
Abstract
As part of the Biology and Mars Experiment (BIOMEX; ILSRA 2009-0834), samples of the lichen Circinaria gyrosa were placed on the exposure platform EXPOSE-R2, on the International Space Station (ISS) and exposed to space and to a Mars-simulated environment for 18 months (2014–2016) to study: (1) resistance to space and Mars-like conditions and (2) biomarkers for use in future space missions (Exo-Mars). When the experiment returned (June 2016), initial analysis showed rapid recovery of photosystem II activity in the samples exposed exclusively to space vacuum and a Mars-like atmosphere. Significantly reduced recovery levels were observed in Sun-exposed samples, and electron and fluorescence microscopy (transmission electron microscope and field emission scanning electron microscope) data indicated that this was attributable to the combined effects of space radiation and space vacuum, as unirradiated samples exhibited less marked morphological changes compared with Sun-exposed samples. Polymerase chain reaction analyses confirmed that there was DNA damage in lichen exposed to harsh space and Mars-like environmental conditions, with ultraviolet radiation combined with space vacuum causing the most damage. These findings contribute to the characterization of space- and Mars-resistant organisms that are relevant to Mars habitability.
1. Introduction
Exploration of the Solar System, as detailed in the AstRoMap European Astrobiology Roadmap (Horneck et al., 2016), encompasses several research areas that include “Life and Habitability” and “Biosignatures Facilitating Life Detection.” Space platforms and laboratories, such as the EXPOSE facility installed outside the International Space Station (ISS), have been essential for the simulation of planetary environments other than Earth, and these are crucial tools for improving robotic and human interplanetary exploration; for example, of Space, Mars, and Titan (Cottin et al., 2008).
Exploration of the limits of terrestrial life in space, focusing on the survival capacity of extremophile organisms in space, is relevant to understanding the effects of the space environment on unprotected biological and chemical material, and on the degradation of organic molecules, especially biomarkers. Several space missions have been performed in Low Earth Orbit (LEO), on satellites and on board the ISS, to study the short- and long-term survival capacity of different extremophile organisms exposed to space and Mars-like conditions (Sancho et al., 2007; Jönsson et al., 2008; de la Torre et al., 2010; Onofri et al., 2012; Wassmann et al., 2012).
For our experiment, which is one of the Biology and Mars Experiment (BIOMEX) (Kappen and Valladares, 2003: de Vera et al., 2019) on the European Space Agency (ESA) EXPOSE-R2 mission, the lichen species Circinaria gyrosa was exposed to both real space- and Mars-like conditions. Many lichen species are regarded as extremophiles in terms of temperature, radiation, and desiccation survival (de la Torre et al., 2007; Green et al., 2011). Some lichens are remarkably resistant to non-terrestrial conditions, a characteristic that is attributed to a range of morphological adaptations (Meeßen et al., 2013), a set of protective secondary compounds (Meeßen et al., 2014), and their ability to pass into anhydrobiosis (an ametabolic state) when desiccated (Ertl, 1951; Crowe et al., 1992; Kranner et al., 2005, 2008; Green et al., 2011). Lichen species have been selected for space experiments, because they are able to colonize terrestrial habitats exposed to high-intensity solar terrestrial radiation (Hughes and Lawley, 2003; Stivaletta and Barbieri, 2009), survive long periods of desiccation, and tolerate wide temperature variation (Sancho et al., 2008; Büdel et al., 2009). Most lichens possess inherent protection mechanisms, for example, the production of secondary metabolites in the uppermost layer of the entire lichen thallus that function as pigments and act as light filters (Gauslaa and Ustvedt, 2003; Solhaug et al., 2003; Gauslaa and Solhaug, 2004; Nybakken et al., 2004; Stivaletta and Barbieri, 2009). As lichens are symbiotic associations comprising a fungus (mycobiont) and a photoautotrophic partner (photobiont), and the photosynthetic capacity of photobionts is crucial for the nutrition of both symbionts (Jahns, 2008), impairment of photosynthetic activity is used to measure the viability of lichen after flight exposure (de la Torre et al., 2010). However, previous experiments have shown a parallel recovery of both symbionts after space exposure (Raggio et al., 2011). The first space exposure experiment with lichens documented the parallel recovery of both symbionts post-mission (Raggio et al., 2011).
In the first experiment with lichens on the ISS, the LIchens and Fungi Experiment (LIFE) (Onofri et al., 2012), samples of the lichen species, Xanthoria elegans and Rhizocarpon geographicum, were exposed to space and Mars-like conditions (Brandt et al., 2015). One lichen species, X. elegans, exhibited high viability after LIVE/DEAD® staining (72% for the photobiont and 82% for the mycobiont) yet a significant reduction in photosynthetic activity (<10%) occurred in response to the environmental space parameters of LEO (extra-terrestrial ultraviolet [UV]-radiation, space vacuum, temperatures [range, −21.7°C to +43°C], and galactic cosmic radiation doses up to ∼300 mGy) (Berger et al., 2012, 2015; Onofri et al., 2012; Rabbow et al., 2012; Brandt et al., 2015). Exposure to the Mars surface simulation conditions, that is, Mars-like UV radiation with wavelengths >200 nm and Mars-like atmosphere, also showed a high viability after LIVE/DEAD staining (79% for the photobiont and 89% for the mycobiont), and the metabolic activity again showed only 10% of the original photosystem II (PSII) activity values. Interestingly, the surviving cells eventually regained 99% of their photosynthetic capability (Brandt et al., 2015), which greatly exceeded the post-flight viability of any of the other organisms tested in the experiment.
The lichen species C. gyrosa, which was previously included in the space exposure experiment LITHOPANSPERMIA, also exhibited remarkable post-exposure viability (de la Torre et al., 2010; Raggio et al., 2011) and demonstrated a conspicuous resistance to space conditions. This high resistance to space exposure was also confirmed in later experiments that simulated conditions on Mars (Sánchez et al., 2012, 2014; de la Torre Noetzel et al., 2018). The combination of high capacity for resistance to space conditions and to a simulated Mars environment led us to propose that C. gyrosa be considered a suitable astrobiological model organism.
C. gyrosa was also used in the recent STARLIFE campaign to study the effects of space-relevant ionizing radiation. For the first time, the survival potential of C. gyrosa was studied as a function of increasing irradiation doses of accelerated helium ions, iron ions, and X-rays. No significant effects were observed on its levels of photosynthetic activity as measured by epifluorescence. In contrast, γ-irradiation elicited significant dose-correlated effects, as revealed by these techniques (de la Torre et al., 2017). These data were corroborated by an additional experiment as part of the STARLIFE project that focused on the C. gyrosa photobiont isolated from the lichen thallus and other aposymbiotic algae/photobionts (Meeßen et al., 2017).
A series of experiments performed or done by the German Aerospace Center (DLR) in Mars simulation chambers and on the ISS have demonstrated that some organisms (Lichens and strains of Chroococcidiopsis sp.) can survive Mars surface conditions, photosynthesizing and metabolizing at low levels in the presence of very limited water (Billi et al., 2011; Onofri et al., 2012; de Vera et al., 2014; de la Torre Noetzel et al., 2018). Other studies, using the Antarctic lichen, Pleopsidium chlorophanum, under Mars-simulated conditions (de Vera et al., 2014), demonstrated that some lichens and cyanobacteria are likely able to adapt to Mars conditions when partially shaded from UV light in niche conditions, probably as a result of adaptive strategies that have developed in their natural habitats in Antarctica, where they are photosynthetically active and growing (Kappen, 2000).
To contribute to a deeper understanding of the resistance of lichens to the environmental parameters at space conditions (UV-radiation and cosmic radiation, vacuum) and at Mars-like conditions (UV-radiation, atmospheric pressure and composition), we exposed the C. gyrosa in parallel to both actual flight conditions and simulated flight conditions in the laboratory during (this was done after the flight, due to the exact simulation of the space conditions at the mission) the BIOMEX experimental campaign (I would say “the BIOMEX space exposure”); then, we evaluated the following: (1) photosynthetic activity was determined by analysis of chlorophyll a fluorescence, to document the maximum quantum yield of PSII; (2) post-flight metabolic activity was documented with confocal laser scanning microscopy; (3) morphological changes were evaluated with field emission scanning electron microscopy and transmission electron microscopy; and (4) DNA damage sustained in the model lichen was documented with the use of a new approach (vs. the comet assay) that involved single-gene amplification via DNA polymerase chain reaction (PCR) and random amplification of polymorphic DNA (RAPD).
2. Materials and Methods
2.1. Biological samples
The vagrant lichen species, C. gyrosa (Sohrabi, 2012), was collected from clay soil in the steppic highlands of Central Spain (Zaorejas, 40°44.691′N, 02°11.109′W, 1293 m a.s.l.), which are characterized by extreme insolation, high-temperature contrasts, and arid summers (Crespo and Barreno, 1978). Vagrant or erratic lichens, living unattached to the substrate, are well known in the continental deserts and arid areas of Middle Asia, Eurasia, North America, and Northern Africa. C. gyrosa typically develops a globular, coralloid fruticose form and compact thallus, up to 2.5 cm diameter, made up of numerous dichotomous or sympodial branching (Fig. 1). In its top branches, C. gyrosa has almost circular pseudocyphellae (structures in lichens that appear as tiny pores or white regions on the outer surface of the lichen organism). Pseudocyphellae develop when there is a break in the cortex of the lichen, and the medullary hyphae extend to the surface (Sancho et al., 2000; Meeßen et al., 2013). Homogeneous thalli with diameters of ∼10 mm were selected for analyses (Fig. 1). As is typical for this lichen species, none of our samples developed fruiting (sexual reproduction) bodies, which eliminated the possibility of cultivating the mycobiont from spores both before and after the flight.

Field photo of the thallus of the vagrant lichen Circinaria gyrosa at the time of collection from Zaorejas (Guadalajara, Central Spain). Color images are available online.
2.2 Sample preparation
Before integration, dried samples of C. gyrosa were sent to the DLR where pellets of the original substrate and substrates of phyllosilicatic martian regolith = early acidic (P-MRS) and sulfatic martian regolith = late basic (S-MRS), which simulate the type of regolith that formed during the Noachian and Hesperian/Amazonian epochs, respectively (Böttger et al., 2012), were prepared. The Mars Regolith Simulants consist of different igneous rock mineral mixtures with a composition similar to that of Mars meteorites (i.e., gabbro, olivine, gypsum, goethite, quartz, and anhydrous iron oxide hematite, the only thermodynamically stable iron oxide under present-day Mars conditions). The Mars-simulated materials were crushed to simulate the size fraction of the martian regolith (Böttger et al., 2012).
A total of 24 pellets were prepared (12 of the original clayey substrates the lichens grew on before collection) and 12 (6 each of S-MRS and S-PRS pellets) of the simulated Mars regolith were prepared. The lichen samples were fixed to the pellets with the space-approved two-component Siliconkautschuk glue RTV-S 691 (Wacker Chemie GmbH) prepared according to the manufacturer's recommendation. The sample pellets were split into three sets of eight pellets (a flight set [FS], a ground-based set [Mission ground reference, MGR], and a laboratory control [LC] set), each of which contained four lichen samples fixed to pellets of the martian regoliths (two to P-MRS and two to S-PRS) and four lichen samples fixed to their original substrate. Each sample set was distributed to identical trays that were designed to fit into the carrier plates of the EXPOSE-R2 mission (de Vera et al., 2019) (Fig. 2A). The cells (type 3 hardware holes) of the trays that hold the sample pellets were 14 mm in diameter. For the flight experiment, the samples were split between a top tray, which was exposed to the UV of space, and a bottom tray, which served as a dark control (Fig. 2B, C). To simulate space-like or Mars-like conditions, half of the pellets for each sample set were either covered with MgF2 windows, which are transparent to the complete spectrum of extraterrestrial solar UV radiation (λ > 110 nm) and simulate space exposure (including vacuum) or covered with quartz windows, which are transparent to wavelengths >200 nm and simulate Mars-like conditions. A Mars-like atmosphere (provided by Praxair Deutschland GmbH), composed of 95.55% CO2, 2.70% N2, 1.60% Ar, 0.15% O2, and ∼370 ppm H2O, was injected into the air space of the pellets covered with the quartz windows of >200 nm to obtain a final atmospheric pressure of 980 Pa.

EXPOSE R2 facility with C. gyrosa flight samples at the International Space Station
2.3. Pre- and post-flight analyses of biological samples
2.3.1. Chlorophyll a fluorescence analysis
Chlorophyll a fluorescence was measured to establish pre-exposure and post-flight viability of the C. gyrosa thalli (diameters ∼10 mm and dry weights of 0.060–0.152 g). The pre-flight fluorescence measurements were made before pellet integration into the EXPOSE R2 flight carrier trays. To determine the PSII activity of lichen photobionts after flight, samples of C. gyrosa were reactivated (Sancho et al., 2007) as soon as they arrived at the laboratory in Madrid. On receipt, the samples were moved to a climate-controlled chamber that operated with a constant temperature (10°C) and a 12 h light/12 h dark cycle for 72 h, during which time the samples were illuminated with photosynthetically active light (400–700 nm, mercury lamp) at a photosynthetically active photon flux density of ∼100 μmol/(m2·s). For rehydration, the samples were sprayed twice per day with 25 mL of deionized water. These parameters mimic the conditions of the native environment of the lichens, hence they were assumed to induce their native physiological activity, which included high water availability during early morning dew formation, low temperatures, and low-light conditions (Lange et al., 1970).
The metabolic activity of the photobiont was measured by using a Mini-PAM fluorometer (Heinz Walz GmbH) (Sancho et al., 2007). The first measurement of the maximum quantum yield of PSII (for nomenclature see van Kooten and Snel, 1990) was assessed following the procedure of Maxwell and Johnson (2000). Before fluorescence measurements, the samples were kept in darkness for 20 min so that all reaction centers would be open during the fluorescence measurement procedure (Schreiber et al., 1995). Maximum quantum yield was calculated (following van Kooten and Snel, 1990) from the equation:
where Fv = variable fluorescence yield, Fm = maximal fluorescence yield, and Fo = minimal fluorescence yield. The maximum quantum yield activity of PSII was calculated to quantify the vitality of the lichen. A comparison of the ratio of the Fv/Fm of the flight samples to that of the ground control pre-flight samples provided a means to determine their relative vitality.
2.3.2. Confocal laser scanning microscopy
Fluorescence and electron microscopy techniques were used to evaluate the structural integrity and cell metabolic viability of the algae and fungi of the vagrant lichens pre- and post-flight.
To assess changes in cell viability, confocal laser scanning microscopy was conducted on all three sets of samples (flight, LCs, and ground controls MGR). On receipt (Table 1), the lichen samples were rehydrated with deionized water and re-activated for 12 h at 10°C in a climate-controlled chamber. Subsequently, samples were incubated for an additional 48 h in a Sanyo MLR-351 plant growth chamber at 21°C, 60–70% relative humidity, and 12 h dark/12 h illumination, with 250 μmol/(m2·s) photosynthetically active radiation. Microtomed cross-sections of reactivated thallus samples (∼100 μm thick) were then prepared by using a vibratome and immediately stained with a 25 μM solution of the fluorescent dye, FUN® 1 (Thermo Fisher Scientific) in 2% glucose and 10 mM HEPES [4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid] buffer (pH 7.0). Samples were incubated at 25°C for 2 h, which was the optimal incubation time for the processing of the FUN 1 dye by the lichen thalli. FUN 1-stained samples were then observed by using a Zeiss LSM 7 DUO Confocal Microscope (Zeiss GmbH) with a 10 × EC Plan-Neofluar objective lens. Images were acquired at 8 bits with an 1024 Å (∼1024 pixels) format, using the 488 nm line of an argon ML laser as the only excitation source and a mean beam splitter of 488/561. Emission bands for each channel were 505–550 nm (for green fluorescence of the dye), 581–620 nm (for red fluorescence of the dye), and 672–758 nm (for chlorophyll fluorescence). Image stacks were taken through the z axis at 4-μm intervals through the entire microtomed section and analyzed with ZEN 2011 software (Zeiss GmbH). When metabolically active fungal cells take up the dye, they exhibit red-orange fluorescence, whereas non-active cells depict yellow-green fluorescence (Millard et al., 1997). Fiji software (Schindelin et al., 2012;
Sample Delivery Period in Days from German Aerospace Center (Cologne) to Each Laboratory for Analyses
BIOMEX = Biology and Mars Experiment; CLSM = confocal laser scanning microscope; DLR = German Aerospace Center; FESEM = field emission scanning electron microscope; PCR = polymerase chain reaction; RAPD = random amplification of polymorphic DNA; TEM = transmission electron microscope.
2.3.3. Scanning electron microscopy
The morphological integrity of cell structures exposed to space and Mars-like conditions was investigated through imaging of flight samples and LCs with the use of a field emission scanning electron microscope (FESEM) and a cryogenic temperature scanning electron microscope (Cryo-SEM). For FESEM, a split piece of the cross-sections of reactivated lichen thalli, previously prepared for confocal laser scanning microscopy, was submerged and fixed in 2.5% glutaraldehyde in 0.1 M cacodylate-buffer (pH 7.4) at 4°C for 2 h. Samples were then washed in cacodylate buffer for 5 min and post-fixed in 1% osmium tetroxide for 1 h at 4°C. The lichen samples were subsequently dehydrated by using an ethanol (30%, 50%, and 70% for 30 min each) and acetone (70% and 90%) dilution series, finishing with 100% acetone for 1 h before drying in a critical point drier (Leica EM CPD300, Wetzlar, Germany) at 34.5°C. Fixed lichen samples were gold sputter-coated (5–10 nm) and examined in a FEI Teneo FESEM (FEI Company, Eindhoven, The Netherlands) by using the secondary electron detection mode with an acceleration voltage of 5 kV for ultra-high-resolution images and 10 kV for elemental microanalysis (Working Distance = 10 mm). Cryo-SEM examination of the flight samples was conducted with a Zeiss EVO LS15 microscope operating at −120°C, with an acceleration potential of 10 kV and a probe current of 290 pA. For cryo-fixation, lichen cross-sections were mounted onto the specimen holder of the cryo-transfer system and plunged into liquid nitrogen. Once frozen, the whole unit was cryo-transferred into the cryo-preparation unit (Leica EM ACE 600), where they were etched for 20 min at −90°C. After ∼5 min sublimation, the etched surfaces were sputtered with platinum and cryo-transferred onto a microscope cold stage.
2.3.4. Transmission electron microscopy
On receipt of the desiccated samples of C. gyrosa in Lisbon (Table 2), they were rehydrated with deionized water for 48 h in a rotator at room temperature (RT) with exposure to natural light. Samples were then fixed overnight at 4°C in the dark in 1% paraformaldehyde and 2% glutaraldehyde in 0.1 M cacodylate buffer (pH 7.4) (EMS, Hatfield, PA, USA). After fixation, the samples were washed three times for 5 min with 0.1 M cacodylate buffer. The samples were then post-fixed with 1% osmium tetroxide (EMS) in the same buffer for 1 h in the dark at 4°C. Then, the samples were washed twice for 20 min at 4°C in 0.1 M cacodylate buffer and twice in deionized water. After this, they were immersed in 2% uranyl acetate in water in the dark for 2 h at RT. Dehydration was performed at RT by using serial dilutions of acetone as follows: once in 50%, 70%, and 90% acetone for 30 min and twice in 100% acetone for 15 min. Samples were then embedded in epoxy resin (EMS) by immersion in mixtures of acetone: epoxy resin (3:1, 1:1; 1:3) and pure epoxy resin for 1 h at RT in a rotator and then they were incubated overnight at 4°C in epoxy resin. After assembling the samples in BEEM® embedding capsules, the epoxy resin was allowed to polymerize at 65°C for ≥48 h. Ultra-thin sections were obtained by using an ultramicrotome (Leica ultracut R) and contrasted with saturated uranyl acetate in water for 30 min, followed by Reynolds lead citrate for 3 min. Transmission electron microscopy observations were performed by using a Hitachi 8100 transmission electron microscope (TEM; Hiatchi, Tokyo, Japan). Digital images were acquired with a Charged-coupled device (CCD) Keen-View camera (Olympus Soft Imaging Solutions GmbH, Munich, Germany).
Sequences of Primers Employed in the Molecular Analysis of Different Fragments of Genomic DNA of Circinaria gyrosa
2.3.5. DNA-based analysis
2.3.5.1. DNA extraction
Lichen samples (Table 2) were pulverized in mineral water and then incubated at RT for 10 min. Next, they were washed with absolute ethanol, sliced, and placed into MagNA Lyser green beads tubes (Roche) with 200 μL of buffer PL1 from the NucleoSpin Plant II Kit (Macherey-Nagel), supplemented with 5 μL of proteinase K solution (Qiagen). Subsequently, the samples were placed into the MagNA Lyser instrument (Roche), and they were subjected to a total of six pulses at 7000 rpm for 99 s each. Between each pulse and the next, tubes were refrigerated for 1 min. Next, the samples were centrifuged at 11,000 × g for 30 s, 200 μL of buffer PL1 supplemented with proteinase K added, mixed well by vortexing, and centrifuged at 24,000 × g, 4°C for 2 min. Supernatants were collected and incubated at 65°C for 1 h. Next, 100 μL of chloroform was added; samples were mixed by vortexing for 30 s and centrifuged (24,000 × g, 4°C, 45 min). Then, the aqueous layer was collected and filtered at 11,000 × g for 2 min with the NucleoSpin filters included in the NucleoSpin Plant II Kit (Macherey-Nagel). Filtrates were collected, 450 μL of buffer PC was added, and the samples were mixed and decanted into NucleoSpin Plant II K columns. Samples were then processed following the NucleoSpin Plant II Kit manual. Buffer PE (55 μL at 65°C) was used to elute DNA samples, which were then incubated for 10 min at 65°C and centrifuged at 11,000 × g for 2 min. The DNA obtained was either used immediately for PCR assays and/or refrigerated at 4°C until use.
2.3.5.2. Single-gene PCR
Internal transcribed spacer, large subunit, and small subunit amplifications were performed as described by Pacelli et al. (2017), using BioMix (BioLine), with 5 pmol of each primer, but using a higher amount of template DNA (20–60 ng), in a final volume of 25 μL. Amplifications were conducted by using a GeneAmp PCR System 9700 Thermal Cycler (Applied Biosystems). Primer sequences are reported in Table 2.
2.3.5.3. RAPD assay
The RAPD protocols described by Selbmann et al. (2011) and Pacelli et al. (2017) for alga and fungi, respectively, were used, except that a higher amount of template DNA (3–16 ng) was included. The primer sequences used in this study are reported in Table 1.
3. Results
3.1. BIOMEX/EXPOSE R2
Samples were exposed to different parameters of space- and Mars-like conditions, as recorded by the ESA (Rabbow et al., 2017). After retrieval, samples were distributed to different laboratories for analysis of the effects of exposure to space- and Mars-like conditions (Table 2).
3.2. Lichen PSII activity after space exposure
The quantum yield activity of PSII, assessed by measurement of chlorophyll a fluorescence, was taken as an indicator of the vitality of the lichens and determined after 72 h of hydration and pre-adaptation of the desiccated specimens. Dark flight controls not exposed to solar UV radiation (space and Mars UV-radiation) showed no significant reduction of the PSII activity both before and after flight. These data demonstrate that the photosynthetic activity of the samples recovered rapidly after the flight (Fig. 3A and Table 3).

Photosynthetic performance of PSII measured by chlorophyll a fluorescence:
Biological Test System Circinaria gyrosa of the Biology and Mars Experiment, the Space Parameters Tested, and Their Effects on the Biological Systems: Samples Were Exposed to Space Vacuum and Solar Extraterrestrial Ultraviolet Radiation (Ultraviolet >110 nm and Visible), and to Mars Surface Radiation (>200 nm), Atmosphere Composition, and Pressure
PSII = photosystem II; UV = ultraviolet.
Exposure to UV irradiation during the space mission reduced the maximum quantum yield of the lichens to between 11.9% and 48.5% of the pre-flight activity (Table 3). Samples exposed to UV radiation >110 nm and to space vacuum exhibited the lowest activity (11.9–30.2%) (Fig. 3A and Table 3), whereas those exposed to Mars-like UV radiation and to a Mars-like atmosphere composition and pressure showed a higher PSII activity range (40–48.5%) (Fig. 3B and Table 3).
Overall, the results from samples exposed to solar UV in LEO demonstrate moderate to high variation in PSII activity among individual samples, with higher values in those that were exposed to Mars-like conditions (mean, 44.3%) relative to Sun- and space-exposed samples (mean, 21.1%). The limited capacity of the EXPOSE R2 flight hardware could not accommodate more than two samples per test parameter; therefore, statistical analysis of the data was not possible.
3.3. Viability of the lichen C. gyrosa determined by CLSM
The CLSM analyses of samples stained with the fluorescent dye, FUN 1 were used to determine the cell viability of the C. gyrosa mycobiont after exposure to space and Mars-like conditions. This technique has previously been used to determine the viability of lichen symbionts after they were subjected to stress in various experiments (Brandt et al., 2015; Meeßen et al., 2015; de la Torre et al., 2017). Fungal and algal cells take up the dye and, when metabolized, the emission properties of FUN 1 change (Millard et al., 1997). The conversion from the original yellow-green fluorescence of the dye to red-orange requires both membrane integrity and metabolic activity of cells, thus red cells observed by CLSM are considered metabolically active and therefore viable.
The density and dispersion of fungal filaments in the cortex and medulla of C. gyrosa thalli exhibited clear differences. To elucidate whether these differences affected cell viability, measurements of cell recovery were conducted separately in both structural areas of the thallus. The mean percentage values of metabolically active cortical hyphae ranged from 62% to 120%, and from 56% to 84% for the medullary hyphae (Fig. 4A). The metabolic recovery of cortical fungal hyphae was greater for samples exposed to space than to Mars-like conditions. The medullary hyphae showed less capacity for recovery than the cortical hyphae for all samples (Fig. 4A).
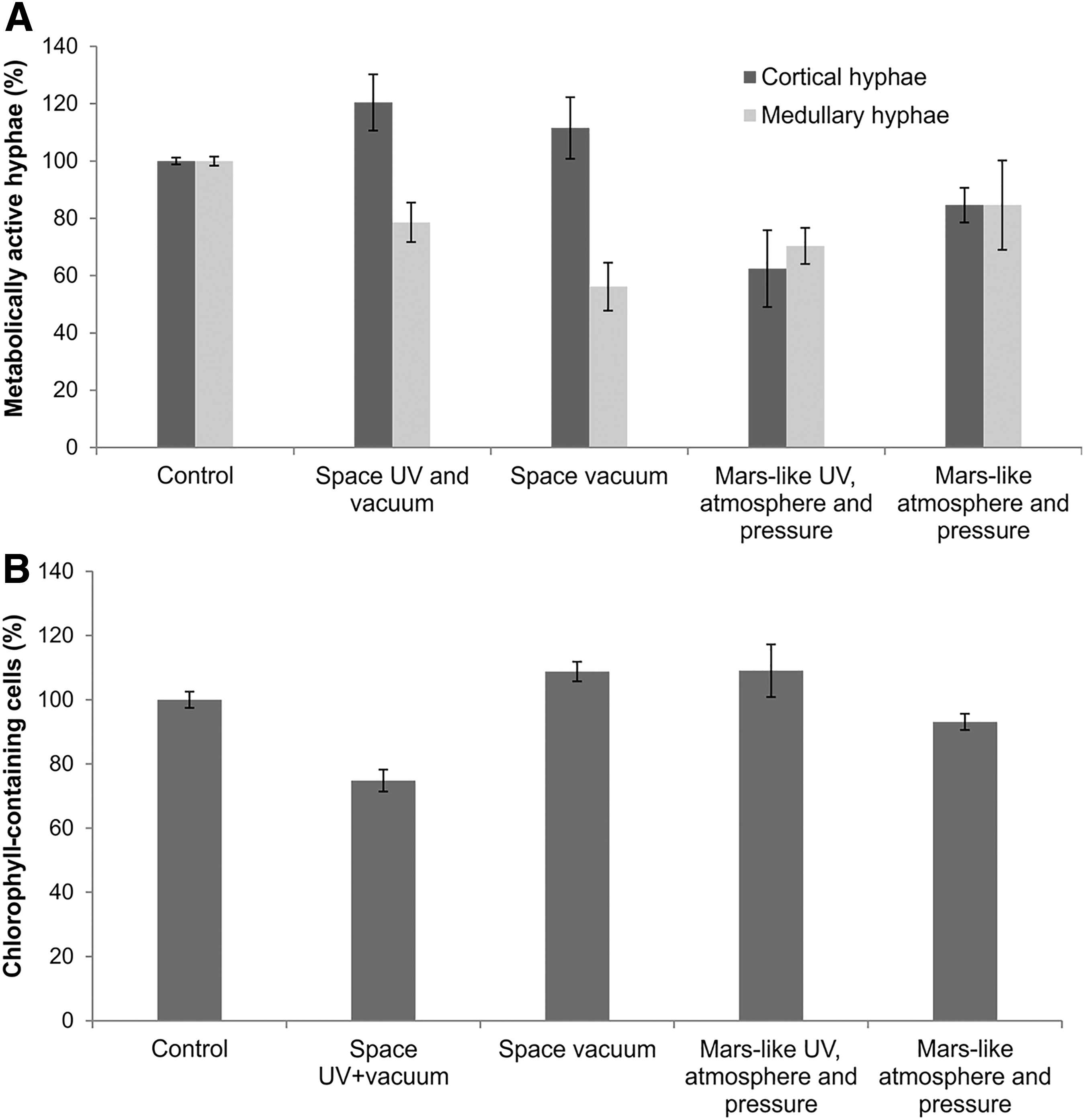
Recovery assays of the mycobiont
Since chlorophyll-induced autofluorescence can be used as an indicator of optimal photosynthetic activity (Osticioli et al., 2013), an assessment of their recovery was performed by counting the number of chlorophyll-containing particles per unit area (Schulze et al., 2011). Though photobiont cells were less efficient at internalization of the FUN 1 dye, the mean percentage value of chlorophyll-containing cells was very similar in all samples, except for samples exposed to space UV radiation and vacuum (mean value of the chlorophyll-containing cells was 75%) (Fig. 4B).
Changes in the shape and autofluorescence of the algal cells of all samples exposed to space UV radiation and vacuum were also compared (Fig. 5). The shape of the algal cells on the flight experiment appeared to be more polyhedral in nature (Fig. 5B). Photobiont cells exposed to space vacuum (Fig. 5C) and Mars-like UV, vacuum, and pressure (Fig. 5D) exhibited good recovery in terms of the number of autofluorescent cells. It is worth noting that the samples exposed to Mars-like conditions exhibited a highly irregular distribution of algal cells, with areas containing densely grouped autofluorescent algae (Fig. 5D) and regions largely devoid of chlorophyll-containing cells (Fig. 5E). Samples exposed to Mars-like atmosphere and pressure also exhibited good recovery of the photobiont (Fig. 5F).
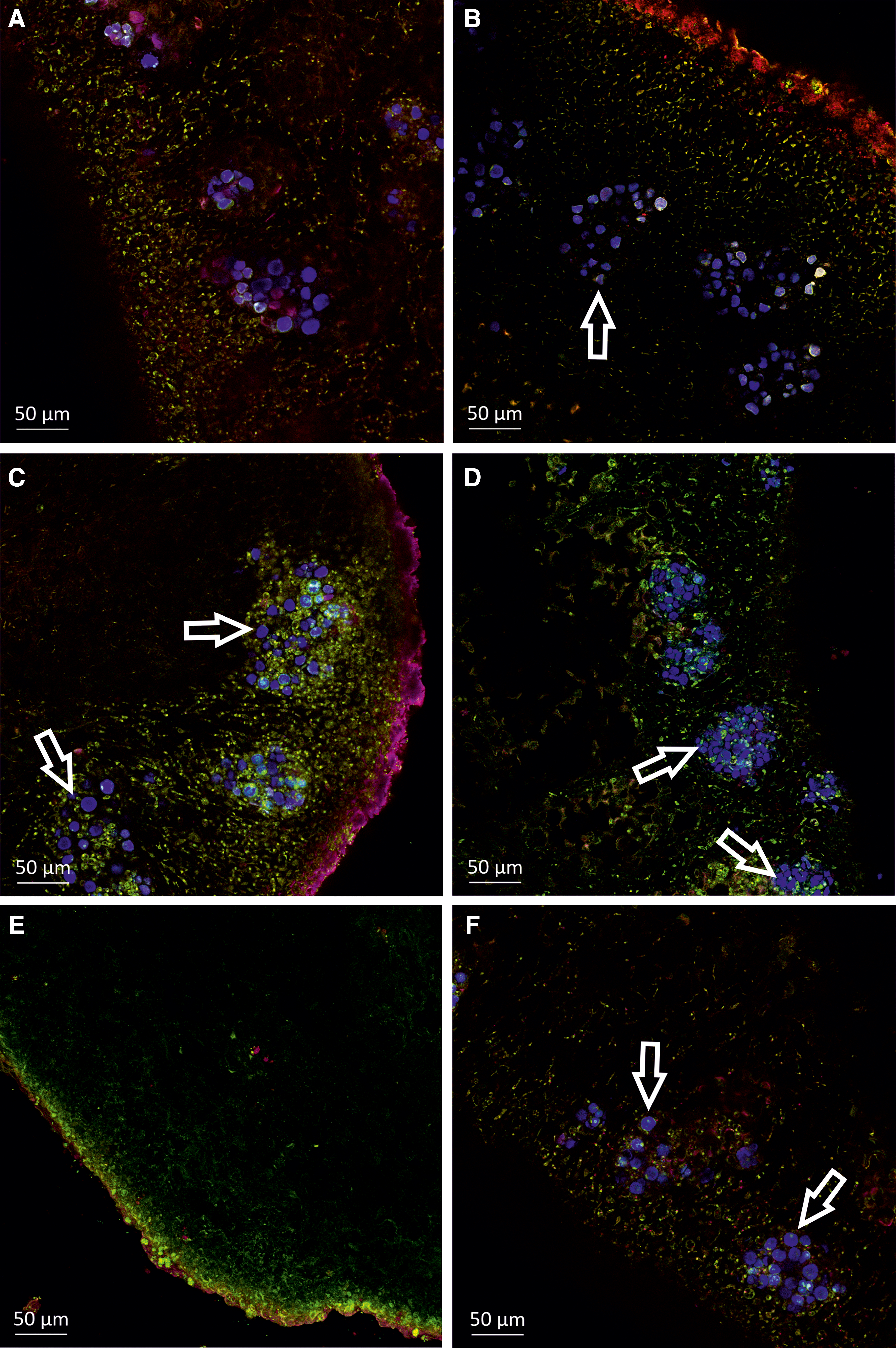
CLSM images of FUN 1-stained C. gyrosa cross-sections of pre-flight control samples
3.4. Morphological features of C. gyrosa examined by microscopy methods
FESEM observations of rehydrated C. gyrosa flight samples (FS) provided insights into the integrity of cell structures after long-term exposure to space and Mars-like conditions on the EXPOSE R2 platform (Fig. 6). In the control samples, healthy intact algal cells arranged in clusters were easily discerned and their localization in the lichen thalli was well preserved (Fig. 6A). There was also good anatomical preservation of medullary hyphae in the control samples (Fig. 6B). In many reactivated samples exposed to space UV radiation and vacuum, the algal layer contained shrunken algal cells (Fig. 6C); however, undisrupted photobiont cells were also frequently found in the same sample sections (Fig. 6D). These cells retained their globular shape and their surfaces were not collapsed (Fig. 6D), which suggested a rapid recovery of the photobiont after rehydration. Similar rapid recovery of medullary hyphae, in which intact fungal cells were detected, was also noted (Fig. 6E). Crystalline lichen secondary metabolites were detected in the pre-flight samples and in all of the flight and laboratory ground control samples. These extracellular lichen products were abundant on fungal cell wall surfaces and voids in the lichen medulla and had the characteristic shape of whewellite (calcium oxalate monohydrate) crystals (Fig. 6B, F). Both shrunken and well-preserved photobiont cells were also detected in the flight dark samples (i.e., those exposed to space vacuum and protected from UV radiation) (Fig. 6G). The shrinkage of mycobiont cells heavily encrusted in whewellite-like crystals was evident in the medulla of these lichen samples (Fig. 6H).

SEM images obtained using FESEM or Cryo-SEM of C. gyrosa cross-sections of pre-flight control samples
In contrast, the lichen samples exposed to Mars-like UV radiation, atmosphere, and pressure revealed abundant cavities devoid of algal cells (Fig. 6I), suggesting that many samples had lost their algal layers (e.g., due to the removal of shrunken cells during sample preparation). Some photobiont cells with well-preserved globular shapes were also observed (Fig. 6J) in the algal layer of the same cross-section of the lichen thallus. Cell shrinkage and disruption of the algal layer were also observed in samples exposed to Mars-like atmosphere and pressure, without UV radiation (Fig. 6K). The medulla contained interlaced hyphae with numerous voids (Fig. 6L).
TEM examination revealed that the photobiont (Fig. 7A) and mycobiont (Fig. 7B) ultrastructure of pre-flight control samples was well preserved, consistent with previous reports (de la Torre et al., 2017; Pacelli et al., 2017). Exposure to space UV radiation and vacuum caused ultrastructural alterations in C. gyrosa, evidenced by cell shrinkage with structural membrane alterations visible in both photobiont (Fig. 7C) and mycobiont (Fig. 7D) cells. Large vacuoles were particularly evident in algal cells (Fig. 7C). Unfortunately, cellular structures were not observed in flight dark samples (samples exposed to space vacuum), which hindered identification of the lichen symbionts (data not shown).
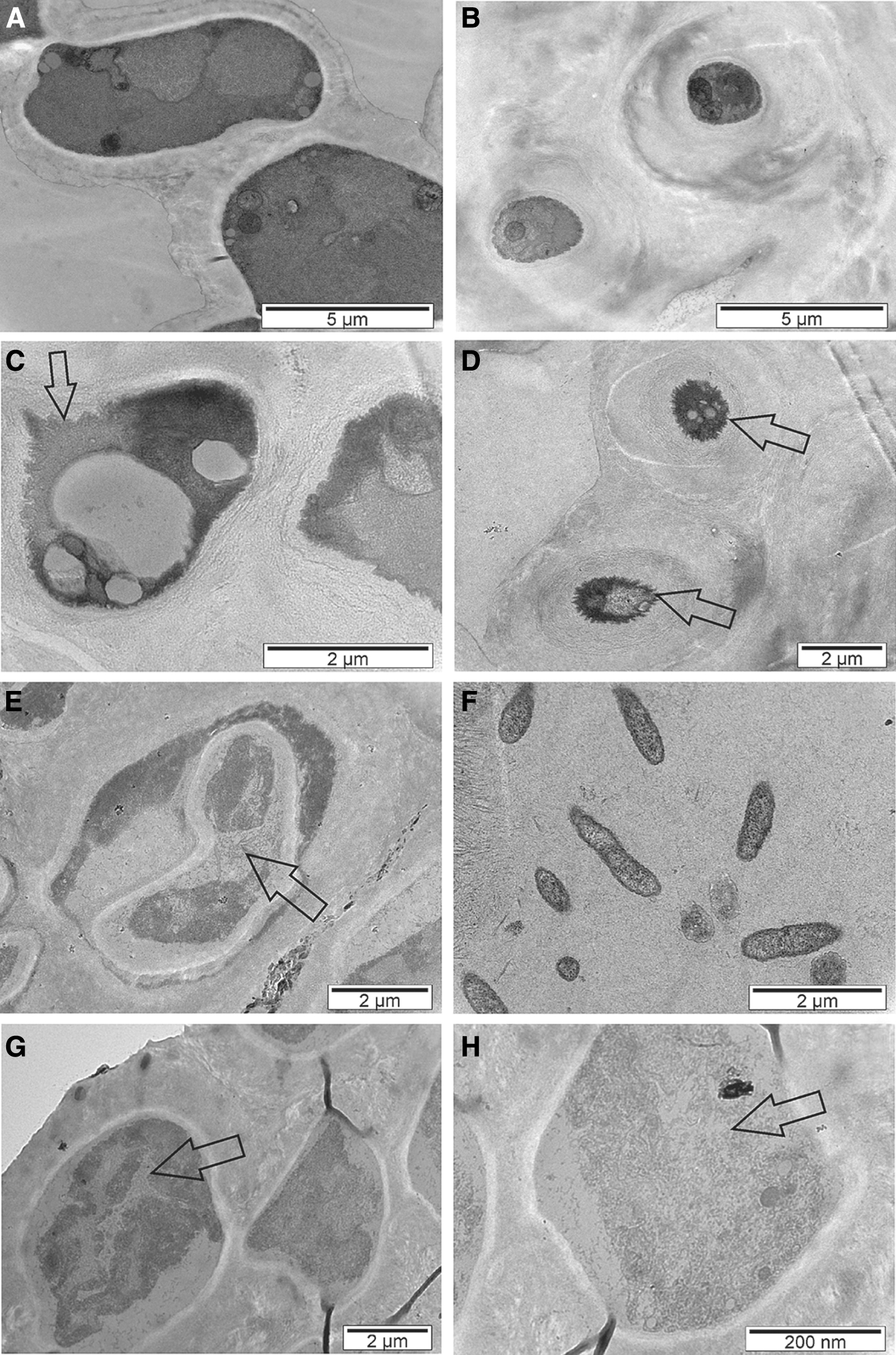
TEM images of C. gyrosa ultrathin sections of pre-flight control samples
The ultrastructural alterations identified by transmission electron microscopy were found to be less evident in lichen samples exposed to Mars-like UV radiation, atmosphere, and pressure when compared with those specimens exposed to space conditions (Fig. 7E–H). Some amount of cell shrinkage with reduction of cellular content was observed for both the photobiont (Fig. 7E) and mycobiont (Fig. 7F). The shape of these cells was irregular and contained large vacuoles filled with cell debris (Fig. 7E, G, H). This dendritic-like cell morphology due to the accumulation of cell debris was observed in both the photobiont (Fig. 7E) and mycobiont (Fig. 7H).
3.5. Polymerase chain reaction
3.5.1. Single-gene PCR
To test the integrity of genomic DNA in the lichen samples, six PCR assays that amplified different ribosomal gene markers were performed. DNA damage was most evident in amplicons with the highest molecular weight (>1100 bp), although it was observed in all PCR amplifications. UV radiation appeared to produce the most DNA damage and appeared to be particularly detrimental in samples exposed to the space environment versus those samples exposed to Mars-like conditions. Some incongruence was observed among samples treated under the same experimental conditions, however, as DNA mutations would have occurred randomly in each sample. Also, the amounts of samples available were very limiting, including the LC. The level of DNA alteration should be further assessed by Sanger sequencing in future studies (Table 4 and Fig. 8).

Assessment of DNA damage in C. gyrosa samples from the BIOMEX: single-gene PCR.
Resume of Sample Name, Experimental Conditions, Sample Size (mg), DNA Concentration (ng/μl), and Single-Gene Polymerase Chain Reaction and Random Amplification of Polymorphic DNA Genomic Fingerprinting Results of Circinaria gyrosa Samples from the Biology and Mars Experiment
DNA concentration was measured in a NanoDrop ND1000 spectrophotometer (Thermo Fisher Scientific).
+ = Positive amplification (no DNA damages); − = negative amplification (DNA damages); ITS = internal transcribed spacer; LSU = large subunit; SSU = small subunit.
3.5.2. RAPD assay
The DNA from one sample exposed to space vacuum was not detected by analysis of RAPD profiles; also, a considerable decrease in the intensity of the bands obtained was observed in one sample exposed to Mars conditions (UV radiation+atmosphere+pressure). According to the results of single-gene PCR, UV radiation appeared to be the most damaging condition, particularly in the space environment. Nevertheless, RAPD profiles were mostly preserved in at least one sample from every condition tested (both space and Mars-like). The same incongruence was observed in the single gene results. Collectively, these results corroborate the occurrence of DNA damage in lichens exposed to the harsh environmental conditions (primarily UV radiation) of space and Mars-like parameters (Table 4 and Fig. 9), though larger sample volumes are recommended for future studies so minimum detection thresholds are exceeded.

Assessment of DNA damage in C. gyrosa samples from the BIOMEX: RAPD genomic fingerprinting.
4. Discussion
Our combined results from this BIOMEX, using the vagrant lichen species, C. gyrosa, clearly demonstrate the resistance of lichens to actual space and Mars-like conditions. The previous experiment LICHENS (Sancho et al., 2007, 2008; de los Ríos et al., 2010) also reported the high resistance of two other lichen species, R. geographicum and X. elegans, to all space parameters (i.e., based on their PSII activity after a maximal exposure of 16 days at LEO on board the Foton satellite, relative to other experiments included in the same Biopan-6 mission, c.f., Jönsson et al., 2008).
Simulation experiments (Sánchez et al., 2012) performed at the “Planetary Atmospheres and Surfaces Chamber” (PASC, Centro de Astrobiologia [CAB], Madrid, Spain) under space- and Mars-like conditions, as well as experiments under real space conditions (de la Torre et al., 2010, Raggio et al., 2011), have previously demonstrated that the lichen C. gyrosa exhibits high resistance and survival capacity. These results showed that neither martian atmosphere and surface UV climate combinations nor LEO vacuum conditions induce a significant decrease in the activity of lichen after exposure for 120 h.
It is important to emphasize that in our experiment the lichen thalli were exposed to simulated Mars and real space conditions. In particular, they were subjected to solar UV radiation, the most deleterious factor in space, which is directly absorbed by the DNA (Horneck, 1993, Horneck et al., 1995). These data, together with previous results, indicate extraordinary levels of resistance in this lichen toward space- and simulated planetary environments (de Vera et al., 2003, 2004; Sancho et al., 2007, 2008; de la Torre et al., 2010; de Vera and Ott, 2010; Raggio et al., 2011; Onofri et al., 2012; de la Torre Noetzel et al., 2018).
The mechanisms underlying the astrobiological tolerance of model lichens to outer space environmental stresses remain incompletely understood. Anhydrobiosis (Sánchez-Iñigo, 2013; Brandt et al., 2015), morphological traits (Meeßen et al., 2013), and secondary lichen compounds (Meeßen et al., 2014) are important factors in the high levels of resistance exhibited by lichens.
The results of the present BIOMEX demonstrate that, after a long exposure period (18 months) at the EXPOSE-R2 facility on the ISS, it remains possible for C. gyrosa to recover PSII activity. Although PSII activity, as determined by measurement of chlorophyll a fluorescence, was further reduced in Sun-exposed samples for space- and Mars-like conditions (Fig. 3), the maximal mean reduction was only 21% for space-like conditions in comparison with pre-flight values under vacuum, whereas PSII activity for Mars-like conditions decreased to a mean of 44.25%. It is interesting to note that the response of the dark-control samples revealed an opposite trend, with PSII activity averaging 45.41% under space vacuum and 85.71% under Mars atmospheric composition and pressure. These data indicate a lower recovery tendency of the photosynthetic system of lichens after exposure to space conditions, including UV irradiation, which is consistent with earlier reports of the results of simulation experiments (Gómez et al., 2010; Sánchez et al., 2012; Gómez-Gómez et al., 2016) that include the LICHENS experiment on board Biopan-5 (Sancho et al., 2007, 2008), the LITHOPANSPERMIA experiment (de la Torre et al., 2010) on board Biopan-6, and the LIFE on the EXPOSE-E of the ISS (Onofri et al., 2012).
Comparison of the data from samples exposed to space- and Mars-simulated UV radiation to that from dark flight dark samples in our experiment supports the suggestion that the compact morphological structure of C. gyrosa, including its thick cortex and cortex, acts as an endogenous protective shield against UV radiation (Sánchez-Iñigo, 2013). The presence of Sun-screening pigments in various lichen species is established (Solhaug et al., 2003), especially among those living in Arctic habitats (Nybakken et al., 2004) and high mountain areas. For example, the cortex protects R. geographicum from the harsh environmental characteristics of high mountain regions (de la Torre et al., 2007). A Sun-screening effect of pigments was also determined in X. elegans flight samples, by depigmentation of a parallel set of samples before the flight (de la Torre et al., 2010); however, because the PSII activity of the depigmented dark flight samples was (76.6% ± 19.6%) lower than that of the intact dark flight samples (100.3% ± 4.0%), it is more likely that depigmentation per se made the lichens more sensitive to the complex matrix of space parameters applied to them. Given that C. gyrosa thalli contain low concentrations of Sun-screening pigments (Sánchez-Iñigo, 2013), this high resistance to UV radiation must depend on other factors (e.g., morphological features) (Meeßen et al., 2013, 2014).
Our results proved our hypothesis that the mycobiont (fungal) part of C. gyrosa provides protection via the cortex cells, whether alive or dead, by surrounding the algal cells, which was demonstrated by the viability data from post-flight C. gyrosa samples that confirmed that UV radiation at real space conditions (>110 nm) combined with space vacuum was the most deleterious environmental factor for the photobiont cells. According to the CLSM analysis, the metabolic recovery of samples exposed in space indicated viability of the fungal cells in the cortex. Hyphae from the photobiont layer typically form short or globose cells with thicker walls than medullary hyphae (Büdel and Scheidegger, 2008), which could explain the higher recovery of the cortex fungal cells. In addition, the hyphae cell walls of the medulla were heavily encrusted with whewellite-like crystals for all the pre-flight and post-flight samples, as was also described in previous studies (Böttger et al., 2013; Meeßen et al., 2013). This encrustation by crystalline lichen secondary metabolites might increase the hydrophobicity of the medullary hyphae, hindering their recovery during re-activation, when compared with the cortical hyphae. These extracellular lichen products were particularly abundant on fungal cell walls from the lichen medulla and had the characteristic shape of whewellite (calcium oxalate monohydrate) crystals (Fig. 6F), as also observed by de la Torre Noetzel et al. (2018), in samples exposed to Mars-like conditions. The morphology of the algal cells in samples exposed to UV radiation, under both space and Mars-like conditions, was polyhedral, rather than spherical (Fig. 5B, D), corroborating the morphological alterations observed with the FESEM (Fig. 6) and TEM (Fig. 7). These electron microscopy findings correlate with the results of PSII function analysis, with a higher recovery of the flight dark samples compared with those exposed to extraterrestrial UV radiation (Fig. 7C, E), which exhibited cell shrinkage caused by plasmolysis, likely attributable to their pronounced decrease in photosynthetic activity (Fig. 3).
The lethal effects and DNA damage caused by UV radiation from 200 to 400 nm, and particularly at 254 nm, have been studied extensively. For example, data from previous experiments under real space conditions (Cockell et al., 2011) showed that a natural phototrophic biofilm augmented with extreme-tolerant cyanobacteria did not survive when exposed to unattenuated extraterrestrial UV radiation, with the exception of the cyanobacterium. The UV flux caused the destruction of carotenoids and bleached the chlorophyll and accessory pigments. In contrast, the dark controls (samples exposed underneath the UV-exposed samples) exhibited superior survival, as was observed in our experiment.
DNA analyses of C. gyrosa were applied for the first time to assess the mutagenic effects of ionizing radiation on DNA, as a key target; consequently, we used novel molecular biomarkers of the effects of ionizing radiation on biological systems. Some similar studies of the DNA integrity of selected eukaryotic microorganisms, including Cryomyces antarcticus, Umbilicaria sp., and Stichococcus sp., after electromagnetic radiation (UVB) or ionizing radiation (gamma-60Co), have recently been reported (Selbmann et al., 2011; Pacelli et al., 2017).
Single-gene PCR analysis revealed DNA damage in the highest molecular weight bands of lichen previously exposed to extraterrestrial conditions, which demonstrated that UV radiation was the most damaging parameter, particularly in combination with space vacuum, although RAPD profiles were well preserved in almost all experimental conditions analyzed. These results are similar to those obtained with simulated space gamma radiation (Keresztes and Kovács, 2002; Pacelli et al., 2017), but they differ from another experiment carried out with simulated UV radiation (Selbmann et al., 2011). It is worth noting that this study was performed with a lichen species in real space and Mars-like conditions during the course of 18 months, which could explain the lower recovery tendency of the photosynthetic system observed in C. gyrosa. PCR analysis confirmed DNA damage in lichen previously exposed to extraterrestrial conditions and demonstrated that UV radiation was the most damaging parameter, particularly in combination with space vacuum.
Collectively, our results confirmed previous findings and provide a foundation for deeper exploration of the limits of terrestrial life in space and planetary environments.
5. Conclusions
Data indicate that the survival of the lichen, C. gyrosa, after exposure outside the ISS in the context of the EXPOSE-R2 space mission, is sensitive to both space UV radiation and Mars-like UV radiation, and less sensitive to vacuum. These findings add to our knowledge of the resistance of life to space conditions and contribute to an understanding of the adaptation potential of extremophiles to the martian surface, and the resistance of lichens in particular.
Footnotes
Acknowledgments
The authors thank the anonymous reviewers for their comments and suggestions.
Author Disclosure Statement
No competing financial interest exists.
Funding Information
This work was supported by grants of the Spanish Ministry of Economy, Industry and Competitiveness (MINisterio de ECOnomía y Empresa, project SUBLIMAS “SUrvival of Bacteria and LIchens on Mars Analogs and Space,” ESP2015-69810-R, 2015, to Rosa de la Torre Noetzel, and Leopoldo García Sancho (PI of the Project CTM2015-64728-C2-1-R) and project Biocomplexity and Functioning of the Cryptogamic Cover in Arid and Polar Regions. a micro- and mesoscale approach . The authors also thank the German Aerospace Center (Deutsches Zentrum für Luft- und Raumfahrt) as well as the European Space Agency (ESA) for supporting the space experiment BIOlogy and Mars Experiment (ESA-International Life Sciences Research Announcement 2009-0834, PI: J.-P.P.V.).