
Abstract
Carbonaceous meteorites provide clues with regard to prebiotic chemistry and the origin of life. Geological Survey of India recorded a carbonaceous chondrite meteorite fall in Mukundpura, India, on June 6, 2017. We conducted a study to investigate the microbial community that survived the meteorite impact. 16S rRNA metagenomic sequencing indicates the presence of Actinobacteria, Proteobacteria, and Acidobacteria in meteorite impact soil. Comparative phylogenetic analysis revealed an intriguing abundance of class Bacilli in the impact soil. Bacillus thermocopriae IR-1, a moderately thermotolerant organism, was isolated from a rock, impacted by the Mukundpura meteorite. We investigated the resilience of B. thermocopriae IR-1 to environmental stresses and impact shock in a Reddy shock tube. Bacillus thermocopriae IR-1 survived (28.82% survival) the effect of shock waves at a peak shock pressure of 300 kPa, temperature 400 K, and Mach number of 1.47. This investigation presents the first report on the effect of impact shock on B. thermocopriae IR-1. The study is also the first report on studying the microbial diversity and isolation of bacteria from impact crater soil immediately after meteorite impact event.
1. Introduction
Shock wave is a high-energy nonlinear wave that propagates at supersonic speed. Shock wave is an abrupt and virtually discontinuous process with an extreme change in pressure, temperature, and fluid field where entropy increases over a nearly small region. Sudden energy release during explosions, earthquakes, lightning, and meteorite/asteroid impacts dissipates chemical, electrical, mechanical, and nuclear energy, resulting in the formation of shock waves.
A meteorite impact on planetary surfaces causes a sudden increase in pressure and temperature and generates shock waves (Shimamura et al., 2016). A meteorite impact causes chemical interactions within the meteoritic material. Previous experimental and theoretical works have reported that these chemical interactions cause the production of volatiles, organic, and inorganic molecules in simulated impact conditions (Fegley et al., 1986; Mukhin et al., 1989; Sekine et al., 2003; Schaefer and Fegley, 2010; Furukawa et al., 2015; Shimamura et al., 2016). These impact events are the only known unique extraterrestrial event that disrupts an ecosystem in time and space (Cockell and Lee, 2007). The biology of impact craters is significant, and impact craters are considered to be sites of unique ecosystems (Cockell and Lee, 2007).
However, there are no reports on the effect of meteorite impacts on the microbial ecosystems in fresh meteorite fall sites. In the present investigation, we studied the microbial diversity that survived shock waves that were generated by an impact event immediately after the meteorite fall. Most studies conducted previously have reported the microbial diversity or organisms present in lakes or aqueous systems formed in meteorite impact craters years after the initial meteorite impact event. The diversity of bacteria in impact crater lakes such as Lonar lake, India (Paul et al., 2016); Tswaing impact crater, South Africa (Cockell et al., 2001); Crater lake, Oregon, USA (Page et al., 2004); and Haughton impact structure, Devon Island, Canadian High Arctic (Parnell et al., 2004) has been reported earlier. However, the effect of allochthonous or anthropogenic nutrient input and other external activities may have played a pivotal role in the development or change in microbial communities over time since the formation of these craters. It is plausible that the current microbial diversity may be different from the microorganisms present during the period of fresh impact. Furthermore, meteorite impacts can render an impact site sterile and cause a complete loss in biota in some cases (Cockell and Lee, 2007). Meteorite impacts also cause a profound disruption of extant microbial communities. These impact events create an extreme change in the ecological balance, which may include dramatic shifts in biological evolution manifested in the fossil records as extinction boundaries. Hence, it is imperative to study the effect of a meteorite impact on microbial communities present in fresh impact craters.
Geological Survey of India (GSI) recorded a carbonaceous chondrite (CC) meteorite fall in the Mukundpura area of Rajasthan, India, on June 6, 2017. In this investigation, we isolated Bacillus thermocopriae IB-5, Brevibacillus borstenlensis IB1, IB3, and IM3 from the Mukundpura meteorite impact crater soil, and B. thermocopriae IR-1 was isolated from a rock impacted by the meteorite. The impact crater soil was collected immediately within 48 h of the meteorite fall (fresh fall) and processed for culturing the microbial flora. Twenty different bacterial strains were isolated from impact crater soil and identified by using morphological, physiological, and biochemical characteristics as well as 16S rRNA gene sequencing. To the best of our knowledge, this study represents the first report on isolation of B. thermocopriae and B. borstenlensis from meteorite impact crater soil.
The microbial community structure of the fresh impact soil in comparison with nonimpact samples was also studied with conventional culturable methods as well as a metagenomic approach. The metagenomic study indicates a statistically significant difference in the microbial community structure in the impact soil and nonimpact soil (NIS). The meteorite impact causes the generation of shock waves in the soil. To assess the resistance of organism that survived the impact event, we exposed B. thermocopriae IR-1 that survived the Mukundpura meteorite impact to impact shock waves in a Reddy shock tube. A Reddy shock tube is routinely used to study the applications of shock waves in aerospace engineering and research (Jagadeesh et al., 2011). We report herewith a novel application of a Reddy shock tube in Astrobiology to simulate meteorite impact shock in a controlled environment and to investigate the effect of these supersonic waves on bacteria for probing implications in lithopanspermia. This study is presumably the first report on the study of surviving microbial communities from fresh impact crater soil and assessment of the resilience of the bacterial impact survivors to controlled simulated meteorite shock wave impact conditions in a Reddy tube.
2. Materials and Methods
2.1. Sampling site and sample collection
A meteorite fall was observed in Mukundpura village (26°52′52.5′N, 75°39′53.7E″) in Bhankrota, Jaipur, Rajasthan, India, on June 6, 2017, at 0515 h IST. The meteorite fall and its characterization were recorded by GSI (
The meteorite had a density of 2300 kg/m3 and it created a circular crater ∼43 cm wide and ∼15 cm deep (Fig. 1A). The Mukundpura meteorite was classified to be a rare fall of CC with typical chondrule granules (Fig. 1B). We visited the site and collected the soil sample from the impact crater pit in triplicate (IS1, IS2, and IS3) in sterile containers. A control soil sample representing NIS was collected ∼100 m away from the impact crater (NIS). The soil was analyzed for pH, sodium, potassium, magnesium, chloride, and salinity as described earlier (Greenberg, 2005; Thombre et al., 2016). Few pieces of fragmented meteorites and rocks were found interspersed with the soil sample. The meteorite samples were collected and used for characterization and detection of amino acids as described earlier (Glavin et al., 2006). The rock sample (Fig. 1C) was probed for the presence of rock colonizing bacteria surviving the impact.

2.2. Isolation and identification of bacteria from meteorite impact crater
Bacteria from the impact crater were isolated by an enrichment culture method. In brief, 1 g soil sample (IS1, IS2, IS3, and NIS) was enriched in 100 mL nutrient broth and incubated at 37°C and 55°C at 100 rev min−1 for 24–48 h until growth appeared. Similarly, for isolation of bacterium from rock collected from impact crater, 1 g crushed rock powder was added to 100 mL nutrient medium incubated as mentioned until growth was observed (Thombre et al., 2017a). After appropriate incubation, a loopful of the enrichment broth was isolated on nutrient agar (NA) and observed for bacterial growth.
Bacterial colonies showing distinctive morphologies were subcultured on NA slants and used for microscopic, physiological, and biochemical identification. Cell morphology was observed by phase contrast microscopy and light microscopy at 1000 × magnification. Endospore formation, gram reaction, and motility were assessed with a commercially available kit (HiMedia, India), according to the manufacturer's instructions.
The biochemical analyses of the bacteria isolated from the Mukundpura meteorite impact crater soil and rock were performed as outlined in Bergey's Manual of Determinative Bacteriology (Holt, 1994). The DNA for 16S rRNA gene sequencing was extracted by using the phenol–chloroform method (Sambrook and Russell, 2001), sequencing was performed by using Sangers method as described earlier (Thombre and Oke, 2015), and identification of the sequences was performed with EzBiocloud as described by Yoon et al. (2017).
2.3. DNA extraction and amplification of 16S rRNA genes for metagenomic sequencing
We extracted the total DNA from soil collected from the Mukundpura meteorite impact crater using the PureLink® Genomic DNA extraction kit (Invitrogen™) according to the manufacturer's instructions. The extracted DNA was checked for purity of nucleic acids with a NanoDrop™ 2000 spectrophotometer (ThermoFisher Scientific) and quantified with Qubit 2.0 Fluorometer (Invitrogen).
Amplification of the microbial 16S rRNA gene fragment (V3–V4 region) was performed with V3–V4 specific primers 314F-5′CCTACGGGAGGCAGCAG3′ and 518R-5′ATTACCGCGGCTGCTGG 3′. A quantity of 10 ng of total DNA was used as a template for polymerase chain reaction (PCR). The PCR conditions were 98°C for 30 s followed by 30 cycles of 98°C for 10 s, 72°C for 30 s, extension at 72°C for 5 s followed by a final hold at 4°C. The PCR product was separated on 2% Agarose gel and visualized with SYBR® Safe DNA gel stain (10 μL/100 mL).
The desired amplicon DNA was purified by using PureLink® Quick Gel Extraction kit (Invitrogen). A second PCR was performed with a set of primers that had Illumina indexed bar code sequences. The PCR Master mix contained 2 μL each of 10 pmol/μL forward and reverse primers, 1 μL of 40 mM dNTP, 10 μL of 5 × Phusion® HF reaction buffers (New England BioLabs, Inc.), 0.4 μL of 2 U/μL F-540 Special Phusion® Hot Start DNA Polymerase (Thermo Fisher Scientific), 10 μL (minimum 5 ng) of amplicon from the previous PCR cycle, and water to make up the total volume to 50 μL.
The amplicons were purified by using PureLink® PCR Spin columns (Invitrogen) and used for library preparation. The library was validated on a 2200 Tapestation® system (Agilent Technologies) and sequenced by paired-end sequencing (2 × 150 bp paired end run) on Illumina MiSeq platform utilizing a 300 cycle Illumina MiSeq reagent kit v.2. as described previously (Caporaso et al., 2010). The raw metagenomic data sets from this study were deposited in NCBI's Sequence Read Archive under BioProject accession no. PRJNA454556 (Submission ID: SUB3969020) with NCBI Accession nos. SRR7155920, SRR7244759, SRR7245280, and SRR7244752 for IS1, IS2, IS3, and NIS, respectively.
2.4. Analysis of metagenomes, annotation of unculturable microorganisms
The raw sequencing reads generated by sequencing of 16S rRNA amplicons were trimmed, and the adapter sequences were removed by using PERL. The sequences were trimmed by quality with Phred score ≥30 (>Q30; error probability ≥0.001). The consensus V3 region sequence was constructed by using FLASH program with default parameter settings (Magoc and Salzberg, 2011). The chimeras were removed by the de-novo chimera removal method, UCHIME algorithm was implemented in the USEARCH package (Edgar et al., 2011). Preprocessed reads from all samples were pooled and clustered into OTUs based on their sequence similarity using UClust program (similarity cutoff = 0.97).
The sequence data were analyzed with QIIME (Quantitative Insights Into Microbial Ecology software package) (Caporaso et al., 2010). Representative sequence was identified for each OTU and aligned against SILVA core set of sequences with PyNAST program (DeSantis et al., 2006; Quast et al., 2013). The taxonomy classification was performed by using Ribosomal Database Project (RDP) classifier 2.2 (Wang et al., 2007) against SILVA 16S rRNA genes (Quast et al., 2013) and Greengenes database (DeSantis et al., 2006).
The taxonomy assignment of phylum, class, order, family, genus, and species distribution was done for each sample based on OTU. Taxa other than the top 10 were categorized as “Others,” whereas the sequences that did not have any alignment against taxonomic database were categorized as “Unknown.” The heat map and taxonomy abundance plots were created with QIIME. SILVA was used for obtaining V3 sequences, taxonomic annotations, and sample wise OTU. A Biological Observation Matrix (BIOM) file was generated for various analyses (McDonald et al., 2012) by using QIIME, and the rare and abundant species were identified from the samples (Galand et al., 2009; Aravindraja et al., 2013).
The genera-level taxonomic statistical analysis of metagenomic sequences was studied with the Statistical Analysis of Metagenomic Profiles (STAMP) v 2.0 software package. Corrected p-values were calculated based on Fisher's exact test method using Storey's false discovery rate method of multiple test correction within STAMP, and the p-values with <0.05 were considered for comparison (Storey and Tibshirani, 2003; Parks and Beiko, 2010; Tang et al., 2013). The microbial diversity within the samples was calculated with Shannon, Chao1, and observed species metrics using QIIME v1.7
2.5. Effect of salinity, temperature, and gravity stress on B. thermocopriae IR-1
The effect of temperature, salinity, and hypergravity stress on growth of B. thermocopriae IR-1 was investigated as described earlier (Thombre and Oke, 2015; Thombre et al., 2016; Thombre et al., 2017a). In brief for temperature stress, log phase culture of B. thermocopriae IR-1 was inoculated in nutrient broth and incubated at a range of temperatures (10°C, 28°C, 37°C, 45°C, 55°C, 60°C, and 70°C), and growth was observed after 24 h. The effect of salinity stress was studied by observing growth in nutrient broth supplemented with different NaCl concentrations (0%, 1%, 3%, 5%, 8%, 10%, and 15%).
For studying the effect of hypergravity on B. thermocopriae IR-1, log phase cells were subjected to hyperacceleration by centrifugal rotation in a centrifuge (Tarson, India) as mentioned earlier (Thombre et al., 2017a). The culture was inoculated in nutrient broth (absorbance at 600 nm = 0.15) in a 1.5 mL microfuge tube and was subjected to centrifugation to achieve hypergravity values of 503 g and 894 g, respectively.
Control consisted of incubation of the culture at static conditions (1 g) (Thombre et al., 2017b). After every 24 h, one microfuge tube was withdrawn, and growth was measured in terms of absorbance at 600 nm (Thombre et al., 2016c) with a UV-Vis spectrophotometer (UV-1800, Shimadzu, Japan). The experimental observations were recorded in triplicate, and the experiment was repeated twice to obtain statistically significant readings.
2.6. Preparation of bacterial cells, spores, and coupons for impact shock experiments
The effect of impact shock on B. thermocopriae IR-1 isolated from rock collected from the Mukundpura meteorite impact crater was studied. The spore suspension was prepared by the method described by Vaishampayan et al. (2012b) with slight modifications. In brief, a single pure colony of IR-1 was inoculated in 1:100 diluted nutrient broth, incubated for 1–2 days, and examined for sporulation by microscopy.
After appropriate incubation, the spore suspension was heat shocked at 80–90°C, and the spore count was estimated using Thoma counter and the spread plate method as described by Roy et al. (2015). Ten microliters of spore suspension and vegetative cell suspension of B. thermocopriae IR-1 was impregnated on sterile stainless steel circular coupons of 5 mm diameter. The steel coupons were sterilized by autoclaving before use. After seeding, the coupons were dried overnight in sterile conditions and then stored individually in sterile microfuge tubes for further use.
2.7. Experimental setup for effect of impact shock on bacteria
A simple, yet novel, methodology was used to study the effect of impact shock on bacteria by using a hand-operated piston-driven single diaphragm Reddy shock tube (Reddy and Sharath, 2013). The shock waves were generated with peak pressures of 1.8–3 bar (180–300 kPa), temperature between 400 and 600 K, and Shock Mach number of around 1.47 for an exposure time of about 250–300 μs. A sudden expulsion of air, generated by way of a piston, was used to rupture the diaphragm with a 31 mm inner diameter that separated the high-pressure (driver section) and low-pressure section (driven section), thus generating a shock wave.
Two piezoelectric sensors (PRESSURE SENSOR ABS AXIAL 6-SIP, MPX5700ASX-ND-NXP Semiconductors, USA, Inc.) of high response value were used to measure the peak pressure and shock speed. With this equipment, the generated shock wave propagated at a speed of 0.5 km/s and impacted the bacteria and spores impregnated on the coupon, which was placed inside the driven section for 250 μs (Supplementary Video S1). After the impact, we collected the coupons and performed recovery of cells by immersing the coupons in 500 μL sterile saline. After recovery, the spore count and viable cell count were estimated by spreading 100 μL of 10-fold serially diluted recovered samples on NA plate. The medium was incubated at 55°C for 24 h, and the total viable count and spore count were recorded as colony forming unit per mL and used for calculation of percentage survival. All experiments were performed in triplicates.
3. Results
3.1. Sampling site and sample collection
According to GSI, the Mukundpura meteorite created an impact crater about 15 cm deep and 40–43 cm in diameter in a dry alluvial field. The elemental analysis of the soil indicated that the concentration of Na, Mg, K, Ca, and Cl was 0.079, 1.668, 2.88, 3.64, and 0.064 g/L, respectively. The trend in the concentration of ions was found to be Ca>K>Mg>Na>Cl. The soil had a neutral pH (7.5–7.8), and it was dry and sandy. The rock sample (IR = imapct rock) collected from the impact crater was identified as quartzite (Fig. 1C). Quartzite is a hard metamorphic rock commonly found in Rajasthan, India. The meteorites collected with impact soil were confirmed to be CC by GSI.
High-resolution liquid chromatography mass spectroscopy (HR-LC-MS) estimated the amino acid content in the meteorite, and the following amino acids were detected: aspartic acid (237.116 pmol/L), glutamic acid (224.207 pmol/L), serine (154.818 pmol/L), glycine (342.339 pmol/L), threonine (74.015 pmol/L), arginine (101.590 pmol/L), alanine (296.670 pmol/L), valine (59.867 pmol/L), methionine (108.972 pmol/L), phenylalanine (48.200 pmol/L), isoleucine (29.281 pmol/L), leucine (129.318 pmol/L), and lysine (75.238 pmol/L).
3.2. Isolation and identification of bacteria from meteorite impact crater
We used the impact crater soil for culturing heterotrophic aerobic bacteria. Out of the 20 predominant bacterial strains present in the Mukundpura meteorite impact crater soil, 6 bacterial isolates exhibited distinct colonies capable of growth at elevated temperatures (55–60°C). We selected these isolates for identification using biochemical tests and 16S rRNA gene sequencing. Interestingly, B. thermocopriae and Brevibacillus borstelensis were dominant in impact soil, whereas Bacillus subtilis, Bacillus cereus, and Bacillus licheniformis were predominant in NIS (Table 1 and Supplementary Fig. S1; see Supplementary Data at
Identification of Microorganisms Isolated from Mukundpura Meteorite Impact Crater Soil and Nonimpact Soil Using 16S rRNA Gene Sequencing
Bacillus thermocopriae IR-1 was isolated from a quartzite rock collected from the impact crater (Fig. 2). This organism demonstrated resistance to higher temperature and salinity compared with other isolates. Strain IR-1 showed typical morphology and phenotypic features of genus Bacillus. IR-1 was gram reaction positive, aerobic, motile, rod shaped, and capable of producing central endospores. Colonies were light, smooth, raised, and showed growth on NA after 24 h incubation at 55°C. The temperature range for growth was 37–60°C, with optimum growth at 55°C. pH range for growth was pH 7.0–9, with optimum growth at pH 7. NaCl concentration for growth was 0–10% (w/v), with optimum growth at 0–1%.

Construction of phylogenetic tree based on 16S rRNA gene sequencing by neighbor-joining method. The evolutionary distances were computed using the Tamura 3-parameter method and are in the units of the number of base substitutions per site. Evolutionary analyses were conducted in MEGA6.
Cells were positive for catalase and reduction of nitrates to nitrites, but they were negative for oxidase test (Supplementary Table S1). We present herewith the first report on isolation of B. thermocopriae IR-1 (NCBI GenBank Accession no.: MH246940) and B. borstelensis IB-1 (NCBI GenBank Accession no.: MH287036) from a meteorite impact crater.
3.3. Effect of salinity, temperature, and gravity stress on B. thermocopriae IR-1
We investigated the effect of salinity, temperature, and hypergravity on the growth of B. thermocopriae IR-1. The results indicate that the organism demonstrated growth in medium supplemented up to 10% w/v sodium chloride at 55°C. The organism could also survive in altered gravity conditions in hypergravity values of 503 and 894 g. B. thermocopriae IR-1 was hence a unique moderately halotolerant thermotolerant spore-forming bacterium that survived the impact event of the Mukundpura meteorite fall. This multiresistant organism thus has potential future application in studies related to understanding adaptation to an extreme space-like environment that limits the principal possibilities of life
3.4. Experimental setup for effect of impact shock on bacteria
We investigated the effect of shock waves on the strain IR-1 in a Reddy shock tube (Supplementary Table S2). The schematic representation of a Reddy shock tube is outlined in Fig. 3A. It consists of a driver section and a driven section that is separated by a diaphragm. The pressure at the driver section is increased by way of a piston until the diaphragm ruptures (which is designed to rupture at a desired higher pressure). This produces a shock wave that impacts the microbial cells impregnated on coupons placed at the end of the driven section. We measured the peak pressure and shock speed using two piezoelectric sensors of high response value. We used the voltage output of an oscilloscope (Fig. 3B) for the calculation of peak pressures. A peak pressure of 300 kPa (3 bar) was impacted on the microbial specimen lasting for about 250 μs. We calculated the Mach number as 1.47 and velocity of shock waves by time of flight method (v = x/t) using the trigger of two sensors placed 15 cm apart as 500 m/s. The Rankine–Hugoniot jump equations for normal shocks were used for the calculation of peak temperature using experimentally calculated Mach number value (Eqs. 1 and 2).

Effect of impact shock on bacteria.
The temperature jump in terms of Mach number due to primary shock wave was calculated using Equation 1:
The temperature jump in terms of Mach number due to reflected shock wave was calculated using Equation 2:
where T 5 → temperature jump of the system due to reflected shock wave,
T 3 → temperature jump of the system due to primary incident shock wave, and
γ → the ratio of specific heats (Cp and Cv) for a particular gas. γ = 1.4 for air.
The results indicate that, in the current experiment, shock waves were generated 1.47 times faster than the local speed of sound in air (Mach number 1.47) with a velocity of 500 m/s. We impacted B. thermocopriae IR-1 at different peak pressures, such as 1.8 bar (180 kPa), 2.5 bar (250 kPa), and 3 bar (300 kPa) at 400 K for 250 μs (Fig. 3C). The control organisms used in this study were the gram positive extremophilic organism Kocuria rosea PRL-1, which was isolated from Himalayan quartzite rock and Staphylococcus aureus MCC 2408 (ATCC 25923). Unexpectedly, we observed complete inhibition of K. rosea PRL-1 and S. aureus when impacted with shock waves. However, IR-1 spores as well as vegetative cells persisted until 3 bar peak pressure.
The inhibitory effects of shock waves on bacteria such as S. aureus, E. coli, Pseudomonas and Mycobacterium tuberculosis, Salmonella typhimurium, and biofilms were described by Divya et al. (2011) and Gnanadhas et al. (2015). Although spores are considered more resistant to environmental stresses, we found that there was no significant difference in the viability of B. thermocopriae IR-1 spores (percentage survival of 18.18) and vegetative cells (percentage survival of 28.82). Hence, the vegetative cells of IR-1 were used in the further experiments for comparison with vegetative cells of K. rosea PRL-1 and S. aureus. The vegetative cells of B. thermocopriae IR-1 survived the impact shock with a percentage survival of 55% at 1.8 bar (180 kPa) peak pressure and a survival percentage of 28.82% at 3 bar (300 kPa) peak pressure (Fig. 3C).
3.5. Analysis of metagenomes, annotation of unculturable microorganisms
We observed the taxonomic complexity of microbial communities in soil from the Mukundpura meteorite impact crater by subjecting the 16S rRNA gene sequences to taxonomic classification using RDP classifier against SILVA 16S rRNA genes database. The 16S rRNA gene sequences of the Mukundpura meteorite impact metagenome that could not be affiliated to any known bacterial class by the RDP classifier were classified as “unknown.” The results of phylogenetic analysis of the16S rRNA gene sequences from the Mukundpura meteorite impact soil (IS1, IS2, and IS3) and NIS are shown in Fig. 4 and Supplementary Dataset S1. The relative phylum abundance of metagenomes of IS1, IS2, IS3, and NIS indicates that Proteobacteria are less in soil affected by impact in comparison with NIS (Fig. 4A). Similarly, the abundance heatmaps at species level (Fig. 4C) depict that the most abundant taxon in impact soil belongs to phyla Proteobacteria and Actinobacteria.
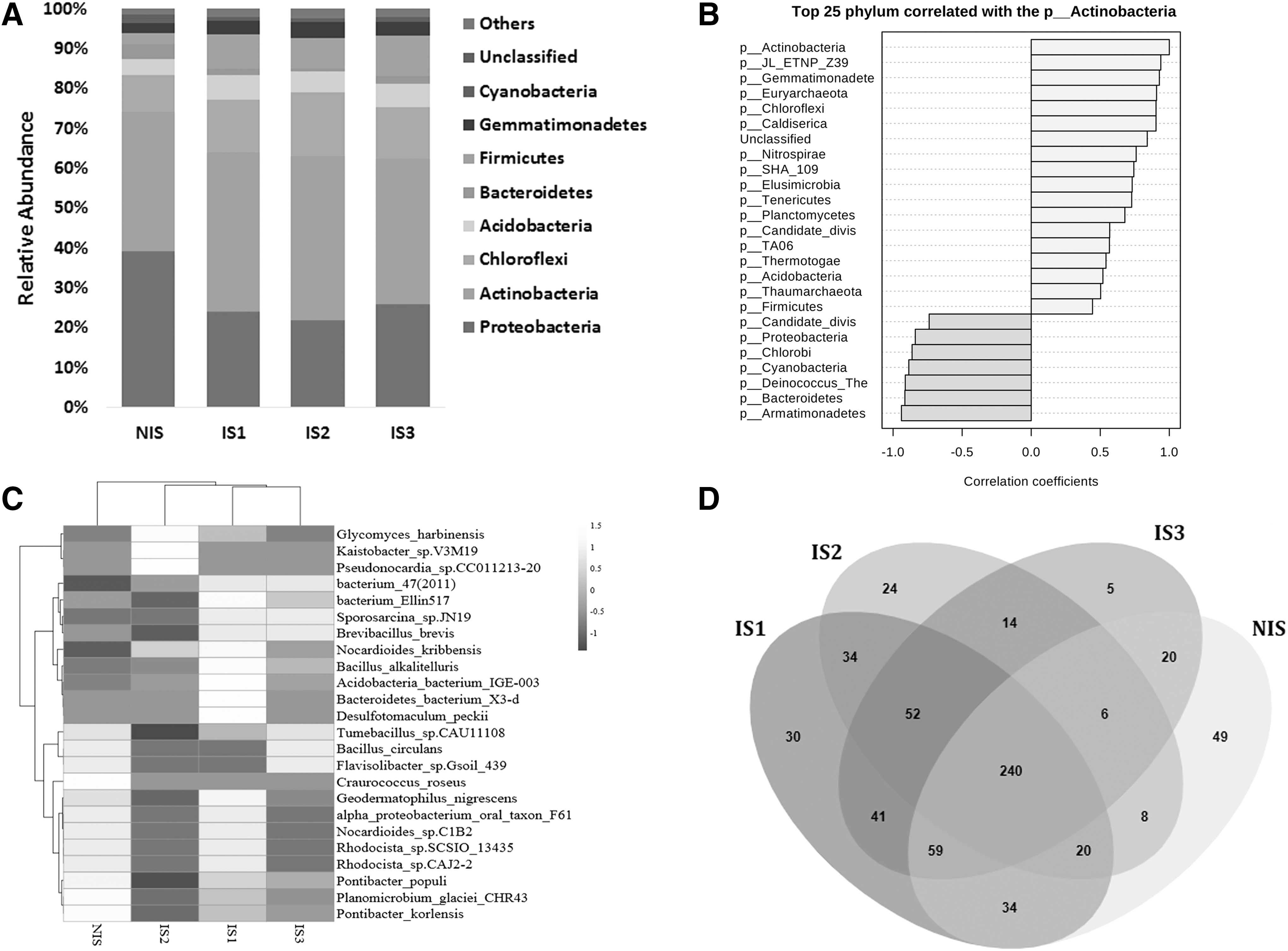
A circular lineage view of the tree of life was constructed with MLTreeMap as shown in Supplementary Fig. S2. The tree represents all the major groups of lineages assigned with different color representations, and the maximum likelihood phylogeny was constructed from nearly full-length 16S rRNA gene sequences. The Krona chart (Supplementary Fig. S3) depicts the relative abundance of microbial diversity from phylum to species level. Krona is an interactive metagenomic data visualization tool that helps in display of hierarchical data with their relative abundances (Ondov et al., 2011). The Shannon's diversity index was used to describe the species diversity, and Chao1 index was used to estimate total species richness in the microbial community of the Mukundpura meteorite impact metagenome (Supplementary Fig. S5A). The Shannon index for NIS, IS1, IS2, and IS3 samples was 8.11, 8.74, 8.64, and 8.58, respectively. The Boxplots (Supplementary Fig. S5B) represent the difference in the diversity measures between the metagenome of NIS and impact soil (IS1, IS2, and IS3). A nonmetric multidimensional scaling coordinate map was generated with a Bray–Curtis distance similarity matrix by measuring pairwise taxonomic profiles of NIS (green dot), IS1 (red dot), IS2 (blue dot), and IS3 (gray dot) samples (Supplementary Fig. S5C). The Beta diversity phylogeny tree was generated based on unweighted UniFrac distances calculated with QIIME v1.9.1 software (Supplementary Fig. S5D).
4. Discussion
Asteroid, meteorite, and comet impact events are the only extraterrestrial mechanisms that generate shock waves on Earth. Meteorite and asteroid impact events have been linked to the origin of life and extinction of biological life on Earth. Fossils, stromatolites, and carbon isotope fragmentation studies suggest that life may have originated around 3.5–3.8 Gya. This period coincides with the bombardment of planets of the inner solar system with remaining debris and planetesimals causing impact craters on martian, lunar, and terran surfaces (Chyba and Sagan, 1992).
It has been suggested that extraterrestrial impactors may have contributed to the organic molecules that directly accrue to qualify as the primal Earth's prebiotic inventories for the origin of life. Although the hypothesis of extraterrestrial implications in the origin of life has garnered a great deal of interest, the underlying principles involved are, to date, speculative. Interplanetary dust, meteorites, and comets are well-known sources of organic material of extraterrestrial origin, all of which could have served as sources of building blocks on primordial Earth that contributed to the evolution of cellular life (Chyba and Sagan, 1992). CCs are rare primitive meteorites that contain amino acids, organic matter, purines, pyrimidines, etc. that may have contributed to the synthesis of macromolecules (Pizzarello et al., 2006).
The Mukundpura meteorite is a rare CC fall as CCs are primordial extraterrestrial rocks that contain presolar grains from different stellar bodies. CCs preserve their chemical and mineral properties that they initially had since the formation of the Solar System. Hence, the Mukundpura meteorite may be useful to our understanding of the origin of the solar nebula, the accretion process of planetesimals, and other asteroidal evolution processes (McSween, 1979; Cronin et al., 1988; Zinner et al., 1989; Septhon, 2002). The detection of 13 different proteinic amino acids in the Mukundpura meteorite is an exciting finding as the organic matter of meteoritic origin is essentially surmised to have originated in interstellar medium or the outer Solar System (Alexander et al., 2007). Similar extraterrestrial amino acids have been reported in other CCs such as the Murchison (CM2) meteorite (Kvenvolden et al., 1970; Cronin et al., 1979; Glavin et al., 2006).
The rare fall of Mukundpura chondrite caused the formation of a rather small impact crater about 15 cm deep. The impact event may have caused significant destruction and change in the microbial community of the soil. We collected soil samples from the crater to investigate the surviving microorganisms that may have been present in the site that was affected by the meteorite impact. It was interesting that B. thermocopriae was isolated from the impact soil as well as from the impact hit rock. The isolation of B. thermocopriae that survived the meteorite impact event indicates the resilience of this hardy organism. At present, no study has reported on microbial population immediately after an actual meteorite impact.
Most studies report the microbial diversity in impact lakes, pits, or craters that have evolved over time due to meteorite impact events that have occurred in the past. A few examples of such meteorite impact crater studies conducted previously include Lonar lake, India (Paul et al., 2016); the Tswaing impact crater, South Africa (Cockell et al., 2001); Crater lake, Oregon, USA (Page et al., 2004); and the Haughton impact structure, Devon Island, Canadian High Arctic (Parnell et al., 2004). These studies provide valuable insights for understanding the environmental constraints that may have selected a particular microbial community in the extreme impact lake (Paul et al., 2016). However, no information is available on a microbial community that may have survived, or was present during, an actual impact event as already mentioned. Fresh meteorite impact craters can, thus, be explored as potential sites with which to gain insights into the effect of natural impact events on indigenous microbial communities.
To the best of our knowledge, this is the first study conducted to probe the microbial community of a fresh impact crater site using metagenomic sequencing. The metagenomic studies indicate the prevalence of Bacilli in impact soil in comparison with the nonimpact sample. Meteorite impacts are known to drastically diminish the biota of an impact site. Surprisingly, the soil metagenome of the impact site in this study had significantly higher microbial richness and diversity in comparison with the NIS metagenome (Supplementay Fig. S5). However, when a culture-based method was used, the total viable count of bacteria in the Mukundpura meteorite impact soil was much less (10−2) in comparison with the total viable count of NIS (105–107). Furthermore, the only organism that could be cultured from the impact site was B. thermocopriae, whereas the organisms from a nonimpact site were normal soil flora such as B. subtilis, B. cereus, and B. licheniformis. This indicates the subtle drawbacks of culturable and nonculturable methods of studying microbial diversity.
It is known that metagenomics can be used to study the presence of unculturable or unknown taxa from a particular ecosystem. However, this method reports the total microbial flora and may overestimate the diversity due to its inability to preclude nonviable cells (Vaishampayan et al., 2012a; Mahnert et al., 2015; Weinmaier et al., 2015). This has highlighted the importance of using different approaches, including live–dead staining, flow cytometry, and advanced methods of molecular biology for assessment of viable communities (Boulus et al., 1999; Ben-Amor et al., 2005; Vaishampayan et al., 2012a).
Nonviable communities in the metagenome can be differentiated by using propidium monoazide (PMA) treatment before the DNA extraction. The application of PMA in distinguishing viable cells from nonviable cells has been reported for food, water, and other environmental samples (Nocker et al., 2006; Cawthorn and Witthuhn, 2008; Vesper et al., 2008; Lin et al., 2011). PMA is a DNA intercalating agent that binds to DNA after photoactivation. The DNA from nonviable cells still persists in the environment and may contribute to the overestimation of microbial population in a metagenomic study (Nocker et al., 2006; Rogers et al., 2008; Vaishampayan et al., 2012a). In this study, the metagenomic diversity of impact soil appears higher in comparison with NIS. Thus, further studies using a PMA treatment of such samples are warranted to eliminate the nonviable organisms and report only the surviving bacterial lineages.
The isolation of B. thermocopriae IR-1 from impact rock was a significant outcome as this spore-forming organism could be one of the hardiest microbial representatives with which to conduct astrobiological studies related to impact survival, panspermia, lithopanspermia, and interplanetary transport. The spores of the genera Bacillus are widely accepted as one of the hardiest microbial representatives that can withstand extreme environmental and space-related parameters (Horneck et al., 2008). Hence, these spores have been previously reported for testing survivability in space or ground-based simulations or for studying lithopanspermia (Horneck, 1993; Nicholson et al., 2000; Horneck et al., 2001b; Benardini et al., 2003; Burchell et al., 2004; Nicholson et al., 2005; Vaishampayan et al., 2012b; Khodadad et al., 2017).
An attempt to investigate the effect of impact shock waves on the survival of B. thermocopriae IR-1 was conducted using a Reddy shock tube (Fig. 3). The spores, as well as vegetative cells of B. thermocopriae IR-1, survived the impact of shock waves with Mach number 1.47 at a peak pressure of 300 kPa (bar) and hypervelocity of 0.5 km/s. The results of the shock recovery experiments on bacterial cells at a peak shock pressure of 300 kPa and a peak shock temperature of about 400 K demonstrate that a substantial fraction of cells (up to 10−3) survived the stress. The physical appearance of the shocked stainless steel coupon remained intact up to 2.5 bar pressure, but the appearance changed at 3 bar pressure. The coupons lost their silver metallic luster and turned dark brown at 300 kPa (3 bar).
The scanning electron micrographs (Supplementary Fig. S4) indicate that shocked coupons developed snowflake-shaped fractal patterns, while the surface of unshocked disks remained intact. In both cases (shocked and control), the morphology of the bacterium remained intact, and slender bacillary forms of the bacterium were observed. Survival of B. thermocopriae IR-1 cells decreased with an increase in shock peak pressure (Fig. 3c).
Different methods have been previously utilized to explore the effect of impact shock, impact ejecta, and shock pressures on bacterial spores. Horneck et al. (2001a) studied the effect of planar shock waves (32 GPa) on spores of B. subtilis placed between quartz plates. Nicholson et al. (2006) impacted Al projectiles at 5.4 km/s into granite targets loaded with B. subtilis spores. Similarly, experimental simulation lithopanspermia was studied by using the launch by spallation at the Ames Vertical Gun Range (AVGR) (Fajardo-Cavazos et al., 2009). Burchell et al. (2001, 2003, 2004) impacted porous ceramic projectiles that were infused with microorganisms. The high explosive and air gun plane wave impact techniques are conventional methods with which to study the effect of impacts (Horneck et al., 2008). In this method, a plane metal plate is accelerated to velocities of 0.5–2.6 km/s that impacts a metal vessel that contains microorganisms between two thin disks of rock (Müller and Hornemann, 1969; Arnold, 1988; Horneck et al., 2008).
In this investigation, a unique method was devised by using a Reddy shock tube to study the impact of shock waves on microbial specimens. A Reddy shock tube is routinely used to study the applications of shock waves in aerospace engineering and research (Jagadeesh et al., 2011). Besides aerospace engineering, this device has now found applications in noninvasive therapy in lithotripsy, wound healing, treatment of avascular necrosis, vaccination, bacterial transformation, biofilm inhibition, and drug delivery (Ludwig et al., 2001; Kuo et al., 2009; Taylor et al., 2010; Divya et al., 2011; Jagadeesh et al., 2011; Gnanadhas et al., 2015). The shock waves in a Reddy shock tube are generated by rupture of the thin paper diaphragm by way of a piston. Hence, shock waves at 1.47 Mach number impact the microbial specimen impregnated on a stainless steel coupon in the driver section. The experimental setup has advantages in that it is simple, cost-effective, and safe as it does not use any dangerous projectiles, explosives, guns, or pistols for the generation of impact shock. Similar impact shock experiments can be conducted by embedding the microorganism in rocks for assessing the potential of interplanetary transfer and experimental validation of lithopanspermia (Horneck et al., 2008).
5. Conclusion
Overall, this study demonstrated a paradigm shift in our understanding of the astrobiological implications of meteorite fall events from a unique perspective. This study shows the potential application of impact ejecta sites as a valuable resource with which to investigate the effects of natural impact shock on indigenous microbial communities. Furthermore, it also provides a unique niche site for isolation of a survival fraction of microorganisms. These survivors may be potential microbial representatives that will aid in the study of the effects of space-related stress and interplanetary travel. Finally, as the quest for a full understanding of the origin of life and limits of organismic survival is still ongoing, these meteorite impact craters provide rare and unique sites for planetary exploration studies related to astrobiology and evidence of life (Cabrol et al., 1999; Newsom et al., 2001; Rathbun and Squyres, 2002).
Footnotes
Acknowledgments
This research was funded by Indian Space Research Organization (ISRO)–Space Technology Cell-University of Pune (STC-UoP), Grant no. 168, India, to R.T. Part of the research described in this publication was carried out at the Jet Propulsion Laboratory, California Institute of Technology, under a contract with the National Aeronautics and Space Administration. We thank Principal, Modern College, Shivajinagar, Pune, for providing necessary facilities for the work. We thank the Director, Physical Research Laboratory, Ahmedabad, India, for shock tube facility. SAIF-IIT Bombay, India, are acknowledged for LC-MS analysis. We thank Dr. Rajadurai CP for his inputs in data analysis.
Ethical Approval
This article does not contain any studies conducted with human participants or animals performed by any of the authors.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
Associate Editor: Petra Rettberg
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.