
Abstract
Antarctica is an ideal analogue for studying the limits of life. Despite severe temperature fluctuations and desiccating conditions, life is commonly found colonizing the structural cavities within Antarctic rocks (i.e., endoliths). Previous studies have speculated that the slow cycling of endoliths in the McMurdo Dry Valleys may be the limit of life on Earth. However, very little is known about the in situ activities of these communities—especially in regions outside the McMurdo Dry Valleys where endoliths are thought to be cycling carbon very slowly (e.g., hundreds of years). Here, we show that East Antarctic endoliths found on nunataks are cycling carbon quickly and are therefore quite active. Through radiocarbon (14C) analyses of the viable cell membrane (as phospholipid-derived fatty acids [PLFA]), we found that the Δ14C composition of these microbial communities was on average predominantly modern, with a few samples signaling older carbon in the system. These findings indicate that endoliths inhabiting inland Antarctic nunataks are cycling carbon on decadal timescales, which support the notion that endoliths in Antarctica are cycling carbon quickly. This work provides new insights into the potential variability of Antarctic endolith activities and demonstrates that, despite the climatic extremes that exist farther inland on the most inhospitable continent on Earth, indigenous life can thrive.
1. Introduction
Life as we know it generally exists within a confined set of parameters. Extremophiles, defined as organisms that survive and even thrive in locations that experience one or more extreme (physical or geochemical), have long been studied to explore the extent of life on Earth with an eye on future astrobiological applications. After plants, microbes are the most abundant organisms on Earth (Bar-On et al., 2018) and, as a result of their ability to colonize advantageous niches, are often the only life-form found in extreme environments.
Endolithic, or “rock-dwelling,” microorganisms are specialized in that they can colonize the interior of any type of rock. They can flourish in temperate climates or persist under harsh conditions and are found across a wide variety of environments (Walker and Pace, 2007). In general, endoliths preferentially colonize rocks with naturally higher translucence and porosity (Friedmann, 1982), such as sandstone, to maximize photosynthesis. In addition, these microbes have been found to colonize rocks that have either been metamorphically altered to possess these traits (e.g., shocked gneisses) or have undergone significant weathering (Cockell et al., 2002).
Endoliths are often the only form of life found in extreme terrestrial environments, and their ability to survive has been attributed to the protection provided by their microhabitats from environmental stressors (Friedmann and Ocampo, 1976; Friedmann, 1982). Colonization of the structural cavities within approximately 1–2 mm of the rock surface is favorable as it can be up to 10°C warmer in these spaces than the surrounding air on a sunny day (McKay et al., 1993) and provides sufficient exposure to sunlight while sheltering inhabitants from harmful ultraviolet radiation (Hughes and Lawley, 2003). Despite harsh katabatic winds, arid conditions, and freezing temperatures, endoliths have been found in one of the most extreme locations on Earth—Antarctica.
Antarctica, a remnant of the ancient supercontinent Gondwana that broke up between 180 and 36 million years ago after the arrival of a mantle plume caused crustal rifting, is primarily divided into East Antarctica, an older continental craton, and the younger West Antarctica, which is actively rifting and includes the Antarctic Peninsula. The Antarctic Ice Sheet, which contains ∼30 million km3 of ice (National Snow and Ice Data Center, n.d.) and covers 98% of the continent, is similarly considered to be composed of two separate masses that are divided by the Transantarctic Mountains: The West Antarctic Ice Sheet and the East Antarctic Ice Sheet. The East Antarctic Ice Sheet is the largest mass of ice on Earth and is extremely thick, reaching up to 5 km in thickness for some regions (Hale, 2014).
The ice-free areas of Antarctica are usually limited to northern coastal areas, isolated rocky peaks (commonly called “nunataks”), and dry valleys extending above the ice and snow. Previous studies have found living endolithic microorganisms colonizing rocks located within these ice-free areas (Friedmann and Ocampo, 1976; Friedmann, 1980; Friedmann, 1982; Friedmann et al., 1988; Vestal, 1988a; Johnston and Vestal, 1991; Wierzchos and Ascaso, 2001; Hughes and Lawley, 2003; Wierzchos et al., 2005; Büdel et al., 2008; Pointing et al., 2010; Colesie et al., 2016; Brady et al., 2018).
Antarctic endolithic communities are considered one of the simplest known ecosystems, with cyanobacteria as the primary producers, fungi as the consumers, and bacteria as the decomposers (Friedmann, 1982). To adapt to unfavorable conditions, it is thought that these microorganisms are able to switch from active to dormant metabolic states (Friedmann and Ocampo, 1976). To partially compensate for the seasonally induced light restriction, Antarctic endoliths preferentially colonize north-facing rocks to augment photosynthetic activity (Friedmann, 1982). Previous observations suggested that these communities are likely slow growing due to short austral summers and long winters, where sunlight is limited and exposure to extremely cold temperatures is typical throughout the year.
Despite previous studies, the viability and activities of Antarctic endolithic microorganisms remain uncertain. Both viability and activity are typically measured through culturing, microscopy, the addition of isotopic labels, and natural abundance radiocarbon (14C) analysis. Previous culture studies were not successful (Friedmann et al., 1988) and were overall inadequate given the low culturability of most microorganisms (Amann et al., 1995). However, the remaining methods—microscopy, isotopic labeling, and natural abundance 14C analysis—have successfully shown that endoliths in Antarctica are viable.
Microscopic analyses found that the physiological states of Dry Valley microbial cells vary as a function of elemental composition (Mulyukin et al., 2002) and fluorescence (Wierzchos et al., 2004), suggesting that these endoliths were metabolically active. Additional evidence of metabolically active endoliths was observed in situ as exfoliative trace fossils left behind by still growing communities (Friedmann and Weed, 1987). Carbon metabolism (primarily via photosynthesis) was observed over a wide range of temperatures, light intensities, and moisture regimes through the addition of various organic and inorganic compounds during in situ and in vitro studies (Vestal, 1988b).
Later, isotopically 14C-labeled bicarbonate studies applied to lichen- and cyanobacteria-dominated colonies found very low rates of photosynthetic carbon incorporation times and extremely long turnover times (Johnston and Vestal, 1991). Recent radiorespiration assays also found that Dry Valley cryptoendoliths had higher respiration rates than local permafrost soil microbiota (Goordial et al., 2016). While these studies were instrumental in understanding the viability of Antarctic endoliths, they have yet to address the activities of these communities.
The analysis of naturally abundant radiocarbon in certain biomarkers can be used to determine microbial carbon sources (Petsch et al., 2001; Slater et al., 2005), thus providing some indication of cycling rates in natural systems. Phospholipid-derived fatty acids (PLFA) are components of cellular membranes that may hydrolyze within days of cell death (White et al., 1979b; Harvey et al., 1986) and are therefore considered a biomarker of the viable microbial community. In general, PLFA structures differ between various microorganisms and, although some types are more ubiquitous, can also be used to get a broad brushstroke of community composition. With regard to the concept of “you are what you eat,” the isotopic content of PLFA is considered representative of the isotopic content of carbon sources to the microbes (Petsch et al., 2001; Slater et al., 2005).
Since Antarctic endoliths have been found to preferentially consume atmospheric CO2 (Vestal, 1988b) and the atmospheric composition of 14CO2 is well known, the 14C content of the PLFA can be used to determine the rate of carbon cycling (i.e., the activity) of an endolithic community (Ziolkowski et al., 2013b), or how much time has passed since the PLFA carbon was last in equilibrium with the atmosphere. Recent work in the McMurdo Dry Valleys found younger 14C ages in endoliths at lower elevations that were proximal to the coast, where warmer temperatures and greater water availability lead to higher productivity within a community (Colesie et al., 2016).
Isotopic analyses of PLFA have also been applied to assess carbon sourcing and cycling by endoliths in the Dry Valleys (Brady et al., 2018). The radiocarbon ages of microbial PLFA were found to be younger than the total organic carbon (TOC)—comprising both dead and living biomass—ages of the rocks they were living in, suggesting fixation of modern atmospheric CO2 (amid some mixing with older carbon sources). Therefore, through use of natural abundance radiocarbon to assess the microbial activity of endoliths, this study found that they were cycling carbon faster than previously thought (Brady et al., 2018). While this work has substantially contributed to ascertaining the activities of West Antarctic endolithic microorganisms, very little is known about communities residing elsewhere across the continent.
Recently, endoliths have been observed colonizing rocks within the Dronning Maud Land region of East Antarctica. Situated at a lower latitude than those found within the McMurdo Dry Valleys, these endoliths are hypothesized to experience less extreme environmental fluctuations—representing a potentially distinct environment for growth. To date, limited work has been carried out to assess the overall viability and carbon cycling within these communities. To address this knowledge gap, this study used 14C analysis of phospholipids to examine the activities (as carbon cycling) of endolithic microorganisms in East Antarctica.
2. Methods
2.1. Study site and sample collection
Encompassing about 2,741,000 km2 of the area in East Antarctica, Dronning Maud (or Queen Maud) Land comprises one sixth of the entire continent and lies roughly between 20° W and 45° E. Rock samples containing endoliths were collected from several nunataks and valleys in the Sør Rondane Mountains of East Dronning Maud Land, ∼200 km inland from the coast (Fig. 1). Field excursions were based out of the Belgian Princess Elisabeth Antarctic Station (PEAS), which is located along Utsteinen Ridge within East Dronning Maud Land. This area is geographically distinct from the McMurdo Dry Valleys in West Antarctica. Not only is there more than 3000 km of distance between the two stations, but the samples in this study are from much further inland than the Dry Valleys (∼200 km vs. 10s of km from the sea in the Dry Valleys) and from distinctly different latitudes (71° S vs. 77° S). Thus, these samples represent a potentially different habitat than the Dry Valleys.
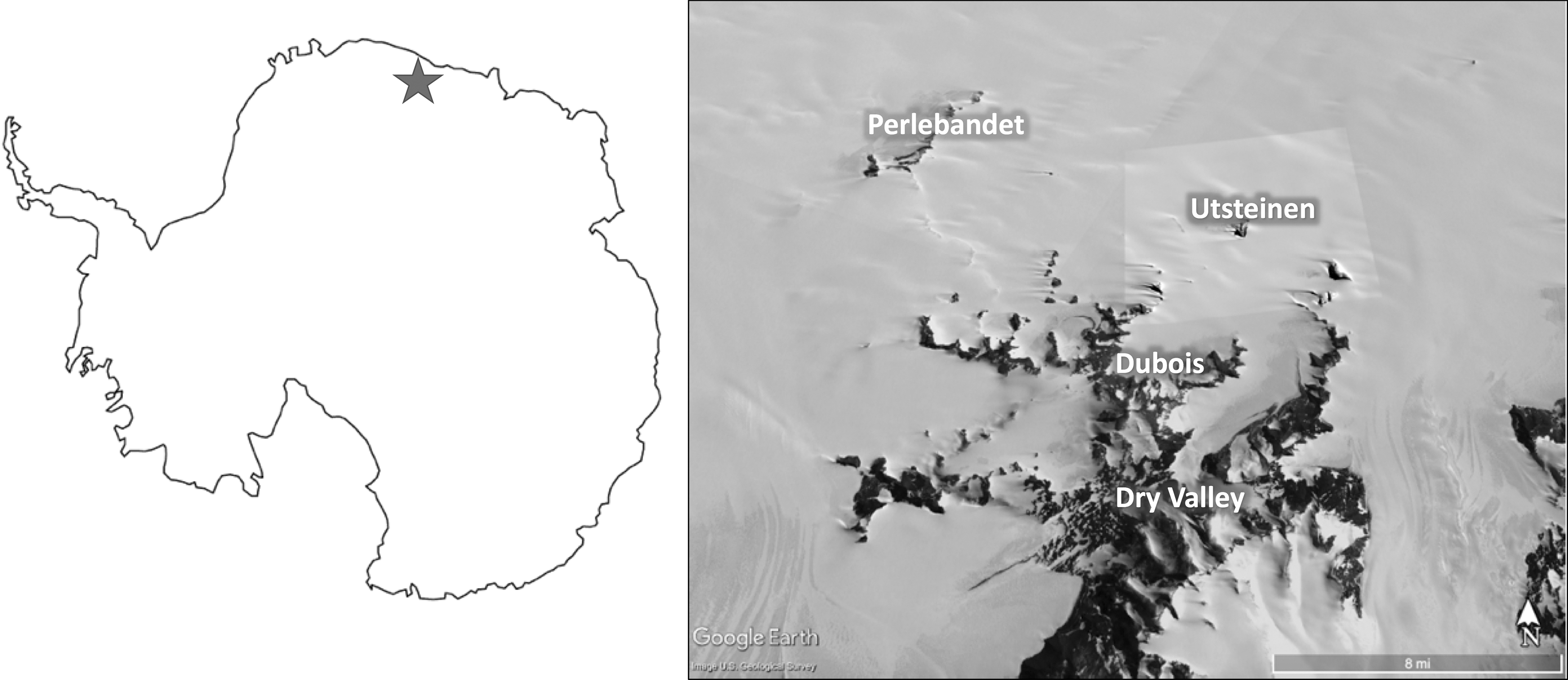
Study area map showing the location of the Sør Rondane Mountains in East Antarctica and sites where endolith samples were collected: Utsteinen, Dubois, Dry Valley, and Perlebandet.
Utsteinen Nunatak, where PEAS is based, is ∼1360 meters above sea level (m.a.s.l.) and located at 71°56′42.26”S, 23°20′42”E. The mean annual air temperature, measured in 2009 and 2010, was approximately −19°C (Gorodetskaya et al., 2013). Perlebandet Nunatak is about 1280 m.a.s.l. and just under 21 km northwest of Utsteinen Ridge, whereas Dubois is about 1350 m.a.s.l. and almost 9 km south of Utsteinen. The Dry Valley in this study is ∼1670 m.a.s.l. and is located about 19 km southwest from Utsteinen. Many of the nunataks in this region are sheltered from the katabatic winds that come from the plateau, which provides a favorable environment for the microbial communities (e.g., Tytgat et al., 2016). Due to its high elevation and proximity to the Antarctic plateau, the Dry Valley site was the windiest location and typically had little snow accumulation as a result.
Endolithic colonies were identified by pigmentation that extended from the rock surface to approximately 1–2 mm into the subsurface (Fig. 2). A total of 23 samples were collected over two austral summers during early 2017 and late 2017 through early 2018. Most samples consisted of highly weathered, extremely friable lithologic fragments. Rock fragments were broken free with a rock hammer and sealed in sterilized plastic bags. Samples were frozen immediately after collection and kept frozen until analysis back at the University of South Carolina. Of the 23 samples collected, a total of 11 rock samples were processed for PLFA analyses and 5 of those 11 samples were selected for isotopic analysis.

Near-surface pigmentation of an endolith sample, indicating microbial colonization, with rock hammer for scale. Color images are available online.
2.2. Extraction and identification of membrane lipids
Colonized rocks were initially crushed with a solvent rinsed impact mortar and pestle and further homogenized with a solvent-rinsed ceramic mortar and pestle. For each sample, ∼80 g of colonized rock was solvent extracted for PLFA twice by using a modified Bligh and Dyer method (White et al., 1979b). Homogenized and pulverized samples were sonicated for 2 min with a 1:2:0.8 dichloromethane (DCM):methanol (MeOH):phosphate buffer solution and allowed to extract overnight at room temperature. The extraction solution was gravity filtered through solvent-rinsed glass fiber filters into a separatory funnel and the solvent ratios adjusted to 1:1:0.9 DCM:MeOH:H2O through the addition of equal parts of DCM and deionized water. The resulting solution was shaken and then allowed to equilibrate into aqueous (upper) and organic (lower) phases. The organic phase was collected, concentrated under a stream of ultra-high purity (UHP) nitrogen, and silica gel chromatography was used to separate the total lipid extract into the neutral lipids fraction (DCM), glycolipid fraction (acetone), and phospholipid fraction (MeOH) (White et al., 1979b). The neutral lipids and glycolipid fractions were stored for future analyses, whereas the phospholipid fraction was evaporated to dryness under a stream of UHP nitrogen then reacted to fatty acid methyl esters (FAME) through mild alkaline transesterification (Guckert et al., 1985). FAME were further purified through secondary silica gel chromatography with 5 mL hexane, 5 mL of DCM, and 5 mL MeOH. The FAME-containing DCM fraction was blown down to 1 mL under a stream of UHP nitrogen.
FAME were quantified and identified on an Agilent 7890B/5977A gas chromatography-mass spectrometer, where 1 μL of FAME was injected by using splitless mode into an HP-5MS column with helium (He) as a carrier gas. The temperature program for separating FAME began at 50°C, increased at 20°C/min up to 130°C, then at 4°C/min up to 160°C and held isothermal for 5 min, and then increased at 8°C/min up to 300°C and held isothermal for 5 min. Quantification of FAME was completed using total ion current and external standards. FAME were identified through comparison of mass fragmentation patterns and retention times to commercially available standards (Bacterial Standards and Supelco 37 FAME mix; Matreya, Inc.).
Fatty acids are composed of long chains of hydrogen and carbon and can either be saturated or unsaturated. Saturated fatty acids (SFA) have a single bond between carbon atoms (resulting in saturation with hydrogen atoms), and unsaturated fatty acids have one or more double bonds. The naming convention for PLFA is based on the number of carbon atoms, followed by how many double bonds are present with some indication of the bond location. For example, 14:0 is a SFA that consists of 14 carbons with single bonds only, whereas 16:1 is an unsaturated fatty acid that consists of 16 carbon atoms with a single double bond.
The observed FAME were grouped into five categories based on these structural differences: saturated, monounsaturated, polyunsaturated, branched, and cyclopropyl. Whereas monounsaturated FAME have only one double bond, polyunsaturated FAME have multiple double bonds (e.g., 16:2, 16:3). Branched chain fatty acids (BCFA) are typically saturated and have methyl branches on the carbon chain, the most common of which are iso- and anteiso-branched fatty acids. Cyclopropyl fatty acids are saturated with respect to hydrogen and contain a ring-like structure within the molecule.
2.3. TOC preparation
Endolith samples for TOC analysis were homogenized and baked at 80°C for 48 h to remove any remaining water. Approximately 1 g of each sample was fumigated with hydrochloric acid for an additional 48 h to remove any trace inorganic carbon. Finally, fumigated samples were baked again (post-acidification) at 80°C for 48 h to remove excess moisture.
2.4. Isotopic analyses
Radiocarbon analysis of the FAME was performed as a compound class rather than on individual fatty acids. Extracted FAME with at least 50 μg C were transferred with 1 mL of DCM to prebaked 6 mm quartz tubes and dried under a stream of UHP nitrogen. Cupric oxide and silver wire were added to the tubes, and they were evacuated on the vacuum line before being flame sealed. The flame-sealed evacuated tubes were baked at 900°C for 2 h. The evolved CO2 was purified and quantified before being sent to the University of Georgia's Center for Isotopic Analysis for 14C analysis with a CAIS 0.5 MeV accelerator mass spectrometer and separate 13C analysis with a stable isotope ratio mass spectrometer. The transesterification cleaves the polar head group from the molecule and replaces it with a methyl. The MeOH used during transesterification was assumed to be radiocarbon-free (Δ14C = −1000‰), and the following equation was used to correct for this:
where N is the number of carbon atoms within the individual fatty acid compounds, Δ14CMeOH is the Δ14C composition of the MeOH used in the transesterification (assumed to be −1000‰), Δ14Cmeasured is the isotopic composition of the FAME, and Δ14CPLFA is the isotopic composition of the PLFA. The radiocarbon data are reported in Δ14C notation and expressed in per mill (‰), whereas the δ13C values are reported in standard delta notation relative to Pee Dee Belemnite (PDB). In addition, the Δ14C values were corrected to the δ13C values of −25% to correct for isotopic fractionation. For the PLFA Δ14C values, the error was estimated to be ±20‰. For the TOC Δ14C values, the error was estimated to be ±5‰.
3. Results
3.1. TOC of East Antarctic rocks
The TOC content within the rock fragments ranged from 0.2% to 1.1% (Table 1). The Δ14C values of the TOC within these samples were overall depleted relative to the recent atmosphere, ranging from −164‰ to −4‰ ± 5‰. In four of the five TOC samples, the Δ14CTOC was significantly depleted relative to the current atmosphere: Dry Valley 1 (−164‰ ± 5‰), Dubois (−138‰ ± 5‰), Perlebandet (−90‰ ± 5‰), and Dry Valley 2 (−70‰ ± 5‰). The corresponding ages for these TOC samples ranged from 1435 to 579 years before present (BP) in uncalibrated 14C years. The Δ14CTOC from Utsteinen was the least depleted (−4‰ ± 5‰), indicating the presence of decadally aged carbon—about 33 years BP in uncalibrated 14C years. The δ13C values of the TOC were depleted and ranged from −34‰ to −26‰ ± 0.5‰. The δ13CTOC from Dry Valley 1 was the most isotopically depleted (−34‰ ± 0.5‰), followed by Utsteinen (−32‰ ± 0.5‰) and Dubois (−31‰ ± 0.5‰). The δ13CTOC values from Dry Valley 2 (−27‰ ± 0.5‰) and Perlebandet (−26‰ ± 0.5‰) were the least isotopically depleted among the endolithic TOC samples analyzed.
East Antarctic Endolith Data Showing Location Elevations, % Total Organic Carbon, δ13C Values for Total Organic Carbon and Viable Membrane Lipid (Phospholipid-Derived Fatty Acids), Δ 14 C Values for Total Organic Carbon and Phospholipid-Derived Fatty Acids, Based on Phospholipid-Derived Fatty Acid Concentrations
C = radiocarbon; m.a.s.l. = meters above sea level; PLFA = phospholipid-derived fatty acid; TOC = total organic carbon; Years BP = years before present.
3.2. Phospholipid-derived FAME: abundance and isotopic composition
PLFA concentrations ranged from 0.9 to 6.9 μg/g (Table 2). The highest abundance of PLFA was observed at Utsteinen (6.9 μg/g), followed by Dubois (3.6 μg/g), Dry Valley 2 (2.8 μg/g), and Dry Valley 1 (1.3 μg/g). Perlebandet had the lowest abundance of PLFA at 0.9 μg/g. A generic conversion factor of 2 × 104 cells/pmol (Green and Scow, 2000) was used to convert PLFA concentrations to viable cell estimates. The calculated cell abundances ranged from 6.5 × 107 to 5.0 × 108 cells/g of sample, where the greatest number of viable cells were observed at Utsteinen (5.0 × 108 cells/g) and were an order of magnitude higher than the estimate for Perlebandet (6.5 × 107 cells/g). While great care was taken to collect areas with the greatest colonization (i.e., more pigment), it is very possible that uncolonized zones within the rocks were incorporated during the sampling process—resulting in diluted lipid concentrations and cell abundances. It is therefore possible that the actual PLFA and viable cell values are higher than what was measured.
East Antarctic Endolith Data Showing Phospholipid-Derived Fatty Acid Concentrations (μg/g), Estimated Cell Abundance (No. of Cells/g Sample) Based on Phospholipid-Derived Fatty Acid Concentrations, and the Distributions of Different Types of Phospholipid-Derived Fatty Acids (mol %) for Each Sample
MUFA = monounsaturated fatty acids; PUFA = polyunsaturated fatty acids.
Overall, the distribution of FAME varied at each location—indicating variability in microbial community composition (Fig. 3). The most abundant FAME were polyunsaturated fatty acids (PUFA), predominantly 18:2 and 18:3, which are fairly ubiquitous types of FAME among many different microorganisms. However, they are generally known to be indicative of fungi (Frostegard and Bååth, 1996; Zelles, 1999), algae (Boschker and Middleburg, 2002), and cyanobacteria (Zelles, 1999; Dijkman et al., 2010) within a microbial community. PUFA comprised 64% of the FAME for Dubois, 63% of the FAME for Utsteinen, 56% of the FAME for Dry Valley 2, 15% of the FAME for Perlebandet, and 9% of the FAME for Dry Valley 1 (Table 2).
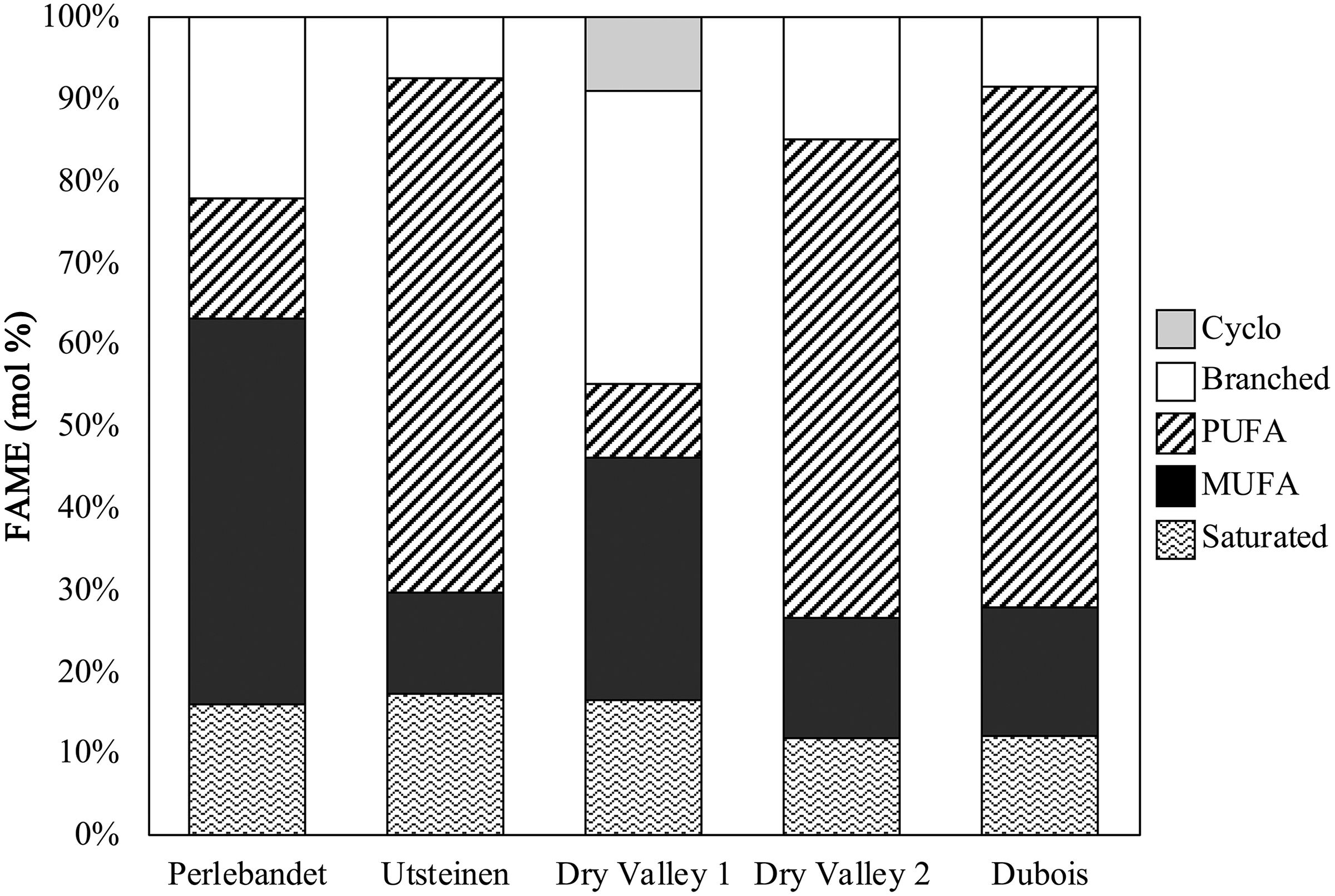
Distribution of different types of FAME in mol % for individual endolith samples. FAME, fatty acid methyl esters.
Monounsaturated fatty acids (MUFA) were the second most abundant type of FAME observed in every sample, mostly 18:1 and 16:1, and are generally considered to be indicative of gram-negative bacteria (Zelles, 1999; Boschker et al., 2005) or methanotrophs (Zelles, 1999; Boschker and Middleburg, 2002; Conrad, 2007). MUFA comprised 47% of the FAME for Perlebandet, 30% of the FAME for Dry Valley 1, 16% of the FAME for Dubois, 14% of the FAME for Dry Valley 2, and 12% of the FAME for Utsteinen. BCFA were nearly as abundant as MUFA and found in every sample, predominantly i-16:0, and are thought to represent gram-positive (Zelles, 1999; Green and Scow, 2000) and gram-negative bacteria (Zelles, 1999) in the microbial community. BCFA comprised 22% of the FAME at Perlebandet, 36% of the FAME at Dry Valley 1, 14% of the FAME at Dry Valley 2, 8% of the FAME at Dubois, and 7% of the FAME at Utsteinen.
SFA, mostly 16:0, are ubiquitous FAME and found in many different organisms, including cyanobacteria (Dijkman et al., 2010). SFA were less abundant than PUFA, MUFA, or branched and comprised 17% of the FAME at Utsteinen, 16% of the FAME at Dry Valley 1, 16% of the FAME at Perlebandet, 12% of the FAME at Dubois, and 11% of the FAME at Dry Valley 2. The least abundant category of FAME—cyclopropyl fatty acids (cyclo)—was only detectable in one sample and consisted predominantly of cyclo-17:0, which may be a stress biomarker (Sulzman, 2007) or indicate the presence of gram-negative bacteria (Zelles, 1999) within the microbial community. Cyclo represented about 9% of the FAME at Dry Valley 1.
The Δ14C values of the viable microbial membranes (as PLFA) ranged from −111‰ to 62‰ ± 20‰ (Table 1). The Δ14C of PLFA from Perlebandet was the most depleted in radiocarbon (−111‰ ± 20‰), followed by Dry Valley 1 (−14‰ ± 20‰). The corresponding ages for these two PLFA samples ranged from 944 to 114 years BP in uncalibrated 14C years—indicating the presence of aged carbon as a source within the endolithic system. The Δ14C of PLFA was enriched within the remaining three samples: Dry Valley 2 (31‰ ± 20‰), Dubois (33‰ ± 20‰), and Utsteinen (62‰ ± 20‰). These signals correspond to modern radiocarbon ages and the presence of bomb carbon, demonstrating some variability in carbon sources to Antarctic endoliths.
The δ13C values of PLFA ranged from −39‰ to −35‰ ± 1‰ (Table 1). The δ13CPLFA of endoliths from Dry Valley 1 was the most isotopically depleted (−39‰ ± 1‰), followed by Perlebandet (−38‰ ± 1‰), then Dry Valley 2 and Utsteinen (both −36‰ ± 1‰). The δ13CPLFA of endoliths from Dubois was the least isotopically depleted (−35‰ ± 1‰) among the samples analyzed.
4. Discussion
4.1. Endolithic microbes are primarily using recent atmospheric carbon
Radiocarbon analyses of lipids from the viable microbial community (as PLFA) show that, overall, East Antarctic endoliths are cycling carbon quickly. While it may be viewed that data presented here support the previous finding that West Antarctic endoliths cycle carbon quickly, we suggest that East Antarctic microbes are cycling carbon even more rapidly than those found in West Antarctica. The most recent Δ14C values of atmospheric CO2 for East Antarctica were 16‰ ± 3‰ in 2016 and 12‰ ± 3‰ in 2017 (I. Levin, pers. comm., March 25, 2019). Atmospheric 14CO2 is steadily declining over time at a rate of about 5‰ per year due to the combustion of radiocarbon-dead fossil fuel carbon and the uptake of bomb carbon by the biosphere and ocean. Three of the five endolith samples in this study appear to be incorporating carbon that was in equilibrium with the recent atmosphere. The amount of carbon in these endolithic systems (TOC %) is also quite different. The East Antarctic endoliths were more carbon rich (0.58% ± 0.34%) than the West Antarctic samples (0.19% ± 0.09%) in the work of Brady et al. (2018). Therefore, we put forward the hypothesis here that the less-southern East Antarctic nunatak endoliths cycle carbon more quickly than the more-southern West Antarctic Dry Valley microbes.
The Δ14CPLFA of endoliths from Dry Valley 2 (31‰ ± 20‰) and Dubois (33‰ ± 20‰) was consistent with recent atmospheric Δ14C values (Table 1). The simplest explanation for the modern carbon in the membrane lipids is that these microbial communities are cycling carbon over yearly or decadal timescales. One way this could occur is through autotrophic fixation of atmospheric carbon and heterotrophic consumption of modern bioavailable carbon. If this carbon had been fixed hundreds or thousands of years ago, it would have undergone radioactive decay and the resultant composition of the PLFA would be much more depleted in radiocarbon. Therefore, for two of the samples in this study, the most plausible explanation for the radiocarbon content of the viable membrane lipids is that these microbes are actively cycling carbon. However, because the history of atmospheric 14CO2 is complex, there could be another explanation for the “modern” 14C composition of the membrane lipids.
The radiocarbon composition of atmospheric CO2 can be altered by variability in cosmic ray flux (natural production) or through artificial production, such as high-energy weapons testing. Nuclear weapons testing in the late 1950s and in the early 1960s introduced an excess of artificial radiocarbon into the atmosphere, resulting in very enriched Δ14C values shortly thereafter (Hua and Barbetti, 2004). This “pulse” has been used as an isotopic tracer for ecosystem uptake of bomb carbon. Thus, bomb carbon can be used to determine the activity of microbes that are slowly growing over decades. If the Δ14C of the PLFA is enriched relative to today's atmosphere, it can be assumed that bomb carbon is present and that the microbes are likely cycling carbon on a decadal scale.
While the Δ14CPLFA of the endoliths from Dry Valley 2 and Dubois (31‰ ± 20‰ and 33‰ ± 20‰, respectively) was within error of the Δ14C of modern atmospheric CO2, it is also possible that these isotopic values could be due to the mixing of old carbon from very slowly cycling cells and bomb carbon taken up by microbes that cycle on decadal timescales. Similarly, the more enriched Utsteinen Δ14CPLFA (62‰ ± 20‰) indicates the incorporation of bomb carbon. The fact that these microbial lipids contain carbon that is within error of the recent atmosphere suggests that East Antarctic endoliths are cycling carbon on a decadal scale or less.
While most of the microbial PLFA contained carbon that was recently in equilibrium with the atmosphere, several PLFA samples contained older carbon. Perlebandet and Dry Valley 1 had the most depleted Δ14CPLFA values in the study (−110‰ ± 20‰ and −14‰ ± 20‰, respectively), indicating the presence of aged carbon consistent with ages of 945-115 BP in uncalibrated 14C years. There are several scenarios that could explain the aged carbon in the PLFA: (1) the endoliths are slowly growing on a 100-year scale (i.e., not very active), (2) heterogeneous distribution of microbial colonization and carbon sources within the rock itself, or (3) the endoliths are actively recycling older carbon in the system.
While previous studies have argued that these extremophilic Antarctic communities are likely slow growing in order survive in such a harsh environment (Friedmann, 1982; Johnston and Vestal, 1991), the predominantly modern Δ14CPLFA values observed in this study, coupled with recent work in the McMurdo Dry Valleys (Colesie et al., 2016; Zazovskaya et al., 2016; Brady et al., 2018), suggest that the scenario in which the microbes are growing on a 100-year scale may be unlikely. Therefore, another explanation for the aged carbon within the PLFA is needed.
Evidence of the second scenario, where heterogeneous distribution of microbial colonization and carbon sources within the rocks could result in aged 14C values, was observed as two samples from the same site yielded different results. Low concentrations preclude the ability to have multiple samples within the same rock, so—rather than sampling the same rock multiple times—two rocks were sampled within 1 km of each other in the Dry Valley. While viable membrane lipids from Dry Valley 2 contained modern carbon (Δ14CPLFA = 31‰ ± 20‰), Dry Valley 1 revealed the incorporation of older carbon (Δ14CPLFA = −14‰ ± 20‰). In addition, the FAME profiles differed between the two samples. For example, Dry Valley 2 had a higher abundance of PUFA (56%), whereas Dry Valley 1 had a notably lower abundance (9%)—inferring distinct microbial communities. The contrast between the 14C of these two samples could be due to a greater abundance of active microorganisms within the sampled region from one rock (Dry Valley 2) versus the sampled region from the other (Dry Valley 1).
As there were no noted differences in what the rocks looked like, another potential cause for the difference in both the 14C and FAME profiles is a recent mechanical loss of stabilized (i.e., preserved) organic carbon (both living and dead biomass) from one or both Dry Valley samples. The more modern and abundant PLFA in Dry Valley 2 also contained more TOC than Dry Valley 1. As endoliths colonize and continue to grow within the structural cavities of a rock, they biogenically weather away lithologic flakes (Friedmann, 1982)—likely resulting in the loss of stored carbon and portions of the microbial community from various areas of the rock. Therefore, the aged 14C value from Dry Valley 1 could be due to less weathering and the resultant accumulation of older carbon in the system.
Evidence of the third scenario, where East Antarctic endoliths could be actively recycling relict carbon from within the system, a variety of carbon sources to these microorganisms exist. Potential sources include CO2 from the atmosphere, CO2 within the rock itself, carbonate, and stabilized organic carbon composed of both living and dead biomass (Ziolkowski et al., 2013a). Endolithic communities within the McMurdo Dry Valleys were found to use significantly variable carbon sources, where the Δ14CPLFA values may reflect a mixture of more than one source (i.e., different isotopic compositions) (Brady et al., 2018).
The availability of carbon sources to endoliths is likely a function of location, where local environmental conditions can impact the stability of carbon within the rock. For example, a horizontally oriented rock is better for preserving endolithic organic matter as opposed to a vertical rock face, which is subjected to enhanced physical weathering due to gravity (Zazovskaya et al., 2016). As weathered lithologic fragments flake away, carbon stored within the rock is gradually lost and may result in heterogeneous distribution of both biomass and carbon sources within the subsurface. Although the rocks collected for this study were generally friable, it is still very possible that relict carbon remains within the East Antarctic endolithic systems and is being actively recycled. The Δ14CPLFA values that resemble modern atmosphere (Utsteinen, Dry Valley, and Dubois) may be a result of isotopically depleted (i.e., aged) carbon being mixed with isotopically enriched bomb carbon, indicating that endoliths are cycling carbon on a decadal, rather than yearly, scale.
The radiocarbon content of the viable microbial membranes from East Antarctic endoliths reported here appears to differ from those of West Antarctic endoliths (i.e., McMurdo Dry Valleys). In the McMurdo Valleys, the Δ14CPLFA of samples from University Valley (UV) had values that ranged from −199‰ to −74‰ ± 20‰, which correspond to uncalibrated 14C ages of 1780–660 years BP, whereas the Δ14CPLFA from Farnell Valley (FV) was modern (40‰ ± 20‰) (Brady et al., 2018). Even though the range of the observed radiocarbon values overlapped, only one of the West Antarctic samples was found to contain modern carbon, whereas three of the East Antarctic samples were found to contain modern carbon. Later in the Discussion section, we address what may be driving the modern age of the carbon in the PLFA in these two environments. Additionally, it seems noteworthy that both the average 14C of the TOC and PLFA for the two studies were ∼90‰ different from each other even though there was a lot of variation between the samples in both environments [e.g., Brady et al. (2018): Δ14CPLFA = −95‰ ± 102‰ and Δ14CTOC = −190‰ ± 58‰; this study: Δ14CPLFA = 0‰ ± 68‰ and Δ14CTOC = −93‰ ± 63‰].
The data presented here could indicate that East Antarctic endoliths are more active (i.e., cycling carbon faster) than West Antarctic endoliths as the average Δ14CPLFA for this study contain carbon that is up to ∼800 years younger than the West Antarctic work (Brady et al., 2018). One explanation for these differences in microbial activities may be a result of geographic location, where the East Antarctic study site is positioned at a lower latitude than the McMurdo Dry Valleys. Given the proximity of the Dry Valleys to the south pole, it is possible that endoliths in this region are exposed to greater seasonal extremes in the form of longer austral winters and shorter austral summers. Therefore, these results suggest that East Antarctic endoliths may be growing under more amenable conditions than those in West Antarctica and are more active as a result.
4.2. TOC variability
The TOC—composed of both living and dead biomass—in East Antarctic endolithic systems is generally more depleted than the carbon within the viable microbial membranes (Fig. 4). Previous work has shown that the Δ14CTOC of endolithic systems is less depleted near sea level (at the coast), where temperatures are warmer and water is more readily available (Colesie et al., 2016; Zazovskaya et al., 2016). Conversely, previous measurements of Δ14CTOC in the Antarctic Dry Valleys, which are even further south than the Sør Rondane Mountains, were even more depleted (Brady et al., 2018). From this study, the Δ14CTOC at Utsteinen (−4‰ ± 5‰), Dry Valley 1 (−164‰ ± 5‰), Dry Valley 2 (−70‰ ± 5‰), and Dubois (−138‰ ± 5‰) was significantly more negative than the corresponding Δ14CPLFA, which ranged from −14‰ to 62‰ ± 20‰. Overall, the depleted TOC relative to PLFA suggests that East Antarctic microbes are incorporating carbon that is younger than the stabilized organic carbon within the endolithic system.
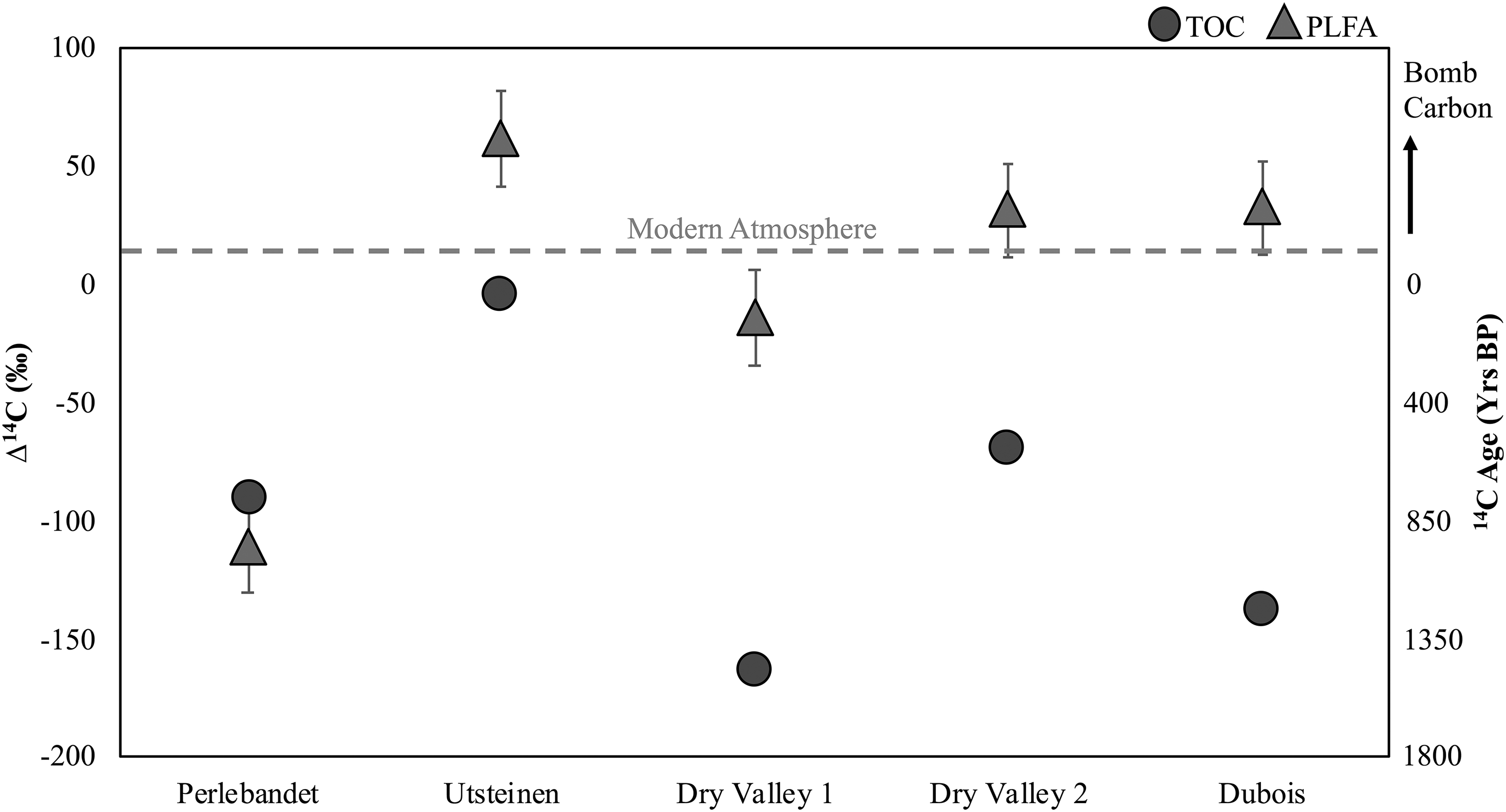
Δ14C ‰ values for the TOC (circles) and PLFA (triangles) of endolithic communities in Dronning Maud Land, East Antarctica. The dashed line represents modern atmospheric CO2 (about 40‰). Error bars are displayed for all points, where TOC errors are too small to be visible. This shows that Utsteinen has the youngest Δ14C age for PLFA. 14C, radiocarbon; PLFA, phospholipid-derived fatty acids; TOC, total organic carbon.
However, one of the PLFA samples contained older carbon than the TOC. For the sample from Perlebandet, the Δ14CTOC (−90‰ ± 5‰) was less depleted than the Δ14CPLFA (−111‰ ± 20‰). While these values are within error of each other, having Δ14CPLFA values that are more depleted than Δ14CTOC is atypical for microbes that are likely using atmospheric carbon. It is possible that once-modern PLFA could have been preserved within the endolithic system, as was thought to be the case for McMurdo Valley endoliths (Brady et al., 2018). It was also hypothesized by Brady et al. (2018) that part of the microbial community could be preserved, there could be large-scale community death, and extended periods of inactivity due to slow or recolonization that could lead to incorporation of 14C-depleted carbon. At this time, it is not clear which of these processes is occurring in these endoliths and warrants further investigation.
4.3. Microbial community composition
There appear to be strong correlations between microbial community composition and Δ14CPLFA. In general, PLFA structures differ between different life-forms. However, some PLFA are ubiquitous and can indicate a suite of organisms. Therefore, identifying the different PLFA structures within an endolithic sample is helpful in determining a broad brushstroke of microbial community composition. To explore the relationship between community makeup and Δ14CPLFA, radiocarbon signals were plotted against different types of PLFA (Fig. 5).

The correlation between PLFA abundance (in mol %) and the Δ14C of the viable microbial membrane (in ‰; error ±20‰) for saturated fatty acids, PUFA, MUFA, and branched fatty acids. MUFA, monounsaturated fatty acids; PUFA, polyunsaturated fatty acids.
There was a substantial statistical relationship between the radiocarbon content of the PLFA and the type of PLFA present, thus indicating that microbial community composition plays a role in how quickly microbes are cycling carbon (Fig. 5). In this study, the endolith samples with the most modern Δ14CPLFA (Utsteinen, Dry Valley 2, and Dubois) have a greater abundance of PUFA (>50%). Conversely, the endolith samples with the least modern Δ14CPLFA (Perlebandet and Dry Valley 1) have a higher abundance of MUFA (47% and 30%, respectively) and branched PLFA (22% and 36%, respectively) (Fig. 3). In addition, the samples with younger Δ14CPLFA had higher concentrations of PLFA (2.8–6.9 μg/g) compared with the samples with older Δ14CPLFA (0.9–1.3 μg/g) (Table 2). When plotted, the relationship between higher PUFA content and younger Δ14CPLFA values appears statistically robust (R 2 = 0.91) as is the relationship between higher MUFA content and older Δ14CPLFA values (R 2 = 0.93). While not as strong as the PUFA and MUFA statistical relationships, a moderate negative correlation (R 2 = 0.64) still exists between the relative abundance of branched fatty acids and Δ14CPLFA. From these trends, greater abundances of PUFA suggest microbial communities dominated by photoautotrophs.
In comparison, while the West Antarctic Dry Valley endolith sample from the work of Brady et al. (2018) with modern Δ14CPLFA (FV) had the highest concentration of PLFA (5.6 μg/g) and a greater abundance of PUFA (>50%), the overall relationship between Δ14CPLFA and PLFA composition is not as clear. For example, the sample with the most depleted Δ14CPLFA (UV3) had a lower abundance of MUFA and higher abundance of PUFA compared with the relatively younger UV2 and UV4 samples (Brady et al., 2018). This suggests that microbial community composition may play a role in how quickly the carbon is cycled in these systems.
4.4. Viable cell abundances within endolithic systems
A strong correlation (R 2 = 0.72) exists between PLFA-derived cell abundances and Δ14CPLFA, where larger cell abundances correlate with younger carbon within the microbial membrane (Fig. 6). Similar to Δ14CTOC values, biomass appears to vary as a function of coastal proximity and elevation, where larger biomasses (composed of living and dead cells) correlate with higher elevations and increased distance from the coast (e.g., Colesie et al., 2016). Calculated cell abundances were 107–108 cells/g of sample, which are comparable to recent estimates from the McMurdo Valleys (Brady et al., 2018). Utsteinen had the highest cell abundance (5.0 × 108 cells/g), followed by Dubois (2.6 × 108 cells/g) and Dry Valley 2 (2.0 × 108) (Table 2). Dry Valley 1 (9.7 × 107 cells/g) and Perlebandet (6.5 × 107 cells/g) had the lowest overall cell abundances in the study. If cell abundances are considered representative of living biomass, the endolithic communities that colonized the Utsteinen, Dubois, and Dry Valley samples are more bountiful than those that colonized the Dry Valley 1 and Perlebandet samples.

Plot showing the strong correlation between Δ14C (in ‰; error ±20‰) and PLFA-derived cell abundances (in cells/g).
5. Conclusions
Antarctic endoliths were previously thought to be some of the slowest growing (e.g., geological timescales) microbial communities on Earth due to climatic extremes, including desiccation, katabatic winds, and freezing temperatures. However, Brady et al. (2018) used natural abundance radiocarbon analysis of biomarkers and found that West Antarctic endoliths were likely cycling carbon on centurial to decadal scales instead of millennial scales, but very little was known about extremophilic communities residing in East Antarctica. By using natural abundance radiocarbon analysis of the viable microbial membrane (as PLFA), this present study illustrates that endoliths from the Dronning Maud Land region of East Antarctica are cycling carbon on decadal scales or less, which is much faster than previous estimates of endolithic microbial activity.
While these data support the premise that Antarctic endoliths are rapidly cycling carbon, the East Antarctic endoliths found on nunataks in this study may be cycling carbon faster than the West Antarctic endoliths found in the Dry Valleys. While the total amount of carbon in this study was higher than those from the McMurdo Dry Valleys, the cellular abundances in this study were comparable to those from the McMurdo Dry Valleys and had a robust statistical relationship to radiocarbon age. These findings suggest that endolith communities with larger biomass are likely more active. Certain FAME distributions also had a strong relationship to radiocarbon age—indicating a greater abundance of photosynthetic microorganisms in PLFA samples containing younger carbon. This work demonstrates that Antarctic endoliths are active on a continental scale, which, in turn, possesses some astrobiological implications. If life ever existed on Mars (or other planets), then evidence of such may be found in one of the last refugium available—inside the rocks.
Footnotes
Acknowledgments
We would like to thank Dr. Liane Benning (GFZ Potsdam), Dr. Stefanie Lutz (GFZ Potsdam), and Dr. Jenine McCutcheon (U. Waterloo) for their assistance in collecting samples and complementary forthcoming analyses. Finally, thank you to the anonymous reviewers and Editor-in-Chief for providing constructive comments that helped improve this article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by funding from the Baillet Latour International Polar Foundation awarded to L.A.Z.