
Abstract
Extreme factors such as space microgravity, radiation, and magnetic field differ from those that occur on Earth. Microgravity may induce and select some microorganisms for physiological, metabolic, and/or genetic variations. This study was conducted to determine the effects of simulated microgravity conditions on the metabolism and gene expression of the probiotic bacterium Lactobacillus reuteri DSM17938. To investigate microbial response to simulated microgravity, two devices—the rotating wall vessel (RWV) and the random positioning machine (RPM)—were used. Microbial growth, reuterin production, and resistance to gastrointestinal passage were assessed, and morphological characteristics were analyzed by scanning electron microscopy. The expression of some selected genes that are responsive to stress conditions and to bile salts stress was evaluated through real-time quantitative polymerase chain reaction assay. Monitoring of bacterial growth, cell size, and shape under simulated microgravity did not reveal differences compared with 1 × g controls. On the contrary, an enhanced production of reuterin and a greater tolerance to the gastrointestinal passage were observed. Moreover, some stress genes were upregulated under RWV conditions, especially after 24 h of treatment, whereas RPM conditions seemed to determine a downregulation over time of the same stress genes. These results show that simulated microgravity could alter some physiological characteristics of L. reuteri DSM17938 with regard to tolerance toward stress conditions encountered on space missions and could be useful to elucidate the adaptation mechanisms of microbes to the space environment.
1. Introduction
The spacecraft environment can be considered a special ecological niche, where some conditions such as gravity, radiation, and acceleration are very different from those on Earth. The focus of recent research has been on the effect of the space environment on human, vegetable, and microbial life-forms (Huang et al., 2018; Lionheart et al., 2018; Strauch et al., 2018; Löbrich and Jeggo, 2019). Microbial life, however, has been found to be more suitable than human or vegetable life because it is easier to manipulate and stable within a wide range of environmental conditions (e.g., temperature, pressure, etc.).
Microbial life is also able to reproduce in a very short period (Senatore et al., 2018). Furthermore, microorganisms negatively or positively affect human life in space, as is the case on Earth. Therefore, the study of space environmental effects on microbial life is crucial to our ability to assess the possible unprecedented interactions between humans and microorganisms. It is, at present, well known that some alterations under spaceflight conditions could cause phenotypic and genotypic variations in microorganisms, concerning modifications on cell morphology and physiology (Nickerson et al., 2004; Li et al., 2015; Huang et al., 2018). Accordingly, altered levels of pathogenicity, viral reactivation, host interaction, and production of metabolites under space conditions have been reported (Leys et al., 2004; Horneck et al., 2010; Taylor, 2015).
It is generally believed that the main factor that determines those changes that occur in microbial response to the space environment is microgravity (Senatore et al., 2018), a term frequently used as a synonym of “weightlessness” and “zero-g,” which indicates that the g-force is not completely neutralized but almost zero (Klaus, 2001; Herranz et al., 2013).
In recent years, there has been a growing interest in the effect of microgravity on pathogenic bacteria as opposed to other microorganisms. In fact, many research studies have documented that this environmental stressor could alter the virulence of potential pathogens such as Salmonella Typhimurium, Escherichia coli, and Staphylococcus aureus (Wilson et al., 2002; Vukanti et al., 2008, 2012; Rosenzweig et al., 2009; Castro et al., 2011). Few investigations have been dedicated to the degree to which spaceflight affects human-associated microorganisms, as reviewed by Saei and Barzegari (2012).
Recently, Turroni et al. (2017) addressed the variation of the gastrointestinal (GI) microbiota of volunteers during a long period in a confined environment like that of spaceflight, while Voorhies and Lorenzi (2016) reported changes in the composition and diversity of gut microbiota of astronauts during long-term space missions. Moreover, several studies have shown that important immune parameters are reduced during spaceflight through the reduction of the number and proportion of lymphocytes, the depression of dendritic cell function and T-cell activation, and reduction in the number of monocytes and precursors of macrophages (Saei and Barzegari, 2012).
It has been suggested that continuous immune alterations in otherwise healthy subjects may have unknown health consequences on long duration missions (Guéguinou et al., 2009). Changes in the GI microbiota and dysregulation of immune cells and cytokines can contribute to cases of gastrointestinal distress (Antonsen et al., 2017), symptoms of anxiety and depression (Slack et al., 2016), skin rashes (Crucian et al., 2014), and viral reactivation (Cohrs et al., 2008). In addition, antibiotic resistance may be increased in spaceflight and make treatment of infections difficult (Taylor and Sommer, 2005).
In this scenario, a regular ingestion of probiotics by space mission crew members should be considered to reduce the risk associated with the presence of microbial pathogens during spaceflight and reduce the antibiotic-associated diarrhea. This would also increase crew resistance to stress and microgravity-induced physiological changes on long duration space missions (Ventura et al., 2012).
Interestingly, the physical forces produced in the spaceflight environment are comparable to those encountered by microorganisms in gastrointestinal, urogenital, and respiratory tracts of the host. In particular some low-fluid-shear areas of the body, such as that between the brush border microvilli of epithelial cells, seem to mimic spaceflight conditions (Nickerson et al., 2004). Accordingly, to investigate the effects of the microgravity environment on microorganisms, some methods in which different technologies are used, such as clinostats, have been implemented (Crabbé et al., 2008; Herranz et al., 2013; Rosenzweig et al., 2014). Such ground-based methods have been developed with the intent to simulate the condition of weightlessness in laboratories (Briegleb, 1992).
One of these bioreactors is the rotating wall vessel (RWV), a clinostat with two rotating axes used instead of one, which was designed at the NASA Johnson Space Center (Houston, TX). The rotation in the RWV occurs on an axis perpendicular to the direction of the gravity force vector (Nickerson et al., 2004).
The other commonly used simulator is the random positioning machine (RPM), an apparatus developed by Hoson at the Osaka City University that changes the position of an accommodated experiment (Hoson et al., 1992, 1997) with two independently rotating frames operating with different speeds and directions and reproducing microgravity in the order of 10−3 g.
These two simulated microgravity systems have in common that the sample is constantly rotated perpendicularly to the gravitational field to hinder a biological system in perceiving the gravitational acceleration vector (Anken, 2013). However, the mixing level differs with respect to the number of rotation axes, the speed, and the direction of rotation (Briegleb et al., 1992; Klaus, 2001). During random movement of the RPM, in fact, continuous acceleration changes experienced by the vessel and consequently by the medium and cells determine an immediate mixing level (Crabbé et al., 2010).
The RWV device can reproduce a constant state of suspension in which cells living in tranquil surroundings are indirectly impacted by changes in the fluid boundary layer that surrounds them (Klaus et al., 1994; Hammond and Hammond, 2001; Yamaguchi et al., 2014). In this system, the gravity vector is randomized, and cells grow with low-fluid-shear (<1 dyn/cm2) under a controlled environment, where the liquid counteracts the slow sedimentation, which creates a constant “free-fall” of the cells through the culture medium. The term low-shear modeled microgravity (LSMMG) has been used to describe the environment produced by the RWV bioreactor in which a dramatic reduction in fluid shear is obtained creating a low-shear stress environment for cell culture, as hypothesized to be a limitation of extracellular mass transport due to the lack of gravity-driven convective flows (Zea et al., 2017).
The RPM system has been less used than RWV for the study of indirect bacterial responses to microgravity, but it is especially suited for studies on plants or mammalian cells in suspension because this system may provide an environment conducive to the successful expansion of stem cell populations in vitro. In fact, it has been reported that this system may provide an environment that is advantageous to the expansion of human stem cell populations in vitro (Anken, 2013). In this context, these two bioreactors are ideal not only for the study of cell behavior under simulated microgravity but also for the evaluation of interactions between the probiotic bacteria and the microvilli of epithelial cells in the intestine and other tissue (Guo et al., 2000; Cervantes and Hong, 2016).
Lactobacillus reuteri DSM17938 has been described as possessing several properties that are believed to be important for its capacity to colonize, adhere to epithelial cells, and produce an antimicrobial compound known as reuterin, 3-hydroxypropionaldehyde, which is produced in the cells by glycerol metabolism. A number of studies have demonstrated the probiotic ability of this microorganism, which was recently reviewed by Srinivasan et al. (2018). Moreover, some papers describe food probiotication by using this microorganism (Malmo et al., 2013; De Prisco et al., 2015; De Prisco and Mauriello, 2016), which is usually ingested by commercial supplements. It is expected that all probiotic properties could be obtained in the spaceflight environment as well and may offer assessment as to the need to protect against gut microbiota imbalance and some aspects of immune dysregulation.
The aim of this work was to investigate the stress response of L. reuteri DSM17938 at the metabolic and transcriptomic levels when cultured under simulated microgravity conditions using the RWV and RPM technologies, and to evaluate whether microgravity condition influences the capability of this probiotic bacteria to exert its positive effect on some human gut disorders.
We investigated growth performance, cell morphology, reuterin production, and resistance to simulated gastrointestinal passage. Moreover, the expression of generic stress genes (groEL, hsp20, dnaK, clpL, ftsH, ctsR, and rpoD), specific stress (i.e., acidic, salt, and bile stress) genes (dpS and rex), genes involved in the intestinal epithelial cell adhesion (map and msa), and genes involved in the synthesis of exopolysaccharides (EPS) (ftf and gtfO) was studied. List and function of selected genes are reported in Table 1. We focused on these genes because they are expressed by L. reuteri during gastrointestinal transit and are involved in the probiotic capabilities.
Selected Genes, Functional Category, and Coding Protein Functions
2. Materials and Methods
2.1. Bacterial strain and growth conditions
The strain L. reuteri DSM17938 was kindly provided by BioGaia (Noos S.r.l.; BioGaia AB, Stockholm, Sweden) and was previously characterized in our laboratory for probiotic properties (De Prisco et al., 2015). It was routinely aerobically cultured in MRS Broth (Oxoid Ltd., Basingstoke, England) at 37°C.
2.2. Simulated microgravity conditions
An overnight culture of L. reuteri DSM17938 was diluted with fresh MRS to reach a cell concentration of ∼106 CFU/mL. Subsequently, eleven 50 mL rotatory cell culture bioreactors (Synthecon, Houston, TX) were filled with the cell suspension taking care that they were completely full and no air bubbles were present to compromise the low-shear condition.
Rotating bioreactors were attached to their base units and rotated on the horizontal axis at 25 rpm at 37°C, here designated as RWV for microgravity treatment (Senatore et al., 2018). Three other vessels were mounted at the center of the bench top of a microgravity simulator through random positioning (RPM 2.0; Dutch Space, Leiden, The Netherlands), the simulator composed of two separated motors that rotate in different directions so as to reach an angular rotation velocity of 60 deg/s, here designated as RPM microgravity treatment (Senatore et al., 2018). Finally, the control cultures (1 × g) were represented by four rotatory bioreactors installed on the base unit of the RWV in vertical axis.
2.3. Growth kinetic
The growth kinetics of bacteria was monitored by measuring absorbance at 600 nm over 72 h.
During the cultivation, 0.1 mL samples were collected from the sampling port, and the same volume was replaced with sterile broth to avoid air bubbles formation. Optical density was recorded at 1 or 2 h intervals up to 30 h, and then two samples were collected at 48 and 72 h. Growth curves were fitted by using DMFit based on the model of Baranyi and Roberts (1994) to calculate maximum growth rate, lag phase duration, and maximum population at stationary phase.
The O2 concentration and pH values were evaluated with 1 mL of 7 h duration, 15 h duration, and 24 h duration samples using the Oxygen Sensor of Knick (Berlin, Germany) and inoLab® pH 7110 (WTW, Weilheim, Germany), respectively. All results are reported as averages ± standard deviation of four replicates treated with RWV, three replicates subjected to RPM device, and four replicates as control treatment. Experiments described below on cell morphology, reuterin production, and GI passage were performed on cultures after 18 h of simulated microgravity exposure.
2.4. Cell morphology by scanning electron microscopy
Samples were analyzed in duplicate with scanning electron microscopy (SEM-Evo 40, Carl Zeiss, Oberkochen, Germany) to examine the morphology of cells after both simulated microgravity treatments. Sample preparation was carried out according to the work of Shao et al. (2017) with little modification.
Two milliliters from the 1 × g, RPM, and RWV cultures was fixed for 2 h at room temperature by adding 1 mL of 2.5% glutaraldehyde. After centrifugation at 6500 g for 5 min, the cell pellet was washed in 1 mL of 0.1 M phosphate-buffered saline (PBS) and dehydrated in an ethanol gradient (30, 50, 70 80, 90, and 100% v/v) for 20 min each. Samples were then processed along three cycles of 30 min contact with 1 mL tert-butyl alcohol (TBA) and centrifugation (6500 g for 5 min). The cell pellet was then resuspended in 100 μL of TBA and incubated overnight at 4°C. After centrifugation the cell pellet was resuspended in 200 μL of PBS for a better dissolution of the pellet, and finally 200 μL of TBA was added. A portion of 20 μL was dropped on a mica plate, air dried at room temperature, coated in gold, and finally observed with SEM.
2.5. Reuterin production assay
L. reuteri was tested for its ability to produce the antimicrobial compound reuterin after RPM and RWV treatments compared with controls. In brief, pellets from 6 mL of cultures were suspended in 250 mmol/L glycerol and incubated at 37°C for 3 h. After that, samples were centrifuged at 6500 g for cell separation, and 20 μL of supernatant was spotted on TSA plates previously inoculated with S. aureus DSM20231. Antimicrobial activity was observed as a growth inhibition zone of the indicator strain, measured by using a calliper and expressed as average of millimeters of the three replicates ± standard deviation.
2.6. Measurement of survival to GI passage
The ability of the cultures to survive simulated GI transit was tested according to the work of Vizoso et al. (2006) as adapted by De Prisco et al. (2015). In brief, 2 mL of cultures was centrifuged (6500 g for 10 min), washed twice with 0.1 M of PBS, suspended in 8 mL of gastric simulated solution (GSS: 5 g/L NaCl, 2.2 g/L KCl, 0.22 g/L CaCl2 and 1.2 g/L NaHCO3, 3 g/L pepsin, pH 2.5), and incubated at 37°C for 120 min under gentle agitation (200 rpm) to simulate peristalsis. After centrifugation, the pellet was suspended in 10 mL of intestinal simulated solution (ISS: 6.4 g/L NaHCO3, 0.239 g/L KCl, 1.28 g/L NaCl, 0.5% bile salts, and 0.1% pancreatin, pH 7.0) and incubated under the same condition of GSS. Cells were enumerated before the start of simulated digestion (T 0) and after GSS and ISS steps, by pour plating on MRS Agar. Results are expressed in percentage of cell survival.
2.7. RNA extraction and Reverse Transcriptase quantitative PCR (RT-qPCR)
L. reuteri cultures at 7 h (initial exponential growth phase), 15 h (end of exponential growth phase), and 24 h (stationary phase) from the RPM and RWV treatments were collected and immediately incubated at a 1:10 ratio (w/v) of RNAlater solution (Ambion, Austin, TX) and stored at −70°C until the RNA isolation.
Three biological replicates of both simulation systems and three control replicates were prepared for the RNA extraction following the Ribo Pure Bacteria kit protocol (Ambion). Cell pellets were washed two times with 1 mL of ice-cold PBS and centrifuged, and the pellets were solubilized in 1 mL of RNA WIZ and transferred in screw-cap tubes containing ∼250 μL of ice-cold zirconia beads (Ambion) and homogenized at maximum speed for 10 min using the Mini-Beadbeater 8 (Biospec Products, Bartlesville, OK).
Each RNA sample was then subjected to a DNase treatment by using the TURBO DNA-free™ Kit (Ambion) and repeating the treatment with 2U of DNase I two times for 30 min. The absence of contaminating genomic DNA in each sample was then determined by using polymerase chain reaction (PCR) and L. reuteri 16S rRNA-specific primers (Table 2). RNA quality and concentration were determined with a Qubit® 2.0 Fluorometer (Life Technologies, Grand Island, NY) and Qubit RNA HS assay kit (Life Technologies) along with NanoDrop 1000 (Thermo Fisher Scientific, Waltham, MA). Synthesis of the cDNA was carried out by starting from 1 μg of total RNA and using a High Capacity cDNA Reverse Transcription Kit (Applied Biosystem, Foster City, CA) according to the manufacturer's instructions.
Primers Used for Reverse Transcriptase quantitative PCR (RT-qPCR) Analysis
RT-qPCR was performed in 96-well plates on a RT-qPCR Chromo4™ System (Biorad, Hercules, CA) using SSoAdvanced Universal SYBR Green (Biorad) as fluorophore. The expression of a total of 13 genes was evaluated.
Amplifications were performed under the following conditions: 95°C for 3 min, 40 cycles of 95°C for 15 s, 58°C for 30 s, and a final step at 72°C for 60 s. At the end of the amplification cycle, a melting analysis was conducted to verify the specificity of the reaction. Differences in gene expression were calculated with the ΔΔCT method after normalizing based on 16S rRNA expression. Data were obtained from three reactions, using RNA from the three independent replicates from each treatment. Data are plotted as average and standard deviation. Genes were considered differentially expressed when they showed a fold change <0.5 or >2.0, and the p-value was <0.05.
The primers for RT-qPCR were constructed with the primer express 3.0 software package (Applied Biosystems; Foster). All primers were verified and aligned with the genome of L. reuteri DSM17938 using Tool BLAST (URL:
2.8. Statistical analysis
The statistical differences were evaluated with t-test and ANOVA test (α = 0.05) using SPSS software (Ver.17.0; SPSS, Inc., Chicago, IL).
3. Results
3.1. Growth profile and growth rate
Results of monitoring and modeling of bacterial growth showed that all cultures started the exponential growth phase after ∼4 h (lag phase duration). Moreover, all cultures showed a maximum growth rate ranging between 0.472 (±0.016) h−1 in RPM and 0.556 (±0.023) h−1 in RWV.
Moreover, all samples showed no significant difference of maximum population level.
3.2. Electron microscopy (SEM) analysis
SEM analysis of cultures subjected to RWV and RPM treatments revealed no differences in shape and size of individual cells (Fig. 1). However, SEM images showed that cells grown under microgravity (Fig. 1B, C) appeared more aggregated compared with the control sample (Fig. 1A), in which cells seemed well separated. A much more homogeneous distribution of cells within the specimen of the control sample was further demonstrated by atomic force microscopy (data not shown) in which no preparation of sample is requested.

Cell morphology images by SEM of Lactobacillus reuteri DSM17938:
3.3. Reuterin production
Results of antimicrobial tests showed that cells grown under both simulated microgravity conditions determined an overproduction of reuterin compared with the control culture. Indeed, the indicator strain showed a growth inhibition halo of 32 ± 0.81 and 31 ± 0.28 mm in RWV and RPM, respectively, against a 22 ± 0.95 mm halo of the control culture (Fig. 2).
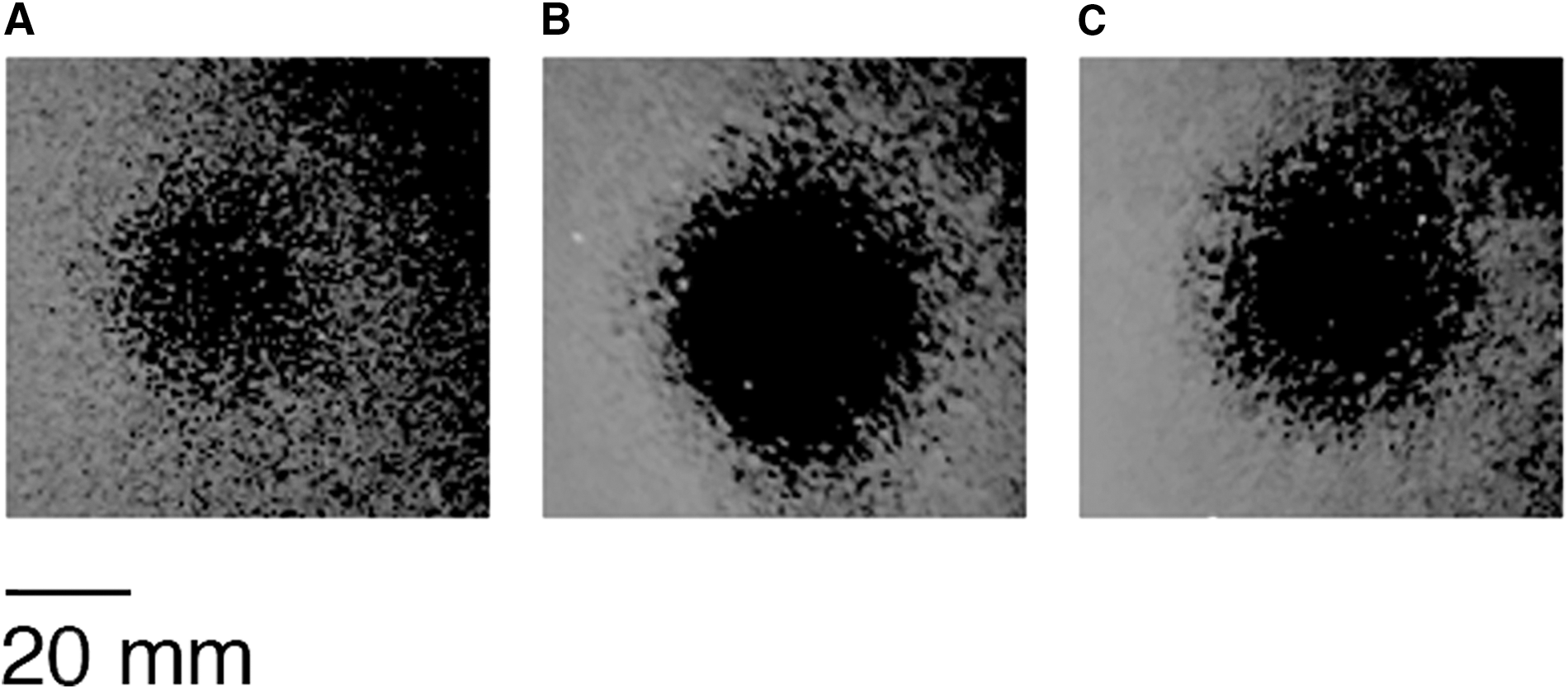
Variation of inhibition halo of Staphylococcus aureus DSM20231 growth on agar plates as effect of simulated microgravity on the reuterin production by the supernatant of Lactobacillus reuteri cultures subjected for 18 h to control condition
3.4. GI stress resistance
The survival rate after 18 h of growing under simulated microgravity and terrestrial gravity conditions was evaluated through the viable counts, and the survival percentage was reduced from 71% after gastric simulating step (GSS) to 31% after the intestinal simulating step (ISS) in the 1 × g samples and from 100% to 62% in RPM samples. The RWV samples seemed to better tolerate the intestine environment because there was less reduction in the survival rate, from 92% to 76% (Fig. 3). Indeed, GSS step determined higher survival rates of cells grown under RPM; whereas ISS exposure determined higher survival rates of cells grown under RWV.
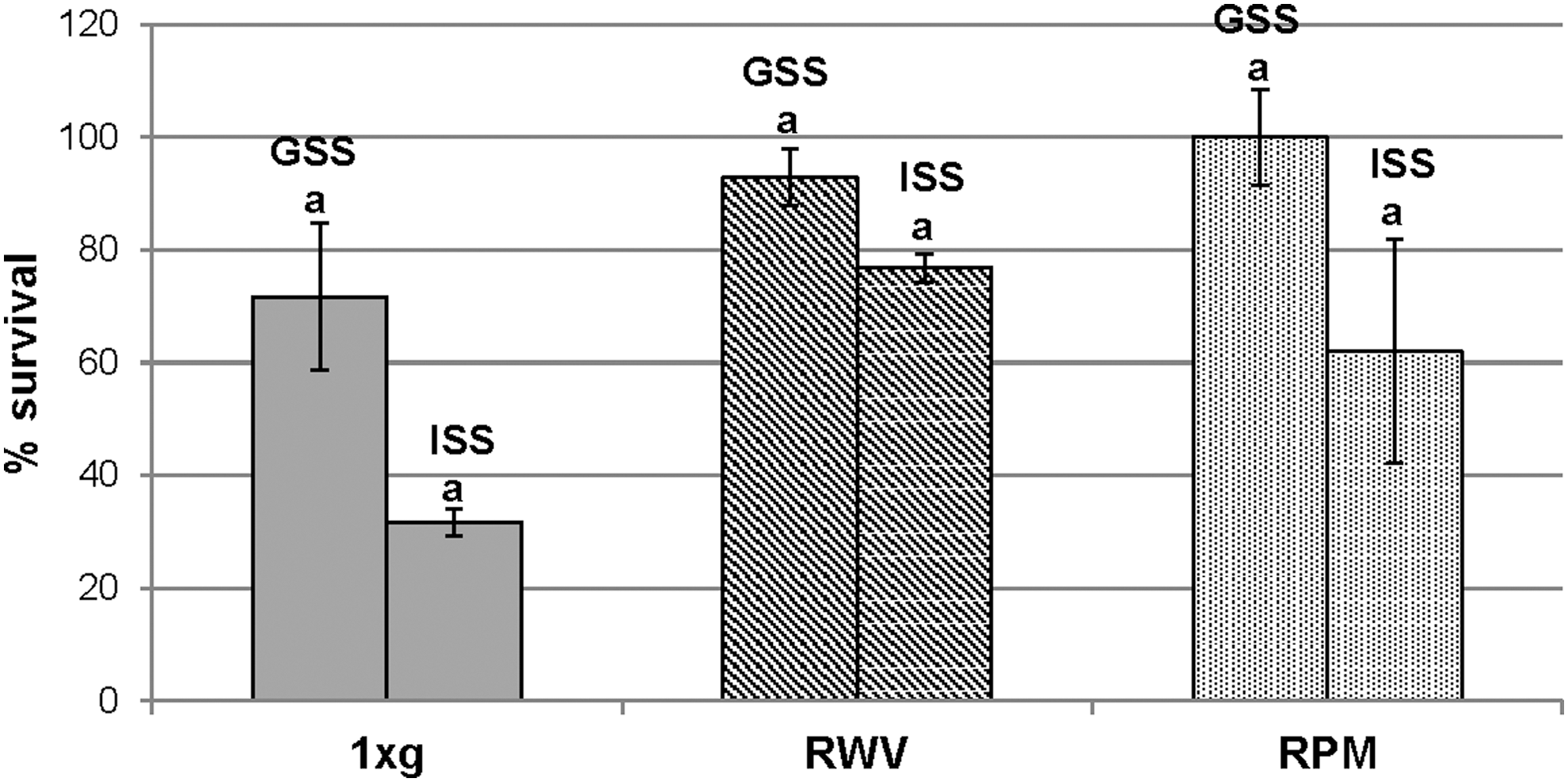
Survival rates of Lactobacillus reuteri exposed to GSS and ISS after 18 h of growth under simulated microgravity conditions and control condition. Different letters indicate significant difference (p < 0.05) between mean values of GSS and ISS as determined by t-test. GSS, gastric simulated solution; ISS, intestinal simulated solution.
ANOVA test was performed comparing GSS step and ISS exposure for the three treatments (1 × g, RPM, and RWV) within the single step of GI passage (time zero, GSS, and ISS) and the three steps of GI passage within the single treatment. Results show no significant difference between the three GI passage steps within RPM samples; on the contrary, a significant difference (p < 0.05) within RWV samples (Table 3) was observed. Results with t-test showed significant differences in cell survival between control and treated samples after GSS exposure, as well as between RPM and RWV samples (p < 0.05) (Fig. 4). Instead, after ISS exposure, a significant difference was shown only between 1 × g and RWV cultures (p < 0.05).

Survival rates of cultures exposed for 18 h to simulated microgravity (RWV and RPM) in comparison with control culture (1 × g) after exposure to GSS and to ISS. Different letters indicate that mean values of GSS and ISS exposure between treatments are significantly different (p < 0.05) as determined by t-test.
Test ANOVA Between the Treatments (1 × g, Rotating Wall Vessel and Random Positioning Machine) for Every Step (Time 0, Gastric Simulated Solution, and Intestinal Simulated Solution) and Between the Three Steps Within Single Treatment
One or two asterisks indicate significant differences between the three treatments or within the single treatment at p < 0.05 or p < 0.01, respectively.
GSS, gastric simulated solution; ISS, intestinal simulated solution; RPM, random positioning machine; RWV, rotating wall vessel.
3.5. Gene expression
The results of the gene expression level experiments are depicted in Fig. 5. RT-qPCR results at 7 h (Fig. 5a) show that most of the investigated genes are not differentially expressed because the fold change remains within the limit of 0.5 and 2, even though all of them showed a significant difference compared with control. However, under RPM microgravity, downregulation of clpL, ctsR, and rpoD genes and overexpression of rex and gtfO genes were shown.
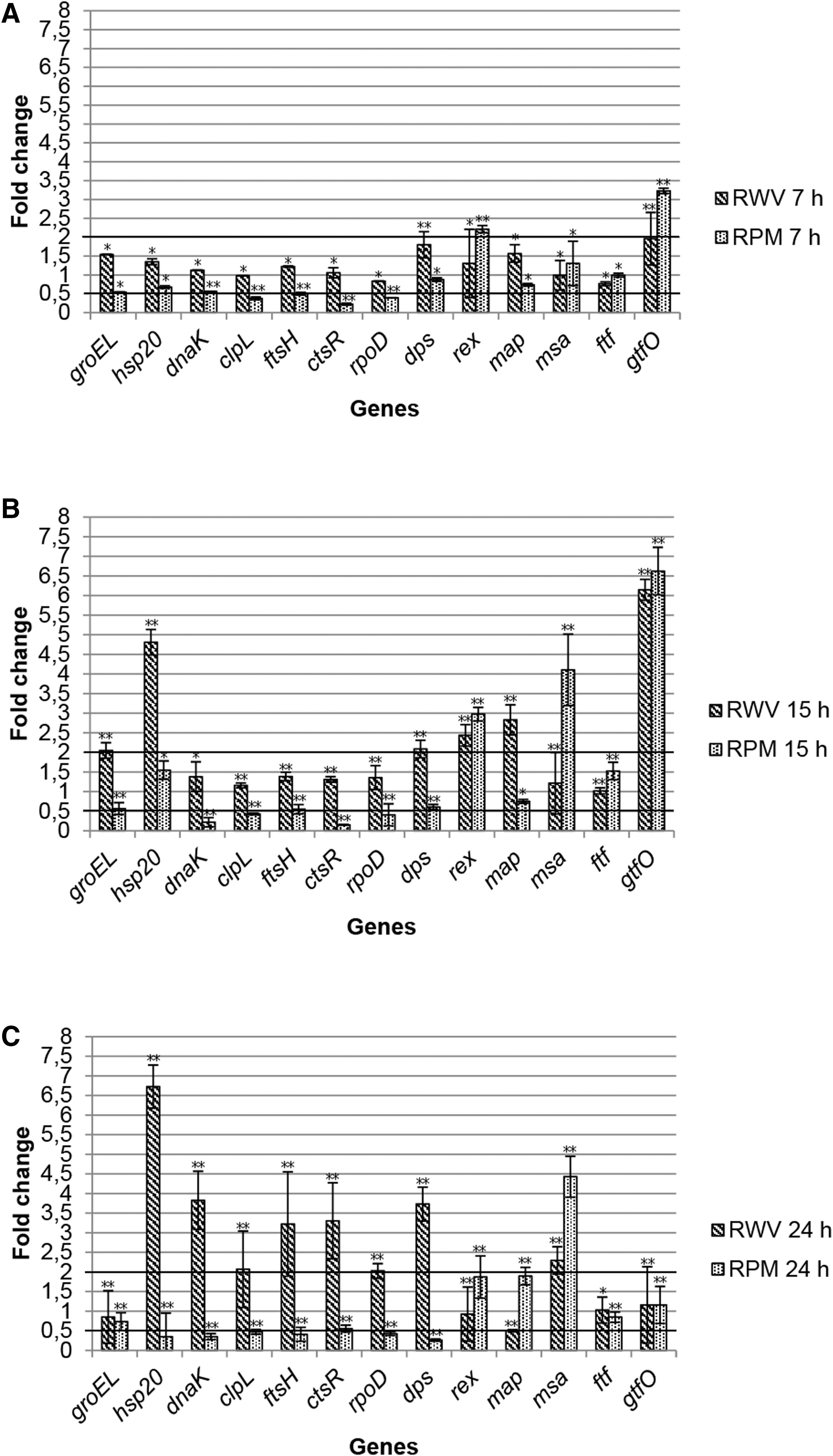
Level of gene expression after 7
Our results show that the overall picture totally changes after 15 and 24 h of growth under simulated microgravity (Fig. 5b, c). In fact, six and seven genes were overexpressed under RWV after 15 and 24 h, respectively. No gene was underexpressed during the growth under the same conditions. Instead, three genes were overexpressed and four underexpressed after 15 h, while one gene was overexpressed and six underexpressed after 24 h under RPM.
After 15 h of growth under microgravity, a significant fold change was registered for most of the genes involved in the generic and specific stress response (hsp20, dnaK, clpL, ctsR, RpoD, and rex) (Fig. 5b), while a significant and out-of-fold change range of 0.5–2 was shown for most of the other genes; that is, some of them involved in the probiosis (i.e., map and msa) of the tested microorganism. Moreover, the high overexpression of gtfO gene—involved in EPS biosynthesis—under both simulated microgravity systems should be underscored.
Results registered after 24 h (Fig. 5c) show a significant fold change in the expression level of most of the tested genes. In particular, the stress genes showed a significant increase and decrease expression level under RWV and RPM conditions, respectively.
4. Discussion
4.1. Growth profile and growth rate
Results of the growth experiments are in agreement with a recent study by Castro-Wallace et al. (2017), which demonstrated that another typical probiotic strain for humans, Lactobacillus acidophilus ATCC 4356, exhibited a growth trend under RWV simulated microgravity, which was similar to that of the control culture. On the contrary, an altered growth profile of the same L. acidophilus strain was reported by Shao et al. (2017), who used a new 3-D RWV system that rotates at 30 rpm. Also, other publications on opportunistic and obligate pathogens have shown alterations in growth profiles when cultivated in LSMMG (Kacena et al., 1997, 1999a, 1999b; Klaus et al., 1997; Leys et al., 2004). Our conflicting results may indicate that every microorganism responds differently to this particular condition.
4.2. Cell morphology and surface characterization
SEM observations did not show any changes in cell morphology, in terms of both shape and size of the cultures treated with RWV and RPM compared with 1 × g cultures. These results are in agreement with a previous work on L. acidophilus in which samples subjected to RWV treatment showed similar morphological and structural features to the control, indicating that on a given day LSMMG treatment had no significant effect on the morphology and structure of these probiotic bacteria (Shao et al., 2017).
The only difference that could be noticed in both treatments was an alteration of the level of cell aggregation, confirming what was reported in the literature for the pathogen Salmonella Typhimurium, which was exposed to real microgravity during the space mission STS-115 (Wilson et al., 2007). This bacterium showed the formation of an extracellular matrix and associated cellular aggregation of spaceflight cells, similarly to that we observed in this study for L. reuteri under RPM condition.
Changes in bacterial biofilm characteristics under a LSMMG environment have been demonstrated for other microorganisms, for which the low-fluid-shear environment stimulates increased EPS production. A culture of Pseudomonas aeruginosa growing in RWV has exhibited self-aggregation, increased alginate production, and a phenotype related to cystic fibrosis pathology in the lung (Crabbè et al., 2008). Moreover, E. coli cultivated under a LSMMG environment produced thicker and more resistant biofilms (Lynch et al., 2006).
4.3. Reuterin production
Reuterin production is increased under simulated microgravity stress, which demonstrates that the results are in accordance with other probiotic bacteria such as L. acidophilus treated with the use of a new 3D RWV system (Shao et al., 2017). LSMMG could also induce increased virulence in pathogenic Salmonella Typhimurium grown aboard the Space Shuttle mission STS-115, which was able to demonstrate an improved percentage mortality in a murine infection model with a shorter time to death and a decreased LD50 (Wilson et al., 2007). On the contrary, most studies on bacterial virulence characteristics under low-shear conditions have indicated no significant differences in virulence, including decreased secondary metabolites production (Taylor, 2015). Indeed, the results obtained from this work are in contrast with previous results on other microorganisms cultured in a RWV apparatus, as was assessed for the first time by Fang et al. (1997). These authors reported that the RWV environment decreased beta-lactam production in Streptomyces clavuligerus, inhibited rapamycin production in Streptomyces hygroscopicus, and prevented microcin B17 production in E. coli.
The reason for the reduced secondary metabolites production could be due to reduced shear stress encountered in the RWV bioreactors (Fang et al., 2000a, 2000b; Demain and Fang, 2001). In our case, the higher production of an antimicrobial substance confirms the previous results for another lactic acid bacteria (Shao et al., 2017), indicating that the low-fluid-shear in which the bacteria grow is perceived as a stimulus in the production of antimicrobial compounds probably because low-fluid-shear reproduces the same conditions as those that occur in the intestinal tract (Nickerson et al., 2004; Arqués et al., 2015).
4.4. Gastrointestinal stress resistance
L. reuteri is a beneficial probiotic organism that inhabits the GI tract. Therefore, it is important to investigate the response of this microorganism to GI passage after exposure to microgravity conditions. The increased resistance of L. acidophilus to acid and bile salts stress (Shao et al., 2017) is similar to the response of L. reuteri in this work, which seems to tolerate the stressful conditions of the stomach and upper intestine.
L. acidophilus showed greater resistance to low concentrations of bile (<0.05%) rather than high concentrations (0.1–0.3%). In our case, L. reuteri cultures treated for 18 h to RPM and RWV systems exhibited resistance to high bile salt concentration (0.5%) because, after the intestinal simulation test, a lower reduction of survival was observed for RPM but more for RWV cultures in comparison with control samples. In fact, in comparison with 1 × g samples, the treated samples to simulated microgravity showed a greater performance in survival immediately after the simulation step of the gastric environment. Moreover, the RWV samples seemed to tolerate the intestinal treatment (ISS) better than the control and RPM samples. These results are in contrast with what was reported in another work on L. acidophilus ATCC 4356 (Castro-Wallace et al., 2017), which highlighted no significant differences in survival between RWV and 1 × g cultures subjected to simulated gastric or small intestinal juice stress.
4.5. Gene expression
Our findings show that the expression level of most genes is affected by simulated microgravity. However, as expected, response is different according to the time of exposition. Moreover, by comparing the RPM and RWV systems, we found substantial differences in the expression level of many selected genes.
The unaltered expression of most genes after 7 h of growth is likely due to the short period of cultivation, during which the modified environment did not alter the mRNA synthesis with respect to control culture. This result is supported by the growth curve, which shows that at 7 h all cultures were still at the beginning of the exponential phase (data not shown). Since the results of gene expression under RWV show a minor difference with respect to RPM conditions, we hypothesize that it is likely RWV conditions can be viewed, at the beginning of growth, as normal for cellular suspension rather than those that occur at the beginning of growth under RPM conditions.
Results of gene expression after 7 h of treatment under both simulated microgravity conditions are similar to those obtained for the other two Lactobacillus strains, which show no alteration or light alteration in the expression level of some stress genes and some key genes of cholesterol metabolism; this is the case as well after 24 h of growth under simulated zero gravity conditions (Castro-Wallace et al., 2017; Shao et al., 2017).
The results at 15 h are analogous to those of two potential pathogens, such as E. coli and Salmonella Typhimurium, that were grown under simulated microgravity conditions, exhibiting a differential level of expression for genes usually known to be subjected to thermal, osmotic, and acid stress, starting as early as the mid-late log phase (16 h) of growth (Wilson et al., 2002, 2007; Vukanti et al., 2008).
In general, our results highlight differences between the two simulated microgravity systems; that is, for RWV conditions there was an upregulation for most of the investigated stress genes that exhibited a general downregulation over time in RPM.
Benoit and Klaus (2007) noted that the two simulations cannot fully reproduce the effects of actual spaceflight. The gravity vector, in fact, is randomized, but fluid convection and shear stress are not completely avoided. Hence, results obtained from cells incubated in real microgravity are frequently similar but not identical (Hemmersbach et al., 2006). Nevertheless, between two systems, the RWV device is believed to be more appropriate in the study of LSMMG-induced response of cells in a suspension environment, as opposed to the RPM. In fact, similarities between spaceflight and LSMMG responses were observed, and can presumably be ascribed to the analogous low-fluid-shear conditions in-flight and in the RWV.
Fluid mixing assessment in both systems is believed to be a pivotal factor that contributes to transcriptional differences between L. reuteri cultures grown in RWV and RPM. These simulation systems can determine different effects due to the different mixing levels. Despite the fact that research objectives when implementing these two devices are often the same, results may be reached in different ways. It is likely that the cells react differently to the growth environment by exhibiting gene variations based on cultivation conditions. Interestingly, fold change of most stress genes after 15 and 24 h emphasized differences when comparing the results of RWV and RPM systems analysis. In particular cells grown under RWV microgravity exhibit a tendency to increase the expression level of these genes over time, whereas under RPM microgravity there is an opposite trend.
The low-fluid-shear conditions reproduced by RWV may be perceived by the cell as an alteration in oxidative state that involves two specific genes such as rex and dps. The change of oxidative conditions is more evident under RWV treatment than under RPM treatment, determining an increase with time of dps expression under the RWV environment. This gene is normally involved in a different type of stress response in E. coli, such as oxidative stress, irradiation, metal toxicity, heat stress, and pH stress (Martinez and Kolter, 1997; Nair and Finkel, 2004).
Our results, in regard to dps gene expression, do not reflect those obtained by Wilson et al. (2007) in another whole transcriptional analysis of a spaceflight culture of Salmonella Typhimurium, which revealed a down expression of the dps gene. The expression level of the rex gene underwent an increase from 7 to 15 h of both growth conditions, which is in agreement with results of RNA-seq analysis with regard to the bacterium Streptococcus mutans, which was subjected to 8 h of simulated microgravity conditions (Orsini et al., 2017). In particular this bacterium showed an overexpression of genes involved in the transport of carbohydrates, in biofilm formation, and in oxidative stress resistance in response to cellular redox, whose expression is regulated by the transcriptional regulator rex. This gene is, moreover, implicated in the transcriptional response of S. aureus to LSMMG by the altered expression of rex-regulated genes under this growth condition (Castro et al., 2011).
Moreover, only one gene involved in EPS synthesis, that is gtfO, showed a surprising increase in the expression level as soon as 7 h after the two treatments. This gene, which encoded a glucosyltransferase, a sucrose-type enzyme involved in EPS synthesis, is probably directly responsible for production of biofilm observed through SEM analysis in both cultures subjected to 18 h treatments.
RpoD is upregulated only after 24 h of RWV condition: this gene codifies a sigma factor that is activated with rpoS upon exposure to oxidative stress signal that puts the cell in a “protective state.” In E. coli AMS6, the stress response to RWV treatment depends on rpoS expression that is higher only in the stationary phase of growth (Lynch et al., 2004), which corroborates our results.
Genes coding for more regulative and/or catalytic functions, that is, ctsr and ftsh, exhibited major values of variation in fold change under RWV after 24 h of treatment. Recently, ftsH was identified as a chaperone-protease that belongs to a novel member of the class three stress gene Repressor (ctsR) regulon in Lactobacillus plantarum (Fiocco et al., 2009, 2010; Bove et al., 2012; Russo et al., 2012). As ctsR, this enzyme is involved in cell protection against environmental stress conditions such as those encountered during the human oro-gastric-intestinal transit (Bove et al., 2013).
A modification of the expression level of these two genes only under RWV treatment suggests that this system is comparable to that encountered in GI environment.
Interestingly, the high induction of the dnaK chaperone gene and the heat shock gene hsp20 might account for their putative additional roles in mechanism of responses to microgravity conditions. These two genes are sensitive to the condition of low-fluid-shear reproduced by the RWV system, and a strong expression increase is evident at 24 h of treatment. Notably, the hsp20 gene exhibited a marked increase in fold change over time under RWV treatment and a decrease under RPM. This gene, like the other hsp genes, responds to varying stress conditions such as heat, cold, acid, osmotic, oxygen, high pressure, and starvation stresses in many Lactobacillus strains (De Angelis and Gobbetti, 2004).
Our results with regard to dnaK gene expression are in agreement with a microarray study on Salmonella Typhimurium grown under simulated microgravity conditions (Chopra et al., 2006) but are in contrast with another transcriptional study on E. coli K12, which showed a downregulation of this gene under modeled reduced gravity (Vukanti et al., 2008).
Considering genes of probiosis, map and msa, only the latter showed an increase of expression over time under the two different treatments. This increment could be indicative of the major ability of this probiotic microorganism to colonize the intestinal surface and to have a competitive exclusion of pathogens even if this capability is regulated by many other genes.
It is interesting to highlight the differences over time at the transcriptomic level of cultures subjected to RPM and RWV conditions, which reflect the different ways of these devices to reproduce the microgravity environment. Differences between RPM and RWV systems were already addressed by other authors who considered the molecular effect induced by both devices (Mastroleo et al., 2013). They evaluated the whole transcriptome of the MELiSSA bacterium Rhodospirillum rubrum under both conditions and showed a significant response to cultivation in simulated microgravity at the transcriptomic, proteomic, and metabolic levels. All 13 genes upregulated under RWV conditions were also included in the more pronounced response to RPM cultivation but with higher fold induction in the latter, which was not the case when comparing the response to both devices at the proteomic level.
This is in contrast with our RPM results, which indicate that the molecular response of L. reuteri was not altered and demonstrate that the responses to simulated spaceflight conditions are species specific and depend on growth conditions. Crabbé et al. (2010) showed conflicting results of the two microgravity simulation methods and reported the microarray analysis of P. aeruginosa PAO1, which was grown in the two different spaceflight-analog culture systems. Analysis of differential gene expression by these authors revealed 134 and 9 genes induced under RWV and RPM, respectively, compared with the normal gravity control, which indicates that RPM and the normal gravity were sensed in a similar manner by P. aeruginosa PAO1, while LSMMG conditions reproduced by RWV led to a more divergent transcriptomic profile.
On the contrary, our divergent results regarding the fold change of many selected genes after 24 h of cultivation under two different microgravity environments show that these conditions and the 1 × g controls were recognized in time as different growth conditions by L. reuteri.
5. Conclusion
In conclusion, this study revealed that simulated microgravity conditions could significantly impact the biological characteristics of the tested L. reuteri, in various aspects, including bile tolerance, antimicrobial property, and gene expression. Overall, the study offers important information for a better understanding of the influence of simulated microgravity on probiotic bacteria, and thus may contribute to protect the health status of astronauts on long-term space missions. Moreover, our results confirm that different simulated microgravity systems can have dissimilar effects on bacterial cells depending on their capability to mimic real microgravity conditions.
These experiments should be considered preliminary to further experiments in the spaceflight environment to develop a comprehensive understanding of the impact of real microgravity conditions on the probiotic bacteria with the final aim to provide these microorganisms to the crew members during long-term missions.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
G.S. is a PhD student supported by a grant from the European Space Agency through the MELiSSA project.
Associate Editor: Petra Rettberg