 10).
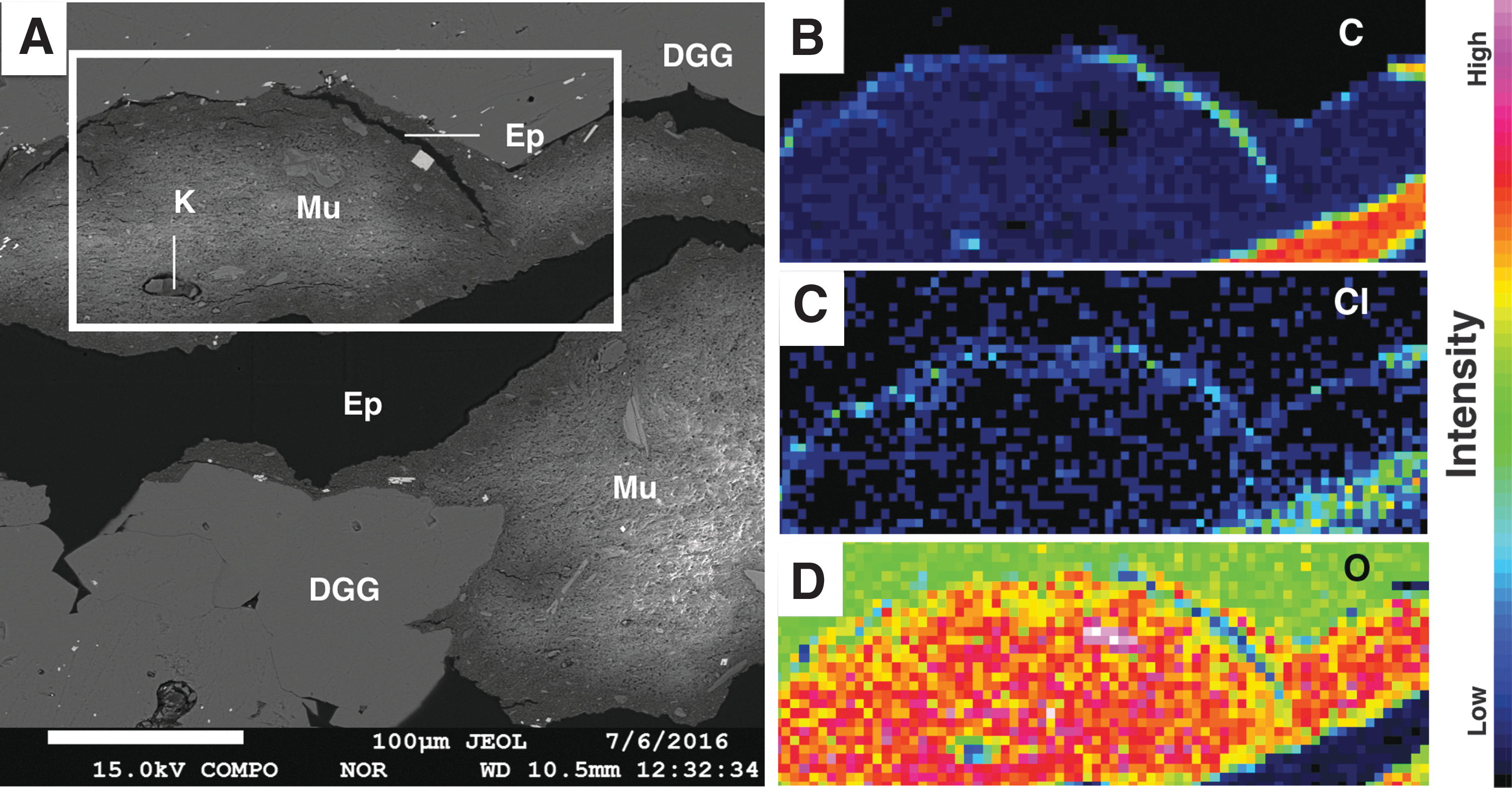
Kerogen characterizations. (
Raman spectroscopy of the mudstone failed to confirm the presence of suspected kerogen grains due to interfering fluorescence. Therefore, we relied on bulk sample methods (which are not spatially resolved) to obtain TOC abundances for each facies.
Total organic carbon results clustered mostly into two groups, as shown in Fig. 11. Values are summarized in Table 3. Both red and gray mudstone facies showed the highest TOC values, while both DGG facies and halite-thenardite pod facies showed slightly lower TOC values. Both mudstone facies showed similar TOC values (Fig. 11; Table 3). These values are consistent with TOC values (<1 wt %) reported for near-surface, bioturbated evaporitic carbonate muds (Warren, 1986). The average TOC values from both DGG facies and the halite-thenardite pod samples fell into the lower TOC group (Fig. 11; Table 3). TOC values for both DGG facies groups are consistent with those of Barbieri et al. (2006), who obtained TOC values of <0.1 wt % using similar methods for the organic-rich laminations of a lacustrine gypsum deposit in Tunisia. Halite-thenardite pod samples and gypsum samples revealed similar TOC values (Fig. 11; Table 3).
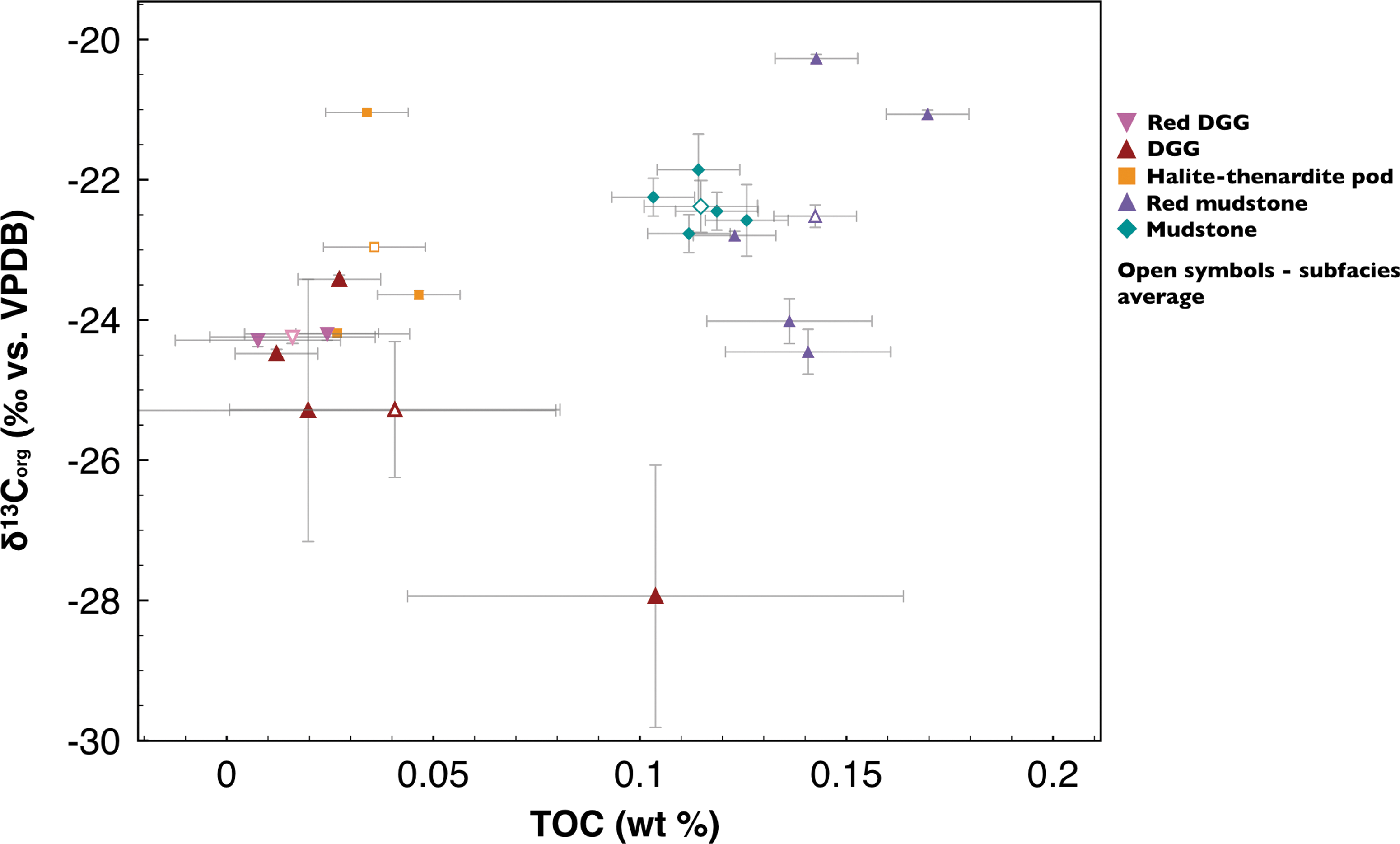
δ13Corg versus TOC measurements with uncertainties plotted for each subfacies (colored symbols), with facies averages shown as open symbols. For error bar calculations, see Methods. The outlier in the DGG facies was one of the sample replicates of sample 04152015.14 (Table 1). It is attributed to acid-organic or acid-sulfate interactions in the silver capsule during acidification.
Isotopes are per mil variation from the PDB standards.
For calculations, see Methods.
Values for δ13Corg ranged from −27.94 to −21.04‰ (VPDB) for all samples (Fig. 11), with average δ13Corg values ranging from −25.28 to −22.38‰ (Table 3). Although the exact input sources of the Corg were undetermined, these results are consistent with a biogenic origin. They are also consistent with values expected for organisms commonly associated with lacustrine carbonates and terrestrial sedimentary OM. These organisms include plants, such as lacustrine algae, that use the C3 carbon fixation pathway, as well as evaporite sediments rich in aquatic autotrophs in Miocene marls, all having δ13Corg values ranging from −5 to −35‰ (Meyers, 1994; Des Marais, 2001; Schouten et al., 2001).
Our δ13Corg values were not consistent with values expected for inorganic carbonates, further supporting an organic C interpretation. Inorganic carbonate values typically range from +12 to −15‰ for a variety of environments similar to those reported here, including shallow lacustrine basins, hydrologically open and closed Miocene-aged lakes, hypersaline settings, and carbonate-rich playas (Talbot, 1990; Bellanca et al., 1992; Utrilla et al., 1998; Aiello et al., 2001; Melezhik et al., 2001).
4. Diagenetic Interpretations
4.1. Diagenetic pathways for OISM evaporites
To address the first goal of the study, we have reconstructed a paragenetic sequence for the facies identified in the OISM quarry. Figure 12 summarizes the sequence detailed below.
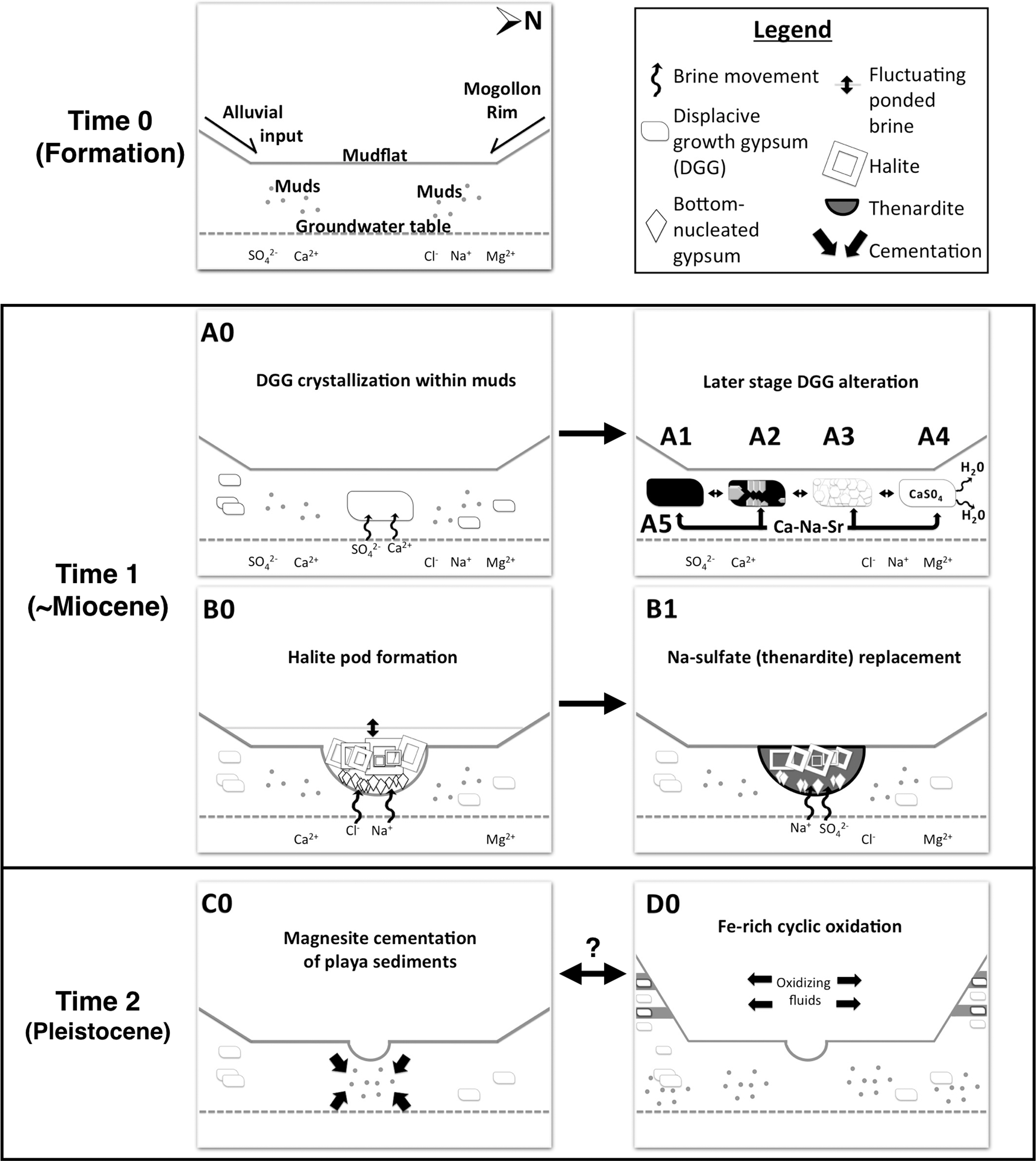
Simplified sequence of diagenetic pathways through time in the OISM quarry. See Discussion for details. The initial depositional environment is depicted at the formation stage (Time 0). Major diagenetic pathways are shown (A0–D0) and arranged through progressing time and diagenesis (arrows). Double-sided arrows imply unconstrained timing. Numbers after letters denote later alteration events associated with those of the same letter. Pathways A0 and B0 may have co-occurred.
4.1.1. Initial depositional context (Time 0 in Fig. 12)
Section 1.5 begins with the early geological history and formation of the Verde Basin, leading up to playa evaporite deposition during the late Miocene. With the onset of playa development, and based on the mineral assemblage seen in the studied samples, we infer a Mg–Na–Ca–SO4–Cl starting brine composition, based on brine evolution models developed by Hardie and Eugster (1970), with modifications by Jankowski and Jacobson (1989).
Mg and SO4 were introduced to the Verde Basin sedimentary system as a result of volcanic weathering (Warren, 1999) and/or from mafic Proterozoic rocks exposed in the Black Hills along the southern margin of the basin. During the late Miocene, to the early Pliocene, crustal extension and volcanic activity resulted in periodic damming of the Verde River, creating a series of smaller sub-basins that formed small ephemeral lakes. Changes in climatic conditions resulted in evaporative conditions that favored the formation of playas, as seen in OISM. This playa system was active during the early Pliocene when evaporite sedimentation alternated with alluvial fan sedimentation. The alluvial fans prograded northward from the southern basin margin, delivering muds to the playa surface. The playa sequence culminated with deposition of Plio-Pliestocene alluvial gravels near the top of the playa section.
Abundant clays were observed within mudstones. However, XRPD could not be used to conclusively confirm their origin (i.e., detrital vs. authigenic), a dilemma also relevant to the interpretation of clays found in mudstones on Mars (Bristow et al., 2015). A predominantly detrital origin can be inferred for the poorly sorted, angular detrital grains and unweathered volcanics observed in studied mudstones, which likely experienced limited transport or alteration following deposition.
4.1.2. Pathway A0, DGG crystallization within playa muds (Fig. 12 Time 1)
Gypsum precipitated displacively within playa muds (Wadell, 1972). DGG was formed by capillary-driven evaporation of CaSO4-enriched bottom sediments in a restricted hypersaline environment where brines accumulated below playa surfaces. Some laboratory experiments have attributed the rosette-like aggregate growth of DGG crystals to gypsum crystal nucleation on specific organic compounds in parent brines (Warren, 1999, and references therein), but it is not well-constrained whether related growth patterns can form in completely abiotic environments. Additional work is recommended to fully understand the astrobiological implications of these crystal mold features seen on Mars (e.g., McLennan et al., 2005).
4.1.3. Pathways A1–A5, later-stage DGG alteration (Fig. 12 Time 1)
DGG was altered by four distinct, postburial diagenetic processes (Pathways A1–A5). In Pathway A1, during periods of lower salinity, DGG was dissolved, leaving behind external crystal cavities within the host mudstone. A second wave of gypsum precipitation infilled the dissolution cavities with secondary bladed gypsum crystals (Pathway A2). In other cases (Pathway A3), gypsum cavities were infilled by secondary gypsum, which then recrystallized to form a mosaic of replacing crystals. In other instances, primary gypsum experienced dehydration (Pathway A4), forming anhydrite-bassanite intermediates, possible at burial temperatures of ∼35°C, shallow depths, and pore-fluid salinity of halite saturation (Warren, 1999).
During diagenesis, cation replacement can create substitution and enrichment within Sr-rich gypsum (Kasprzyk, 2013). Pathway A5 represents Na-Ca-Sr cation-substitution in DGG crystals, including partial or total Sr-substitution to form celestine. Sr was derived from brines that came into contact with the dissolution fluids and may have caused gypsum recrystallization. The Na-Ca sulfate endmember, glauberite, was a minor phase observed in some samples (Table 2).
Pathway A5 was discovered through Raman analyses. Previous Raman analyses of Ca sulfates and Na sulfates have reported only rare cation substitutions (e.g., Dharmasena and Frech, 1993). We recommend further studies on the effects of cation substitutions on Raman-based sulfate identification, especially for phases known to exist on Mars. One example of this line of research was published for carbonates (Rividi et al., 2010). Such studies could better inform the use of cation substitution as paleoenvironmental proxies (Butler, 1973; Baker and Bloomer, 1988; Donovan and Rose, 1992). This could provide useful insights into diagenetic effects on BPP.
What can be said about the timing of DGG alteration events based on our analyses? Aside from the fact that dissolution preceded infilling and recrystallization in the playa mudstones, and that Sr and Na enrichment were likely contemporaneous with gypsum replacement and/or recrystallization in those environments, the timing of Pathways A1–A5 remains relatively unconstrained (indicated by the double-sided arrows in Fig. 12).
4.1.4. Pathway B0, halite pod formation (Fig. 12 Time 1)
Ca consumption by gypsum led to the progressive enrichment of Na and Cl, along with an increase of Mg and Ca in ponded brines (Jankowski and Jacobson, 1989). Pathway B0 is characterized by halite precipitation within localized brine ponds, visible as halite-rich pods exposed on mid-slope quarry walls (Fig. 2).
Our outcrop observations and textural relationships are consistent with halite pod formation by localized upwelling and karstification of subsurface evaporites accompanied by the shallow subsurface circulation of saturated brines. The halite pods are aligned laterally, suggesting that their formation occurred concurrently, midway through the stratigraphic time sequence exposed in the quarry walls. Pods are also elongated vertically (perpendicular to bedding), suggesting that they persisted for some time.
Periodic depositional events on playas commonly deposit fine beds or laminations. The absence of such fine sedimentary layering in the mudstones is consistent with the destruction of laminations by haloturbation. Other evidence for haloturbation in mudstones comes from clasts seen in thin section (Fig. 3B), which may have become deformed by compaction after diagenetic burial of evaporite beds after being flushed by undersaturated subsurface waters (Warren, 1999). Similar features have been described as overlying and/or becoming entrained by collapse during the dissolution of salt beds (Warren, 1999).
4.1.5. Pathway B1, replacement of halite by Na sulfate (Fig. 12 Time 1)
Following formation of the halite pods, depletion of Ca, Cl, and increases in alkalinity of subsurface playa brines enriched Mg and SO4 relative to other cations (Jankowski and Jacobson, 1989). In OISM evaporites, when Mg concentrations exceeded SO4, neutral C1-SO4 brines enriched in Na and Mg were predominant, and this led to the precipitation of Na sulfates. The main Na sulfate associated with the halite pods is thenardite, which has locally replaced halite. Evidence for this is based on two thin section observations: (1) thenardite always surrounding halite (Fig. 8A) and (2) corroded remnants of halite within thenardite crystals (Fig. 8C, 8D).
Other Na-rich sulfates besides thenardite may have been deposited via cyclical evaporation to near dryness (Wadell, 1972). An additional Na-rich phase, glauberite, was identified in the quarry in minor abundances. Mirabilite was also expected, as it reportedly appears in efflorescent crusts on the playa during cooler seasons (Wadell, 1972; Anthony et al., 1982; Phillips, 1987) but was unidentified in our study. This is likely because it is stable at cooler temperatures and dehydrates (in situ or in the lab) to thenardite, which is stable at warmer temperatures (Tursina et al., 1980).
4.1.6. Pathway C0, magnesite cementation of playa sediments (Fig. 12 Time 2)
During the Pleistocene, an extensive perennial lake formed in the Verde Basin, depositing lacustrine carbonates above the older playa sequences (Donchin, 1983). During this time, OISM mudstones in the subsurface were further lithified by widespread precipitation of magnesite-rich diagenetic cements. The source of the carbonates remains unconstrained but may have been derived from the dissolution of Colorado Plateau marine limestones north of the Verde Basin.
4.1.7. Pathway D0, Fe-rich oxidation (Fig. 12 Time 2)
A late-stage change in depositional and diagenetic conditions is implied by playa mudstones exposed in the upper quarry walls where gray and red mudstones are interbedded. Interbedding suggests that environmental conditions during deposition alternated between reducing conditions and oxidizing conditions. Red mudstones could have provided favorable conditions for the circulation of Fe-rich oxidizing pore fluids through the mudstones, cementing them with Fe oxides and imparting their red color, while gray mudstones were cemented by magnesite during reducing conditions. This is consistent with observations of red mudstones as being coarser (Fig. 5B, 5C) and more permeable than the gray mudstones.
During this period, groundwater-mediated chemical weathering may have produced authigenic Fe-oxides (and perhaps clays). It remains unconstrained whether Fe oxidation occurred during transport of detrital grains or clasts to the playa or during chemical weathering in situ.
Fe-oxide phases were later altered, as suggested by embayment features seen in red mudstone samples (Fig. 5D). Four hypotheses, which remain to be tested, may explain this alteration: (1) migration of fluids through coarser and more permeable sediments, (2) nucleation and growth of Fe-oxide cements on grains during diagenesis, (3) partial removal of Fe-oxide coatings (e.g., during transport or by wind abrasion), and/or (4) acidic chemical dissolution.
Fe-oxide cementation postdates the displacive growth of gypsum in red mudstones, since the exteriors of DGG crystals from red mudstone units were coated with Fe oxide.
It may be possible that Fe-metabolizing bacteria played a role in the Fe-rich cementation in the outcrop. This study did not explore possible contributions of Fe-based bacterial metabolism, such as iron reduction, to the diagenetic pathways discussed here.
Near the end of playa deposition, alluvial fans prograded northward, unconformably depositing poorly sorted gravels on top of the mudstone sequence. These alluvial gravels, thought to be derived mostly from the Black Hills exposed along the southern margin of the Verde Basin, cap the Verde Formation sequence studied at OISM as a <1 m thick pebbly conglomerate unit (Fig. 2H).
4.2. Impacts of diagenesis on long-term BPP in the OISM quarry
To address our study's second goal, here we present an empirical model for the BPP of playa evaporite facies at OISM across diagenetic pathways recognized. The model integrates our diagenetic interpretations with TOC results (Fig. 11; Table 3), which are used as a proxy for evaluating BPP.
The earliest stage of biosignature capture and highest BPP during the playa's earliest history cannot be directly measured today, since the samples we collected represent the combined endpoint of all diagenetic stages in the playa's history. To provide starting-point estimates for BPP in the oldest facies, the gray mudstones, we refer to published literature on TOC values obtained from a variety of similar modern environments (Warren, 1986, and references therein; Carroll and Bohacs, 2001; Schreiber et al., 2001). Modern saline and hypersaline sediments, marine carbonate-rich evaporites, and hypersaline evaporative facies of freshwater, and suboxic environments are all known to have average TOC values between 0.2 and ∼30 wt % (Warren, 1986, and references therein; Carroll and Bohacs, 2001; Schreiber et al., 2001), up to 3 orders of magnitude higher than our mudstone TOC values. Based on this, we assume that initial deposition in our studied environment had the highest BPP, and it was reduced as diagenesis and alteration progressed.
Of the five facies sampled, TOC results revealed two data clusters: highest values in both mudstone facies, and lower values in both DGG facies (associated with both mudstone facies) and the halite-thenardite pod facies. The high BPP in gray and red mudstone facies may be explained by their association with abundant phyllosilicates (especially smectites; Table 2), whose preservation potential is well documented (e.g., Farmer and Des Marais, 1999, and references therein; Kennedy et al., 2002). A second factor may be the pervasive early diagenetic cementation of the playa mudstones by fine-grained, carbonate-rich cement, providing two conditions often cited as enhancing preservation (Farmer, 1995; Farmer, 1998; Farmer and Des Marais, 1999; Westall and Cavalazzi, 2011).
It was anticipated that Fe oxidation in the red mudstone facies would reduce BPP relative to gray mudstones (Allison and Briggs, 1991; Farmer and Des Marais, 1999; Westall and Cavalazzi, 2011). However, an important result of this study is that Fe-rich mudstones exhibited comparable TOC values, and by extension, BPP, as compared to the less oxidized gray mudstones. One reason for this difference may be that Fe-oxidizing fluids came into contact with finer-grained components after carbonate cementation, which protected OM from oxidation.
The BPP of DGG and halite-thenardite pod facies were similar to each other but lower than both mudstone facies. During displacive growth of gypsum, host muds (along with any entrapped organics) were captured inside gypsum and halite crystals. We could not constrain how the BPP of early precipitated gypsum compares to that of DGG after undergoing this later alteration (Pathways A0–A5) because our TOC values reflect a combined BPP over both time and diagenesis, represented by the modern samples we collected. Nevertheless, we assume that later alteration pathways are likely to have reduced the BPP of our DGG samples, relative to the BPP of the original gypsum. Gypsum recrystallization (Pathway A3), dehydration (Pathway A4), Na or Sr cation substitution (Pathway A5) could all have reduced the BPP of the original gypsum, as diagenetic fluids associated with these processes could have flushed and oxidized any OM preserved in the primary gypsum (Warren, 1986; Farmer, 1999; Allwood et al., 2009). The comparable BPPs in both red and gray DGG facies are unsurprising since Fe-oxide interactions with the red DGG facies postdated DGG growth.
Biosignature preservation potential within pods was expected to be high, based on the fact that when ponded brines reach halite saturation, natural waters approach anaerobic conditions (Warren, 1999). This condition is known to enhance BPP. However, OM originally entombed in halite was lost likely during halite dissolution and re-precipitation cycles, causing the lower BPP in pods relative to both mudstone facies.
This finding may contradict studies that have concluded that saline evaporite basins have high BPP (Powell, 1986; Warren, 1986, and references therein). Our lower TOC values in both DGG and halite-thenardite pod samples are consistent with results from Katz et al. (1987), who concluded that evaporite facies rarely yield higher OM quantities relative to associated carbonate members. They cited three reasons for this: (1) density contrast between hypersaline brines and suspended OM retards its settling rate and prolongs exposure to oxidation, (2) concentrated brines and diagenetic fluids support oxidizers that degrade labile organics, and (3) rapid evaporite precipitation rates may dilute suspended OM reaching the sediment-water interface. Any of these factors may have contributed to the lower BPP of both the halite-thenardite pod and DGG facies.
Additional factors that may impact BPP on Mars were not considered in this study, such as the destructive impact of ionizing radiation and heavy particle bombardment on the martian surface. Ionizing radiation is thought to degrade organic molecules (Dartnell, 2011; Dartnell et al., 2012), thus reducing their preservation potential.
4.3. Microscale, nondestructive analyses to understand BPP
Although we infer BPP from our bulk TOC and C isotope results, our analyses are limited to bulk sample analyses and not representative of in situ, nondestructive methods. Summons and Hallman (2014) point out that when interpreting biogenicity analyses, carbon isotope values alone cannot be taken at face value. Additionally, for TOC analyses, alteration pathways (e.g., cation-exchanged domains) cannot be isolated from bulk crystals. Our study highlights recommendations of the astrobiology and Mars communities (Des Marais et al., 2008; MEPAG, 2015) that defining BPP requires microscale, nondestructive analyses. Future studies are encouraged to refine such methods in order to improve the understanding of BPP.
Our study also illustrates the importance of using multiple, complementary analyses when assessing the BPP of any complex environment. For example, EPMA confirmed a halite composition when Raman could not because halite lacks Raman-active vibrational modes. VNIR confirmed Fe-oxide compositions when XRPD could not due to detection limits. For the most successful biosignature searches on Mars, rover instruments should obtain complementary data sets to constrain interpretations when selecting samples with the highest BPP for MSR.
One additional point regarding analyses for BPP studies should be noted. Our Raman analyses were sometimes challenged by a fluorescence background, a common problem when using 532 nm excitation Raman spectroscopy, especially in the types of Mars analog samples we studied (Shkolyar et al., 2018). Our choice to use a CW 532 nm Raman spectroscopy instrument was based on the ubiquity of this type of Raman facility as well as its small laser spot size which allowed us to target micron-scale features, like kerogen. Raman is valued for providing nondestructive, sensitive analysis of organics and minerals, and three Raman instruments will soon fly on board space missions for the first time on the ExoMars and Mars 2020 payloads. Many strategies have been employed in Mars analog biosignature studies to mitigate fluorescence. These studies are briefly reviewed by Shkolyar et al. (2018), who make recommendations for future studies seeking to optimize Raman analyses for planetary surface biosignature applications. Shkolyar et al. also highlight the effectiveness of a Raman instrument, similar to Mars 2020 rover's SHERLOC (Scanning Habitable Environments with Raman and Luminescence for Organics and Chemicals; Beegle et al., 2015), for reducing fluorescence in Raman spectra in a suite of diverse natural Mars analog samples, including a Verde Formation mudstone sample.
5. Summary and Conclusions
This study examined playa evaporites from the ∼15-million-year-old Verde Formation of central Arizona as a potential analogue for past martian environments having a high BPP. This study presents an approach for building BPP models that consider the complex diagenetic histories of sedimentary systems. Through fieldwork, we identified and obtained a representative suite of endmember samples. Outcrop-scale observations were integrated with data sets from six complementary lab analyses to identify five facies making up the Miocene-Pliocene evaporite playa system. Using this approach, our aim was to (1) improve BPP models for terrestrial playa evaporite systems; (2) enhance our understanding of the highest-priority samples to be cached for return to Earth in an MSR scenario; and (3) improve biosignature search strategies in similar ancient environments on Mars.
Our inferred diagenetic history of the OISM evaporites may be summarized as follows. First, fine-grained sediments were deposited onto playa surfaces. DGG then precipitated within playa muds. DGG later underwent later diagenetic pathways, including dissolution, formation of external crystal cavities within mudstones, infilling of crystal cavities with secondary gypsum, recrystallization of that gypsum, as well as dehydration to anhydrite-bassanite intermediates and Na-Ca-Sr cation substitution. Another facies was represented by surface depressions where brine ponds formed and where bottom-nucleated halite grew and was later partly replaced by Na sulfates. Finally, during the Pleistocene, a perennial lake system developed in the Basin. During a later lacustrine phase, magnesite was deposited as a sedimentary cement that partially lithified playa sediments. Environmental conditions later cycled between reducing (with gray mudstones being cemented by magnesite) and oxidizing (where red mudstones were cemented by Fe oxides).
Total organic carbon values were obtained from bulk samples. A BPP model was proposed based on this inferred diagenetic history of playa facies and TOC measurements obtained for each facies as a proxy for BPP. In this model, initial sedimentation of fine-grained muds onto playa surfaces was inferred to be the earliest stage of biosignature capture with the highest BPP. Both gray and red mudstone facies exhibited a higher BPP, compared to the DGG and halite-thenardite pod facies, which showed comparable and lower BPPs. The higher BPP in mudstones was attributed to the abundance of smectites and fine-grained carbonate cements, both factors promoting OM preservation in gray and red mudstones. Analyses of red mudstones showed that Fe oxidation did not reduce BPP relative to gray, non-oxidized mudstones. Low BPP in both halite-thenardite pod and DGG facies was attributed to oxidation, evaporite recycling, and/or OM dilution due to rapid evaporite precipitation.
For similar habitable environments on Mars, this framework may help guide the identification of samples with the highest BPP, supporting in situ analyses and/or caching selections for MSR. With the approach of Mars 2020 and ExoMars, further analog studies are warranted to refine and quantify the definition of BPP, especially studies employing in situ, nondestructive, and microscale techniques to enhance the success of biosignature sampling strategies employed on these missions.
Beyond Mars, our BPP model may potentially be extended to additional high-priority astrobiology targets. These include Europa's putative ocean and icy surface, which may contain evaporite assemblages similar to those in this study, including Mg-Na-SO4-Cl brines, Na sulfate hydrates, and NaCl (McCord et al., 1999; Zolotov and Shock, 2001; Prieto-Ballesteros et al., 2003; Hand and Carlson, 2015).
Footnotes
Acknowledgments
We gratefully acknowledge the use of the Raman facility within ASU's LeRoy Eyring Center for Solid State Science through the help of Emmanuel Soignard. Brian St. Clair, Natalya Zolotova, Gwyneth Gordon, and especially Candace Ashley (ASU) are thanked for help with TOC measurements. Philip Gopon (UW-Madison) is thanked for EPMA sample preparation and useful insights on EPMA methods. Axel Whittmann (ASU) is thanked for help with EPMA measurements. Christopher Haberle (ASU) is thanked for help with VNIR spectroscopy analyses. ASU's NASA's Mars Program and the NASA-SETI Astrobiology Institute are acknowledged for financial support. S.S. thanks ASU's Graduate and Professional Student Association (GPSA) for conference travel and support of analytical equipment through ASU's Graduate Education Graduate Research and Support Program (GRSP). The Postdoctoral Fellowship at Carnegie Institution for Science is acknowledged for support. Thanks are also given to two anonymous reviewers for their encouragement and reviews that improved this manuscript.
Author Disclosure Statement
No competing financial interests exist for any of the authors.