
Abstract
Unambiguous detection of chemical and physical signatures of microbial life on Mars or other solar system bodies requires differentiation between signals produced by biotic and abiotic processes; instruments aimed at generalized in situ extant life detection would therefore increase the science return of a life-detection mission. Here, we investigate Bioelectrochemical Systems (BES) as a technique to measure microbial metabolism (which produces electrical current and redox changes) and distinguish between potential abiotic and biotic responses in environmental samples. Samples from inhabited niches should contain everything necessary to produce current, that is, catalysts (microorganisms) and fuel (nutrients). BES can also probe for inactive organisms in less energetically rich areas by adding a fuel to drive metabolism. A commercial potting soil and a Mars simulant soil were inoculated in the anodic chamber of microbial fuel cells, and current was monitored over time. Addition of a fuel (electron donor) source was tested for metabolic stimulation of endogenous microbes. Redox reactions between Mars simulant soil and the introduced electron donor (lactate) produced false-positive results, emphasizing the importance of careful interpretation of signals obtained. The addition of lactate to both soils resulted in enhanced biologically produced current, allowing stimulation and detection of dormant microbes. Our results demonstrate that BES provide an approach to detect metabolism in samples without prior knowledge of the organisms present, and that thorough electrochemical analyses and experimental design are necessary to determine if signals are biotic.
1. Introduction
The first attempt to detect microbial life and metabolism on Mars was through the biological experiments conducted on the Viking mission in 1976. These experiments focused on analysis of organic chemical signatures and metabolic activity determined through the consumption and transformation of introduced organic compounds (Brown et al., 1978; Klein, 1978; Levin and Straat, 1979). Results from these types of analyses must be interpreted with caution due to difficulties of distinguishing biotic chemical signatures from similar signatures produced by abiotic processes. The activity observed during the Viking biological experiments can likely be attributed to oxidizing agents such as perchlorate and its radiolysis products produced by the intense ionizing radiation on the martian surface (Quinn et al., 2013). The inconclusive biological experiment results combined with the absence of organics in the gas chromatography–mass spectrometry analysis of soil samples (Klein, 1978) are what drives the consensus view that no life was detected by the Viking mission. Since that time, very little has been attempted in terms of measuring metabolic activity, directly or indirectly in subsequent missions (Nealson and Conrad, 1999; Conrad and Nealson, 2001; Dorn et al., 2003). Habitability is often defined more broadly in a “follow the energy” approach that searches for useful redox couples in the environment (Chyba and Phillips, 2001; Johnson et al., 2003; Hand et al., 2007; Kounaves et al., 2009; Blake et al., 2012; Grotzinger et al., 2012; Hays, 2015), identifying likely inorganic and organic components that could be either redox reactants for, or waste products of, metabolism. The identification of habitable or formerly habitable environments on other worlds (Greenberg et al., 2000; Parkinson et al., 2007, 2008; Stoker et al., 2010; Grotzinger et al., 2014) has provided us with sites to start looking for life with a strategy that builds upon what we have learned from former missions (e.g., the Viking mission) but is also novel in its approach. To overcome this challenge and be capable of detecting life with a different origin than life on Earth, focus must be placed on designing instruments that are capable of detecting a biosignature that is a shared and distinguishable feature of all living organisms while being as non-Earth-centric as possible (Nealson and Conrad, 1999; Conrad and Nealson, 2001; Dorn et al., 2003). Metabolism is a universal characteristic of (earthly) life involving electron flow (redox) reactions among organic and/or inorganic substrates. All metabolic modes use the same basic processes of differential redox and electron transfer to convert energy from the environment into biologically functional energy in the form of adenosine triphosphate (ATP) (Demirel and Sandler, 2002; Mentel and Martin, 2008). This process involves oxidation of an electron donor (fuel), liberating electrons and protons. The electrons are eventually transferred to an electron acceptor (oxidant) reducing it in the process. The electron flow from an electron donor to an electron acceptor results in cations (usually protons) being transported across a cellular membrane to produce an electrochemical gradient that drives the synthesis of ATP, the energy currency used for all cellular processes.
Since metabolism always involves redox reactions, it can be detected through either chemical or electrical means. Changes in oxidation state in a reaction can be analyzed by using electrochemical techniques such as fuel cells, which measure electrical current produced by oxidation and reduction half-reactions occurring in separate reservoirs divided by a proton exchange membrane. In a fuel cell, the electrodes (where oxidation or reduction occurs) are often coated in a catalyst (e.g., platinum or other metals) to increase surface area and lower the energy barrier for the oxidation or reduction reaction. Bioelectrochemical Systems (BES) operate similarly to standard fuel cells, but instead of an inorganic metal catalyst in the anode compartment, microorganisms catalyze the oxidation reactions. In the anode compartment, microorganisms oxidize an electron donor, releasing both electrons and protons. These electrons are directly transferred to the anode and subsequently travel through an external conductor (e.g., a resistor) to reach the cathode compartment. Reduction reactions (e.g., reduction of O2 to H2O, anaerobic reduction of nitrite to nitrogen) occur at the cathode, utilizing the electrons and the protons that were released. When an external conductor is placed between the anode and cathode, current is produced, and an increase in electrical potential can be measured. The current produced is proportional to the oxidative processes occurring in the anode compartment, either from microbial metabolism or other redox reactions. The physiological process of transferring electrons directly to an electrode is known as extracellular electron transport (EET). Though once thought to be rare, EET capability has been found in a wide variety of environmental microbial communities (Holmes et al., 2004; Wrighton et al., 2010; Ishii et al., 2012), including those in extreme environments (Miller and Oremland, 2008; Rojas et al., 2017; Rowe et al., 2017), and is a biological process that should be considered when looking for life.
Electrochemical devices have been proposed as promising life-detection instruments capable of measuring altered chemistry and ionic properties caused by biological activity (Kounaves et al., 2002) and current produced by microbial metabolic activity (Abrevaya et al., 2015a, 2015b). Environmental samples from inhabited niches should contain all the components necessary to produce current flow, that is, catalysts (microorganisms), electron acceptors, and fuel (electron donors). BES can also probe for starving and/or dormant microorganisms by adding exogenous fuel to drive metabolism and/or growth. BES have been shown to successfully measure the potential generated by pure cultures of microorganisms (Abrevaya et al., 2010; Li et al., 2017) including photolithoautotrophs (Figueredo et al., 2015). Despite the success demonstrated in detecting metabolic activity in a wide variety of monocultures, BES have rarely been applied to measure metabolic activity in environmental samples. Power and current density values generated by a humus-rich soil were much larger than those obtained with a sterilized soil control (Abrevaya et al., 2010). To this end, we applied BES, specifically microbial fuel cells (MFCs), to environmental samples (a commercially available soil and a Mars simulant soil) that are less than generous in terms of the nutrients present to support life and the overall biomass present. We hypothesized that the untreated soil samples would produce greater current than sterilized controls, and that by providing an additional electron donor source (lactate) increased current density values would be observed by stimulating microbial metabolic activity.
In this paper, we demonstrate that current density values did not differ significantly between sterile and nonsterile soils in long-term MFC experiments. Enhanced current density was observed in nonsterile soils with the addition of lactate, demonstrating the increased sensitivity of BES by stimulating dormant/starving microbes. We also characterized the experimental soils electrochemically before and after sterilization procedures and show that the redox properties of soils are altered after autoclaving, emphasizing the importance of appropriate control measures. BES improve upon the detection of metabolism approach utilized by the Viking missions by looking for temporal patterns indicative of metabolism as well as identifying electrical signatures that demonstrate active metabolism rather than the chemical (waste) by-products of metabolism. The use of two different soil mixtures broadens the perspective on what electrochemical signals can be produced abiotically and perhaps biotically.
2. Materials and Methods
Bioelectrochemical Systems (BES) experiments were performed by using MFC devices that contain two separated reservoirs across a proton exchange membrane (Fig. 1). For MFC experiments, one reservoir contained the anode immersed in a volume of wet soil (soil and anolyte), and the other reservoir contained the cathode immersed in the catholyte solution (Table 1). MFC experiments either used unsterilized soil or sterilized soil for controls. Injections of a fuel (electron donor in the form of lactate) for microbial growth were added at periodic intervals. Voltage was measured, and current was calculated between the two electrodes to determine the presence and activity of microbial metabolism.
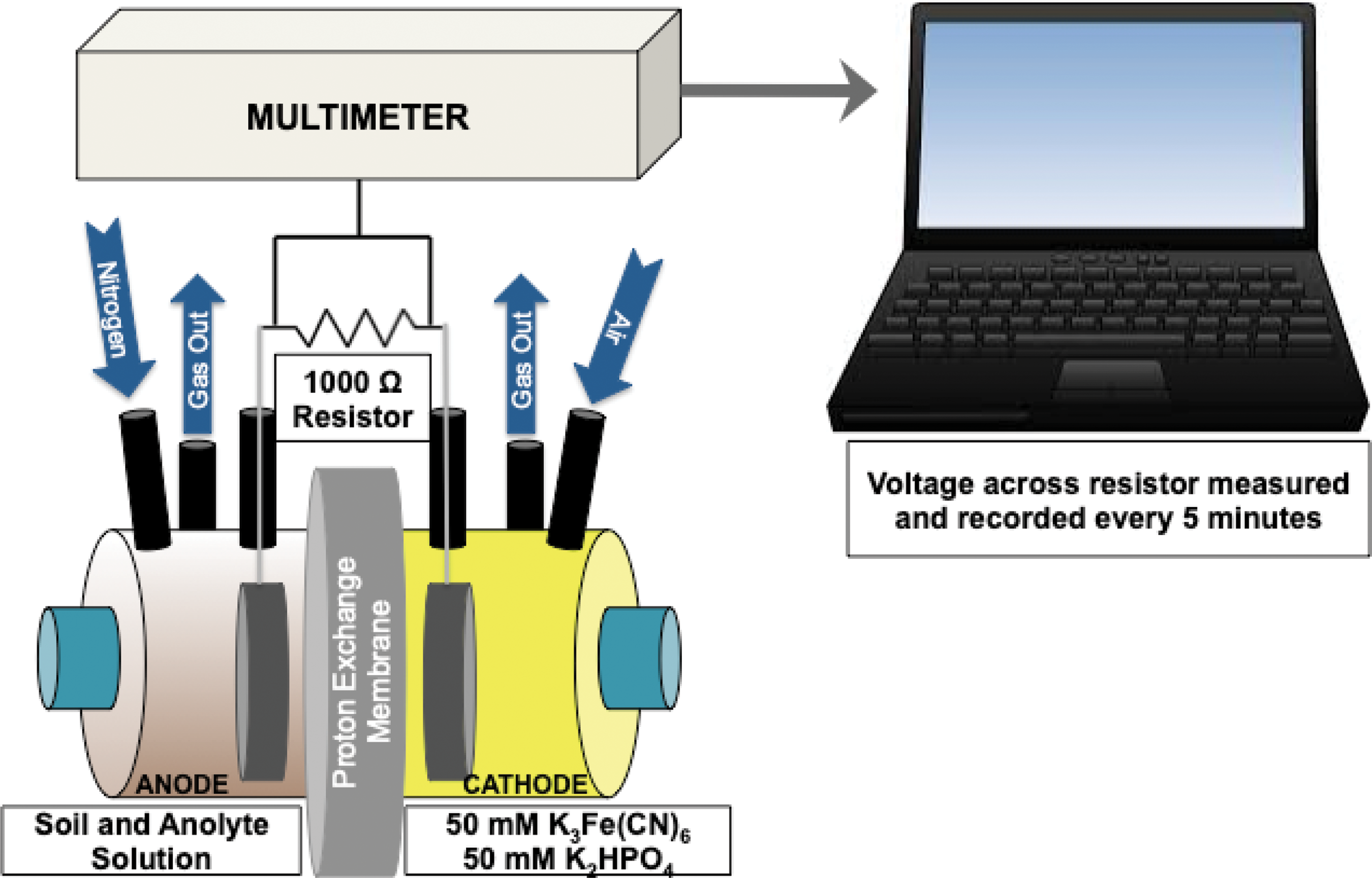
General schematic of the BES used. For MFC systems, the anode and cathode chambers were separated by a proton exchange membrane. Anode compartments contained the soil hydrated in anolyte solution while the cathode compartments contained the catholyte (50 mM K3Fe(CN)6, 50 mM K2HPO4).
Summary of Substrates Used in BES Experiments
2.1. Soil, anolyte, and catholyte preparation for BES experiments
The soil mix in the anode compartment contained either standard potting soil or Mars simulant soil. Standard potting soil mix (referred to here as HF, Happy Frog brand) was procured from FoxFarm Soil & Fertilizer Company, Samoa, California. The HF soil was chosen compared to other commercial potting soils because it omits standard presterilization procedures that eliminate the endemic microbial communities present in soils. Mars simulant soil (referred to here as MSS) was JSC-Mars-1A (M-JSC1A; Orbitec, Madison, WI), which is accepted as a spectral and physical simulant for testing various instruments and hardware in preparation for Mars missions (Chevrier et al., 2007; Hudson et al., 2007; Pommerol et al., 2009), though it is not an exact chemical analog. JSC-Mars-1A soil simulant has been characterized in detail (Allen et al., 1998), and it is sourced from the Pu'u Nene cinder cone in Hawaii. Soil samples were aliquoted into 20 g batches into sterile, acid-washed flasks, and some were sterilized by autoclaving with two cycles: one cycle at 121°C for 1 h and 45 min and a second cycle with the same conditions 24 h after the first cycle. Other samples of MSS and HF soil were not sterilized and were used as is. Soils (sterile and nonsterile) were hydrated up to 30 mL total volume with anolyte solutions. Sterile 0.6 M NaCl was used as the anolyte for all HF soil experiments. Sterile Mars simulant salt solution was used as the anolyte for all MSS experiments. The Mars simulant salt solution contained 1.2 mM Mg(ClO4)2, 1.8 mM MgSO4, 3.6 mM CaSO4, 0.3 mM KCl, 89.7 μM NaCl, 0.6 mM Na2CO3, and 20 mM CaCO3, and is derived from a mixture of salts detected at the Mars Phoenix mission landing site (Hecht et al., 2009). The sterile catholyte solution contained 50 mM potassium ferricyanide (K3Fe(CN)6) and 50 mM K2HPO4 (pH = 7.2). Potassium ferricyanide was chosen as a catholyte as it does not require O2 for cathodic reactions to proceed and would function in oxygen-free atmospheres as part of a Mars instrument. JSC-Mars-1A is a spectral analog rather than a chemical analog, so although basaltic soils do occur on Mars and some mineralogical properties may be consistent with the soil fraction of our MSS, other types of soils may be a better chemical match for different martian environments. The salt fraction of our MSS is based on the soluble chemical composition of martian regolith measured by the MECA instrument suite on the Phoenix lander, and was included to more realistically simulate the redox activity of one potential type of martian sample.
The fuel (electron donor) injection solution contained 50 mM lactate, 50 mM piperazine-N,N'-bis(2-ethanesulfonic acid) (PIPES) buffer and was prepared by using traditional anaerobic techniques (Miller and Wolin, 1974). To account for responses to fuel injection being due to the PIPES buffer and not the lactate (electron donor), 50 mM PIPES buffer solution was also prepared as a control for fuel addition experiments.
2.2. Bioelectrochemical Systems experiments
Microbial fuel cells (MFCs) consisted of dual-compartment 30 mL custom-designed glass chambers (Fig. 1). Anode and cathode chambers were separated by a pretreated Nafion 424 ion exchange membrane (DuPont, Wilmington, DE). Briefly, ion exchange membranes were soaked in 0.5% HCl and rinsed with ultrapure H2O, and subsequently boiled in 3% H2O2 and 0.5 M H2SO4 with ultrapure H2O wash steps in between and after each boiling treatment. All electrodes were made from 6 mm thick graphite felt (Electrolytica, Alpharetta, GA) cut into circles with a 39 mm diameter and treated as previously described (Hsu et al., 2012). Graphite felt was bonded to platinum wire (Alfa Aesar, Tewskbury, MA) with EPOX-4 graphite adhesive (Electrolytica, Alpharetta, GA). The anode chamber was continuously purged with filtered N2, and the cathode chamber was continuously purged with filtered air at a rate of 15 mL/min. Voltage across a 1000 Ω resistor was measured and recorded every 5 min by a high-impedance digital Keithley 2700 Integra Series multimeter (Keithley, Cleveland, OH). The multimeter utilized has a 0.1 μV resolution at the 100 mV range. Factoring in the resistance (1000 Ω) and surface area of the electrodes (20 cm2) of our MFC systems, a current resolution of 0.005 nA/cm2 was determined. For organic acid (electron donor) stimulation experiments, fuel cells were run for 24 h to allow stabilization and collection of baseline voltages. Lactate (3 mM final concentration) or PIPES buffer was injected into the anode compartments after 24 h, and two additional injections were performed at the 48 and 72 h mark. Current (I) was calculated according to I = V/R, and current density (j) was determined by dividing I by the surface area of the electrode (20 cm2). Open circuit potentials of the anode were determined by measuring the voltage difference between the anode electrodes and 1M Ag/AgCl reference electrodes (BASi, West Lafayette, IN) placed in the same anode compartment. Open circuit anode potentials were determined for both sterile and nonsterile soil substrates (HF and MSS) with the same anode and cathode compartment components as listed in Table 1. Voltage measurements were taken at 8 time points over a course of 120 h, and a lactate injection (3 mM final concentration) was conducted at 18 h.
2.3. Cyclic voltammetry of test substrates
Cyclic voltammetry (CV) tests were conducted on both hydrated soils (sterile and nonsterile) and anolyte solutions to observe electron transfer reactions (oxidation and reduction peaks) of the samples. Tests were performed in the same chambers as in the fuel cell experiments configured to have a port for a 1M KCl Ag/AgCl reference electrode (BASi, West Lafayette, IN). Graphite felt electrodes (as described above) served as the working electrode, and platinum wire was used as a counter electrode. The three-electrode chamber was continuously purged with filtered N2 at a rate of 15 mL/min. CV tests were performed with a Gamry Reference 600 potentiostat and Gamry Framework software (Gamry Instruments, Warminster, PA). Each CV analysis was conducted at a scan rate of 10 mV/s over a range of −1000 to 1000 mV vs. Ag/AgCl for 3 cycles.
2.4. Analysis of organic acids and proteins
Samples taken from the anode chambers were filtered through 0.2 μm Acrodisc Supor membrane syringe filters (Pall Corporation, Port Washington, NY) for organic acid analysis. Samples were stored at −80°C until analysis. To concentrate cells for total protein analysis, 2 mL aliquots of removed soil anolyte was centrifuged at 13,000 rpm for 5 min. Pellets were stored at −20°C until analysis. After fuel cell experiments were done, electrodes were sectioned in half to conduct both total proteins quantification and scanning electron microscopy (SEM).
Organic acid concentrations were measured as described previously (Bretschger et al., 2010) by using a high-performance liquid chromatograph (HPLC) with a Hi-Plex PLRP-S 300 mm × 7 mm column (Agilent Technologies, Santa Clara, CA). A fluorescence detector (Agilent Technologies, Santa Clara, CA) was used with an excitation wavelength of 450 nm and an emission wavelength of 520 nm. The HPLC was operated with 0.1 M sulfuric acid as the mobile phase (0.5 mL/min). To measure total proteins, pellets were hydrolyzed in 1 M NaOH at 50°C for 30 min with vigorous mixing by vortex at 10 min intervals. Electrode samples were hydrolyzed in 200 mM NaOH at 100°C for 90 min with vigorous mixing by vortex at 15 min intervals. Soluble protein in the extracts was determined spectrophotometrically using Folin phenol reagent (Lowry et al., 1951) with bovine serum albumin (BSA) as a standard.
2.5. Scanning electron microscopy sample preparation of electrode samples
Electrode samples were fixed in distillation purified electron microscopy grade 2.5% glutaraldehyde (Electron Microscopy Sciences, Hatfield, PA). Samples were then processed through an ethanol dehydration series (30, 50, 70, 80, 90, 95, 100% v:v ethanol, three 10 min intervals for each concentration) and critical point drying (Autosamdri 815 critical point drier, Tousimis Inc., Rockville, MD). Samples were mounted on aluminum stubs and coated with Au (Sputter Coater 108, Cressington Scientific, Watford, UK). Images were captured at 5 keV with a JOEL JSM 7100F low-vacuum field emission scanning electron microscope (JEOL USA, Inc., Pleasanton, CA).
3. Results
3.1. Cyclic voltammetry of test substrates
Cyclic voltammetry (CV) analysis was used to detect different oxidation and reduction reactions in the test substrates (soils and anolyte solutions) under the same anaerobic conditions utilized in MFC anode chambers. For both soils, commercial potting soil (HF) and Mars simulant soil (MSS), the sterilized soils showed much more activity in terms of reduction and oxidation reactions (Fig. 2). For the HF soil, there were no responses observed in the nonsterile soil, while the sterile soil exhibited substantial anodic (oxidative) current at +250 mV and cathodic (reductive) current at −250 mV. The MSS possessed a similar trend, with the sterile soil showing pronounced anodic current at +500 mV and cathodic current at −250 mV. These data suggest that the sterilization process of autoclaving results in the production of redox active species in both soils.

Cyclic voltammograms of the various experimental test substrates: (
3.2. Long-term baseline fuel cell tests
Endemic microbial metabolic activity of both soils was investigated with long-term fuel cell experiments. Sterile and nonsterile soils were hydrated and placed in a standard MFC setup (Fig. 1). Long-term fuel cell tests were run for 336 h after preliminary tests (Appendix Fig. A1) indicated a longer run time was necessary to allow for current density readings to stabilize. Nonsterile HF soil exhibited enhanced current density in comparison to sterile HF soil (Fig. 3A). The delineation between the sterile and nonsterile HF soil was seen after 80 h of incubation. With the long-term fuel cells tests for the MSS, the sterile MSS had a higher current density than the nonsterile MSS starting at 25 h (Fig. 3B). There was a large increase in current density seen in the sterile MSS that began at the 25 h mark, which started to decay after 30 h.
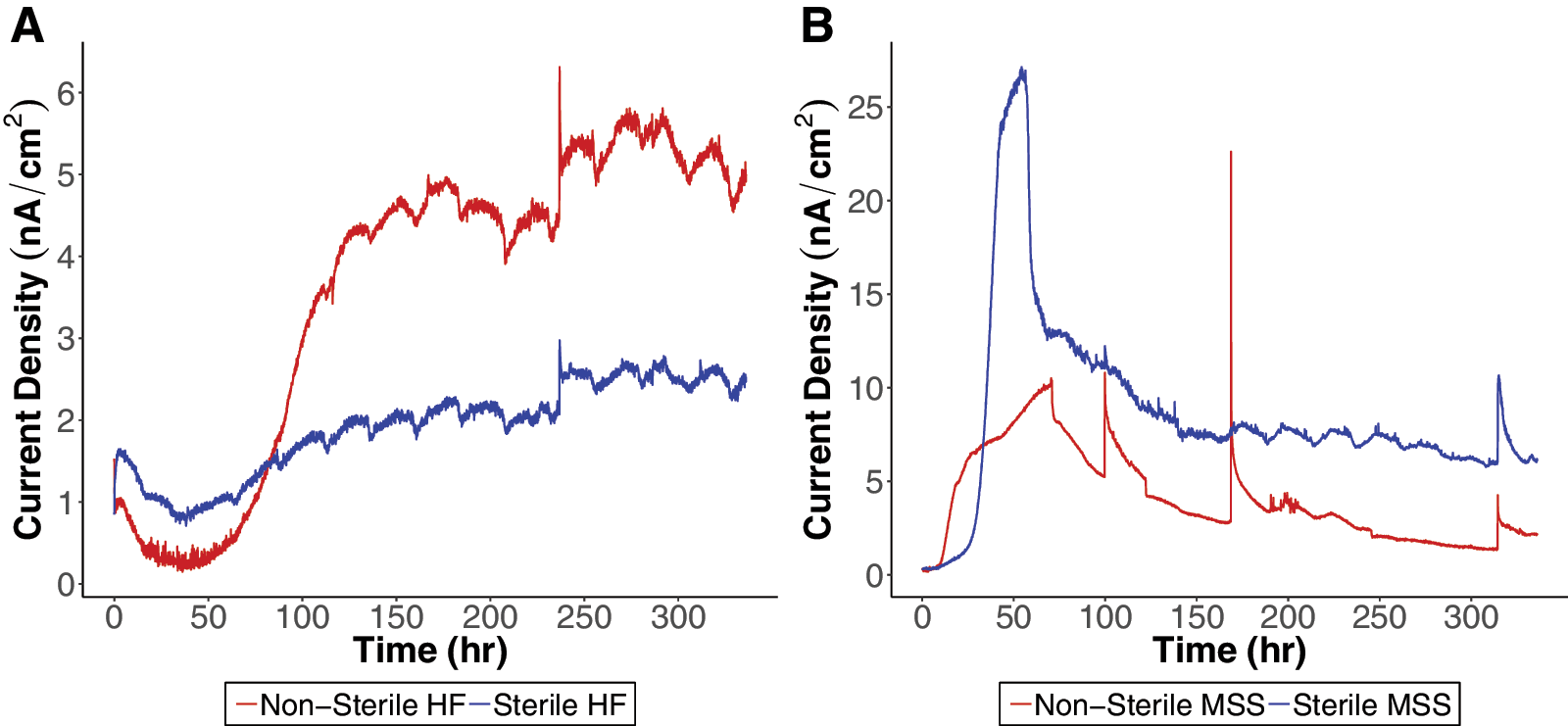
Long-term (2-week) baseline full cell tests conducted on (
3.3. Fuel cell tests with organic acid addition
Organic acid addition to both soils was explored as a way to enhance microbial metabolic activity and observe the subsequent responses in terms of current density. Lactate was chosen to serve as the organic acid as it is a common monovalent organic acid found in soils that is weakly adsorbed to the solid phase (Jones et al., 2003). In addition, lactate is a by-product of complex carbon substrate oxidation that can be utilized as an electron donor by many microbial groups, and has been shown to increase metal reduction rates of anaerobic microbial communities in situ (Brodie et al., 2006; Faybishenko et al., 2008; Burkhardt et al., 2010). Furthermore, 2.4 × 108 g of reduced carbon arrives on Mars each year through meteoric deposition (Hayatsu and Anders, 1981; Mullie and Reisse, 1987; Flynn, 1996), and this organic carbon is most likely to be oxidized to carboxylic acid derivatives such as lactate (Benner et al., 2000). CO and CO2 could also be reduced on catalytic iron minerals to form pyruvate at high pressure and temperatures (e.g., in ancient martian seas or martian subsurface), which could readily be reduced to form lactate in the presence of Fe2+ (Roldan et al., 2015; Camprubi et al., 2017). Lactate injections in replicate fuel cell experiments with HF soil led to a sustained and increasing current density in the nonsterile soil (Fig. 4A). The control experiments, nonsterile soil with PIPES addition and sterile soil with lactate addition, demonstrated a spike in current density with injections that decayed and was not sustained with time. Similar trends were also observed in the lactate addition experiments done on MSS, with the exception of the sterile MSS with lactate addition (Fig. 5). The nonsterile MSS had an increasing amount of current density with each lactate injection (Fig. 5A). The sterile MSS replicate experiments with lactate injections also showed a gradual increase in current, but the readings never reached the level of the nonsterile MSS. The nonsterile MSS experiments exhibited an increase in current density after the first PIPES injection, but the current dropped off during the time course of the rest of the experiments. The maximum stable level of average current obtained for the nonsterile MSS was 37.73 nA/cm2 and 19.71 nA/cm2 for the nonsterile HF soil.
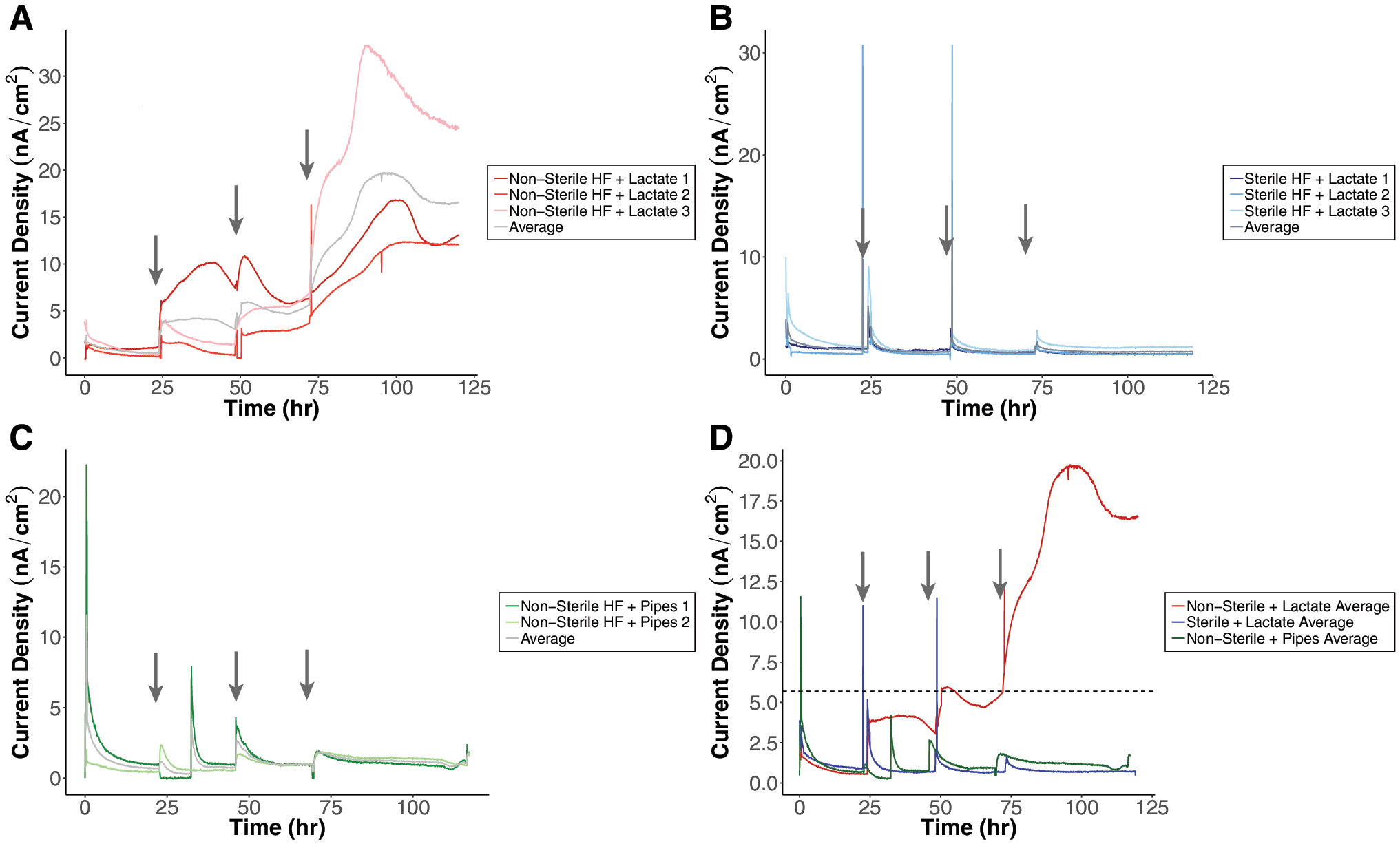
Fuel cell experiments with organic acid additions conducted on HF soil. Each arrow corresponds to either a 3 mM lactate or PIPES addition. (

Fuel cell experiments with organic acid additions conducted on MSS. Each arrow corresponds to either a 3 mM lactate or PIPES addition. (
3.4. Stable anode potentials in fuel cells
Since environmental samples and especially soils are naturally very heterogeneous, the microbial communities will vary between experiments, and no uniform and/or replicable anode potential may be measured across samples. However, we measured the anode potentials over 5 days in nonsterile and sterile HF and MSS to evaluate if anode potentials change due to the reducing power of the endogenous microbial communities. Open circuit anode potentials for fuel cells with sterile and nonsterile soils were monitored, and stable anode potentials in nonsterile soils were significantly more negative compared to sterile soil anode potentials (Fig. 6). After a lactate injection at 18 h, the potentials of the nonsterile soil anodes decreased while anode potentials in the sterile soils remained relatively stable. The nonsterile MSS had the most negative stable anode potential at -0.37 V (vs. Ag/AgCl), and the nonsterile HF soil reached a stable anode potential of -0.26 V (vs. Ag/AgCl). The sterile HF soil and sterile MSS anode potentials were much more positive (0.23 V and 0.20 V vs. Ag/AgCl, respectively). As differences in anode potentials in nonsterile and sterile soils were rapidly established and observed after one lactate injection, no further lactate injections were conducted.
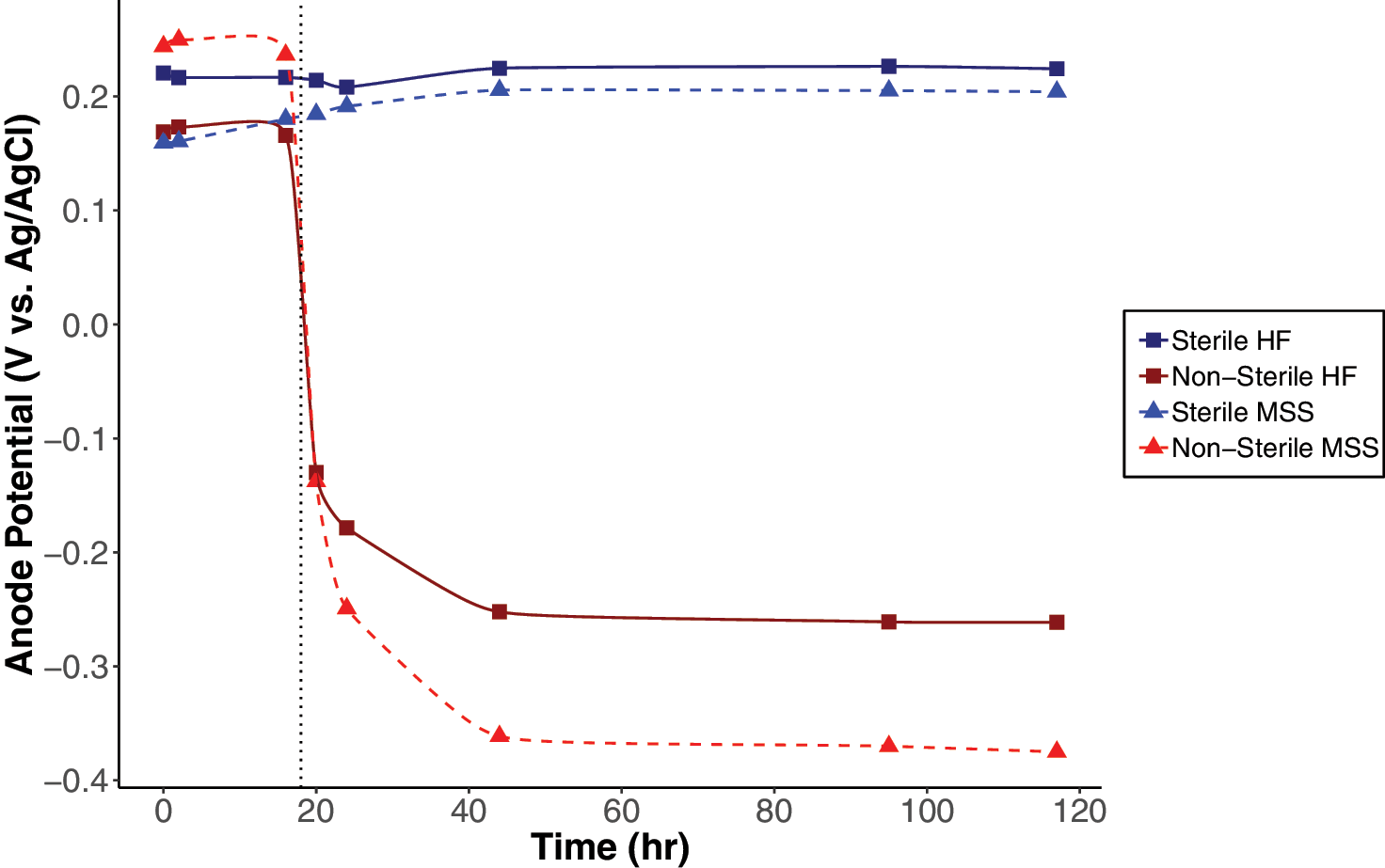
Open circuit anode potentials (vs. Ag/AgCl) measured with fuel cells containing nonsterile and sterile soils. A 3 mM lactate addition was done at 18 h indicated by the dotted line.
3.5. Analysis of organic acids and protein concentrations during fuel cell experiments with organic acid addition
Samples were taken during the time course of the organic acid addition fuel cell experiments to measure organic acid (lactate, acetate, and pyruvate) concentrations. Acetate and pyruvate were measured as they are both by-products of lactate oxidation. At each injection time point, 1.5 mL of a 50 mM lactate solution was injected into the anode compartment. Table 2 summarizes the measured organic acid concentrations for a representative set of the lactate addition experiments done on each soil. Accounting for all the lactate additions, a final concentration of ∼9 mM lactate would be expected if all of it remained in solution. For both soils, the sterile experiments showed an incremental increase in lactate concentrations. At the end of the experiments, 3.11 mM lactate was detected in the nonsterile HF soil, and 4.61 mM lactate was detected in the nonsterile MSS. An incremental increase in pyruvate was observed in the nonsterile HF soil experiment. For the nonsterile MSS experiment, acetate was only observed at the end of the experiment.
Concentrations of Lactate and Potential By-products from Lactate Oxidation, Acetate and Pyruvate, during the Time Course of Representative Fuel Cell Experiments with Lactate Addition
Time points T1, T2, and T3 occurred before ∼3 mM lactate injections. T4 was the final time point at experiment end. ND indicates not detected.
Total proteins were measured in samples taken from the anode compartment to evaluate changes in biomass during the time course of the organic acid addition experiments (Fig. 7). For the HF soil experiments, the initial protein concentrations across all experimental conditions were in a similar range. The sterile HF soil experiments displayed a decrease in protein concentrations with each subsequent sample, with the exception of the final time point that displayed a slightly elevated protein concentration. The nonsterile HF soil experiments with PIPES injections exhibited decreases in protein concentration during the experimental time course. In the nonsterile with lactate addition replicate experiments, protein concentrations remained constant or increased slightly during the initial time points until dropping off at the T3 time point. For the MSS experiments, protein concentrations in the nonsterile lactate addition experiments had markedly higher protein concentrations initially and throughout the time course of the experiments compared to the other conditions. In the nonsterile MSS PIPES addition replicate experiments, protein concentrations generally remained constant. With the sterile MSS lactate addition experiments, a decrease in protein concentrations was observed during the time course after an initial increase at the T1 time point. Protein concentrations from the electrodes were also measured to account for any biomass that was attached to the electrode. For both soils, the experimental conditions with lactate had similar values in protein concentrations, while the PIPES addition experiments had markedly less protein concentrations (Fig. 8). Protein samples taken from the anode compartment were generally higher in the MSS experiments compared to HF soil experiments (Fig. 7), while the reverse is true for the electrode protein concentrations (Fig. 8). Cell biomass attached to the electrodes was also evaluated with SEM imaging (Appendix Fig. A2). No dense biofilms were detected, although cells were observed attached to the electrodes from nonsterile soil experiments.

Average protein concentrations from soil anolyte samples taken during the time course (T0: experiment start to T4: experiment end) of replicate fuel cell with organic acid (lactate) addition experiments. The protein concentrations measured reflect any biomass in the planktonic phase in the soil anolyte. Error bars indicate standard deviations of triplicate samples.

Average protein concentration from electrodes harvested after replicate fuel cell with organic acid (lactate) addition experiments. The protein concentrations measured reflect any biomass that attached to the electrode. Error bars indicate standard deviations of replicate experiments.
4. Discussion
Bioelectrochemical Systems (BES) such as MFCs have been suggested to be a viable method for the in situ detection of microbial life (Abrevaya et al., 2010; Figueredo et al., 2015; Li et al., 2017). These studies utilized pure cultures of different organisms, including an archaeon (Abrevaya et al., 2010), eukaryote (Abrevaya et al., 2010), and photolithoautotrophs (Figueredo et al., 2015), grown under ideal growth conditions. The pure culture investigations of heterotrophs used rich media supplemented with carbon sources (glucose and/or yeast extract), and cultures were incubated at optimal growth temperatures (37°C) (Abrevaya et al., 2010; Li et al., 2017). The one application of BES to an environmental sample was with a humus-rich topsoil (Abrevaya et al., 2010). The detection of life will likely not occur in environments that are optimized for the growth of one organism or in environments that are replete in organics. Microorganisms found in the most oligotrophic subsurface environments on Earth are estimated to catabolize 104 to 106 times more slowly than organisms grown in the laboratory, with biomass turnover occurring at timescales of hundreds to thousands of years (Hoehler and Jørgensen, 2013). In our study, we chose to explore the efficacy of BES for life detection by using two different soils that were not altered by the addition of nutrients, including a Mars simulant soil. In long-term MFC tests, there was minimal differentiation between sterile and nonsterile HF soil, and high variability in the response of MSS (Fig. 3, Appendix Fig. A1). These results suggest simple hydration of the soils was not enough to stimulate metabolic activity and differentiate between electrochemical signals produced biotically or abiotically.
Many microorganisms in the natural environment are not actively metabolizing and actually exist in a state of dormancy. Ecosystems experience states of nutrient flux, and starvation due to nutrient limitation is a cue that regulates dormancy in natural communities (Braeken et al., 2006). Theoretically, benefits afforded by dormancy should be greater in environments with limited or low resource availability (Gardner et al., 2007). Soil environments can be especially heterogeneous, with changes in soil properties affecting water flow and nutrient cycling (Schlesinger et al., 1996). Metagenomic analysis of sequencing data from different environments revealed a high abundance of dormancy genes present in soil environments (Lennon and Jones, 2011). An effective life-detection strategy must be able to detect not only metabolically active life but potentially starving or dormant life as well. As part of our study, we investigated how the addition of an electron donor stimulated microbial activity of dormant populations and consequently could be detected by a MFC system. Nonsterile fuel cell experiments where no lactate (Fig. 3) was added looked very similar to sterile MSS experiments with lactate addition, indicating that redox reactions between the soil and organics can provide false-positive results (Fig. 5B). The sterile HF soil with lactate addition did not show the same response as the sterile MSS, suggesting that perhaps the salts present in the MSS salt solution may react with some of the lactate, causing the gradual increase in current density. When lactate was added to the nonsterile fuel cells, a large and unmistakable response occurred, demonstrating the usefulness of supplying an additional electron donor and not just relying on what is in the soil itself (Figs. 4D and 5D). The enhanced current density (four times the current observed in long-term tests) in both nonsterile soils highlights the reproducibility of microbial activity profiles, with increased current that is maintained over time. Additionally, differences between measured anode potentials in nonsterile and sterile soil fuel cells suggests microbial metabolic activity was a driver of producing reducing conditions/reactions altering nonsterile anodic potentials to more negative values over time. Sterile soil fuel cells had anodic potentials that remained relatively constant and did not deviate from baseline levels. Decreasing anode potentials tracked well with increasing overall voltage for nonsterile soils, particularly after the lactate injection at 18 h (data not shown). The use of lactate for metabolic activity is likely when compared to the controls; adding a buffer (PIPES) did not change the current density signal in nonsterile soils, and with the sterile soils no sustained current density was observed at the level of the nonsterile experiments with the addition of lactate. HPLC results also revealed an accumulation of lactate in the sterile soils compared to the nonsterile soils. Pyruvate and acetate were both measured in the nonsterile soil lactate addition experiments, and both organic acids are the by-products of lactate oxidation. Their presence indicates the potential biotic oxidation of lactate. These metabolic by-products of lactate oxidation could also be used by other organisms in the microbial community, stimulating further metabolic activity.
It is important to note that to be able to distinguish between abiotic and potentially biotic signals, BES experiments should run on longer timescales. The feasibility of long-term BES experiments in an in situ (Mars) context is facilitated by the fact power would only be needed to record potential measurements and maintain fuel cell temperature, both of which would be relatively lower power activities that should be able to proceed while the rover performs other operations. Potential can be continuously monitored after adding a salt solution to soil samples, and/or after the addition of substrates at determined intervals. If these experiments were only run for a few hours, it would be easy to misinterpret some of the current density results observed. In addition, the current density from abiotic reactions usually occurred in large spikes that decayed rapidly, while current density from metabolic activity exhibited a sustained response indicative of cellular activity and growth. These abiotic spikes often occurred when there was an increased rate of mixing by injecting test substrates into the anode chamber. The only consistent mixing in the MFC systems was contributed by the purging with N2 gas. Future investigations should be designed to employ a mode of constant mixing of both the anolyte and catholyte chambers. Evaporation of the anolytes could also disrupt the electrical signals detected as seen toward the end of the time course in Fig. 5B. Additionally, determining the appropriate controls for these experiments is complicated by the fact that standard sterilization through autoclaving seemed to change the electrochemical baseline of the soils (Fig. 2). CV tests on the soil substrates showed that oxidants were released upon sterilization of the soils, which can further complicate interpretation of the electrochemical signals observed in full cell tests. Electrochemical analysis of environmental samples with techniques such as CV is essential to understand baseline redox properties and how they may change after control treatments. Other methods of sterilization such as heat sterilization with a vacuum oven (Schuerger et al., 2012) or gamma irradiation (Berns et al., 2008) should be explored in future studies, as they may minimize impacts and alteration of the chemical composition of soils. The organic acid addition tests conducted with an organic acid and a control buffer demonstrated that this experimental design might be sufficient to distinguish between abiotic and biotic electrochemical signals. For future instrumentation, implementing these types of controls rather than heat sterilization may help simplify and reduce the overall required instrument payload.
Quantification of the proteins present in the soil anolyte solution and on the electrodes allowed us to approximate changes in biomass. Overall soil anolyte protein concentrations were higher in the nonsterile soil with lactate addition experiments. Nonsterile MSS with lactate anolyte protein concentrations was much higher than the concentrations for other conditions at all time points. Accordingly, the current density observed for the nonsterile MSS lactate addition experiments was also the highest achieved throughout the time course of the MFC experiments. Electrode protein concentrations were highest for both soils under nonsterile conditions with lactate addition, although concentrations were also high with the lactate additions to sterile soil experiments. The lack of biofilms evident in SEM images of the electrodes, along with the anolyte protein data, suggests that the planktonic community might have contributed the most to current density. Many soils are rich in humic acids that can serve as electron shuttles (Klüpfel et al., 2014). Electron shuttles such as humic acids are known to mediate electron transfer between microbes and insoluble electron acceptors/donors (Kappler et al., 2004; Wolf et al., 2009). It is of note that some microbes in the soil samples could have been nonmotile and attached to the soil particles. Soil biofilms confer many advantages including protection from variations in environmental conditions (Burmølle et al., 2012) and resistance to antibacterial compounds (Burmølle et al., 2006). Soil particle–associated biofilms may have been a factor in current density through endogenously produced shuttles or through use of naturally occurring shuttles present in the soil. We also explored introducing exogenous electron shuttles (riboflavin and flavin mononucleotide) into our MFC systems, but no enhancement in current density was observed (data not shown). Future investigation is needed to see if other potential electron shuttles (e.g., anthraquinone-2,6-disulfonate) might increase metabolic activity and subsequently electrochemical signal. Introduction of a known EET-capable microbe, Shewanella oneidensis MR-1, to sterilized soils was investigated to determine the relationship between electrochemical signal and cell number; however, the organism did not tolerate conditions provided by the different soil environments. Future experiments that characterize the microbial communities in the soils through metagenomic sequencing with BES implementation could give insight into changes in the microbial communities and the functions present contributing to anode reduction (electrochemical signal).
5. Conclusion
Bioelectrochemical Systems (BES) are a promising, novel approach to life detection that can be independent of the metabolic modes present in an environment and complementary to biochemical approaches. The appeal of BES comes from the ability to apply these systems to environmental samples without any prior knowledge of the existing microbes and metabolisms, making it ideal for life detection on other planetary bodies. In concert with habitability studies, BES experiments can also be designed to probe for specific metabolisms. Habitability studies might indicate what dominant electron donors or acceptors are present in the environment, but typically if life is using a particular substrate in its metabolism, concentrations should be changing and/or these compounds may be limiting and thus elude detection. For example, a site might have ample electron acceptors but no measurable electron donors. By determining what might be a likely or favorable electron donor in an environment, addition tests similar to our lactate experiments can be conducted to stimulate metabolic activity. Similar to an approach used by the Viking experiments, a mixture of electron donors could also be first utilized to assess for a positive response with subsequent testing of individual compounds. In addition, BES like a three-electrode chemical cell can potentially test various redox potentials (electron donors or acceptors) in an environment in situ. BES can be configured as a three-electrode system, where the working electrode can be poised at a biological redox potential of interest. The redox potential can be controlled so that the electrode can serve as either an electron donor or acceptor, and current at the working electrode can be measured and is indicative of active redox reactions. Future experiments should be designed to test the three-electrode system approach with different environmental samples. If a particular redox potential can be used for energy by life, current should be easily detected and measurable.
Footnotes
Acknowledgments
The authors would like to thank Katie Lee and Kelsey Shane Davis for their assistance in BES experiments, and Richard Quinn for suggesting the MSS anolyte solution. A portion of this research was carried out at the Jet Propulsion Laboratory, California Institute of Technology, under a contract with the National Aeronautics and Space Administration and funded through the internal Research and Technology Development program.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
A. Appendix
Associate Editor: Victor Parro