
Abstract
The hydrogen peroxide (HP) crucible hypothesis proposed here holds that life began in a localized environment on Earth that was perfused with a flow of hydrogen peroxide from a sustained external source, which powered and mediated molecular evolution and the protocellular RNA world. In this article, we consolidate and review recent evidence, both circumstantial and tested in simulation in our work and in the laboratory in others' work, for its multiple roles in the evolution of the first living systems: (1) it provides a periodic power source as the thiosulfate–hydrogen peroxide (THP) redox oscillator, (2) it may act as an agent of molecular change and evolution and mediator of homochirality, and (3) the THP oscillator, subject to Brownian input perturbations, produces a weighted distribution of output thermal fluctuations that favor polymerization and chemical diversification over chemical degradation and simplification. The hypothesis can help to clarify the hero and villain roles of hydrogen peroxide in cell function, and on the singularity of life: of necessity, life evolved early an armory of catalases, the continuing, and all-pervasive presence of which prevents hydrogen peroxide from accumulating anywhere in sufficient quantities to host a second origin. The HP crucible hypothesis is radical, but based on well-known chemistry and physics, it is eminently testable in the laboratory, and many of our simulations provide recipes for such experiments.
1. Introduction
Over recent decades, our understanding of biological evolution, and the theory itself, has made giant strides (Müller, 2017), but the same cannot be said for its beginnings, the origin of life. Even specific breakthrough advances, such as nonenzymatic RNA replication and growth and division of vesicles (Szostak, 2017), are but fragments of the greatest story never told. A sociological hindrance may be that origin of life research is inherently interdisciplinary and multifaceted so that there is little common language and no unified framework to guide theory and practical developments.
The problem may be rooted at a deeper level, though. That relatively little progress has been made on understanding abiogenesis, despite best efforts over many decades by top laboratories, suggests that we may have been missing or overlooking some fundamental insight, and that a radical conceptual leap may be needed. Similar or related sentiments from the recent literature are collected in Supplementary Data Section S1 (Supplementary Data are available online at
The potential importance of hydrogen peroxide in pre- and protocellular evolution is illustrated in a recent work by Springsteen et al. (2018). The authors report experiments that generate a nonenzymatic simpler analogue of the citric acid cycle (the most fundamental and universal metabolic cycle), in which glyoxylate is oxidized by hydrogen peroxide. They hypothesize that this cycle could have emerged prebiotically, giving references for possible sources and occurrence of hydrogen peroxide that we have also cited previously (Borda et al., 2001; Liang et al., 2006). Their experiments allow us to draw protometabolism under the umbrella of the HP crucible hypothesis, together with polymerization and replication. Furthermore, Lineweaver and Chopra (2012) and Stueeken et al. (2013) recognized that the environment for abiogenesis was not necessarily inhabitable by the first organisms. The HP crucible hypothesis supports a stronger statement that the physical environment where life began must have been radically different from any that has existed since and most likely was uninhabitable by cellular life.
What was this environment like? Liquid water may have persisted on Earth since ∼4.3 Gya (Kasting and Catling, 2003; Sleep et al., 2011); there is evidence for fossilized microorganisms that may be 4280 Gy old (Dodd et al., 2017), and at least parts, if not most, of Earth's near-surface were likely teeming with varied cellular life by 3.7 Gya (Nutman et al., 2016). Thus, abiogenesis may have occurred within a time window of only a few hundred thousand to some millions of years during which chemical evolution yielded pre-RNA (Kaur et al., 2017) and the RNA world, lipids and peptides, and simple metabolic cycles (Springsteen et al., 2018), which coevolved to protocellular and fully cellular organisms. To us this suggests that life arose in a localized, relatively ephemeral highly abiogenesis-favoring environment, and, having arisen, perforce rapidly evolved catalases that prevent any emergence of “life 2.0” (Davies, 2011), by effectively “pulling up the ladder” (Bains and Schulze-Makuch, 2015).
The HP crucible hypothesis constrains the general milieu or even the locality on Earth where life likely originated. Possible sources of hydrogen peroxide on the early Earth were discussed and reviewed in the work of Ball and Brindley (2014). If produced by intense ionizing radiation from a natural fission reactor (Draganic, 2005; Adam, 2007), there may be an outside chance, in future, of locating the actual crucible of life because such leave a (diminishing) signature. The hydrothermal vent setting for the origin of life is supported by the H2O2-producing experiments of Borda et al. (2001) and Foustoukos et al. (2011), with significant H2O2 produced even in anoxic environments (Gil-Lozano et al., 2017). The ionizing radiation that bathed the early Earth would have produced dilute H2O2 rain on land and sea and in the ocean surface layer (Draganic, 2005; Dartnell, 2011), but we lack hard evidence, at present, that H2O2 from these sources concentrated locally in continuous flow by, for example, preferential evaporation of water, although this is amenable to experimental investigation.
In the following sections, we elaborate on the three key words in the title, with respect to the HP crucible hypothesis. Section 2.1—the power: What was the driving force for the transitions from geochemistry to biogeochemistry to biochemistry? We review and interpret simulations in which the thiosulfate–hydrogen peroxide (THP) redox thermochemical and pH oscillator was used to drive RNA processes. Section 2.2—the glory: Although hydrogen peroxide has the power—the enthalpy of the peroxide bond—it has been denied the glory, until recently being cast as life's villain. We restore it to its rightful place as a likely driver and mediator of life's origin. Section 2.3—multiple roles: We place the stochasticity of THP oscillations at center stage in the origin of life drama. We then discuss the properties of hydrogen peroxide with respect to their possible effects on incipient life. In Section 3, we draw the tree of life into the HP crucible hypothesis. We sum up briefly in Section 4. Mostly, we excluded references that were cited in our previous works, instead citing newer relevant literature.
2. H2O2—“Villain with a Smiling Cheek”
2.1. The power
The evolution of templated replication and biochemistry required power from a sustained source, as well as a continuous supply of nutrient, to counter the otherwise inevitable drift to equilibrium and collapse of biomolecules to tars and tholins (Deamer and Weber, 2010; Pascal et al., 2013; Russell et al., 2013; Sutherland, 2017). Molecular evolution and the RNA world most likely drew on thermal power directly (Muller and Schulze-Makuch, 2006), if the milieu was porous rock around a heat source such as a submarine hydrothermal vent (Sleep et al., 2011; Lane and Martin, 2012) or a natural nuclear reactor (Ebisuzaki and Maruyama, 2017). That such a power source must have been periodic in temperature is now well recognized (Spitzer, 2013; Spitzer et al., 2015), because in the absence of fully developed cellular machinery, thermal cycling is required for processes such as adsorption/desorption, polymerization/hydrolysis, and RNA replication. Several mechanisms have been proposed: diurnal and seasonal cycles (Mamajanov et al., 2014), tidal flows periodically activating natural nuclear reactors (Adam, 2016), periodic entrainment by hot geothermal streams (Szostak, 2016), and thermoconvection in rock pores (Kreysing et al., 2015).
In Ball and Brindley (2014, 2015), we tested the novel idea that the thermal cycling needed to drive the RNA world may have been provided by thermochemical oscillations. In particular, we identified the well-known THP redox oscillator, encouraged by the substantial evidence of existence on the prebiotic world of the necessary reactants [see Reactions (S1)–(S2) in Supplementary Data Section S2]. We used experimental rate data to show that the THP oscillator could power replication (by complementary strand pairing) of short RNA species and further that the associated pH cycling may have played an important adjunct role. The reliance of all life on pH gradients to drive metabolism has prompted many researchers to investigate how this arose. Lane and Martin (2012) asked: “Some form of chemiosmotic coupling probably evolved very early in the history of life, arguably before LUCA (Lane et al., 2010); the question is how, and why?” Lane's hypothesis is that natural proton gradients across alkaline hydrothermal vents were harnessed by the earliest living cells. The HP crucible hypothesis provides a viable alternative (or perhaps complementary) possibility since in a bounded spatially extended system, thermo-pH oscillations manifest as travelling waves and patterns. The gradients of a travelling thermo-pH wave powered by the THP oscillator can provide a proton-motive force that may have driven metabolic processes in the RNA world.
This adds another dimension to the HP crucible hypothesis—it releases the RNA world and protocells from the narrow confined environment of a hydrothermal vent and, at the same time, provides the first intimation that
The universal dependence of all life on the power of transmembrane pH gradients may have originated in the precellular world.
The vitally important functions of biologically produced hydrogen peroxide in cells may be rooted in the dynamic association of pre- and protocellular life with an abiotic hydrogen peroxide-rich medium.
Additionally, we found that pH cycling provided by the THP oscillator [see Reactions (2) in Supplementary Data Section S2] improved ribozyme activity significantly over constant pH (Ball and Brindley, 2016b). Such pH cycling also provides a possible resolution of the “replication versus ribozyme activity” paradox (Ivica et al., 2013), which is that
folded structures are necessary for ribozyme activity, but replication requires unfolded sequences.
In the dynamical THP medium, a high temperature and low pH phase of the cycle may favor destructuring of a ribozyme for replication, and then, replicated duplexes may adopt active ribozyme structures during a cooler higher pH phase.
It is important to stress that in none of these functions is thermochemical cycling necessarily exclusive; it could operate in a complementary, even resonant, manner with the nonchemical cycling mechanisms, which have been proposed by others and referred to earlier.
2.2. The glory
We have shown that hydrogen peroxide could have provided the “power,” and much more, for the origin of life. However, the “glory” it deserves has not been accorded. Indeed, for more than a century, until the early 2000s, hydrogen peroxide was more sinned against than sinning, reviled and vilified as anti-life, its vital role in life from origin to present day misunderstood and even rejected. Biologists saw it as toxic to cell metabolism and gene transcription fidelity.
Chemists see it as destructive to molecular bonds: “If we drop some RNA into a flask of hydrogen peroxide it would be consumed within minutes!” No doubt that is true, but the origin of life environment was not, could not have been, flask-like. The dynamic far-from-equilibrium flow environment we are contemplating here, with competing substrates for oxidation, is utterly different from the naysayers' static flask that is left alone to equilibrate. In the THP oscillator–RNA system simulated in Ball and Brindley (2015), we quantified that a hydrogen peroxide molecule is ∼65,000 times more likely to encounter a thiosulfate ion than a G residue, the oxidizable base of the ribonucleotides. Given the intermittent spikes produced by the THP oscillator, RNA contact is likely to be even more fleeting. [We note also that the protometabolic cycles of Springsteen et al. (2018) were catalyzed, rather than destroyed, by hydrogen peroxide.]
Since the 2000s, there has been a steady rehabilitation of hydrogen peroxide (and other reactive oxygen species [ROS]) with respect to its role in cellular life, and a much more nuanced view of its essential place in biology has emerged (Roy et al., 2017). Its essential roles in metabolic processes and redox signaling in aerobic cells were reviewed by Sies (2017), who pointed out that, significantly, it is a transcription-independent signal molecule, and by Kuksal et al. (2017), and it is known to be produced by facultative anaerobes (Lisher et al., 2017; Sekar and DiChristina, 2017).
Therefore, rather than dismissing hydrogen peroxide as being anti-life, important and valid questions that we urge to be investigated experimentally are: Taking as given the persistence of a localized environment, rich in hydrogen peroxide and the building blocks of life, in dynamic nonequilibrium flow, what other components, conditions, and structures must be present, absent, or ongoing to kick-start molecular evolution? What types of nucleotide sequences are resistant to oxidation? Could exposure to hydrogen peroxide select for such sequences? How might the system dynamics evolve so as to minimize such exposure? What stabilizing agents may have been present?
As recommended by Spitzer (2013) and Agerschou et al. (2017), more experimentation using open driven systems should be encouraged, representing better the sustained far-from-equilibrium conditions necessary for the origin and continuing existence and evolution of living systems.
2.3. Multiple roles
In addition to its influence as the oxidant of the THP oscillator, hydrogen peroxide may play other roles in the drama of life's origin and early evolution. An indirect role—but one of crucial importance—comes about because the THP oscillations, in any realistic setting, are not perfectly uniform in amplitude and period but are subjected to random perturbations. Hydrogen peroxide also may have had more direct effects, chemical and structural in nature. In this section, we focus on these aspects of the HP crucible hypothesis.
2.3.1. In the beginning was disorder
Although the necessity of an open far-from-equilibrium flow environment to create and maintain life is well understood (Branscomb et al., 2017), the effects of random fluctuations of a nonequilibrium flow on prebiotic chemistry have received much less attention. Plausible environments for life's origin were temporally messy because input flows through an inhomogeneous matrix were subjected to random fluctuations and chemically diverse because the resulting output fluctuations produced a wide range of compounds (Guttenberg et al., 2017). Branscomb et al. (2017) stated that “In this world, in fact, fluctuations are everything,” and we agree. But their model contained only Brownian motions or normally distributed random thermal fluctuations, which are not special to far-from-equilibrium systems—they occur regardless of equilibrium status. Something more than their Brownian ratchets is needed to drive chemical processes into living ones.
The simulated nonequilibrium flow experiments reported in the work of Ball and Brindley (2017) mimic conditions in a hydrothermal porous rock setting by (1) using a “toy train” model for reacting flows through a system of interconnected inhomogeneous rock pores (Supplementary Fig. S1 in Supplementary Data Section S3) and (2) setting the inputs as normally distributed random quantities, that is; subject to Brownian motions. Within this paradigm, we simulated surface-catalyzed dimerization of prebiotically relevant monomers, driven by the THP oscillator. The dynamical system model for the toy train is reproduced in Supplementary Data Section S3. We summarize the results in Figure 1.

Of relevance here is the following result obtained rigorously by Bertola and Cafaro (2015): Gaussian perturbations through the boundary of a near-equilibrium, nonreactive, closed system do not induce Gaussian fluctuations in the outputs.
Their output temperature and entropy fluctuations followed a gamma probability density function (Fig. 1b). This is not surprising. A gamma distribution has positive skewness S and is left-weighted (the median M < the mean). In the form of the Maxwell–Boltzmann distribution, it applies to the realm of dilute ideal gases at or near equilibrium.
What does the output fluctuation spectrum of our far-from-equilibrium, reactive open flow system look like? The output fluctuations of temperature oscillation maxima, as experienced by the monomers and dimers, yielded the distribution shown in Figure 1c, and the smoothed envelope of the histogram is shown in Figure 1d. The relevant summary statistics for the distribution are: median M = 0.090, skewness S = 0.27, and kurtosis κ = 0.19 (as calculated by Excel).
The contrast between the distributions shown in Figure 1b and d is remarkable. Evidently, the reactive mass flows, strong nonlinearity, and dynamic far-from-equilibrium drive—all necessary (but not sufficient) conditions for life—alter the distribution (Fig. 1b) profoundly. What do these results mean?
Fluctuating inputs are necessary for a growing, diversifying prebiotic system. Such a system must sample a wide range of temperature extrema to produce a variety of products from reactions having different activation energies.
The positive median and negative skewness of the output fluctuations weight the system toward higher temperature perturbations. Thus, the precious, high activation energy synthetic reactions that the prebiotic nonenzymic world must access to grow, polymerize, and diversify are favored.
But the temperature perturbations must not be too high, and must not occur too often! There would be optimum values of the summary statistics such that the system spends enough time at high enough temperatures to carry out essential high activation energy reactions, but not so much that the reactant is totally consumed or destroyed, or surfaces degraded. Characterizing this distribution in explicit form could effectively give us a “fundamental equation of life,” which may provide useful guidance in designing molecular evolution experiments: they should be messy, as suggested by Szostak (2011), but not too messy. It also suggests a new strategy in the search for habitable worlds and emergent extraterrestrial life: sample the temperature fluctuations. Applying this diagnostic to potential extraterrestrial habitats could easily and cheaply eliminate unlikely ones—those that are not sufficiently out of equilibrium to support life and those where the temperature fluctuations are too extreme.
We can see why a system close to equilibrium cannot give rise to or sustain life. Under output fluctuations that follow the gamma distribution shown in Figure 1b, a high activation energy reaction would undergo its low activation energy reverse with greater probability, and development of a more complex prebiotic world would be impossible.
Branscomb et al. (2017) stated that “There is no way to selectively cause the thermal fluctuations you want …,” but as evinced by the weighted fluctuation distribution rendered in Figure 1c and d, life may have found a way of doing just that! Furthermore, this distribution adds support to the notion that life may be able to produce the fluctuations it requires simply by operating far (but not too far) from equilibrium.
2.3.2. Two (oxygen atoms) are better than one
Let us take a closer look at hydrogen peroxide, as medium and molecule, in terms of how its structure and properties may have facilitated abiogenesis.
The observed homochirality of the nucleic and amino acids of life on Earth has attracted much research (Plasson et al., 2007; Blackmond, 2011). Proposed mechanisms for the emergence of homochirality were reviewed from a physics perspective by Walker (2017) and Goldenfeld et al. (2017). The hydrogen peroxide molecule too is chiral, due to quantum mechanically hindered rotation about the O–O bond (Hunt et al., 1965; Zhang et al., 2009). In Ball and Brindley (2016a), we used a simplified mathematical model to demonstrate a proposed two-tiered mechanism by which this axial chirality may have affected the
Here, we present a probabilistic argument for hydrogen peroxide-mediated emergence of homochirality, illustrated qualitatively in the schematic drawing in Figure 2. At t
0, there is a marginal excess of
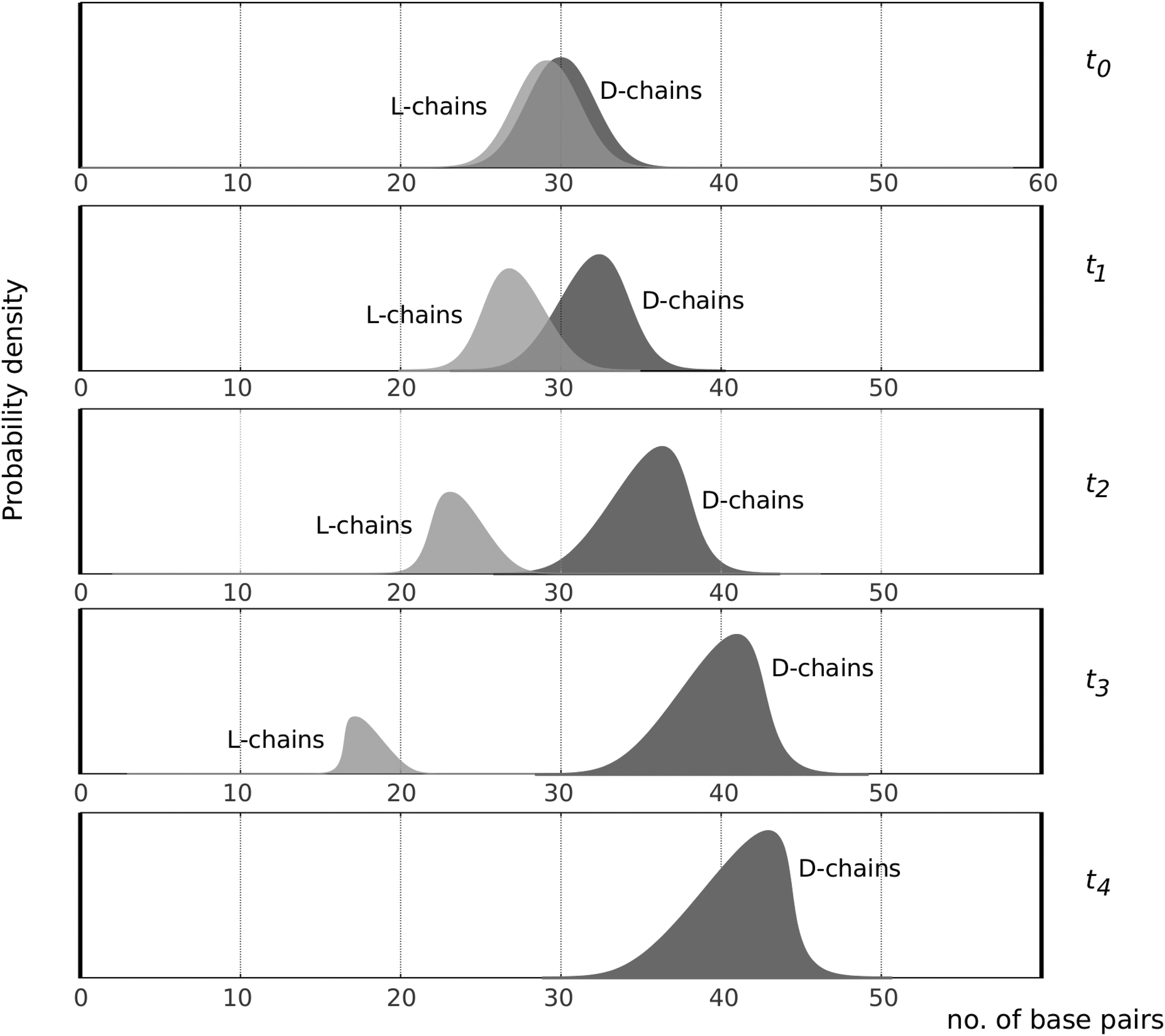
Schematic drawing of hypothetical distributions of chain lengths of
The mean chain length at t
0 of around 30 is arbitrarily chosen. On the basis of this discussion, we predict that the “true” evolving distributions, which could be obtained by simulation experiments, would evolve left-skewed and right-weighted for the
Let us assume enantiospecific D–P and L–M interactions. Then, the molecular fraction D/P increases while the fraction L/M decreases over time because facile interconversion means that M and P must remain constant and equal. When the enantiomeric excess
This mechanism is compatible with and complementary to some other mechanisms that have been proposed for prebiotic homochiral amplification, for example, breakage and fusion of homochiral chains (Blanco et al., 2011).
We also collected, from primary sources, and tabulated in Ball and Brindley (2016a) many of the relevant physical, thermodynamic, and electrical properties of hydrogen peroxide and its aqueous solutions and found that, in particular, the higher surface tension and viscosity favor vesicle formation and division, that hydrogen peroxide solutions are more polar, H-bonded, and highly structured than pure water, and that homochiral clusters may persist. None of these effects is dramatic, but we do not want high drama in the RNA world. (A dramatically larger surfactant effect would really gum up the prebiotic works!) In a continuous flow system, small effects are nondestructive but cumulative over time—this is the essence of the mediation aspect of the HP crucible hypothesis.
3. Catalases and the Singular Tree of Life
3.1. Burning bridges
If we accept that life may have required an HP crucible to originate, we must also ask: What aspects of subsequent evolution, that currently are a mystery, can be explained by the HP crucible hypothesis? What does it say about the tree of life? Let us pay attention to this, then, and see where it takes us.
That all life descended from a common ancestor has been widely accepted as a fundamental dogma of biology since Darwin and Pasteur. The homochirality of life supports a single origin, and by extension at a single locality, as does the near-commonality of the genetic code and basic metabolism; and independent tests using model selection theory provide strong corroboration (Theobald, 2010). Phylogenetic interrogations of genomic and protein databases have built up a solid picture of this last universal common ancestor (LUCA) at the bifurcation to Archaea and Bacteria around 4 Gya and its functionality and capabilities (Woese et al., 1990; Hedges et al., 2014; Becerra and Delaye, 2016; Weiss et al., 2016).
The “tree of life” is widely used both as metaphor and cartoon for the homology of life. Its branches vanish as they grow, so we can see and study directly only its existing tips and such vanished tips for which we have found fossils; the antecedent structure mostly is deduced. This deduced tree of life is sketched in Figure 3a in minimal form, where branches representing only the three domains, Bacteria, Archaea, and Eukarya, arise from a single origin.

Known
This fundamental dogma has been questioned though. There would seem to be no good reason, prima facie, why cellular life should have arisen just once on Earth. The existence of microbial “shadow biospheres” that arose from independent origins was proposed by Cleland and Copley (2005) and discussed further by Davies et al. (2009) and Davies (2011). Surely, a shadow biosphere could coexist with the known biosphere—after all, the three domains of life, Bacteria, Archaea, and Eukarya, rub along in more-or-less peaceful, often interdependent, coexistence to this day. Could there even exist a remnant RNA world ecosystem in some niche on Earth?
A few imagined shadow trees of life are sketched in Figure 3b, c, d, and e (also in minimal form, for obvious reasons). In Figure 3b, shadow life (perhaps of opposite chirality) originated contemporaneously with known life and persists (unobserved by us) to this day. No evidence for such a shadow biosphere has ever turned up, although we do not rule it out entirely. In Figure 3c, an independent contemporaneous origin of life led to a shadow biosphere that coexisted for some time with the early stages of the known biosphere before becoming extinct, but not before some lateral gene transfer (LGT) had taken place. This assumes the shadow genetic code was nearly identical to the known genetic code, so we could never corroborate this scenario. In Figure 3d, a weak shadow tree was born when evolution was well advanced and died before present, and Figure 3e shows a recently originated shadow tree that persists (notionally) to this day.
The HP crucible hypothesis can explain why scenarios such as those shown in Figure 3d and e are impossible: Quite simply, the all-pervasive presence of catalases (and peroxidases) produced by cellular life, since before LUCA, in all habitable niches would forestall any possible second origin. Since the HP crucible hypothesis holds that a medium rich in hydrogen peroxide was necessary (although not sufficient) for molecular evolution to protocellular structures, the unceasing action of catalases in oxidizing hydrogen peroxide effectively would prevent the development and persistence of such an environment again.
The evolution of catalases was usually assumed to have coincided with oxygenation of the atmosphere 2.5 Gya, attributed to rapid spread of oxygen-evolving photosynthetic organisms. But this view is shifting backward in time (Slesak et al., 2016; Olson et al., 2017). Phylogenetic analysis was used by Slesak et al. (2016) to show that enzymatic antioxidant systems were functioning in early anaerobic protocells at the time of LUCA. The HP crucible hypothesis lends powerful support to the early evolution of catalases. Calvin (1959) proposed that catalases developed from simple free Fe2+, through iron-heme complexes to associate with peptides to the active site of the fully translated catalase protein, and there has been no contradictory finding since.
Evolution can be thought of as the burning of a succession of small bridges: the results of a transformational evolutionary step usually destroy the preconditions for its own occurrence. This is described as “pulling up the ladder” in the work of Bains and Schulze-Makuch (2015), and in nontechnical terms in the book by Schulze-Makuch and Bains (2017). But the first cellular life destroyed utterly the bridge across the biggest divide of all, which spanned the cellular and the RNA worlds! Any chance of re-building it was permanently rubbed out by the necessary early evolution and subsequent persistence of catalases. Living cells themselves forbid the possibility that a competing biosphere, Life 2.0 (Davies, 2011), could arise and out-compete, coexist with, or devour Life 1.0!
3.2. Bottlenecks and bifurcations
If the tree paradigm applies to evolution of the biosphere from LUCA onward, by continuity it must also apply to pre-LUCA evolution. What do the branches and twigs on a pre-LUCA tree of life actually represent? A single branch is the surviving coarse-grained thread, identified a posteriori, of a myriad successive bifurcations to smaller twigs, none of which survived competition with the lineage that became the living thread. A pre-LUCA tree of life is postulated in Figure 4. It is rudimentary, and undoubtedly important details are lacking or incorrectly placed in the hierarchy, but it is a first pass and, anyway, its function is heuristic. Consider the branch, distinguished as a thick dark thread, that gives rise to LUCA.
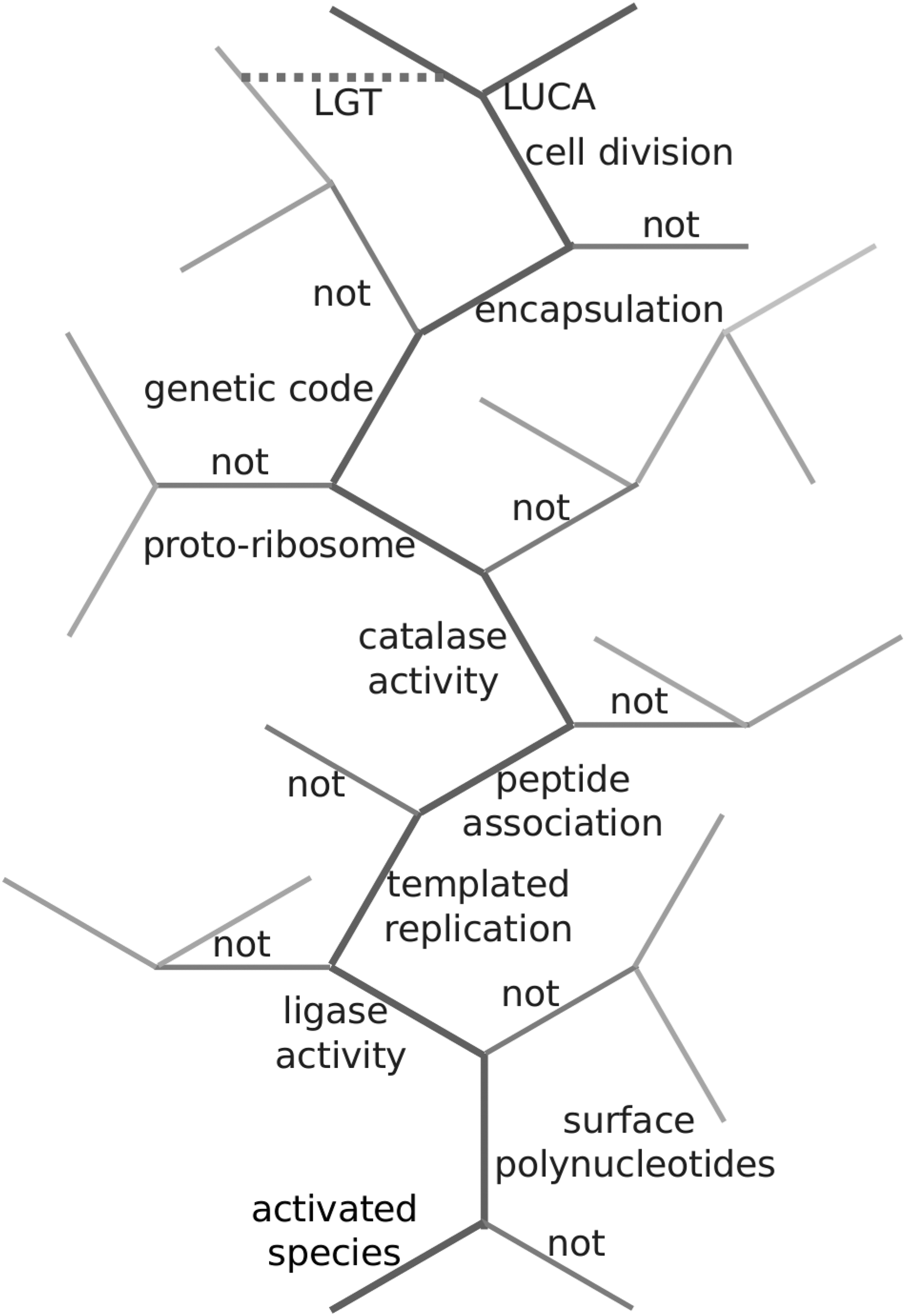
A postulated partial tree of emergent life leading to LUCA. The branch that produces LUCA is a thick dark thread; gray or pale twigs do not survive. Possible LGT is indicated by the dotted line. LGT, lateral gene transfer; LUCA, last universal common ancestor.
A full picture of this branch would look quite a confusing mess, an open network of uncountable extinct twigs with, postgenetic code, LGTs, but the simplified picture is helpful in guiding our thoughts about the tree of life concept in terms of the HP crucible hypothesis.
Imagine the tree growing upward through a fluctuating hydrogen peroxide-rich flow, emanating at its source, which powers, directs, and mediates the biomolecular species and functionalities that evolve, the homochirality, surface polymerization, replication, catalases, and so on, annotated in Figure 4. As the tree grows and bifurcates the flow also becomes more dilute, due to source attrition, and action of catalases, enzymes must take over from hydrogen peroxide's direct drive and actions, and protective encapsulation is selected for, where salts are included and hydrogen peroxide excluded from the protocell. But living protocells had become dependent on the depleting nonbiotic hydrogen peroxide, not least the pH gradients provided by the THP oscillator to power them! So abilities to exploit transmembrane pH gradients were selected for.
We note that there may have been a period during which the intracellular solvent of protocells was an H2O2/H2O solution. Houtkooper and Schulze-Makuch (2007) hypothesized that living organisms evolved on Mars that used a water/hydrogen peroxide mixture as an intracellular liquid, based on the presence of hydrogen peroxide in the martian soil, which they proposed is of biogenic origin. Bonfio et al. (2018) explored experimentally a proposed path whereby iron–sulfur peptides in protocells pass electrons to nonbiologically produced hydrogen peroxide (0.5 mM), which is then reduced to provide a pH gradient across primitive membranes. Hydrogen peroxide is singled out in particular by Taverne et al. (2018) as having played a major role in evolution of life on Earth, being both a driver and a consequence of evolution. These works strengthen the HP crucible hypothesis, adding experimental support and new evidence for a significant participation of hydrogen peroxide in very early cellular life.
This discussion is purposely (and necessarily) speculative. It is intended to stimulate further ideas concerning possible feedbacks between an evolving biomolecular system and its energetic medium, and how these interactions themselves may have shaped emergent life. It is by setting out these speculations, too, that we can see, in principle, how hydrogen peroxide may have played an active role in shaping the tree of life by providing a pregenetic selection pressure—as do biologically produced ROS to this day on genomes and epigenetically (Sies, 2017).
4. Conclusions
The HP crucible hypothesis can explain or provide a mechanism for several necessary (but not sufficient) conditions and processes for the origin of life on Earth, or so far unexplained universal characteristics of life: In a primordial medium rich in hydrogen peroxide, produced continuously by a localized source, the THP oscillator can provide the far-from-equilibrium drive and thermal periodicity necessary to power chemical evolution and the RNA world. The experiments of Springsteen et al. (2018) provide a possible example of this driving role. Since the THP redox system also behaves as a pH oscillator, it can provide pH (or chemical potential) gradients in a spatially extended bounded matrix, which can augment the thermal drive. The universal dependence of all life on transmembrane pH gradients may have its origins in the THP oscillator. The Gaussian inputs that drive the THP oscillator in a natural setting produce a non-Gaussian distribution of output thermal fluctuations that is highly enabling for chemical evolution. This distribution is a “fundamental equation of life,” which can guide experiments and searches for habitable worlds. As a chiral molecule, hydrogen peroxide has the potential ability to beget chiral segregation of biomolecules (by the Curie principle) and effect homochirality of the nucleic acids. Its structural and molecular properties in solution may facilitate chemical evolution and the RNA world. It effectively grooms the evolving pre- or protocellular world for survival in the absence of its external supply. Evolution of living systems in an HP crucible may have stimulated the early (pre-LUCA) evolution of catalases and peroxidases and may help explain how life's conflicted relationship with ROS arose. Life's origin in a hydrogen peroxide-rich medium can explain the singularity of the tree of life. The rapid proliferation of cellular life in all habitable niches on Earth means that all-pervasive efficient catalases have prevented any subsequent buildup of hydrogen peroxide.
There is much more work to be performed on the HP crucible hypothesis. Inevitably, we will be shown to be amiss, or lacking, in many details. There are also large gaps—for now. (For example, Where, exactly, on Earth did the origin of life take place? We simply don't know.) But we stress that many aspects are amenable to experimental investigations, and the simulations we have reported in Ball and Brindley (2014, 2015, 2016a, 2016b, 2017) in fact have provided recipes for experiments that are quite feasible in the hands of competent students and postdocs. Such experiments cannot of course prove anything about the origin of life, but they can demonstrate likely scenarios and guide our thinking on missing parts of the story.
The HP crucible hypothesis restores the full glory of hydrogen peroxide in life's long story: it was there in the beginning, and life—ancient and modern—cannot exist without it.
Footnotes
Acknowledgments
This work was partially funded by Australian Research Council Future Fellowship FT0991007 (R.B.). We thank the referees for their helpful and constructive comments, which have enabled us to produce a greatly improved article.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
Associate Editor: David Deamer
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.