
Abstract
Hot spring environments are commonly dominated by silica sinters that precipitate by the rapid cooling of silica-saturated fluids and the activity of microbial communities. However, the potential for preservation of organic traces of life in silica sinters back through time is not well understood. This is important for the exploration of early life on Earth and possibly Mars. Most previous studies have focused on physical preservation in samples <900 years old, with only a few focused on organic biomarkers. In this study, we investigate the organic geochemistry of hot spring samples from El Tatio, Chile and the Taupo Volcanic Zone, with ages varying from modern to ∼9.4 ka. Results show that all samples contain opaline silica and contain hydrocarbons that are indicative of a cyanobacterial origin. A ∼3 ka recrystallized, quartz-bearing sample also contains traces of cyanobacterial biomarkers. No aromatic compounds were detected in a ∼9.4 ka opal-A sample or in a modern sinter breccia sample. All other samples contain naphthalene, with one sample also containing other polyaromatic hydrocarbons. These aromatic hydrocarbons have a thermally mature distribution that is perhaps reflective of geothermal fluids migrating from deep, rather than surface, reservoirs. These data show that hot spring sinters can preserve biomolecules from the local microbial community, and that crystallinity rather than age may be the determining factor in their preservation. This research provides support for the exploration for biomolecules in opaline silica deposits on Mars.
1. Introduction
Biosignatures are distinctive suites of durable textural, mineralogical, and chemical indicators of life (Bock and Goode, 1996; Farmer, 1999, 2000; Farmer and Des Marais, 1999; Jones et al., 2001a; Jones and Renaut, 2003; Philippot et al., 2007; Glikson et al., 2008; Preston et al., 2008; Van Kranendonk et al., 2008; Campbell et al., 2015a, 2015b; Westall et al., 2015a, 2015b; Morag et al., 2016; Djokic et al., 2017). The search for biosignatures on Mars has been guided by our knowledge of preservational pathways on Earth (Walter and Davies, 1999). For macroscopic fossils, microfossils, and many types of textural biosignatures, silica is the best preserving mineral, effectively entombing organisms and their constructed edifices within the fine matrix of various silica phase minerals (e.g., opal-A, opal-CT/C, microcrystalline quartz or chert: Walter and Des Marais, 1993; Guido et al., 2010; Brasier et al., 2011; Campbell et al., 2015b; Tarhan et al., 2016). For example, the preservation of ancient life signatures in Archean rocks from the Pilbara Craton of Australia is overwhelming in cherts (e.g., Sugitani et al., 2009, 2010, 2013) or in diagenetically silicified carbonates (e.g., Hofmann et al., 1999). Carbonates from throughout the geological record, on the other hand, typically do not preserve microfossils unless silicified early in diagenesis (Guido and Campbell, 2012). This is because of the high crystallization energy of carbonate, which has been reported to cause the destruction of prokaryotic cell-sized fossils, as well as organic matter readily becomes remineralized to carbonate through diagenesis (e.g., Altermann and Kazmierczak, 2003).
Siliceous hot spring deposits, or sinters, precipitate due to the rapid cooling and evaporation of silica-saturated fluids, as well as from biomediation (e.g., Kaur et al., 2008). As with Archean cherts, it has been well documented that young (i.e., Phanerozoic) sinters preserve not only textural biosignatures but also microfossils (e.g., Campbell et al., 2001, 2015a; Jones and Renaut, 2001a; Konhauser et al., 2001; Ruff and Farmer, 2016). To date, however, most studies on sinter have focused on textural preservation, with only a small proportion having evaluated the preservation of organic geochemical biomolecules from the microbial communities that inhabited the silicifying hot spring waters (e.g., Siljeström et al., 2017; Sanchez-Garcia et al., 2019). Even fewer studies have prospected for fossilized biomolecules encased in well-preserved mineral deposits (i.e., those with opaline silica: Kaur et al., 2008, 2011; Gibson et al., 2014; Campbell et al., 2015a). To date, the oldest identified biomolecules in sinters are 900 years old (Kaur et al., 2008; Hays et al., 2017).
1.1. Astrobiological implication for siliceous hot spring sinters
Siliceous hot spring sinters are promising targets in early life studies and in the search for life on Mars. When silica first precipitates to build up a sinter deposit, it comprises noncrystalline (amorphous) opal-A. As sinter undergoes diagenesis, opal-A transforms to paracrystalline opal-CT, followed by opal-C, and finally to microcrystalline quartz, or microquartz (Herdianita et al., 2000). Although age can sometimes be used as an estimate of the crystallinity, these silica stages are not always spatially or temporally uniform. The degree of crystallinity among sinter deposits can depend on other factors such as variable fluid components and the degree of fluid mobilization (Campbell et al., 2001).
Considering this, the presence of opaline silica observed on Mars suggests that minimal diagenesis/recrystallization has occurred, for at least some inferred hydrothermal deposits, such as Columbia Hills (Squyres et al., 2008; Ruff et al., 2011). H2O-poor opaline silica also has been detected at Nili Patera on Mars, where Skok et al. (2010) suggested that the presence, distribution, and chemical composition of this silica on a mound immediately adjacent to a volcanic center indicates a hydrothermal origin.
Since modern to ancient (∼3.5 Ga) terrestrial hot spring systems on Earth that display varying degrees of crystallization preserve signs of microbial life (e.g., Ruff and Farmer, 2016; Djokic et al., 2017, 2020; Ruff et al., 2019), the well-preserved martian hydrothermal deposits become promising astrobiological targets. Hot springs have also been suggested as candidate environments for the origin of life on Earth (Damer and Deamer, 2015; Deamer and Georgiou, 2015; Van Kranendonk et al., 2017). If life originated in a terrestrial hot spring setting, there are implications for the search for life beyond Earth.
NASA's Mars life exploration strategy, which includes a requirement to “follow the water,” is reflected by the Mars Science Laboratory (Grotzinger et al., 2014), which has focused its efforts on determining habitable environments by investigating fluvial and lacustrine settings in Gale crater. These environments will continue to be studied with the Mars2020 mission in Jezero crater (announced by NASA, 2018). So far, results from Gale crater have revealed the presence of organic molecules, but without definitive proof that any of these compounds have a biological origin (Freissinet et al., 2015; Eigenbrode et al., 2018).
In addition, laboratory simulation experiments (Royle et al., 2018) show that chlorinated hydrocarbons detected in Gale crater likely became chlorinated during analysis, and that these compounds have a low likelihood of survival under martian conditions. Further, there is no information on the preservation of organic molecules in silica from Mars, and, indeed, this topic is not well studied, even on Earth.
Research into the preservation of fossilized biomolecules from microbial cell membranes has primarily concentrated on data from the opaline sinter terrace of Champagne Pool in the Taupo Volcanic Zone (TVZ), New Zealand (Kaur et al., 2008; Gibson et al., 2014), which occupies a 900-year-old hydrothermal explosion crater (Lloyd, 1959). However, these data do not necessarily reflect other hydrothermal systems, because the primary controls on lipid preservation in hydrothermal sinters have been suggested to be thermal alteration, high temperatures, and variable pH values of the hot spring waters, which vary from pool to pool (Pancost et al., 2006). This highlights the need for not only cross-locational studies but also ones that span a range of ages, pH, and silica mineral phase crystallinity.
Here, we investigate preserved biomolecules in samples of siliceous hot spring sinters from El Tatio, Chile (Fernandez-Turiel et al., 2005), and from the TVZ, New Zealand (Houghton et al., 1995). Samples analyzed here span an age range from actively forming, to fossil deposits (∼9.4 ka), and from rocks that preserve primary opal-A to those that are recrystallized to microcrystalline quartz. This study examines the potential for sinters to preserve biomolecules of their entombed microbial community over time, and it provides an additional context for the exploration of biosignatures in hot spring deposits on Mars.
2. Sample Acquisition
2.1. El Tatio, Chile
Three modern opal-A sinter samples were collected from the El Tatio hydrothermal field in the high Andes of northern Chile (22°20′S, 68°01′W), which experiences high evaporation and low precipitation rates at an elevation of ∼4300 m at the edge of the Atacama Desert (e.g., Nicolau et al., 2014). All samples were collected with permission issued in 2017 by the presidents of the indigenous communities of Caspana (Jilberto Anza) and Toconce (Abel Bartolo), which oversee El Tatio.
Sample ET2-1 (22°20′43.71″S 68°00′43.97″W) is a nodular sinter with poorly developed digitate structures collected from a discharge channel with green and nearly black biofilms (Fig. 1A, D). Its location was ∼7 m downstream of a ∼1-m-diameter spring mound discharging water with a temperature of ∼77°C and near neutral pH, although cooled to <40°C at the position of the sample.

Sample locations at El Tatio, Chile.
Sample ET3-2 (22°20′32.74″S 68°00′46.90″W) is also a nodular sinter with poorly developed digitate structures that came from a dry, but recently flowing, discharge channel with no apparent biofilm (Fig. 1B, E). Temperature and pH were measured at the source, a geyser pool ∼5 m from the sampling location, with ∼85°C water (boiling point at the elevation of the location) and near neutral pH.
Sample ET4-4 (22°19′56.51″S 68°00′41.93″W) is a sinter breccia located ∼10 m downstream of an active geyser producing shallow discharge flows at near neutral pH (Fig. 1C, F). The sample was broken off from a larger piece of loose sinter breccia (∼30-cm-long dimension) resting on damp regolith.
2.2. The TVZ, New Zealand
The TVZ is a 60-km-wide by ∼300-km-long rifted arc basin, in North Island, New Zealand (Wilson and Rowland, 2016). It is the largest and most active rhyolitic magmatic system on Earth, and it is accompanied by more than 20 large hydrothermal fields (Houghton et al., 1995). Four samples from the TVZ were analyzed. All samples were collected with permission from the landowners and/or the New Zealand Department of Conservation.
First, a modern conical tufted, opal-A sinter (Tokaanu) was collected from the Hipua-Waihi field, ∼50 km SW of Taupo. This field consists of ∼5000 m3 of sinter deposited over 6000 years (Soto et al., 2019). Conical tufted samples are found in the mid-apron, where temperatures were between 45°C and 65°C (Fig. 2).
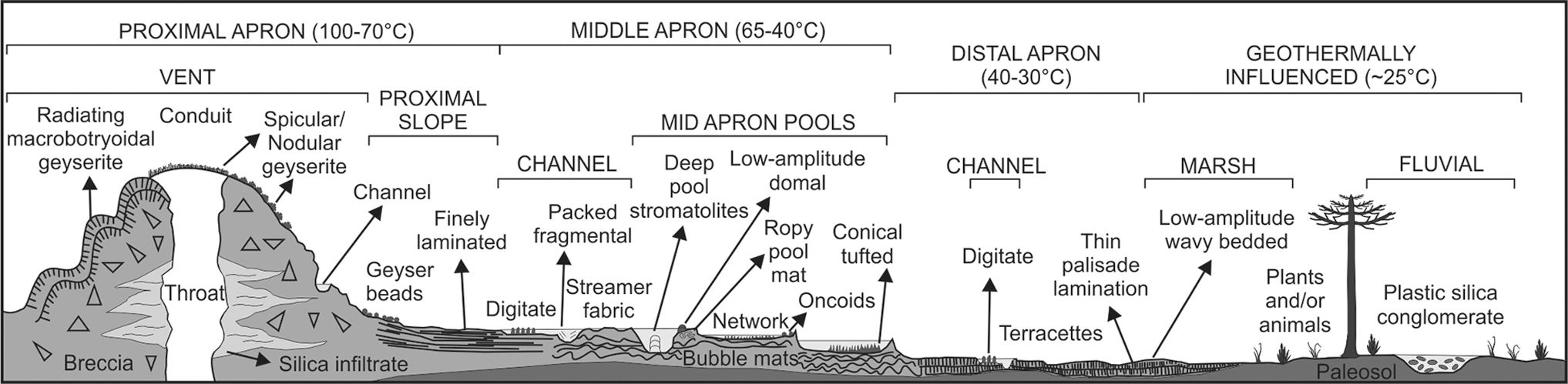
Schematic cross-section of a near-neutral pH, alkali chloride, Si-bearing hot spring. The connected sinter morphologies and temperature gradients are shown. Figure modified from Campbell et al. (2015b).
Second, a Subrecent sample from Te Puia is an opal-A sinter with distinct microbial palisade texture (e.g., Campbell et al., 2015a) and is from a part of the terrace with a pH 8.5–9.15 (Jones et al., 2001b). Palisade textures occur when fine filaments are densely packed in vertical pillar structures (Campbell et al., 2015b). Palisade textures are typically formed from photosynthetic cyanobacteria, which are associated with the low-temperature environments in the distal apron (Fig. 2; Campbell et al., 2015b).
Third, a ∼9.4 ka sample of packed fragmental opal-A sinter was collected from Mangatete, which is a fossil hydrothermal field of ∼2 km2 area located in the Taupo Fault Belt (Drake et al., 2014). Packed fragmental sinter (PFS) textures are known from the Jurassic in Patagonia, and even the Paleoarchean of Australia (Djokic et al., 2020). Modern PFS constitutes a porous sinter matrix with stacked, broken sheets of irregular, angular, mostly highly tabular, silicified microbial fragments, and which has been interpreted to form where fast flowing, warm (50°C), alkali-chloride thermal streams break up silicified microbial mats (Guido and Campbell, 2011; Drake et al., 2014).
Finally, a ∼3.5 ka sample comes from a sinter buttress in the Te Kopia hydrothermal field (38°24′S 176°13′). It is composed of microcrystalline quartz and contains microbial sinters that have a streamer fabric and microbial palisade fabric (Martin et al., 2000), both of which are observed from channels and in the distal sinter apron at temperatures of <40°C (Fig. 2; Campbell et al., 2015a).
3. Methods
3.1. Sample preparation
The samples in this study, except the one from Te Puia, were broken off from outcrop by use of a geological hammer and placed in sealed plastic bags until analysis. The Te Puia sample was obtained from historical collections stored in rock trays in the University of Auckland petrological collection.
A mixture of dichloromethane (DCM, Honeywell) and methanol (MeOH, Honeywell) (9:1 v/v) was used to extract surficial contaminants before crushing, as these samples were not collected cleanly (e.g., George et al., 2007, 2008; Sherman et al., 2007). The samples were placed in beakers and covered with a mixture of DCM and MeOH (9:1 v/v). The beaker was sonicated for two cycles of 10 min and rested for 5 min in between. This outside rinse solution was reduced by a rotary evaporator until less than 2 mL remained and was then transferred by pipette into a 2-mL vial. The vial was spiked with three compounds as internal standards by adding 1 mL of a DCM solution (solution-1) containing about 117 ng each of anthracene-d10 (98 atom%D, Isotec), p-terphenyl-d14 (98 atom %D, Isotec), and tetracosane-d50 (98 atom %D, Isotec).
The outside rinse was then checked to ensure there were minimal contaminants by gas chromatography-mass spectrometry (GC-MS). Once the chromatograms obtained from the GC-MS were checked, the samples were cut with a diamond blade and crushed with a tungsten carbide ring mill crusher. Before cutting and crushing, a blank was performed on both the saw and the crusher. The outer 3–5 mm of the sample was cut off and not extracted, so as to minimize contamination from sample collection. These external offcuts from the samples were made into thin sections. Petrographic examination of thin sections was undertaken with a Nikon Eclipse Ci POL microscope in plain polarized light, with magnifications from 2× to 50× zoom at the University of New South Wales (UNSW).
3.2. Solvent extraction, fractionation, and derivatization
The rock powder was solvent extracted by using a mixture of DCM and MeOH (9:1 v/v) and sonication for 10 min, resting for 5 min, and sonication for a further 10 min (e.g., Hoshino and George, 2015; Hoshino et al., 2015). Between each sonication, the solution was stirred and left to resettle for 5 min. The solvent solution was then decanted into a round bottom flask, and the process was repeated twice more for each sample. The second and third solvent mixtures were added to the first for each sample.
Before rotary evaporation, a blank was performed and the machine was confirmed to be free of contamination. The collected solvent mixtures were reduced by rotary evaporation to ∼40 mL. The samples were centrifuged at 2000 rpm for 10 min to remove fine rock particles, and then the solution was decanted into a round bottom flask. If, after centrifugation, there were still fine particles suspended in the mixture, the mixture was filtered through silica gel (silica gel 60, 0.063–0.200 mm), which was activated at 120°C for >4 h before use.
The samples were fractionated on small scale silica columns in pipettes, using a mixture of hexane and DCM (4:1) to elute an apolar fraction. The apolar fraction was further fractionated into aliphatic and aromatic hydrocarbons by using hexane and the 4:1 solution. The hydrocarbon fractions were spiked with the internal standard solution-1, further evaporated by hot plate and nitrogen blow down, pipetted into a 100-μL vial insert, and finally analyzed by the GC-MS.
3.3. Gas chromatography-mass spectroscopy
The samples were analyzed by GC-MS using a two-dimensional gas chromatograph (Agilent 7890A) operating in one dimension, coupled to a Pegasus time-of-flight-mass spectrometer (GC × GC-ToFMS). The interface was held at 310°C, and the MS source temperature was 280°C. Fractions were injected (1 μL) in a split/splitless injector operating at 310°C in splitless mode onto a J&W DB5MS column (60 m × 0.25 mm i.d., 0.25 mm film thickness) coated with modified 5% phenyl 95% methyl silicone, with 1.5 mL/min He carrier gas flow. The GC column oven temperature was programmed from 40°C (hold for 2 min) to 310°C at 4°C/min and held for 40.5 min.
Aliphatic and aromatic fractions of an international external standard (North Sea Oil) were run by using the same program to aid component assignment. Peak areas were integrated with LECO Chromatof software, and they were quantified relative to the peak areas of the internal standards. This quantification was based on ng/g of rock (after French et al., 2015; Hoshino and George, 2015; Aharonovich and George, 2016). The limit of detection for n-alkanes was 0.5 ng/g, and for aromatic hydrocarbons it was 0.02 ng/g.
4. Results
4.1. Petrography
Thin-section petrographic analysis of sample ET2-1 from El Tatio reveals a polymict clastic sediment, containing fine-grained (up to 300 μm) clasts in an opal-A matrix (Fig. 3A). Some sand-sized clasts are themselves an aggregate of fine-grained clastic material, including angular volcanic glass shards, mica grains (biotite and white micas), and angular quartz and feldspar mineral fragments. These aggregate clasts are rimmed by uneven layers of opal (i.e., point a on Fig. 3A). Other parts of the sample contain multiple, thin, opaline silica rinds that coat bundles of aligned, clear, completely silicified microbial filaments. The opaline silica rind coats, and abuts against, a matrix of opaline silica packed with diatoms (point b on Fig. 3B) and individual sand grains. Well-preserved microbial filaments with orange pigment (point c on Fig. 3B) appear to cut across the boundary between textural domains within the sample, suggesting that these less well-oriented orange filaments grew after those incorporated within the coated grains.
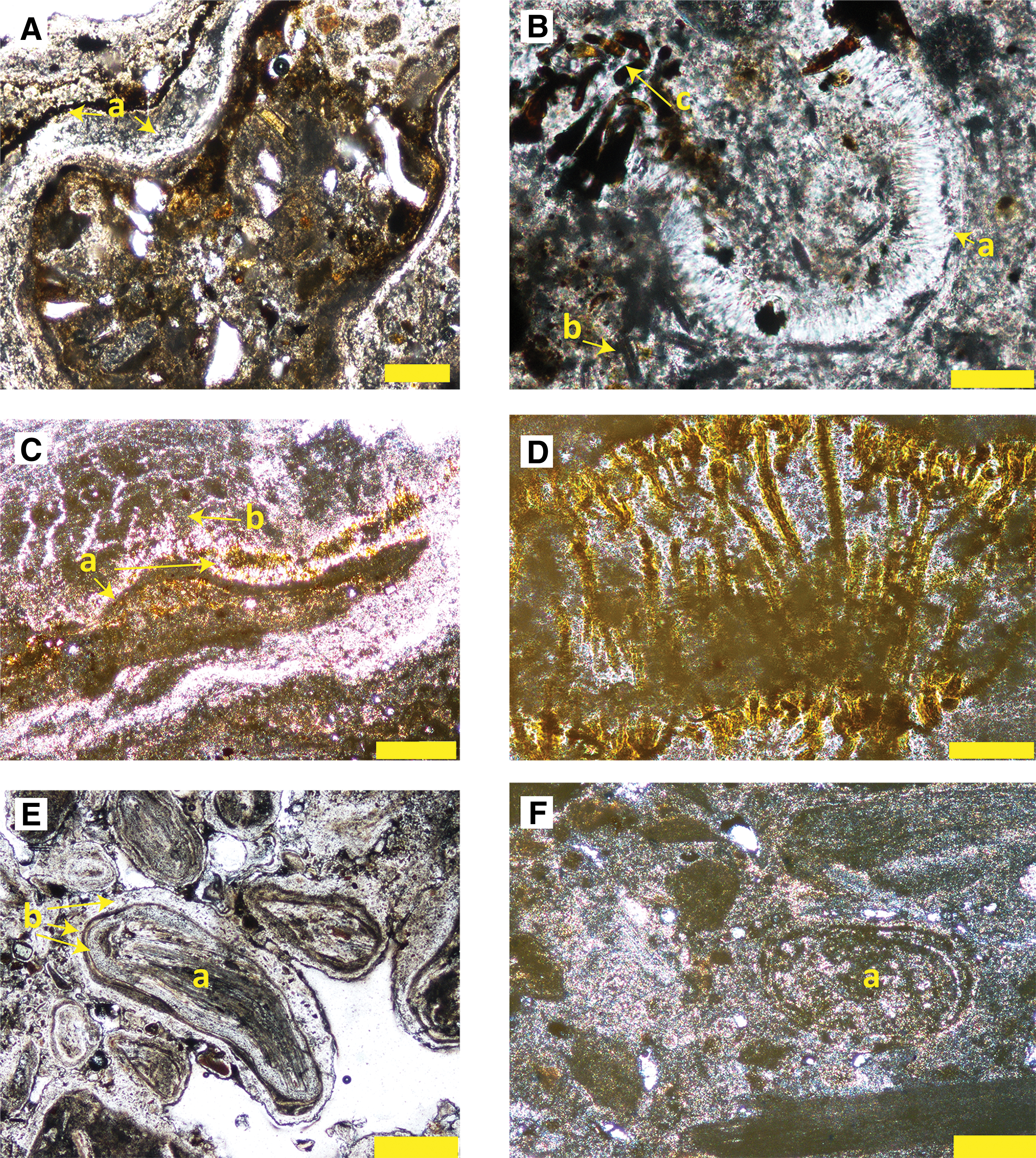
Thin-section photomicrographs of analyzed samples, in plane polarized light.
Sample ET3-2 from El Tatio contains mottled opaline silica layers with microbial palisade texture comprising bright orange cyanobacterial filaments aligned vertically along bedding planes (point a in Fig. 3C, D). Microbial palisade layers are overlain by digitate columnar microstromatolites that are characterized by dark opaline silica in the lower parts of the columns (point b in Fig. 3C), and by fine-scale laminations in the upper parts of the columns.
Sample ET4-4 from El Tatio contains densely packed and poorly sorted oncoids that are 250–500 μm in diameter, which form by wetting and drying cycles of mid-temperature thermal spring discharge on the mid- to distal sinter apron (c.f. Jones and Renaut, 1997; Guido and Campbell, 2011). These show concentric laminations with both angular and rounded nuclei. Some of the oncoids are discoidal, whereas others have lobate edges. Some elongated oncoids have cores composed of aligned microbial filaments that are coated by a series of very thin opaline silica coatings (point b on Fig. 3E). These rinds include a granular, mottled lamina coated by an irregular, dark brown rim, and then by a lighter lamina with detrital infill, which is again coated by a thin dark rind.
The Mangatete sample has a mottled, gray, siliceous matrix with layered, broken, silicified microbial sheets that contain flat lamination throughout, as well as sub-rounded to lobate oncoids (point a on Fig. 3F). The rock matrix is recrystallized to a fine-grained, mottled, silicified patchwork incorporating volcaniclastic silt and sand grains. The embedded oncoids constitute a series of concentric coatings that are preserved despite recrystallization.
The Te Kopia sample is a sinter of fine-grained opaline silica in which no distinctive textures were visible in thin sections. Thin sections were not made for the samples from Te Puia and Tokaanu, as there was insufficient material after organic geochemical analysis.
4.2. Organic geochemistry
4.2.1. n-Alkanes
A series of n-alkanes were detected in all samples (Fig. 4; Supplementary Table S1). The two modern digitate samples from El Tatio (ET2-1 and ET3-2) have bimodal n-alkane distributions. The first mode is from n-C13 to n-C20, with strong enrichments of n-C15 relative to n-C14 and n-C16, and n-C17 relative to n-C16 and n-C18. The second mode is from n-C20 to n-C30, with a very strong odd-over-even carbon number predominance. In ET2-1, the maximum is at n-C30; whereas in ET3-2, the maximum is at n-C17. El Tatio sinter breccia (ET4-4) has n-alkanes ranging from n-C15 to n-C32. This sample has similar n-C15 and n-C17 enrichments relative to the neighboring alkanes, a maximum at n-C27, and a strong odd-over-even carbon number predominance. Carbon preference indices (CPI22–32) for all three El Tatio samples are 1.5–1.8 (Table 1).
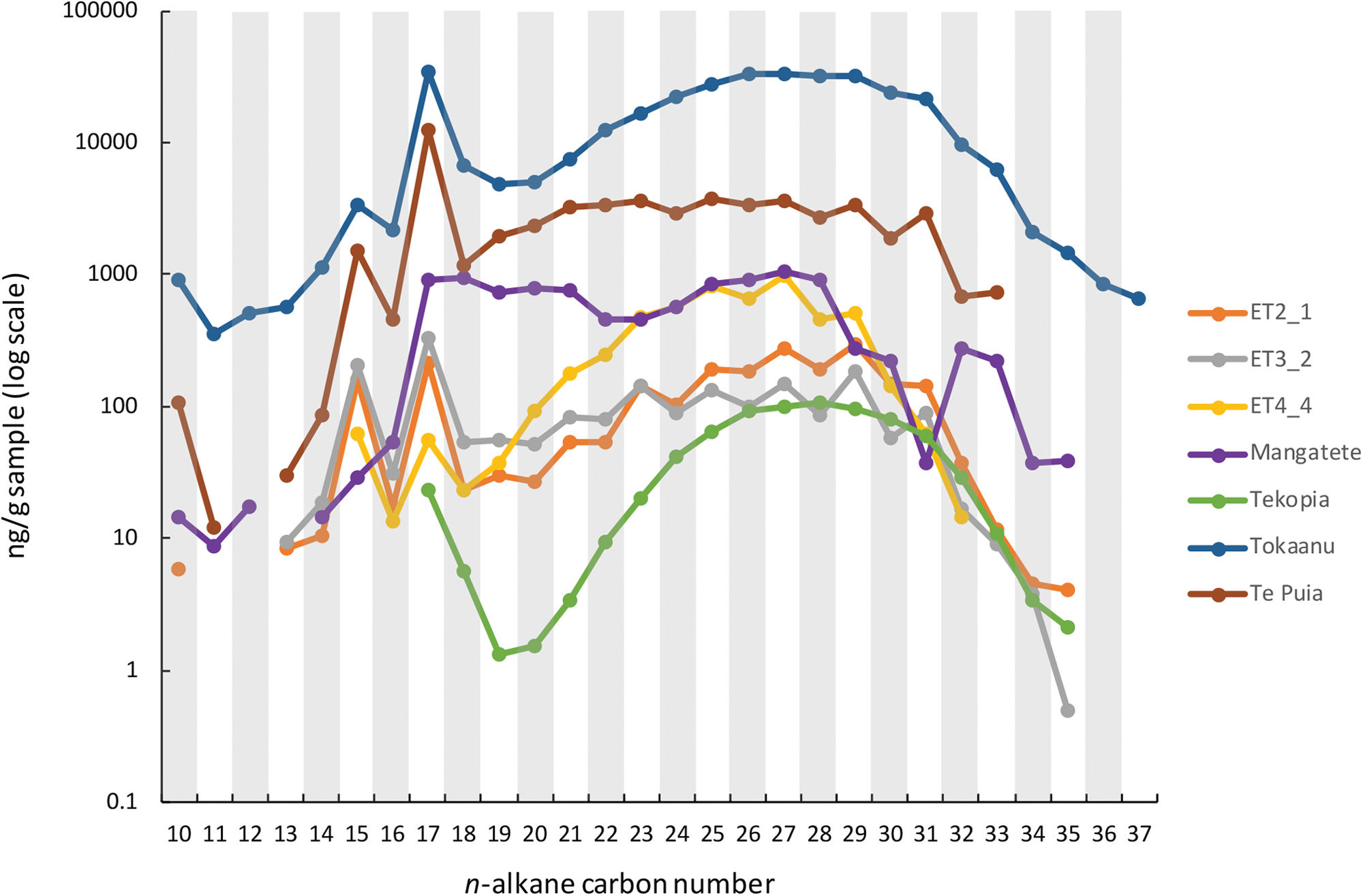
Amount (ng/g sample) and distribution of n-alkanes in all samples from this study. Procedural blanks yielded no n-alkanes, except a very small amount of the C10 n-alkane. Color images are available online.
Weights of Samples and Aliphatic Hydrocarbon Ratios for the Samples from El Tatio (Chile) and the Taupo Volcanic Zone, New Zealand
Pr = pristane, Ph = phytane, CPI22–32 = carbon preference index of n-alkanes: (2 × (C23 + C25 + C27 + C29 + C31))/(C22 + 2 × C24 + 2 × C26 + 2 × C28 + 2 × C30 + C32).
nd = not detected.
The four New Zealand samples have similar bimodal n-alkane distributions as those from El Tatio, with n-alkanes ranging from n-C10 to between n-C33 and n-C37. In the fossilized Mangatete and Te Kopia sinter samples, n-C13 was not detected, and n-C12 was not detected in the Te Puia sample. Te Puia has a strong odd-over-even predominance similar to the El Tatio samples, whereas the ∼9.4 ka Mangatete sample has a slight even-over-odd predominance (Table 1). Both the modern Tokaanuu and the ∼3.5 ka Te Kopia samples have CPI22–32 near unity. The two modern samples (Tokaanu and Te Puia) have maxima at n-C17, whereas the maximum in the ∼9.4 ka Mangatete sample is n-C27, and in the ∼3.5 ka Te Kopia sample it is n-C28. The Te Kopia sample is a recrystallized, microcrystalline sample with the lowest total abundance of n-alkanes among all samples (Fig. 1). In contrast, the older ∼9.4 ka Mangatete sample that preserves opal-A has an intermediate abundance of n-alkanes relative to the other samples (Fig. 4).
All samples are enriched in n-C17 compared with neighboring n-alkanes, as measured by the 17/(16 + 18) n-alkane parameter (Table 1; Hoshino and George, 2015), although the strength of that enrichment varies between samples. This n-C17 enrichment is strongest in the digitate structures from El Tatio, and in the modern samples from New Zealand (Te Puia and Tokaanu) (Table 1).
The isoprenoids, pristane (Pr) and phytane (Ph) were detected in all samples except Te Kopia, and phytane was not detected in ET3-2. All samples have Pr/Ph ratios of <1.1, with the ∼9.4 ka recrystallized Mangatete sample having the lowest ratio (0.4; Table 1).
4.2.2. Methylalkanes
Monomethylalkanes (MMAs) were detected in all samples. ET3-2 has MMAs from C12 to C32 (Fig. 5), whereas ET4-4 and ET2-1 have MMAs from C13 to C32. MMAs are present in the Tokaanu sample from C13 to C33, in Te Puia from C14 to C32, in Mangatete from C16 to C31, and in Te Kopia from C23 to C32. For all samples that were found to contain methylheptadecanes (MHeD), it is the mid-chain MMAs (especially including 6-MHeD and 5-MHeD) that are more predominant than 2-MHeD and 3-MHeD (Fig. 5).
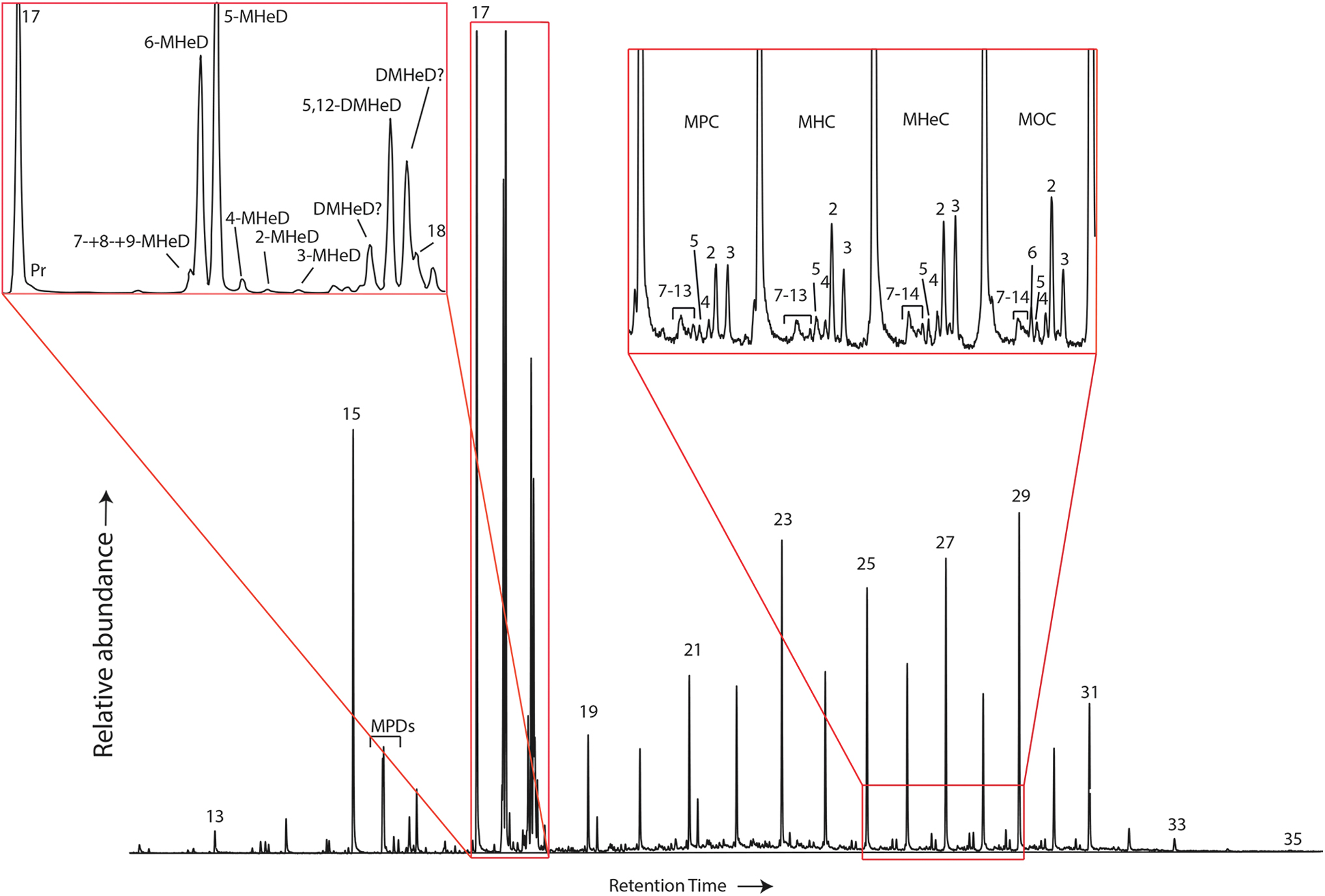
Partial m/z 57 mass chromatogram of sample ET3-2, showing the n-alkane range from C12 to C35, with zoomed-in segments for monomethylalkanes, including the MHeD, and the C26–C29 MPC, MHC, MHeC, and MOC. Numbers in the insets refer to methyl positions on the methylalkanes. DMHeD = dimethylheptadecane; MHC = methylhexacosanes; MHeC = methylheptacosane; MHeD = methylheptadecanes; MPC = methylpentacosanes; MPD = methylpentadecane; MOC = methyloctacosanes. Color images are available online.
The distribution of MHeDs varies between the samples (Fig. 6). The sample ET3-2 is dominated by 5-MHeD with abundant 6-MHeD, whereas the Mangatete sample contains 6-MHeD as the most abundant, followed by 5-MHeD and the peak due to the co-eluting mid-chain isomers 7-, 8-, and 9-MeHeD.

Histogram of mono MHeD distribution in all samples from this study. Abundance is relative to n-C17 (100). Color images are available online.
The most abundant MHeD peak in the ET2-1, ET4-4, and Te Puia samples is due to the mid-chain isomers, followed by 6-MHeD (Fig. 6). In the sample from Tokaanu, the most abundant is 3-MHeD, and for all other MMAs, the 2- and 3-MMAs are more predominant than the mid-chain isomers. In ET3-2 and ET2-1, the mid-chain methylpentadecanes (MPD) are also more predominant than 2-MPD and 3-MPD. In Tepuia, the mid-chain MMAs are more predominant than the 2- and 3-MMAs for the methylhexadecanes, MHeDs, and methyloctadecanes.
There is a strong increase in the relative abundance for the MMAs between C24–C30 for ET4-4, C24–C32 for Tokaanu, C24–C31 for Mangatete, and C25–C30 for Te Kopia. Two unidentified compounds that are tentatively assigned as dimethylheptadecanes (DMHeDs) have a base peak at m/z 57 and an m/z 268 molecular ion (labeled DMHeD? in Fig. 5). A third DMHeD was identified as 5,12-DMHeD, based on retention time and spectral comparison with the spectrum published in the work of Shiea et al. (1990).
4.2.3. Aromatic hydrocarbons
No aromatic compounds were detected in samples ET4-4 or Mangatete, but naphthalene is present in all other samples. The methylnaphthalene (MN) isomers 2-MN and 1-MN are present in both ET2-1 and ET3-2, whereas Tokaanu contains a suite of MN, ethylnaphthalenes, dimethylnaphthalenes, and trimethylnaphthalenes (Fig. 7; Table 2). Fluorene and the 2-, 3-, 1-, and 4-methylfluorene (MF) isomers were detected in both Tokaanu and El Tatio sample ET3-2, with the 2-MF/1-MF ratio of Tokaanu = 1.49, and ET3-2 = 0.49. These samples were the only ones that contained phenanthrene and the four methylphenanthrene (MP) isomers (Fig. 8). Anthracene, 1-MA, and biphenyl were also detected in the Tokaanu sample. The methylnaphthalene ratio for Tokannu (1.21) is much higher than that of ET3-2 (0.76; Table 2). Tokaanu has an ethylnaphthalene ratio of 1.32, a dimethylnaphthalene ratio-1 of 2.8, a trimethylnaphthalene ratio-1 (TNR-1) of 0.67, and a trimethylnaphthalene ratio-2 (TNR-2) of 0.96 (Table 2).
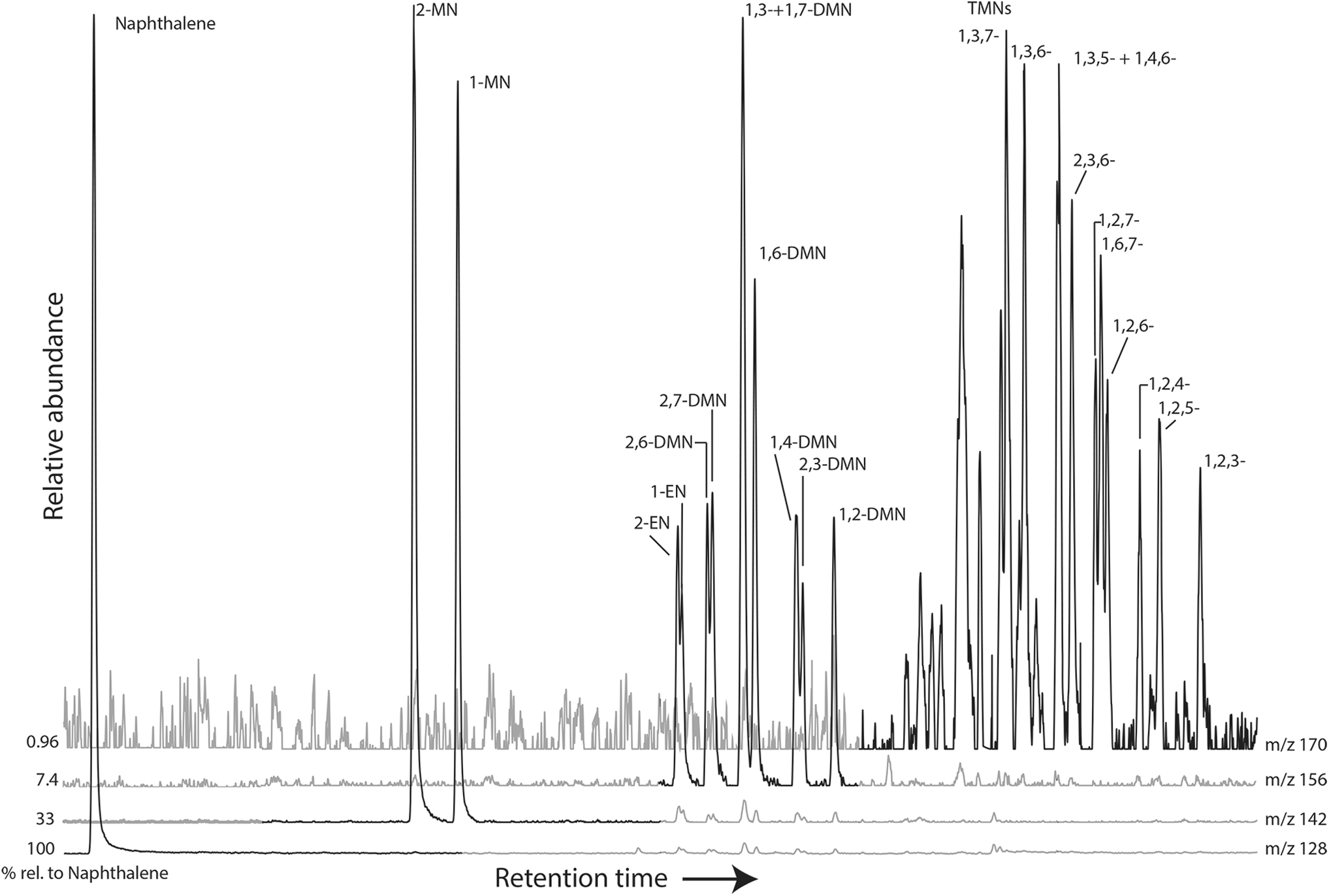
Partial m/z 128, 142, 156, and 170 mass chromatograms of the Tokaanu sample. Abundance of each chromatogram relative to naphthalene is shown. DMN = dimethylnaphthalene; EN = ethylnaphthalene; MN = methylnaphthalene; TMN = trimethylnaphthalene.

Partial m/z 178 to 192 mass chromatograms of the Tokaanu sample, showing phenanthrene, anthracene, and their methyl derivatives. MA = methylanthracene; MP = methylphenanthrene.
Aromatic Hydrocarbon Ratios for Samples from El Tatio (Chile) and the Taupo Volcanic Zone, New Zealand
MNR = methylnaphthalene ratio (2-MN/1-MN); ENR = ethylnaphthalene ratio (2-EN/1-EN); DNR-1 = dimethylnaphthalene ratio ([2,6-DMN + 2,7-DMN])/1,5-DMN); TNR-1 = trimethylnaphthalene ratio-1 (2,3,6-TMN/[1,4,6- + 1,3,5-TMN]); TNR-2 = trimethylnaphthalene ratio-2 ([2,3,6- + 1,3,7-TMN]/[1,4,6- + 1,3,5- + 1,3,6-TMN]); MPI = methylphenanthrene index (1.5 × (3-MP + 2-MP)/(P + 9-MP + 1MP)). Rc = calculated reflectance; MPR = methylphenanthrene ratio (2-MP/1-MP). nd = not detected.
The amount of phenanthrene relative to MPs increases with increasing degree of thermal alteration, and there is also preferential preservation of the β-substituted isomers 3-MP and 2-MP (Radke et al., 1982). Phenanthrene is >5 times more abundant than the MPs (Fig. 8), >8 times for Tokaanu, and almost 6 times for ET3-2. The methylphenanthrene ratio (MPR) for the ET3-2 and Tokaanu samples is 1.42 and 1.32, respectively (Table 2). As a result, the methylphenanthrene index (MPI) was used to derive calculated reflectance (Rc) by using the lower maturity calibration (Rc = (0.6 × MPI) +0.4) (Radke et al., 1982). Rc values are 0.82 for ET3-2 and 0.73 for Tokaanu, showing that both samples have a thermal maturity in the middle of the oil window (Table 2: see Section 5).
5. Discussion
5.1. Depositional environments
All samples have Pr/Ph ratios of <1.1. Values <1.0 are indicative of hypersaline and evaporative conditions (Clark and Philp, 1989), which are common in hydrothermal fluids of alkali chloride composition. Low Pr/Ph values in the El Tatio samples are likely related to the presence of saline waters associated with evaporites (Fernandez-Turiel et al., 2005). The Pr/Ph ratio is higher in ET2-1, which is from a flowing thermal stream, but lower in ET4-4, where the sinter breccia was exposed to moisture but no fluid outflows.
Tokaanu hosts chloride-rich springs (Soto et al., 2019), as does Te Puia (formerly Whakarewarewa, Lloyd, 1975). The Pr/Ph ratios of 0.98 and 0.90 for these two samples, respectively, are consistent with these environments.
Mangatete, however, has a much lower Pr/Ph ratio than the other samples (0.38). Low Pr/Ph ratios have been associated with high sulfur content (Peters et al., 1995), and low values of Pr/Ph (0.4) have also been associated with green and purple sulfur bacteria (Bhattacharya et al., 2017).
5.2. Organic matter inputs
5.2.1. Bacterial input
The high values of the 17/(16 + 18) n-alkane parameter indicate the presence of cyanobacteria in all samples. However, the Mangatete sample differs slightly from other samples studied here in its n-alkane distribution, which appears more similar to the distribution in the green sulfur bacteria Chlorobidium sp. (Shiea et al., 1991). The possibility of a green sulfur bacteria source is supported by the fact that it was necessary to remove elemental sulfur from the Mangatete sample, and because of the low Pr/Ph ratio from this sample, as discussed next.
In the samples studied here, except Te Kopia, we report the presence of mid-chain MMAs, which are the direct products of cyanobacteria (Schirmer et al., 2010). Cyanobacteria are the only microorganisms known to produce mid-chain MMAs, and they are found in most species of cyanobacteria (Shiea et al., 1990; Schirmer et al., 2010; Hoshino and George, 2015).
The most commonly reported organic components derived from cyanobacteria are the 7- and 8-methylalkanes, which have been previously reported as significant components of hot spring cyanobacterial mats (Robinson and Eglinton, 1990; Shiea et al., 1990). Indeed, Robinson and Eglinton (1990) noted that hydrocarbons present in mats in varied environments globally have a varied carbon number range and distribution of mid-chain branched alkanes, but that the composition of the microbial population and environmental effects (e.g., water temperature) may govern the distribution of mid-chain branched alkanes in specific mats.
In modern cyanobacterial monomethylalkane synthesis, 6-MHeD is not a major product (Coates et al., 2014), although some cyanobacterial species (such as Fischerella—a streamer-fabric-producing cyanobacterium found in hot spring environments) produce more 6- and 5-methylalkanes than the 7+-methyl isomers (Coates et al., 2014; He et al., 2015). This trend is similar to the results reported here from two of our samples (Mangatete and ET3-2). In addition, Hoshino et al. (2015) reported 6-MHeD having a higher abundance than 7-MHeD in endolithic bacteria found on the surface of 2.7 Ga stromatolites from limestones in the Pilbara Craton, Western Australia, which they suggested could be related to relative thermal stabilities of the hydrocarbons. Alternatively, wild cyanobacteria may have produced different branched alkanes than their laboratory-cultured counterparts, or perhaps Hoshino et al. (2015) discovered cyanobacteria that had not yet been identified. In the samples from our study, the outer slices were removed, so endolithic cyanobacteria living on the outside of the samples cannot be a source here.
5.2.2. Algal and terrigenous input
The El Tatio samples all have pronounced odd-over-even carbon number predominance (CPI >1) in the long chain n-alkanes. This signature, especially for C27, C29 and C31, is commonly associated with the waxes of vascular plants (Eglinton and Hamilton, 1967). In contrast, the TVZ samples have a CPI near 1.0, which may reflect an algal signal, or a source from sulfate-reducing bacteria. Some of the latter are known to produce long-chain n-alkanes ranging from n-C25 to n-C35 (Ladygina et al., 2006; Melendez et al., 2013). This difference in microbial community composition could explain the differing signals obtained from the TVZ versus the El Tatio samples, where no sulfur is present in the studied materials.
In addition, the high-molecular-weight methylalkanes do not have the same mid-chain predominance in the TVZ samples (Fig. 2), but they rather have 2- and 3-methyl isomers as the predominant input. This signal has been observed in lichen (consisting of an alga and a fungus; Huang et al., 2012), although the source of the compounds remains poorly defined (Huang et al., 2011). Samples with a weak even-over-odd carbon number predominance sometimes also produce these high-molecular-weight methylalkanes, and this signal has been associated with algal organic matter, if the thermal maturity is not high (Cheng et al., 2018), which is not the case for any of the samples studied here (Section 5.3).
5.3. Thermal maturity
The mostly low Ph/n-C18 ratios of 0.14–0.89 are consistent with a thermal maturity in the oil window, as are the low Pr/n-C17 ratios (0.03–0.17) (Table 1; Ten Haven et al., 1987; Aharonovich and George, 2016). The MPI value, which changes with thermal maturity (Radke et al., 1982; Radke, 1988), is 0.55 for Tokaanu, and 0.70 for ET3-2 (Table 2), placing these samples in the early oil window (Rc of 0.73% and 0.82%). The MPR is in agreement that ET3-2 (1.42) has a higher thermal maturity than Tokaanu (1.32). The alkylnaphthalene data and the smooth n-alkane distribution (CPI22–32 near unity) for Tokaanu are also consistent thermal maturities within the oil window (Table 2).
These data, indicative of temperatures typical in petroleum systems, are unusual for recent sediment/rock samples that have not been buried. The Tokaanu sample comes from the mid-slope of the hot spring sinter apron, an area that is generally a mid-temperature environment (60–45°C; Campbell et al., 2015a), so it seems unlikely that the temperatures indicated by the thermal maturity values are related to the surface-temperature gradient. Rather, several boreholes in Tokaanu show that the surface hydrothermal fluids originate from higher temperatures in the subsurface throughout the field, some of which are higher than 130°C (Soto et al., 2019). The Tokaanu sinter comes from the dissolution of Si-bearing minerals at high temperatures beneath the hydrothermal field (Soto et al., 2019). Thus, these aromatic hydrocarbons with a thermally mature distribution may be reflective of organic matter sourced from subsurface hydrothermal fluids migrating from deeper immature source rocks that have been hydrothermally altered, rather than from the in situ material preserved within the samples. The presence of a significant amount of anthracene, and a small amount of 2-methylanthracene, is consistent with hydrothermal alteration of immature organic matter (e.g., Kawka and Simoneit, 1990; Leif and Simoneit, 1995).
This may also be the case for the ET3-2 sample, which has a slightly higher maturity than Tokaanu based on the alkylphenanthrene distributions. The waters at El Tatio have been interpreted to be heated by the Laguna Colorado ignimbrite caldera complex (Zandt et al., 2003). The primary aquifer has a temperature of 265°C at a depth of 800 m, whereas the surface waters discharged at El Tatio have a temperature of ∼86°C (which is the boiling point at the El Tatio hydrothermal field; Lahsen and Trujillo, 1975). However, this does not explain why these two samples are the only ones studied that have these aromatic compounds. Analysis of further samples is required to ascertain why this is the case.
5.4. Biomarker preservation in hot springs
The fossilized opal-A sample from Mangatete has comparable abundances and a similar compound range to the modern opal-A-bearing samples, even though it is 9.4 ka. However, the younger, ∼3.5 ka, fossilized microcrystalline quartz sample from Te Kopia has fewer, and lower abundance, compounds. It, thus, would appear that the degree of hydrocarbon preservation is, at least in part, related to sample crystallinity arising from the degree of diagenesis. The pristine opal-A sinters preserve more hydrocarbons than samples affected by a greater degree of diagenesis. The reason for this is that, typically, the density of silica minerals increases as diagenesis proceeds (Lynne and Campbell, 2004), resulting in occlusion of porosity and expelling of interstitial organic compounds. During deposition, opal-A encrusts or replaces microbes, and it then continues to precipitate as a cement framework. This process continues as long as the silica saturated waters are flowing, and it can reduce porosity by up to half (Jones and Renaut, 2003). As pore fluids continue to evolve during burial, hydrocarbons may migrate and be carried out of the host rock to imprint a hydrocarbon signature on the mineralogically maturing sinter deposit. In some of our samples, high-temperature signals, such as the alklylnaphthalenes, appear to be overprinting the primary biogenic signal. In these samples, we can detect the low maturity signal due to the cyanobacteria, and the signal from the high maturity migrating fluid.
5.5. Life on Mars
The results from this study have implications for the search for life on Mars. Currently, the search for fossil organics on Mars is focused on lacustrine-deltaic sediments and the regolith. However, this study shows that other strong candidates in the search for biomolecules would be the opal-A deposits at Columbia Hills identified by the Miniature Thermal Emission Spectrometer (Mini-TES; Squyres et al., 2008), hydrothermal silica at Nili Patera identified by the Compact Reconnaissance Imaging Spectrometer for Mars (CRISM; Skok et al., 2010), or perhaps among the high-Si minerals identified by the Chemistry and Camera (ChemCam) instrument, and the Chemistry and Mineralogy (CheMin) instrument in Gale crater (Morris et al., 2016; Frydenvang et al., 2017).
The results presented here are biased by the overwhelming prevalence of photosynthesizers on Earth today and in the recent past (e.g., Hays et al., 2017; Michalski et al., 2018). Although the types of organics we may detect on Mars could be radically different than the cyanobacterial and plant signatures discovered here, hot springs remain some of the most ancient analogs for life on Earth (e.g., Djokic et al., 2017), and thus they provide a window to when Earth and Mars would have been the most similar. Given that silica is known to be an exceptional preserver of microfossils and other biosignatures (e.g., Knoll, 1985; Trewin, 1996), and can act as a potential shield to UV radiation (Phoenix et al., 2002), it becomes a very attractive beacon by which to guide the search for life on Mars, whether or not photosynthesizers existed there. At Columbia Hills, the presence of nodular opaline silica with digitate structures, similar to those from El Tatio sinter known to have been microbially mediated (Ruff and Farmer, 2016; Ruff et al., 2019), provide exciting opportunities to investigate whether life may have ever existed on Mars (Westall et al., 2015a; Cady et al., 2018).
6. Conclusions
Three modern opal-A sinter samples from El Tatio, two modern opal-A sinter samples from the TVZ in New Zealand, a ∼3.4 ka microcrystalline quartz sinter sample from Te Kopia in the TVZ, and a ∼9 ka opal-A sinter sample from Mangatete in the TVZ were investigated for their potential biomolecular preservation state. All samples contain biomarker distributions that are consistent with the presence of cyanobacteria. The samples collectively have Pr/Ph ratios that are reflective of evaporative conditions, consistent with the sinter apron environments from which they were collected. Thermal maturity parameters indicate the presence of organic compounds in the oil window, derived from temperatures higher than those present in the surface environment, suggesting a contribution of organic components from fluids that have undergone convective circulation in the subsurface and, thus, are not solely from microbial remnants indigenous to the rock sample. Overall, hydrocarbon preservation in these samples appears to be affected more by crystallinity—reflecting degree of diagenesis—than by age. These findings are favorable for the search for traces of ancient life in opaline silica deposits on Mars.
Footnotes
Acknowledgments
B.L.T. acknowledges helpful discussions with Bryan Drake and Ayrton Hamilton about sinter aprons. B.L.T. would like to acknowledge and pay her respects to the Bedegal people (UNSW) and the Darug Nation (Macquarie University), the traditional custodians of the land on which this research took place. The authors thank the communities of Caspana and Toconce, Chile for allowing fieldwork and sample collection at El Tatio. Thanks are due to Diego Guido for providing an updated sinter facies-temperature gradient diagram.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
B.L.T. was supported by an Australian Government Research Training Program Scholarship. Funding for this project comes from an Australian Research Council Discovery Grant to M.J.V.K., and samples were collected during research conducted by K.A.C. supported by a Royal Society of New Zealand Marsden grant.
Supplementary Material
Supplementary Table S1
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.