
Abstract
Microbial ecology of permafrost, due to its ecological and astrobiological importance, has been in the focus of studies in past decades. Although permafrost is an ancient and stable environment, it is also subjected to current climate changes. Permafrost degradation often results in generation of thaw ponds, a phenomenon not only reported mainly from polar regions but also present in high-altitude permafrost environments. Our knowledge about microbial communities of thaw ponds in these unique, remote mountain habitats is sparse. This study presents the first culture collection and results of the next-generation DNA sequencing (NGS) analysis of bacterial communities inhabiting a high-altitude permafrost thaw pond. In February 2016, a permafrost thaw pond on the Ojos del Salado at 5900 m a.s.l. (meters above sea level) was sampled as part of the Hungarian Dry Andes Research Programme. A culture collection of 125 isolates was established, containing altogether 11 genera belonging to phyla Bacteroidetes, Actinobacteria, and Proteobacteria. Simplified bacterial communities with a high proportion of candidate and hitherto uncultured bacteria were revealed by Illumina MiSeq NGS. Water of the thaw pond was dominated by Bacteroidetes and Proteobacteria, while in the sediment of the lake and permafrost, members of Acidobacteria, Actinobacteria, Bacteroidetes, Patescibacteria, Proteobacteria, and Verrucomicrobia were abundant. This permafrost habitat can be interesting as a potential Mars analog.
1. Introduction
On Earth, permafrost, that is, soil or sediment frozen for at least 2 years, is widely distributed, representing ∼26% of the terrestrial soils and found in the polar regions as well as high altitudes (Steven et al., 2006). Despite the harsh circumstances represented by prolonged exposure to subzero temperatures often coupled with extreme low nutrient content and high background gamma radiation, permafrost is an ancient and stable environment harboring relatively diverse communities of microorganisms, mainly bacteria, adapted to specific environmental factors (Jansson and Taş, 2014).
Besides providing unique niches for microorganisms (Gilichinsky, 2002), permafrost has great astrobiological importance (Gilichinsky et al., 2007) given that in our solar system, most planets and moons, due to their distance from the Sun, have a cryogenic environment (Gilichinsky, 2002). Biomarkers harbored by permafrost can be used in the search for possible extraterrestrial ecosystems, for example, culturable microorganisms can be used as biosignatures (Gilichinsky, 2002; Vishnivetskaya et al., 2006).
Permafrost and its degradation as a potential source of meltwater can be important analogs for Mars where this type of water body might have been present in the past (Cabrol and Grin, 1999; Fassett and Head, 2008; Kereszturi, 2010). Various thermokarst features (Hargitai and Kereszturi, 2015) point to the formation and source of lakes on Mars, including recent periods (Soare et al., 2008), while degradation of ice-cemented dust happened in the form of latitude-dependent mantle recently (Kreslavsky and Head, 2002), connected to various surface features (Séjourné et al., 2015) where mainly gullies, but not lake sediments, could be observed.
Past lakes on Mars have high astrobiological relevance (Domagal-Goldman et al., 2016), especially if they are connected to the melting of permafrost; however, there is a debate on the potential lifetime (Kereszturi, 2012) of such lakes with the possibility of not only ephemeral (Coleman et al., 2007) but also long-duration ones (Ori et al., 2000). Some potential Mars analog lakes have also been analyzed on Earth (Gómez et al., 2006; Prieto-Ballesteros et al., 2006); however, no detailed information exists on the biological characteristics of permafrost thaw ponds at a high-altitude terrain.
Nowadays, the rapidly changing climate has been reported to cause degradation of permafrost (Cheng and Wu, 2007; Schuur et al., 2015) and shifts in both microbial community compositions and activities (Vincent, 2010; Mackelprang et al., 2011). Accumulation of permafrost meltwater results in thaw ponds, unique environments present in the Canadian subarctic (Laurion et al., 2010) or at high-altitude permafrost sites of the Dry Andes (Kereszturi et al., 2020; Nagy et al., 2018).
In the past decades, many studies focused on the microbiology of permafrost (Steven et al., 2006) and drew attention to its significance for astrobiology. Bacterial communities in the permafrost of Antarctica (Cary et al., 2010), the High Arctic (Steven et al., 2007, 2008; Wilhelm et al., 2011), high-altitude sites (Zhang et al., 2007, 2009), and communities supported by cryopegs in Siberia (Gilichinsky et al., 2003) were examined, including cultivation-based studies on psychrophilic bacteria of permafrost (Shi et al., 1997; Bakermans et al., 2003; Jakosky et al., 2003; Vishnivetskaya et al., 2006; Steven et al., 2008).
Still, our knowledge about the microbial ecology of permafrost thaw ponds is sparse. In Alaska, bacterial communities of permafrost thaw ponds were characterized (Hultman et al., 2015), while in the Canadian High Arctic, a biogeographical analysis was performed, which revealed that a large-scale environmental gradient, especially climate, has the main influence on bacterial community composition (Comte et al., 2016). High-altitude sites with continuous permafrost subjected to climate change are rare and often hard to access. Thaw ponds above 5000 m a.s.l. are less frequent than other periglacial lakes (e.g., moraine lakes and polar lakes). High-altitude, permafrost thaw ponds are current, but most probably ephemeral habitats, where—due to low organic material content—mainly simplified bacterial communities can exist.
As in other extreme environments, bacteria can be the most important inhabitants; therefore, these lakes offer a great opportunity to analyze how a fast change in climate influences bacterial diversity. Nevertheless, they offer a natural laboratory for astrobiological examinations since the effects of both high altitude and permafrost are represented. The Dry Andes have long been known as a Mars analog (Escudero et al., 2007; Cabrol et al., 2009a, 2009b, 2018; Farías et al., 2011; Dorador, et al., 2013), and sites at more than 5000 m a.s.l. are getting more attention in the past years (Schmidt et al., 2018). The highest volcano on Earth, Ojos del Salado, is in this region, harboring several permafrost-derived high-altitude lakes suitable for astrobiological investigations.
Therefore, the aim of this study was to assess the bacterial community composition of a high-altitude, permafrost thaw pond located at 5900 m a.s.l. on the Ojos del Salado volcano. Our main goal was to evaluate the diversity of bacterial communities harbored by the thaw pond by using cultivation and next-generation DNA sequencing (NGS) methods. During this study, bacterial diversity of the water and sediment of the thaw pond, as well as the permafrost sediment surrounding the pond, was analyzed in parallel. The bacterial diversity of the Ojos del Salado thaw pond was also compared with other permafrost or high-altitude periglacial environments.
2. Materials and Methods
Several permanent, shallow freshwater ponds (27°09′S, 68°53′W) can be found north of the summit of Ojos del Salado, in Chile, in the proximity of Tejos camp. Meltwater originated mainly from thawing permafrost (and partially from the snow cover or penitentes) accumulates in shallow endorheic basins, forming thaw ponds that are among the highest altitude lakes on Earth. Located in the remote and barren Dry Andes, the ponds are affected by extremely arid climate (Nagy et al., 2018), along with high daily temperature fluctuations across the freezing point, low mean temperature, and, due to altitude, intense ultraviolet radiation, which increases 4–10% by every 1000 m (Cabrera et al., 1995). The permafrost active layer extends to a depth of around 60 cm at this altitude (Nagy et al., 2018).
Sampling took place in February 2016 during the expedition of the Hungarian Dry Andes Research Programme. Sediment samples from the pond and permafrost active layer around the pond were collected with sterilized spatulas into sterile Falcon tubes. Three water samples were also obtained by filtering 250 mL of water onto a Millipore cellulose membrane filter with 0.22-μm pore diameter using a sterilized syringe.
Altogether, seven groups of triplicate samples were taken from the permafrost thaw pond at 5900 m a.s.l. on the Ojos del Salado. Among these, one group consisted of water samples (F1–F2–F3) and the other six were from the lake and permafrost active layer sediment samples (S7–S12): from 0, 50, 100, or 150 cm far from the lake water, from the upper 5 cm of the sediment, or from a depth of 10 cm, just above the frozen layer at the time of sampling. The location of the sampling sites is pictured in Fig. 1. The description of samples is summarized in Table 1. All samples were kept frozen until processing in the laboratory.

Location of sampling sites in the environment of the permafrost thaw pond (5900 m
Description of Samples Derived from a Permafrost Thaw Pond at 5900 m Above Sea Level on Ojos del Salado, Chile
Temperature, pH, and electric conductivity of the lake water were measured on the spot with the PCSTestr 35 multimeter (EuTech Instruments, Thermo Scientific). Further chemical parameters of water (NH4 +, NO2 −, NO3 −, PO4 3−, SO4 2−, and Fe) were measured in the laboratory.
Cultivation, isolation, and identification of bacterial strains and estimation of colony-forming units per gram (CFU g−1) were carried out as described by Aszalós et al., 2020. (submitted). Briefly, samples were serially diluted and plated onto peptone-yeast extract-glucose medium (DSMZ 1140) and R2A (DSMZ 830) agar plates (pH 7). Colonies formed after 2 weeks of incubation at 8°C were counted and average CFU g−1 were estimated. DNA of the isolates was extracted with the bead-beating method (Krett et al., 2017), and the 16S rRNA gene was amplified with primers 27F and 1401R (Borsodi et al., 2015). Isolates were grouped by their ARDRA (amplified ribosomal DNA restriction analysis) patterns, and the 16S rRNA genes of the representatives were Sanger sequenced by LGC Genomics (Germany).
Sequences obtained were deposited at GenBank under accession numbers MH667812–MH667859. For identification of strains, the EzBioCloud database was used (Yoon et al., 2017). A phylogenetic tree was constructed with MEGA7 (Kumar et al., 2016) using the neighbor-joining method (Saitou and Nei, 1987). The evolutionary distances were computed with the Kimura 2-parameter method (Kimura, 1980). All positions containing gaps and missing data were eliminated.
Community DNAs from the water and sediment samples were extracted according to the manufacturer's protocol of the DNEasy PowerSoil kit (Qiagen). To test whether the triplicate samples are similar, denaturing gradient gel electrophoresis (DGGE) as a molecular fingerprint method was applied according to Aszalós et al. (2016). Based on the 16S rRNA gene banding patterns of the samples, a UPGMA (unweighted pair group method with arithmetic mean) similarity dendrogram was constructed using the Total Lab software (TotalLab Ltd.).
Before NGS, composite samples were created from pooling aliquots of the triplicates using same amounts of community DNAs. Based on the DGGE similarity dendrogram, only one of the water samples (F1) was chosen for NGS analysis. NGS was performed on the Illumina MiSeq platform by Genomics Core, Research Technology Support Facility, Michigan State University, as presented in the work of Aszalós et al., 2020 (submitted). Bioinformatic analysis was carried out as described by Aszalós et al., 2020 (submitted). Sequence reads were deposited in the NCBI SRA database and accessible through the BioProject ID PRJNA484891.
Mothur (Schloss et al., 2009) was used to create operational taxonomic units at 97% sequence similarity and to calculate diversity indices. UPGMA cluster analysis using the Bray–Curtis similarity index based on the subsampled OTU table was carried out in PAST3 (Hammer et al., 2001). A principal component analysis (PCA) was performed using PAST3 software. PCA was used to cluster bacterial community compositions of different sites based on relative abundance of corresponding phyla in each sample.
3. Results and Discussion
Description of the obtained samples is shown in Table 1. Physical–chemical characteristics of the lake water are presented in Table 2.
Water Physical–Chemical Parameters in the Thaw Pond of Ojos del Salado at 5900 m a.s.l Measured at the Time of Sampling (Asterisk) and in the Laboratory
3.1. Average CFU g−1 and cultivable diversity
The average CFU g−1 of the obtained six sediment sample groups were 3.52 × 104 ± 2.0 × 104 on nutrient-rich PYG medium and 1.91 × 105 ± 9 × 104 on nutrient-poor R2A. These values are consistent with the number of viable, aerobic heterotrophic bacteria reported from other permafrost active layer sediments (e.g., Canadian High Arctic, Antarctica) and Antarctic lake sediments with low nutrient contents (Steven et al., 2006; Wilhelm et al., 2011). It is also important to note that the applied medium had a strong effect on the estimated CFU g−1 since it was one order of magnitude higher on the poor R2A compared with the rich PYG medium.
A culture collection with altogether 125 bacterial strains was established. Phylogenetic affiliations and relationships of the representatives and their closest match in the EzBioCloud database are shown in Fig. 2. Although no noteworthy differences occurred between cultivable diversities of the studied samples, regarding the affiliation of bacterial isolates, a more diverse culture collection was established on R2A than on PYG medium, as shown in Fig. 3, and several bacterial species described from environments with low nutrient content (karst caves, freshwater, and drinking water) were detected from the studied thaw pond. The isolates belonged to Proteobacteria, Bacteroidetes, and Actinobacteria, the most common phyla detected in permafrost environments. However, members of Firmicutes were not detected, although representatives of this phylum are also commonly isolated from permafrost (Jansson and Taş, 2014).
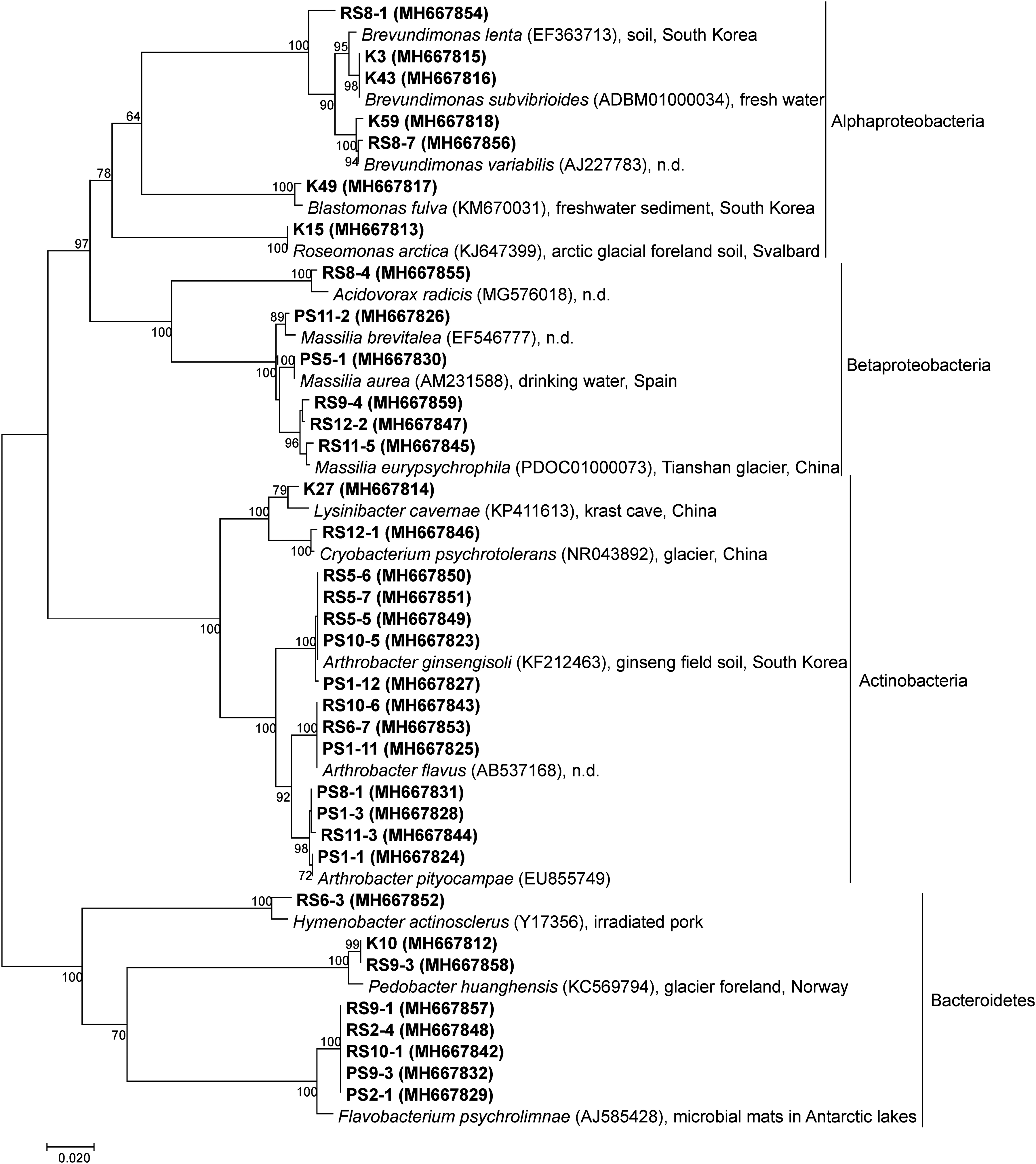
Neighbor-joining phylogenetic tree of bacterial isolates derived from the permafrost thaw pond of Ojos del Salado at 5900 m a.s.l. The optimal tree with the sum of branch length = 1.07 is shown. The percentage of replicate trees, in which the associated taxa clustered together in the bootstrap test (500 replicates), is shown next to the branches (Felsenstein, 1985). The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The analysis involved 52 nucleotide sequences with a total of 539 positions in the final dataset.
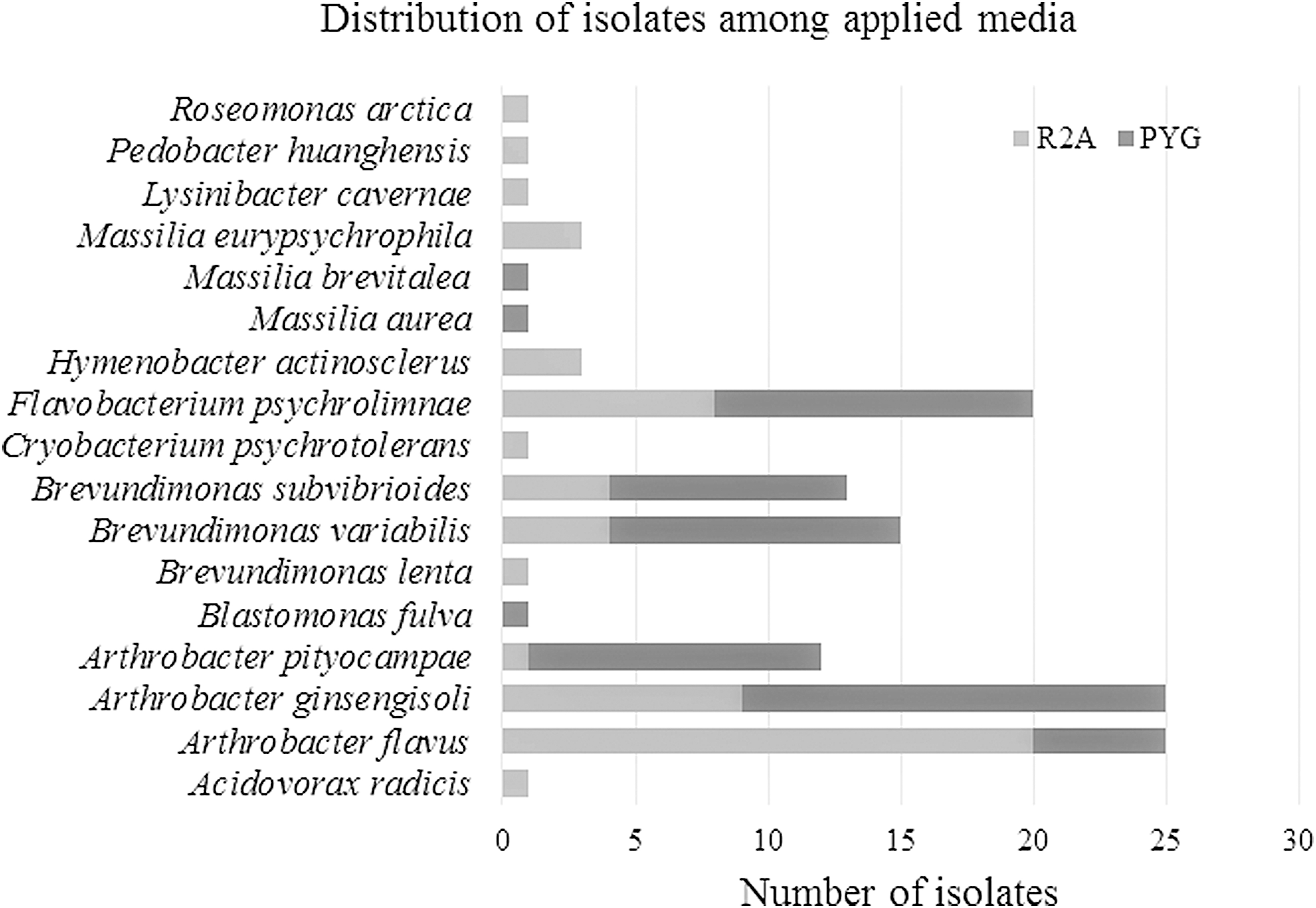
Distribution of bacterial isolates derived from the permafrost thaw pond of Ojos del Salado at 5900 m a.s.l. among the applied media R2A and PYG, peptone-yeast extract-glucose medium.
The isolated strains belonged to altogether 11 genera (Brevundimonas, Blastomonas, Roseomonas, Arthrobacter, Acidovorax, Massilia, Lysinibacter, Cryobacterium, Hymenobacter, Pedobacter, and Flavobacterium). Most of them represented psychrophilic species well adapted to temperatures below 20°C (Flavobacterium psychrolimnae, Cryobacterium psychrotolerans, Massilia eurypsychrophila, and Roseomonas arctica). The type strains of these species were isolated from high-altitude or polar (Svalbard, Antarctica) periglacial environments, which confirms the capability of these species to be active under the circumstances of this high-altitude, permafrost thaw pond.
The Gram-negative F. psychrolimnae was isolated from microbial mats in freshwater lakes of the Antarctic Dry Valleys and grows at temperatures between 5°C and 25°C with the optimum at 20°C (Van Trappen et al., 2005). The facultative psychrophilic species, M. eurypsychrophila, isolated from a glacier ice core (China), is another example of a Gram-negative species from this environment (Shen et al., 2015). Several obtained isolates affiliated with bacterial species described from glacial environments: type strain of C. psychrotolerans was derived from the foreland of the No. 1 glacier in the Tian Shan mountains, a high-altitude environment surrounded by dry climate (Zhang et al., 2007), while Pedobacter huanghensis and R. arctica were both isolated from glacier foreland in the Svalbard archipelago, Norway (Qiu et al., 2014, 2016).
Regarding cultivable diversity, the number of species in the culture collection was higher than that reported from the crater lake of the Ojos del Salado at 6480 m a.s.l., where an acidic pH and warm (40.8°C) temperature caused by fumarolic activity coupled with the high altitude (Aszalós et al., 2020 submitted). Surprisingly, taxon diversity of the strains was also higher than that of a culture collection established from permafrost active layer samples from the Canadian High Arctic, where only Arthrobacter sp. was detected (Wilhelm et al., 2011). A great proportion of strains from the Ojos del Salado thaw pond belonged to this genus, which is commonly reported from other permafrost environments, for example, in the Tian Shan mountain (Bai et al., 2006) and Siberia (Bakermans et al., 2003; Vishnivetskaya, et al., 2006).
3.2. DGGE molecular fingerprinting
On the UPGMA dendrogram based on the 16S rRNA gene molecular fingerprints of samples (Fig. 4), sediment (S7–S8–S9–S10–S11–S12) and water (F1–F2–F3) samples were distinctly separated, while the triplicate samples from each sampling site showed the highest similarity to each other. Sample S7, an upper layer thaw pond sediment sample, grouped separately from the rest of the sediment samples, which is probably the effect of the connection with water. This is confirmed by the fact that those sediment samples (S8–S9–S10–S11–S12) that were not in direct connection with the lake water grouped together and harbored slightly different bacterial communities. However, the sampling depth (upper 5 cm or 10 cm deep) does not seem to influence bacterial community composition, implying that the dominant bacterial taxa were the same in the whole depth of the permafrost active layer.
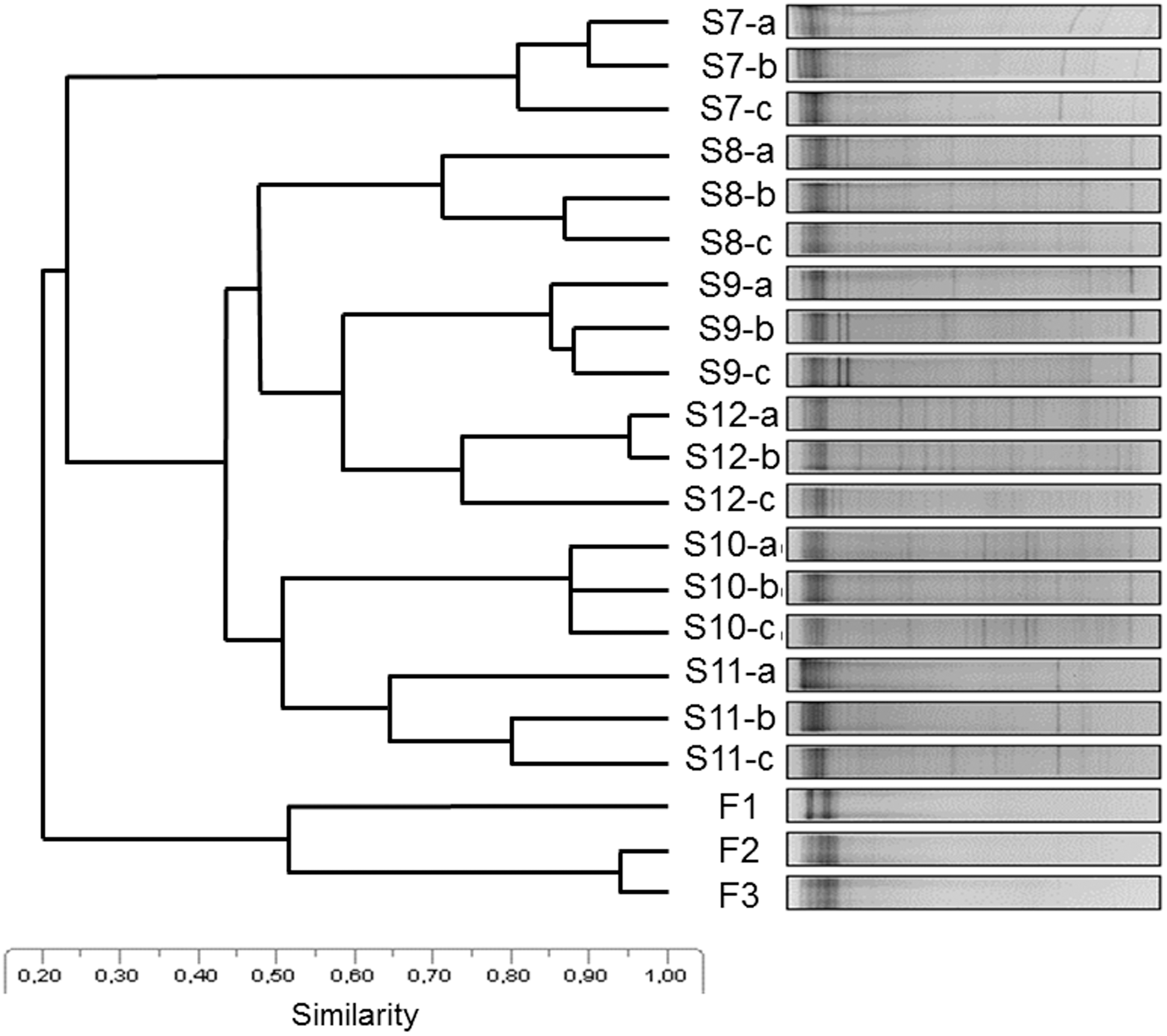
UPGMA similarity dendrogram of 16S rRNA gene-based DGGE molecular fingerprints showing similarities of bacterial community composition harbored by the water and sediment of the studied permafrost thaw pond on Ojos del Salado at 5900 m a.s.l. DGGE, denaturing gradient gel electrophoresis; UPGMA, unweighted pair group method with arithmetic mean.
3.3. Illumina MiSeq next-generation sequencing
For an in-depth analysis of the community composition of bacteria inhabiting the high-altitude permafrost-influenced environment, Illumina MiSeq NGS was performed on composite community DNA samples. Altogether, 447,253 sequences were obtained (24,372 to 94,052 sequences per sample), which based on Good's coverage values (Table 3) were enough to describe the bacterial diversity of the studied environment.
Summary of Next-Generation Sequencing Results
Number of phyla, genera (relative abundance of at least 1%), and sequences in each sample along with OTU-based beta diversity indices.
OTUs, operational taxonomic units.
Community composition of bacteria revealed with NGS from the water and sediment samples was remarkably different. The number of observed OTUs (Sobs) showed that species richness was lower in the water sample compared with the lake and permafrost sediment samples (Table 3). The Shannon index (H) implied the presence of a more complex bacterial community in the sediments, especially in samples derived from the deeper layers (S8 and S10). This might be the effect of increase in water activity with depth: evaporation mainly affects the surface of sediments, and only a few centimeters deeper the sediment can remain moist, resulting in a more favorable environment for microbial life. Alternatively, intensive solar radiation affecting the surface can also be a limiting factor for bacterial growth.
Although glaciers are not represented in the Dry Andes, high-altitude aquatic environments are often connected to them in other regions. Based on Shannon diversity indices, bacterial communities harbored by the Ojos del Salado thaw pond were more complex than those reported from the periglacial lakes of Mount Everest (Liu et al., 2006).
The results of the phylogenetic analysis, including a UPGMA dendrogram based on Bray–Curtis similarity indices showing relationships of bacterial communities harbored by the studied water and sediment samples, are summarized in Fig. 5. Similar to the dendrogram constructed based on DGGE molecular fingerprints, the water (F1) and sediment samples (S7–S12) were distinctly separated, while bacterial communities in the pond sediments (S7 and S8) showed the highest similarity to each other and grouped together with the upper layer permafrost sediment samples (S9 and S11).
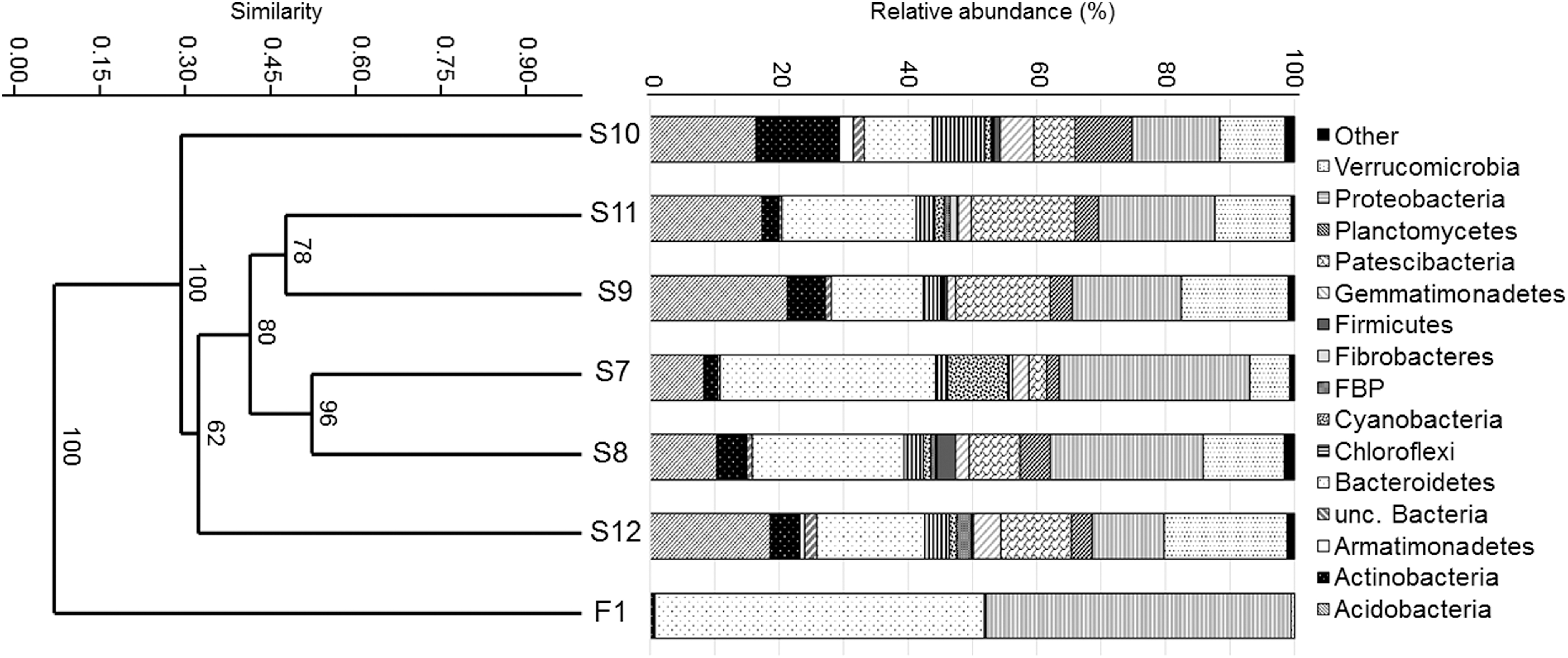
Relative abundance of phyla revealed from the water (F1) and permafrost sediment (S7–S12) samples of the permafrost thaw pond of Ojos del Salado at 5900 m a.s.l. along with the UPGMA similarity dendrogram constructed based on Bray–Curtis similarity.
The most surprising result regarding the phylogenetic affiliation of the sequences was the high abundance of hitherto unknown species and candidate taxa in this remote environment. Applying the 16S rRNA gene sequence similarity of 97% to delineate species (Stackebrandt and Goebel, 1994), the ratio of sequences affiliated with uncultured, unclassified, or candidate genera was between 46% and 79% in the analyzed sediment samples and 12% in the water sample.
Many of the detected uncultured or unclassified genera were dominant in the communities, belonging to phyla Patescibacteria, Acidobacteria, Actinobacteria, Bacteroidetes, Chloroflexi, Gemmatimonadetes, Planctomycetes, Proteobacteria and Verrucomicrobia. This corroborates the results of a preliminary analysis with 16S rRNA gene clone libraries (Aszalós et al., 2016) where most molecular clones belonged to hitherto uncultured taxa and confirms the idea that various habitats in the remote area of the Ojos del Salado volcano harbor hitherto unknown microorganisms. Sequences affiliated with the recently described phylum Abditibacteriota, formerly candidate phylum FBP (currently consisting of one described species, obligate oligotrophic Abditibacterium utsteinense) (Tahon et al., 2018), were also detected (S8, S11, and S12) along with members of the candidate genus Udaeobacter (Verrucomicrobia), a taxon known to be ubiquitous to soil environments (Brewer et al., 2017).
It is important to note the high proportion of hitherto uncultured species and candidate phyla detected by other researches carried out in astrobiologically relevant regions of the Dry Andes (Costello et al., 2009; Lynch et al., 2012; Solon et al., 2018).
The water was dominated by members of phyla Bacteroidetes and Proteobacteria, and only a few sequences of Acidobacteria, Actinobacteria, and Verrucomicrobia were detected. From the sediment samples, phylotypes related to phyla Acidobacteria, Verrucomicrobia, Patescibacteria, Actinobacteria, Planctomycetes, Chloroflexi, Cyanobacteria, Firmicutes, Armatimonadetes, Fibrobacteres, Gemmatimonadetes, and Abditibacteriota were detected. Nevertheless, there are great differences between bacterial communities of other permafrost sediments regarding minor taxa.
On the PCA plot, bacterial community structures in water (F1) and permafrost active layer sediments (S9–S12, Fig. 6) separated completely. Along both principal components, three groups were formed. PC1 was responsible for almost 90% of the variance between bacterial community compositions. According to the biplot, PC1 correlated with relative abundance of phyla Bacteroidetes and Proteobacteria. This principal component separated bacterial communities of the water (F1) from the lake sediment (S7 and S8) and permafrost sediment (S9, S10, S11, and S12), implying that the presence of water might be a driving force to cause differences in relative abundance of Proteobacteria and Bacteroidetes.
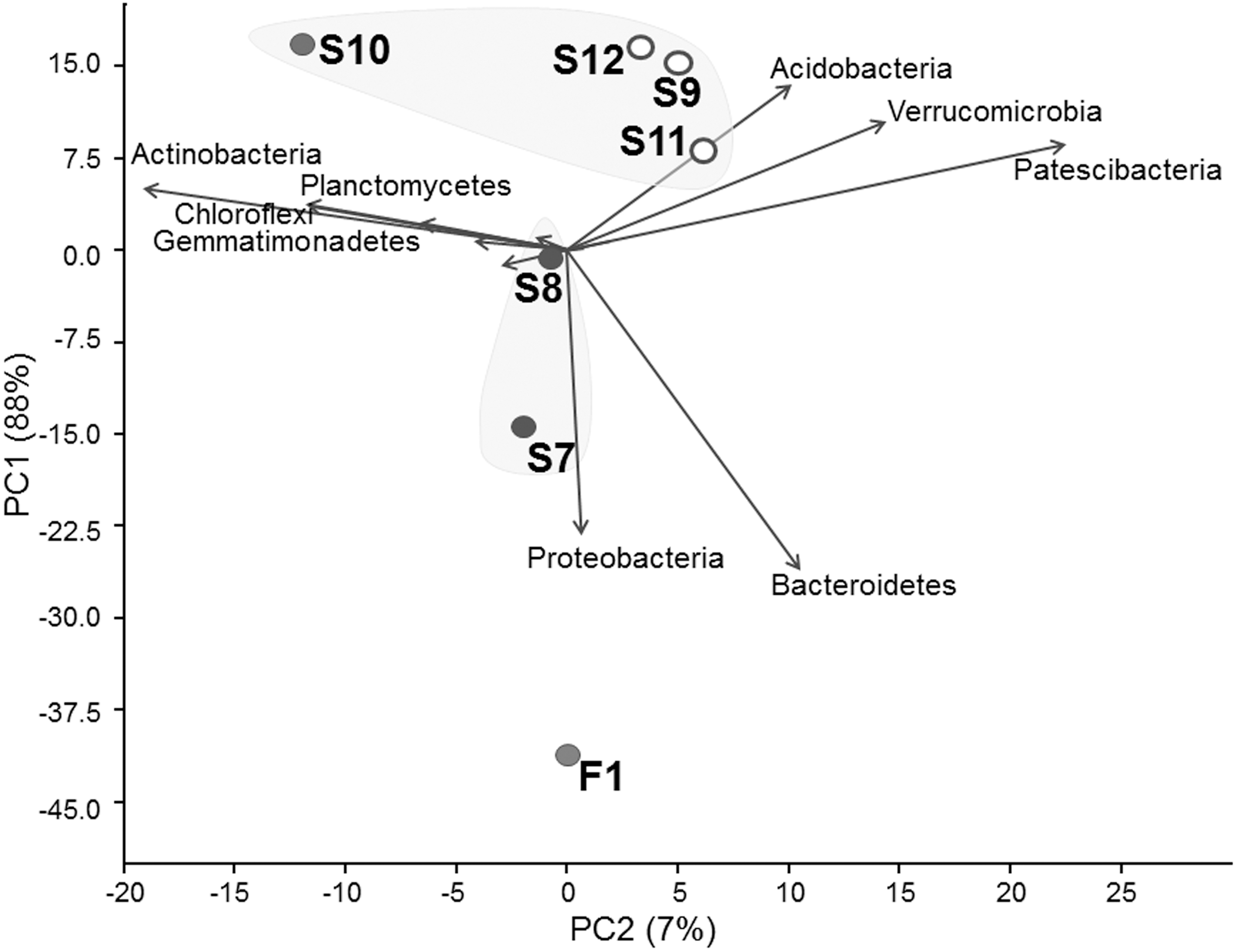
Principal component analysis of bacterial communities detected from the water (F1) and permafrost sediment (S7–S12) samples of the permafrost thaw pond of Ojos del Salado at 5900 m a.s.l. The biplot shows the main phyla contributing to variance between communities.
Along PC2, surface permafrost formed a distinct group (S9, S11, and S12), from which subsurface sediment (S10) and bacterial communities of the lake (F1, S7, and S8) separated. The biplot shows that several phyla (mainly Actinobacteria, Planctomycetes, Acidobacteria, Verrucomicrobia, and Patescibacteria) corresponded to the variance along PC2; however, this component accounted for only 8% of the variance.
Phylogenetic diversity revealed by NGS presented in this study was similar to that reported in a previous 16S rRNA gene clone library analysis (Aszalós et al., 2016). First, due to the better resolution of NGS, the presence of some minor phyla, for example, Patescibacteria, Planctomycetes, Chloroflexi, Cyanobacteria, Abditibacteriota, Firmicutes, Armatimonadetes, and Fibrobacteres, was also revealed; on the other hand, the presence of Aquificae was only detected with 16S rRNA gene clone libraries from a sample taken in 2014. In comparison, permafrost microbial communities in the Canadian Arctic slightly differed from those detected in the thaw pond of Ojos del Salado since members of Acidobacteria were present only in a moderately acidic permafrost wetland in Nunavut (Wilhelm et al., 2011), but were not detected in other Nunavut permafrosts (Steven et al., 2006) or the Canadian subarctic permafrost thaw ponds (Comte et al., 2016).
Similar to our results, most studies on bacterial communities in the Dry Andes at altitudes higher than 5000 m a.s.l. revealed simplified communities mirroring the low organic material contents in the studied sites (Costello et al., 2009; Lynch et al., 2012; Solon et al., 2018). This feature is especially important because low organic material content resembles martian environments (Domagal-Goldman et al., 2016).
Although there are only a few in-depth analyses of high-altitude sediments in the Dry Andes at similar altitudes of volcanoes Llullaillaco and Socompa, most studies described the lack of Cyanobacteria in cold and dry sediments (Costello et al., 2009; Lynch et al., 2012), while their presence is often detected in warmer settings, for example, fumarolic sediments (Solon et al., 2018). However, Cyanobacteria were detected in low abundance in all the sediment samples, with the highest ratio in the pond sediment (S7). Among Cyanobacteria, Nodosilinea nodulosa PCC-7104 was predominant in the sediment of the pond (S7, 9%). This strain was originally isolated from a marine rock in Montauk Port, NY, and capable of N fixation (Perkerson et al., 2011).
Other autotrophic bacteria were also detected, for example, members of the chemolithoautotrophic genus Hydrogenophaga that is capable of oxidation of H2 and utilizes CO2 as a sole carbon source (Kämpfer et al., 2005).
Regarding lower taxonomic levels, there were only 12 genera with at least 1% relative abundance in the water sample, with Massilia (25.5%) and Flavobacterium (36.0%) being the most abundant. The rest of the genera were Algoriphagus (1.3%), Hymenobacter (2.2%), Spirosoma (1.6%), Brevundimonas (4.6%), Porphyrobacter (2.8%), Sphingorhabdus (6.7%), and Hydrogenophaga (1.1%) and two uncultured genera belonging to phyla Bacteroidetes and Proteobacteria (4.4% and 2.9%, respectively).
Bacterial communities of the sediment samples consisted of 21–29 genera. Altogether, six of them (Blastocatella and Sphingomonas and four unclassified genera) had at least 1% relative abundance in all six sediment samples. Besides these, the genera with the highest relative abundance (>4%) in one or more sediment samples were Fluviicola (10%), Algoriphagus (5%), Luteolibacter (5%), and uncultured genera of various phyla.
Many of the detected genera are described from oligotrophic environments (Sphingorhabdus, Brevundimonas, or the formerly mentioned Abditibacterium), and others are known to withstand arid circumstances (Blastocatella). Many of the genera were known to be present in polar or periglacial environments (members of Massilia) (Shen et al., 2015). The genus Algoriphagus contains one species, the psychrophilic Algoriphagus ratkowskyi isolated from sea ice and saline lake microbial mats, which shows growth at temperatures between −2°C and 25°C (Bowman et al., 2003). Members of the strictly aerobic Hymenobacter (Hirsch et al., 1998) genus are often isolated from dry and cold environments, for example, Antarctic Dry Valleys (Hirsch et al., 1998).
Several genera revealed by Solon et al. (2018) on Llullaillaco and Socompa volcanoes in the Dry Andes were also detected on the Ojos del Salado (Hymenobacter, Segetibacter, Sphingomonas, Blastocatella, and genera belonging to Chitinophagaceae or Gemmatimonadaceae), although some of the most abundant genera detected by us were not reported from Socompa or Llullaillaco sediments. This could be the effect of higher pH values of water present in this lake than is Socompa and Llullaillaco environments.
It is noteworthy to compare the revealed cultivable diversity with the results of NGS. Although most of the cultivated taxa were detected by NGS, it is surprising that the presence of Arthrobacter, one of the most commonly cultivated genera in periglacial environments, was only sporadically revealed in the samples by NGS. This raises the question whether this taxon can be active, and present in low abundance, or inactive in the environment.
4. Conclusions
This is the first study that reports on bacterial communities of a high-altitude, permafrost thaw pond, considered to be a Mars analog lake, revealed by a combination of cultivation and NGS. The permafrost thaw pond of the Ojos del Salado volcano harbors simplified bacterial communities, similar to other high-altitude, permafrost-influenced aquatic environments. The dominance of oligocarbophilic and psychrophilic species revealed from the studied remote lake water and sediments indicates that these bacteria are adapted to the harsh environmental circumstances of the Dry Andes.
In addition, the high proportion of hitherto uncultured bacteria revealed by NGS shows the importance of cultivation-based analysis of extremophiles for a future potential biotechnological application. Moreover, due to the rare combination of environmental settings, a more detailed analysis of bacterial communities in this environment would provide astrobiologically relevant information about the supposed lakes on the Red Planet formed by ephemeral melting.
Footnotes
Acknowledgments
Sampling in Chile was performed with the permission of CONAF (Corporación Nacional Forestal, Chile). The authors would like to thank Tamás Felföldi at ELTE Eötvös Loránd University for his help in next-generation DNA sequencing and Ákos Kereszturi for his helpful comments and advice for preparing the manuscript.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this research.