
Abstract
Earth's ocean-atmosphere system has undergone a dramatic but protracted increase in oxygen (O2) abundance. This environmental transition ultimately paved the way for the rise of multicellular life and provides a blueprint for how a biosphere can transform a planetary surface. However, estimates of atmospheric oxygen levels for large intervals of Earth's history still vary by orders of magnitude—foremost for Earth's middle history. Historically, estimates of mid-Proterozoic (1.9–0.8 Ga) atmospheric oxygen levels are inferred based on the kinetics of reactions occurring in soils or in the oceans, rather than being directly tracked by atmospheric signatures. Rare oxygen isotope systematics—based on quantifying the rare oxygen isotope 17O in addition to the conventionally determined 16O and 18O—provide a means to track atmospheric isotopic signatures and thus potentially provide more direct estimates of atmospheric oxygen levels through time. Oxygen isotope signatures that deviate strongly from the expected mass-dependent relationship between 16O, 17O, and 18O develop during ozone formation, and these “mass-independent” signals can be transferred to the rock record during oxidation reactions in surface environments that involve atmospheric O2. The magnitude of these signals is dependent upon pO2, pCO2, and the overall extent of biospheric productivity. Here, we use a stochastic approach to invert the mid-Proterozoic Δ17O record for a new estimate of atmospheric pO2, leveraging explicit coupling of pO2 and biospheric productivity in a biogeochemical Earth system model to refine the range of atmospheric pO2 values that is consistent with a given observed Δ17O. Using this approach, we find new evidence that atmospheric oxygen levels were less than ∼1% of the present atmospheric level (PAL) for at least some intervals of the Proterozoic Eon.
1. Introduction
The mid-Proterozoic (1.9 to 0.8 billion years ago, Ga) is generally regarded as a transitional time between the anoxic surface conditions of the Archean and the relatively well-oxygenated conditions that characterized much of the late Neoproterozoic and Phanerozoic (Canfield, 1998, 2005; Holland, 2006; Kump, 2008; Derry, 2015; Laakso and Schrag, 2017). However, there exists a wide range of estimates for surface oxygen during the mid-Proterozoic, from <0.1% to 40% of the present atmospheric level (PAL) (Kump, 2008; Lyons et al., 2014). If estimates of mid-Proterozoic pO2 levels near or below ∼1% (PAL) are correct, environmental conditions characteristic of the majority of Earth's history may have limited ecosystem complexity and could have impacted the emergence and ecological expansion of mobile multicellular heterotrophs (Reinhard et al., 2016). The potential for low pO2 levels during the mid-Proterozoic also has important implications for oxygen-based frameworks for remote life detection on terrestrial planets. In particular, if we use Earth as an analogue and take lower Proterozoic pO2 estimates, it is possible that terrestrial planets can stabilize for billion-year timescales at weakly oxygenated states, with implications for atmospheric biosignature detection (Reinhard et al., 2017). The importance of the mid-Proterozoic for the secular history of Earth's biotic and environmental evolution thus provides strong motivation to critically assess claims of low surface oxygen levels through this interval.
Historically, mid-Proterozoic atmospheric oxygen levels were assumed to be ∼10% PAL based on geochemical signatures in paleosols and marine redox records (Holland, 2006; Kump, 2008). There are several reports of Proterozoic paleosols with quantitative iron oxidation, implying atmospheric oxygen levels above ∼1% PAL (Pinto and Holland, 1988; Zbinden et al., 1988). The presence of anoxic deep oceans (e.g., Poulton et al., 2010; Lyons et al., 2014) has been assumed to roughly constrain maximum atmospheric oxygen levels at ∼40% (Canfield, 1998; Kump, 2008). However, these traditional bounding pO2 estimates have recently been questioned (Planavsky et al., 2018). Critically, the only definitive mid-Proterozoic paleosols that have formed directly from parent rock are characterized by iron loss (Mitchell and Sheldon, 2009, 2010, 2016), pointing to low oxygen levels. A suite of other proxies has also suggested atmospheric oxygen levels less than ∼10% PAL or even ∼1% PAL, for at least intervals of the mid-Proterozoic. For instance, sedimentary Cr isotopes and carbonate trace element patterns have been suggested to indicate atmospheric oxygen levels less than 1% PAL, perhaps as low as 0.1% PAL (Cole et al., 2016; Gilleaudeau et al., 2016; Liu et al., 2016; Bellefroid et al., 2018), consistent with updated currently available mid-Proterozoic paleosol records. However, records of both low and high mid-Proterozoic oxygen levels (Zhang et al., 2016b) have been questioned and critiqued (Planavsky et al., 2016).
Some of the debate surrounding Proterozoic oxygen levels emerges naturally from the diverse suite of proxies being applied to the problem. The majority of paleo-oxygen barometers do not directly track an atmospheric signal; instead, they track an oxidation reaction occurring in a soil (or the oceans), or they attempt to link a marine redox structure to an atmospheric oxygen level. These approaches, even those that rely on fossilized soil horizons in more direct contact with the overlying atmosphere, can only indirectly track pO2 and rely on a large number of assumptions. For instance, to link a paleosol record to an atmospheric oxygen level one must assume a background pCO2, local rainfall and infiltration rates, and a denudation rate, in addition to assumptions regarding the presence/absence and/or activity of a local terrestrial biosphere. To link a marine redox-sensitive metal signature to an atmospheric pO2 estimate one needs to link local or global ocean redox to atmospheric composition and is thus reliant on a series of assumptions regarding ocean circulation and nutrient inventories. Not surprisingly, different researchers have reached very different conclusions about Proterozoic oxygen levels using essentially the same numerical frameworks and empirical records. The strong potential for diagenetic overprinting of depositional signatures in paleoredox proxies only adds to uncertainty and continuing debate.
The sedimentary triple-oxygen isotope record presents a means to more directly track mid-Proterozoic atmospheric oxygen levels. The majority of reactions on Earth induce stable isotope fractionations that are dictated almost entirely by mass differences between isotopes (Urey, 1947). However, for systems with more than two isotopes, certain processes can create pools that are anomalously enriched or depleted in a given isotope relative to what would be expected for a purely mass-dependent process, leading to so called “mass-independent” or “non-mass-dependent” (NMD) isotope effects (e.g., Thiemens, 2006). Ozone formation is one of the most intensively studied of the these NMD isotope effects (Thiemens and Heidenreich, 1983). Ozone formed in the stratosphere is anomalously enriched in 17O, denoted as a positive Δ17O value. This positive Δ17O signal is transferred via photochemistry to atmospheric CO2, while the complementary negative Δ17O signal is stored in residual stratospheric O2. The magnitude of the negative Δ17O anomaly in atmospheric oxygen is dependent most strongly on the amount of atmospheric O2 and CO2 and how quickly the anomalous O2 produced during photochemistry is reset by biological cycling (Luz et al., 1999; Cao and Bao, 2013). This residual negative Δ17O signal can then be transferred to the rock record via oxidation reactions during weathering (e.g., pyrite oxidation). Therefore, the Δ17O composition of sedimentary sulfate minerals at any given time and location primarily reflects (1) atmospheric O2 levels; (2) atmospheric CO2 levels; and (3) biospheric O2 recycling (e.g., primary productivity) (Luz et al., 1999; Cao and Bao, 2013). Importantly, once atmospheric oxygen isotope signatures are imparted to sulfate diagenetic processes, microbial sulfur cycling and/or metamorphic overprinting should only act to erase Δ17O anomalies, making this proxy in many cases more robust than traditional redox proxies.
Here, we revisit previous modeling of the Proterozoic sulfate mineral Δ17O record (Crockford et al., 2018a, 2019; Hodgskiss et al., 2019) in order to more precisely reconstruct atmospheric pO2 during an interval of mid-Proterozoic time. We utilize a stochastic approach, building from statistical analysis of output from the global biogeochemical model CANOPS (Ozaki et al., 2011, 2019), to filter the large number of mathematically possible solutions for any given Δ17O value according to only those solutions that allow for mass balance between biospheric productivity and atmospheric pO2. This results in a dramatically reduced number of possible O2-CO2-productivity combinations consistent with a given Δ17O measurement, and a much more precise range of atmospheric pO2 values.
2. The Mid-Proterozoic Δ17O Record
Sulfate minerals extracted thus far from mid-Proterozoic carbonates have been found to be heavily contaminated with recent atmospheric sulfate and thus cannot be used to track atmospheric signals (Peng et al., 2014). However, sulfate from a limited number of mid-Proterozoic evaporite successions appears to preserve Δ17O signals from atmospheric O2, presumably acquired during terrestrial pyrite oxidation (Crockford et al., 2018a). Although Δ17O signals in examined evaporite successions are highly variable, there are distinctively negative Δ17O signals relative to the Phanerozoic sulfate record (Crockford et al., 2018a) (Fig. 1). These signals are thought to capture negative Δ17O anomalies from tropospheric O2. Here we assume that between 5% and 15% of the oxygen atoms that become incorporated into sulfate during pyrite oxidation are atmospherically derived (Balci et al., 2007; see discussion in Crockford et al., 2018a; Kohl and Bao, 2011). This range is slightly lower than a recent estimate of ∼18% atmospheric oxygen in sulfate from the strongly anthropogenically influenced Mississippi River system (Killingsworth et al., 2018).
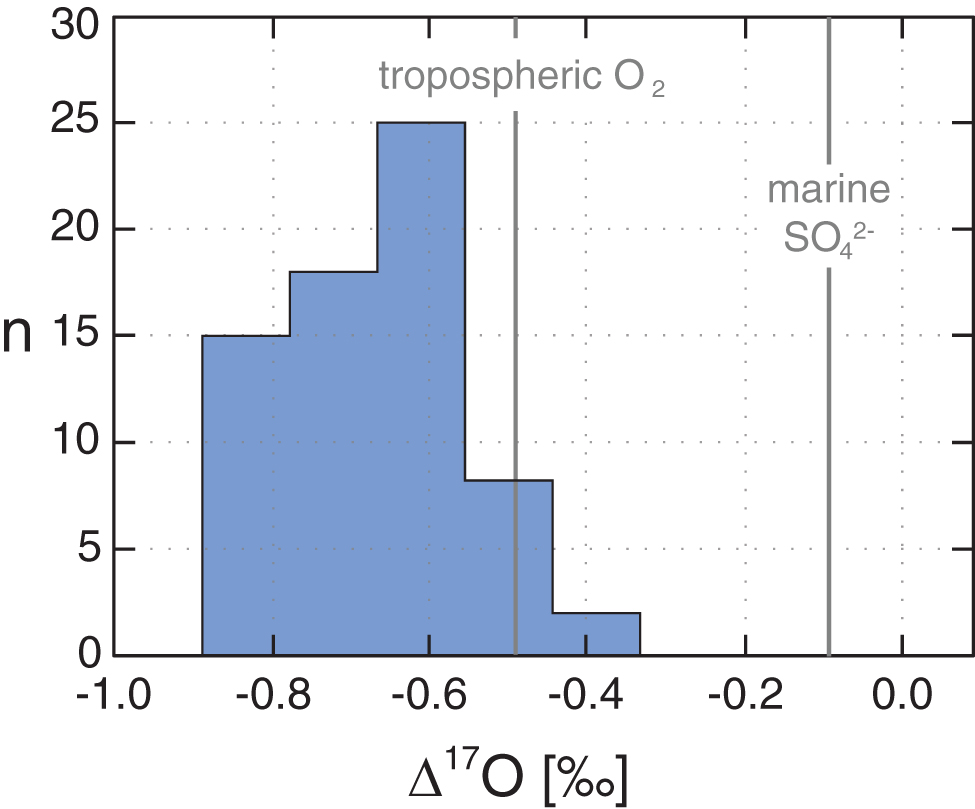
Histogram of gypsum Δ17O values from the ca. 1.4 Ga lacustrine Sibley Basin in Ontario, Canada. This lacustrine setting seems to be characterized by limited sulfur cycling (sulfur reduction and re-oxidation; see Crockford et al., 2018a), allowing for the effective preservation of atmospheric Δ17O anomalies. For comparison, the gray lines show the compiled values of modern marine sulfate and modern tropospheric O2. Note that only a portion (roughly 10%) of atmospheric oxygen is incorporated into sulfate during pyrite oxidation. Data adapted from Crockford et al. (2018a) and Hodgskiss et al. (2019). Color images are available online.
Variability on the formation scale in existing mid-Proterozoic evaporite Δ17O data sets is expected, given that any surface sulfur cycling will tend to homogenize and erase Δ17O signals. In particular, activation of S to SO3 2- during microbial sulfate reduction leads to extremely rapid O isotope equilibration with isotopically “normal” ambient H2O within the cell (e.g., Antler et al., 2013). Therefore, sulfate reduction and sulfide oxidation will tend to decrease the magnitude of Δ17O signals. Marine environments, even those that are strongly redox stratified (e.g., the Black Sea), are characterized by extensive sulfide re-oxidation (Yakushev and Neretin, 1997). Evaporative successions can similarly be marked by high rates of sulfur redox cycling (e.g., Petrash et al., 2012). With these observations in mind, it is not surprising that there is variation within a given evaporite unit and that the most anomalous Δ17O signals have thus far been found in lacustrine—rather than marine—evaporite successions (Crockford et al., 2018a). The most negative Δ17O signals found to date with the exception of Cryogenian barite and CAS (cf. Bao et al., 2008; Crockford et al., 2016)—Δ17O values of -0.9‰ (Fig. 1)—are observed in the lacustrine Sibley Formation in Canada, deposited at ∼1.4 Ga (Crockford et al., 2018a). However, similarly anomalously negative values (Δ17O < -0.75‰) are also found in the ca. 1.7 Ga marine Myrtle Shale in the MacArthur Basin in Australia (Crockford et al., 2019) and the ca. 1.9 Ga marine Belcher Group in Canada (Hodgskiss et al., 2019). Therefore, although the mid-Proterozoic sedimentary Δ17O record is still sparse, motivating additional work, there is a clear signal from several formations for markedly different sulfate Δ17O values than those observed from the modern and recent Earth system. Because minimum values are likely our best estimate of a true atmospheric signal (Crockford et al., 2018a; Planavsky et al., 2018), in what follows we develop an approach that is designed to translate these anomalously low values into a quantitative estimate of atmospheric pO2. However, we stress that current records are unlikely to capture “true” minimum sulfate Δ17O values, indicating that our approach should be viewed as providing maximum pO2 estimates.
3. Translating Δ17O Signals to pO2 Estimates
The first step in our approach uses a 4-box atmosphere-ocean-biosphere model derived from that of Cao and Bao (2013) (Fig. 2) to estimate all combinations of atmospheric pO2 and biospheric O2 recycling that are consistent with a given Δ17O value. The model is designed to predict tropospheric Δ17O solutions with different pO2, pCO2, and O2 residence times (e.g., biospheric O2 recycling rates). Δ17O values of stratospheric O2 and CO2 vary with changing pO2/pCO2 ratios, following experimental results from the O2-O3-CO2 photochemical reaction system (Shaheen et al., 2007). The pO2/pCO2 ratio affects both the lifespan of Δ17O anomalies in the troposphere and the magnitude of the stratospheric Δ17O anomaly (Cao and Bao, 2013). When O2 with a Δ17O anomaly is mixed down into the troposphere, it can be consumed by aerobic respiration and replaced by isotopically “normal” O2 produced from photosynthesis (e.g., Luz et al., 1999). It is important to point out that our approach here, as well as that of Cao and Bao (2013), implicitly assumes a constant vertical structure of the atmosphere, and in particular rates of oxygen exchange between the troposphere and stratosphere. This is unlikely to be correct below a certain atmospheric pO2, as production of O3 scales nonlinearly with ground-level atmospheric O2 abundance (Kasting and Donahue, 1980), and this should in turn impact the exchange rates of oxygen between the troposphere and stratosphere. In addition, these models do not fully account for O atom exchange between all atmospheric species (e.g., water vapor). A full evaluation of these issues will require a photochemical model that explicitly tracks O atoms and allows for variable vertical atmospheric structure. Nevertheless, this approach is in keeping with current and previous work, and we consider it a useful starting point from which to move forward.
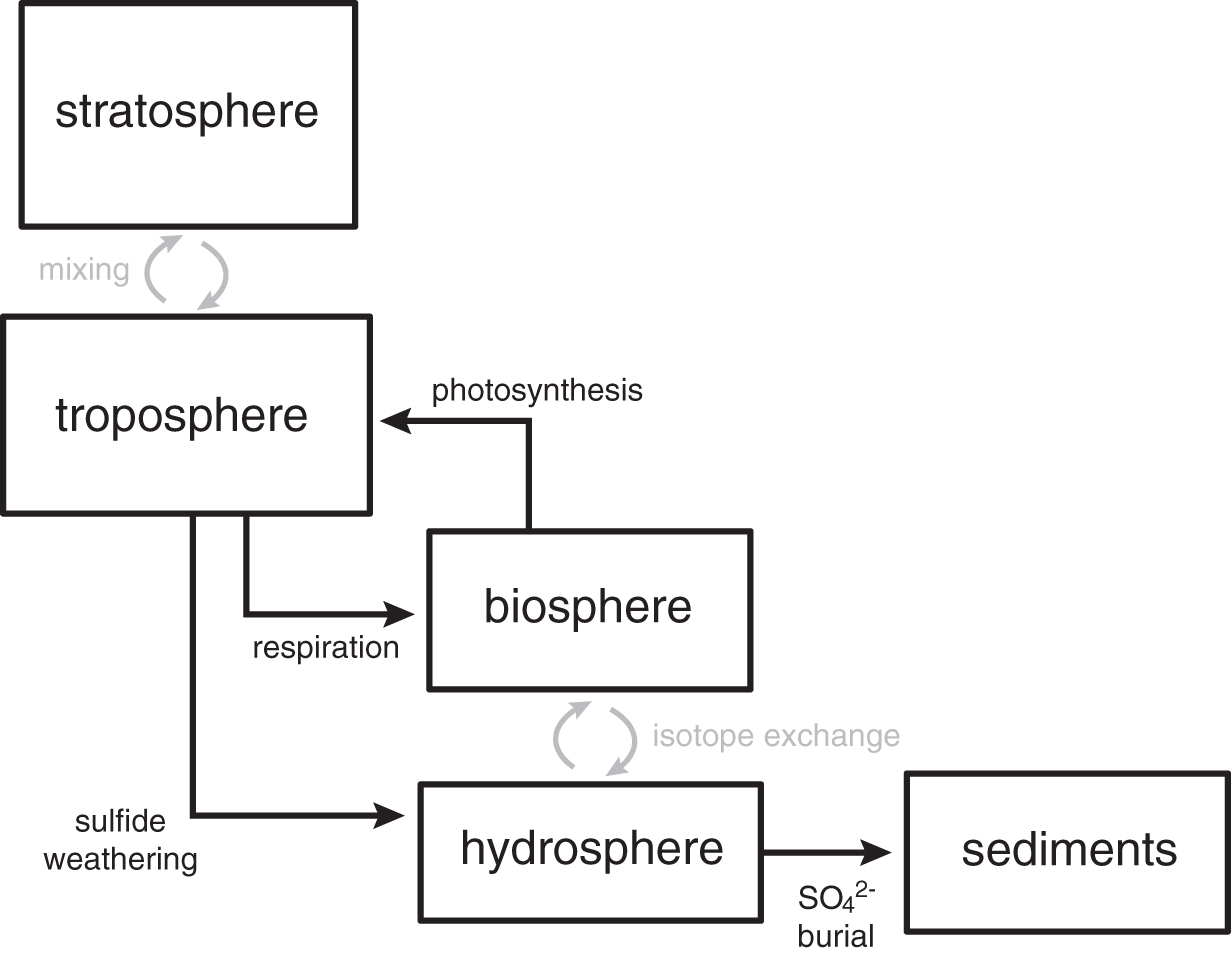
Schematic overview of the box model used in the first step of our technique for inverting Δ17O anomalies for atmospheric pO2. 17O excesses in ozone (O3) are created in the stratosphere through the Chapman cycle and subsequently transferred to CO2. Corresponding negative Δ17O anomalies are transferred to O2, which can be mixed down into the troposphere where it can be erased through photosynthesis and aerobic respiration (reset to the terrestrial mass fractionation line) or transferred to the rock record via weathering of reduced mineral phases (e.g., pyrite oxidation to form sulfate). The magnitude of Δ17O transferred to sulfate will be dependent on the atmospheric CO2/O2 ratio and the extent of global biospheric O2 recycling.
Because we assume steady state, the model effectively links the overall rate of biospheric O2 recycling to the oxygen released through primary production within the biosphere. It is important to note that in general biospheric O2 recycling will likely be less than gross primary productivity (GPP) (with the potential for some deviations induced by photorespiration and the Mehler reaction; see Hodgskiss et al., 2019, for a discussion). As the fraction of photosynthetic O2 that is exchanged with the environment before being used to recycle gross photosynthate by autotrophic respiration increases, biospheric O2 recycling approaches GPP. As this fraction drops—for example, most or all of the O2 used in autotrophic respiration is recycled before leaving the cell, or is produced in an adjacent region of the surface ocean before exchanging with the atmosphere—biospheric O2 recycling approaches net primary productivity. Because the range of pO2 values consistent with a given Δ17O value must increase as the rate of biospheric O2 recycling rises, we consider it conservative for our purposes to use GPP as our metric for biospheric O2 recycling. We note, however, that the factors regulating this relationship in differing environments and at significantly different O2 and CO2 levels are important topics for future work.
Although the amount of primary productivity in the terrestrial realm during the Precambrian is debated, we include a recent lower-end estimate of the oxygen efflux from terrestrial cyanobacterial mats (Planavsky et al., 2018; Zhao et al., 2018). However, it is worth noting that this terrestrial cyanobacterial productivity likely results in an insignificant amount of net organic carbon burial, given the high potential for oxidation during transport of this organic matter to the marine realm. Within this framework, the Δ17O value of O2 in the troposphere reflects the mass balance between sourcing of anomalous O2 from photochemistry in the stratosphere (the magnitude of which varies with the size of the atmospheric CO2 and O2 reservoirs) and O2 recycling within the biosphere (Fig. 2). The tropospheric Δ17O anomaly can then be transferred to the rock record during pyrite oxidation and subsequent sulfate burial, linking Earth's stratosphere with the rock record.
There are numerous non-unique O2, CO2, and GPP combinations that could be consistent with a given sedimentary Δ17O value (Cao and Bao, 2013; Crockford et al., 2018a). However, not all of these combinations of pO2, pCO2, and biospheric recycling will represent mechanistically accessible Earth system states—for example, states that are stable on geologic timescales and satisfy both mass balance within the coupled C-O-S-P cycles and global redox balance. To delineate which combinations of atmospheric O2, CO2, and global primary productivity represent plausible alternative Earth states, we rely on previous estimates of atmospheric CO2 (Sheldon, 2006; Kah and Riding, 2007; Crockford et al., 2018a; Isson and Planavsky, 2018) and combine these with results from a stochastic analysis of the biogeochemical Earth system model CANOPS (Ozaki et al., 2011, 2019) to filter out combinations of pO2, pCO2, and GPP that may satisfy Δ17O mass balance but do not provide redox-balanced Earth system states. The rationale behind this approach is simple—on geologic timescales there are mechanistic links within the Earth system between the globally integrated rate of biospheric primary productivity, organic carbon burial, and atmospheric oxygen levels (Laakso and Schrag, 2014, 2017; Derry, 2015; Ozaki et al., 2019), and these ensure the mass balance and redox balance on geologic timescales.
In practice, our approach occurs in two steps. First, pO2 and pCO2 combinations are randomly selected (assuming uniform distributions for both; pO2 0–100% PAL; pCO2 2–500 × PAL), and GPP is computed for each pCO2 and pO2 combination using our observed minimum Δ17O value (Δ17O = -0.9‰) as a constraint on our atmosphere-ocean-biosphere model (Fig. 2). This step of the inversion is performed following Crockford et al. (2018a) using the parameter ranges listed in Table 1 and results in a wide range of possible atmospheric oxygen levels. To refine these posterior atmospheric pO2 estimates further, we then filter the data set for combinations of atmospheric pO2 and GPP that yield mass- and redox-balanced solutions according to a recent stochastic analysis of the CANOPS global biogeochemical model (Figs. 3 and 4; Ozaki et al., 2019). Specifically, we filter the synthetic data set generated from the first resampling analysis for results that fall within the 90% credible interval of mass- and redox-balanced CANOPS results (Fig. 4). This additional filtering step dramatically refines the posterior distribution of atmospheric pO2 values consistent with the Δ17O constraint, reducing both the range of pO2 estimates and their central tendency (Fig. 5). The median pO2 value yielded by our analysis is 3.4 × 10−3 PAL, with a 90% credible interval of 1.4–8.7 × 10−3 PAL (Fig. 5). Although the posterior distribution of pO2 values still spans over an order of magnitude, this range is considerably more precise than that conventionally invoked for the mid-Proterozoic Earth, which spans well over 2 orders of magnitude (Lyons et al., 2014).
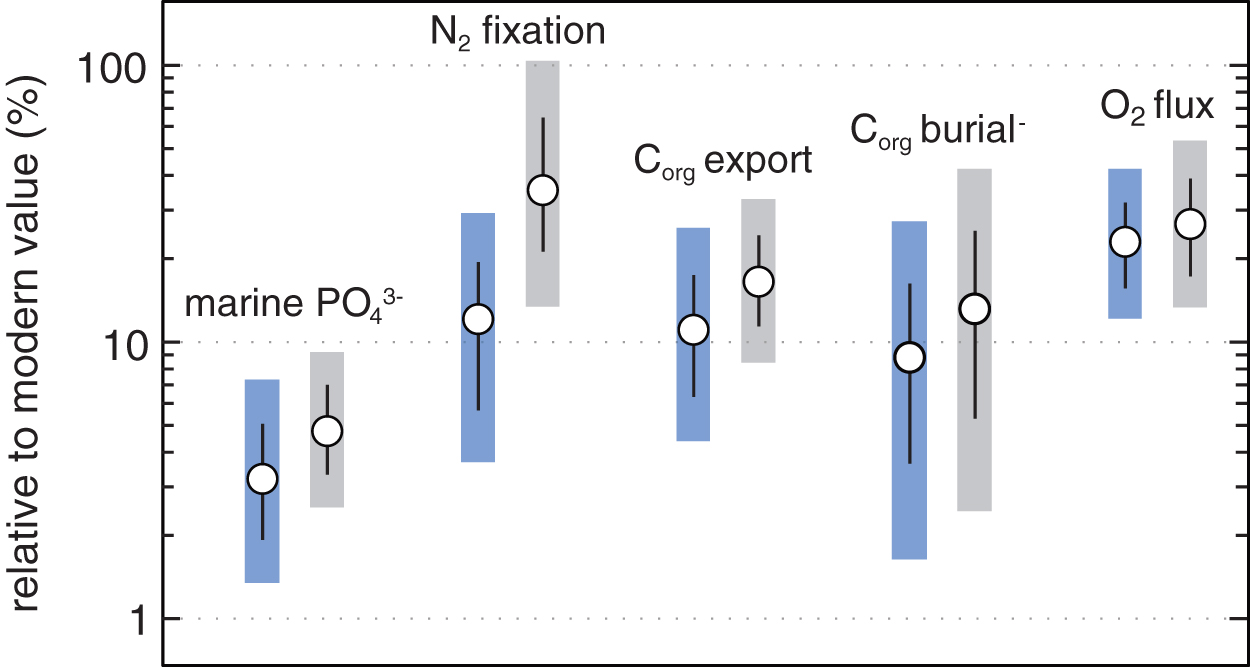
Example of the steady state environmental parameter space from the CANOPS Monte Carlo simulation with pO2 ranges of 0.01%–1% PAL (light blue) and 1%–10% PAL pO2 (light gray). Marine phosphate levels, the extent of N fixation, the marine export flux, organic carbon burial, and the O2 are all significantly less than those of modern Earth (see Ozaki et al., 2019). Color images are available online.
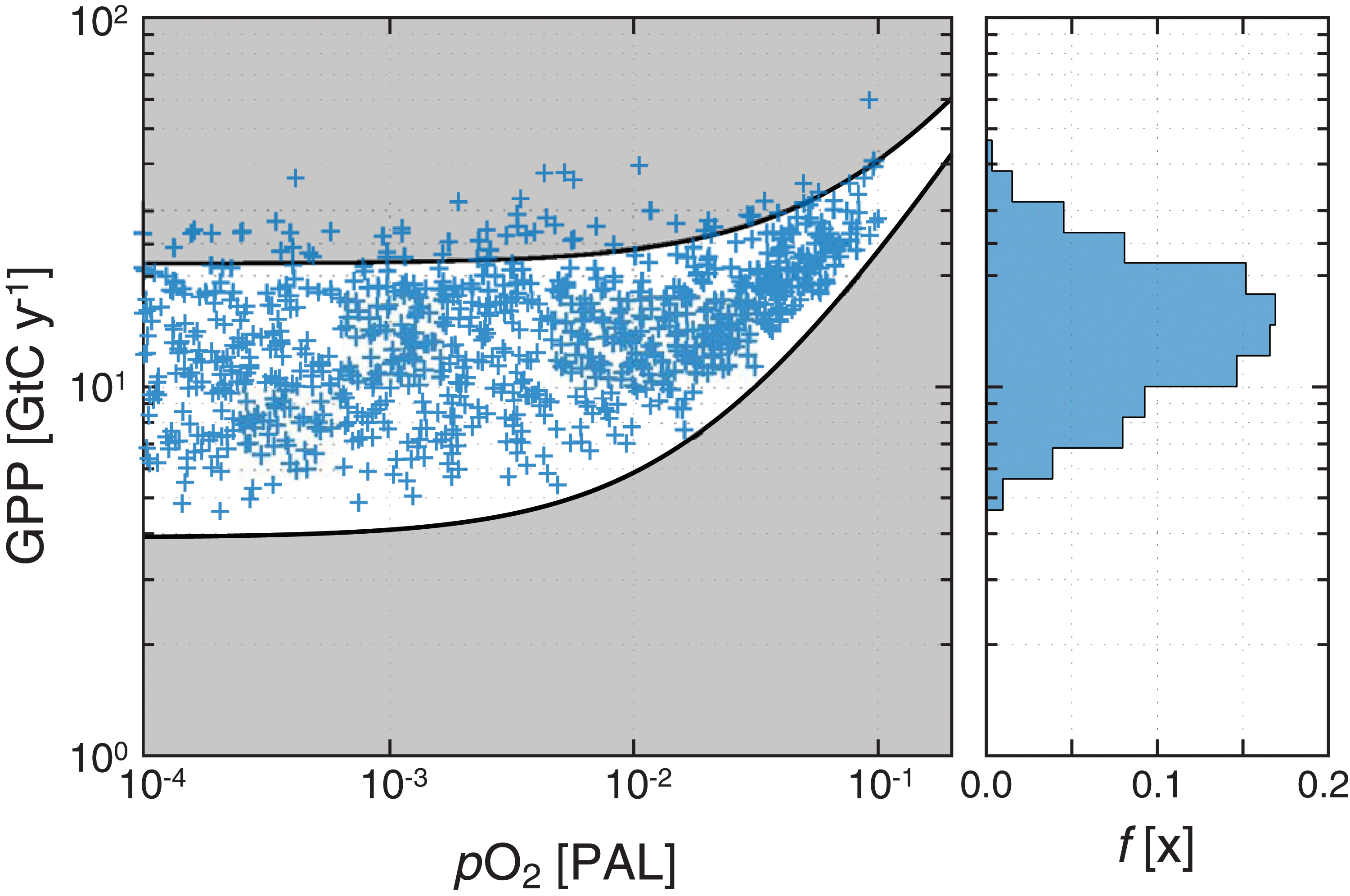
Mass- and redox-balanced Earth system states from the CANOPS global biogeochemical cycle model. Shown at left are individual CANOPS model runs from the Monte Carlo analysis of Ozaki et al. (2019), with the 90% credible interval for combinations of pO2 (relative to the present atmospheric level, PAL) and gross primary productivity (GPP, in gigatons of carbon per year, GtC y−1) yielding mass- and redox-balanced Earth system states shown by the black lines. The second step of the inversion analysis discussed in the text filters out possible pO2/pCO2/GPP combinations that fall within the gray shaded area. Shown at right is a relative frequency distribution of GPP. Color images are available online.
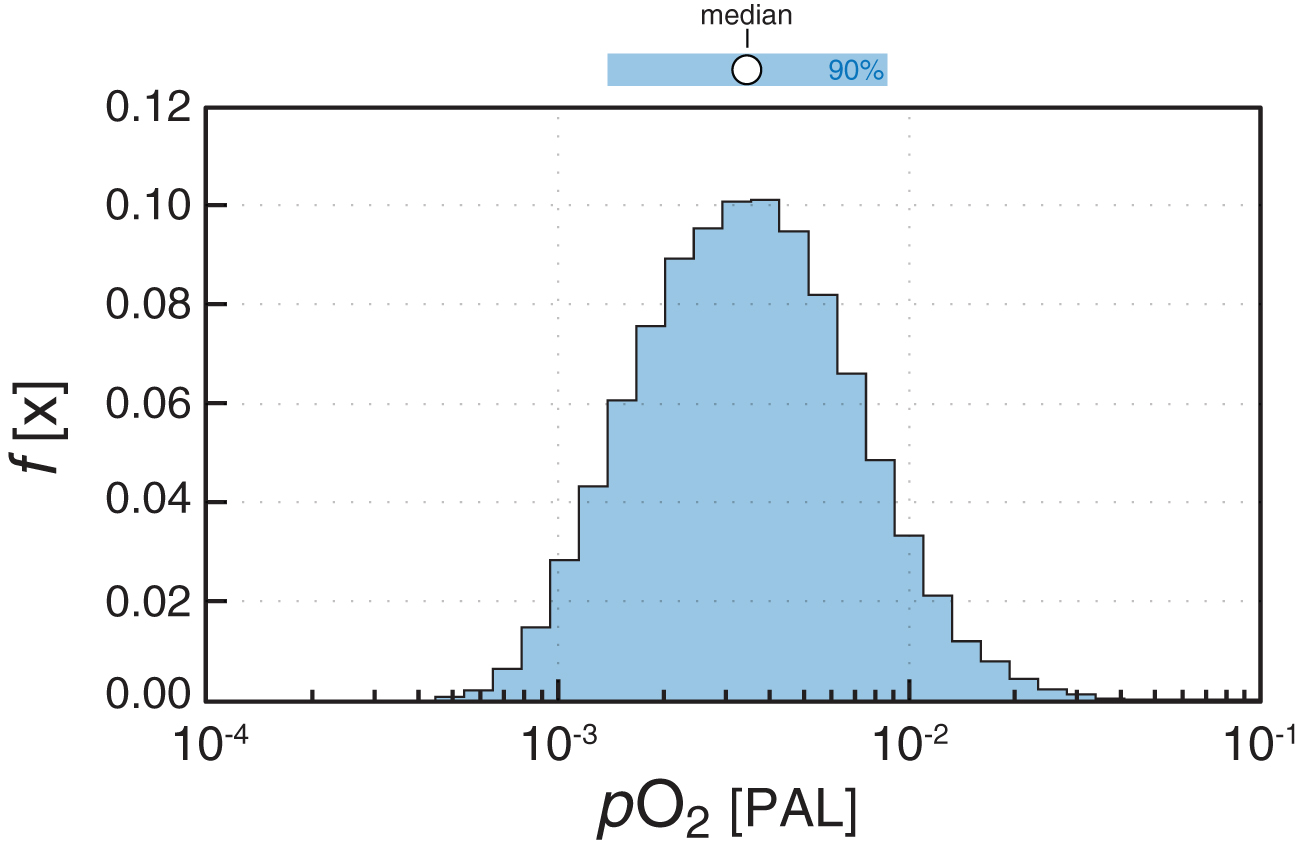
Atmospheric oxygen estimates consistent with the CANOPS-filtered sulfate Δ17O values from the inversion analysis. Although there are a wide range of numerically possible O2, CO2, primary productivity combinations that are consistent with a single sedimentary Δ17O value, most of these combinations are not mass- and redox-balanced Earth system states. The stable Earth system states that are consistent with the Sibley Δ17O data generally have low (<1% PAL) atmospheric oxygen levels. The median value and 90% credible interval for pO2 are shown above the figure. Color images are available online.
Parameters Used in the Monte Carlo Simulation
4. Discussion
Given the strong potential to erase Δ17O anomalies (through surface sulfur cycling) and the stratigraphic rarity of rocks that best preserve atmospheric Δ17O signals—lacustrine evaporites—we can only reconstruct meaningful maximum pO2 levels from snapshots of mid-Proterozoic time using this approach. However, we suggest that this snapshot may offer the most robust and precise constraint on surface oxygen levels during mid-Proterozoic time. First, later diagenetic alteration should erase Δ17O signals for low oxygen, which is in strong contrast to most oxygen proxies for which signals implying low oxygen can potentially be caused by burial diagenesis, alteration, or basin restriction. That is, the Δ17O system is the only currently available geochemical tracer applicable to the post-Archean rock record that should not be susceptible to “false positives” for low O2. Further, as stressed above, the Δ17O system is the only known oxygen barometer that records a direct atmospheric signal—with the notable exception of the Δ33S system, which has transformed our understanding of atmospheric composition during Archean time (4.0–2.5 Ga) (Farquhar et al., 2000; Pavlov and Kasting, 2002; Bekker et al., 2004). The translation of a sulfate Δ17O value into an atmospheric pO2 estimate requires a model, and thus the leveraging of several assumptions and consideration of uncertain model parameters. Nevertheless, the use of computationally tractable biogeochemical models allows for formal assessment of uncertainty, and in certain cases should yield significant refinement of pO2 estimates during Proterozoic time. We consider this approach a useful strategy to build on moving forward, notwithstanding the additional insights likely to emerge from modeling Δ17O systematics explicitly in 1-D models of atmospheric photochemistry.
Our new atmospheric pO2 estimate for ∼1.4 Ga contrasts with some recent, roughly contemporaneous pO2 estimates. For example, our estimate is higher than estimates inferred from an early Cr isotope study (<0.1% PAL; Planavsky et al., 2014) but much lower than estimates from several marine redox records (<5% PAL; Zhang et al., 2016a). It is possible that the disagreement among proxies simply reflects time-variable atmospheric oxygen levels, as temporally variable oxygen levels are excepted with a small atmospheric oxygen reservoir. However, we emphasize that the empirical and quantitative approaches utilized in reconstructions of mid-Proterozoic pO2 that yield estimates outside of 0.1% to 1% PAL range have been strongly critiqued (e.g., Planavsky et al., 2016; Zhang et al., 2016b; Diamond et al., 2018). In any case, our estimate is consistent with a number of other proxy reconstructions. For instance, more recent interpretations of the Proterozoic Cr isotope record, surface oxygen estimates based on cerium oxidation kinetics, and other works revising classical estimates from Proterozoic paleosols are all consistent with a mid-Proterozoic atmospheric pO2 between 0.1% and 1% PAL (Cole et al., 2016; Gilleaudeau et al., 2016; Liu et al., 2016; Bellefroid et al., 2018).
Robust evidence for low Proterozoic oxygen levels carries important implications for our understanding of long-term ecosystem evolution and the development and maintenance of remotely detectable atmospheric biosignatures. For instance, it has been argued that an Earth system with surface oxygen levels less than ∼1% PAL may not be well suited to foster the diversification and expansion of animal-rich ecosystems (Reinhard et al., 2016). Minimum oxygen requirements for many animals are likely less than 1% PAL (Sperling et al., 2015, 2016) but at such low baseline levels of oxygen and productivity would render most marine environments challenging for early metazoan organisms at some stage of the their life history (Reinhard et al., 2016). Second, this work supports the notion that terrestrial planets can maintain “weakly oxygenated” (pO2 < 1% PAL) states on geologic timescales, rendering certain spectral features of O2 (and O3) potentially difficult to discern (see Reinhard et al., 2017, 2019). This provides an impetus to bolster the observational capabilities associated with the most sensitive O2/O3 spectral features in future direct-imaging exoplanet characterization missions (Schwieterman, 2018).
5. Conclusions
Here we have provided new constraints on atmospheric composition during mid-Proterozoic time by developing a new quantitative technique for interpreting the sedimentary Δ17O record. Building from previous work that has provided evidence for a less productive mid-Proterozoic biosphere (Crockford et al., 2018a), we translate sulfate Δ17O values into a range of possible atmospheric oxygen levels by solving for combinations of pO2, pCO2, and GPP that are consistent with minimum mid-Proterozoic Δ17O values for the ∼1.4 Ga Sibley Group. Following upon several models that have stressed the mechanistic links between marine nutrient levels, biospheric productivity, and atmospheric oxygen (Laakso and Schrag, 2014, 2017; Derry, 2015; Ozaki et al., 2019), we filter these pO2/pCO2/GPP combinations for pO2 and productivity combinations that yield stable Earth system states in the global biogeochemical model CANOPS. Using this approach, we arrive at an estimate of atmospheric pO2 at ∼1.4 Ga of 3.4+5.3 -2.0 × 10−3 times the PAL. Although this approach provides only a snapshot of Earth's evolving atmospheric chemistry, it represents the first quantitative Proterozoic pO2 estimate from an isotopic signal directly exported by the atmosphere. This bolsters the case that environmental conditions inhibited the rise and diversification of complex multicellular heterotrophs during at least portions of mid-Proterozoic time, and provides impetus for developing observational tools for characterizing weakly oxygenated terrestrial planets beyond the solar system.
Code Availability
The inversion code, all necessary input data, and documentation are hosted on GitHub (
Footnotes
Acknowledgments
This work was funded by the NASA Alternative Earths Astrobiology Institute. The authors thank Bryan Killingsworth and two anonymous reviewers for constructive comments that improved the manuscript and Devon Cole, Dave Johnston, Jim Kasting, and Jingjun Liu for critical discussion.