
Abstract
Hydrothermal spring deposits host unique microbial ecosystems and have the capacity to preserve microbial communities as biosignatures within siliceous sinter layers. This quality makes terrestrial hot springs appealing natural laboratories to study the preservation of both organic and morphologic biosignatures. The discovery of hydrothermal deposits on Mars has called attention to these hot springs as Mars-analog environments, driving forward the study of biosignature preservation in these settings to help prepare future missions targeting the recovery of biosignatures from martian hot-spring deposits. This study quantifies the fatty acid load in three Icelandic hot-spring deposits ranging from modern and inactive to relict. Samples were collected from both the surface and 2–18 cm in depth to approximate the drilling capabilities of current and upcoming Mars rovers. To determine the preservation potential of organics in siliceous sinter deposits, fatty acid analyses were performed with pyrolysis–gas chromatography–mass spectrometry (GC-MS) utilizing thermochemolysis with tetramethylammonium hydroxide (TMAH). This technique is available on both current and upcoming Mars rovers. Results reveal that fatty acids are often degraded in the subsurface relative to surface samples but are preserved and detectable with the TMAH pyrolysis-GC-MS method. Hot-spring mid-to-distal aprons are often the best texturally and geomorphically definable feature in older, degraded terrestrial sinter systems and are therefore most readily detectable on Mars from orbital images. These findings have implications for the detection of organics in martian hydrothermal systems as they suggest that organics might be detectable on Mars in relatively recent hot-spring deposits, but preservation likely deteriorates over geological timescales. Rovers with thermochemolysis pyrolysis-GC-MS instrumentation may be able to detect fatty acids in hot-spring deposits if the organics are relatively young; therefore, martian landing site and sample selection are of paramount importance in the search for organics on Mars.
1. Introduction
Siliceous sinters are terrestrial geothermal spring deposits often formed in volcanic settings from silica-supersaturated fluids heated to high temperature by magmatic reservoirs. These fluids rapidly ascend to the surface where the water discharges at geysers and hot-spring vents, precipitating silica as the pressure and temperature of the fluids rapidly decrease (Renaut and Jones, 2011; Campbell et al., 2015). The precipitating silica can nucleate and coat all surfaces in the spring discharge flow path, resulting in unique mineralogic textures that are dependent on temperature and fluid composition when the silica precipitates.
Microorganisms that thrive in these extreme temperature, flow rate, and fluid compositions include bacteria, archaea, phages, and eukaryotes (e.g., diatoms) (Skirnisdottir et al., 2000; Breitbart et al., 2004; Leira et al., 2017; Saini et al., 2017; Pumas et al., 2018), and the cells of these organisms can also be entombed by silica precipitation, preserving both the physical structure of the cell and cellular organic matter (Cady and Farmer, 1996). These domains of life inhabit specific microenvironments at the hot springs conducive to the organisms' ideal growth conditions. Hyperthermophiles and thermophiles are found near the higher temperature spring vent, mesophilic organisms such as cyanobacteria inhabit fluids <73°C between the vent, proximal slope, mid-apron, and distal apron, and plants and other mesophiles inhabit the lower temperature environments (commonly proximal slope, mid-apron, or distal apron) (Fig. 1).
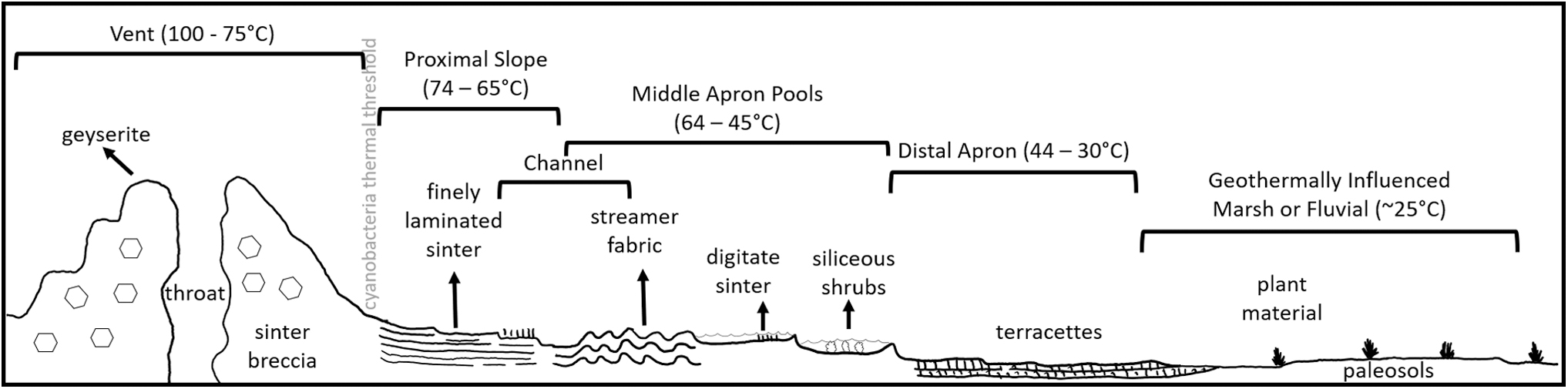
Environmental settings and facies of a generalized geothermal spring system. Modified from Campbell et al. (2015).
The preservation of organic biosignatures in geothermal spring deposits is of particular interest to the astrobiology community, as several siliceous hot-spring deposits have been identified on Mars with both remote sensing and in situ exploration. Hydrothermally driven siliceous sinter deposits have been identified on Mars from orbital observations in Nili Patera (Skok et al., 2010) and Arabia Terra (Allen and Oehler, 2008; Rossi et al., 2008). Hot-spring opal-A deposits with nodular and digitate stromatolitic morphologies have been identified in Columbia Hills, Gusev crater, by the Spirit rover (Squyres et al., 2008; Ruff et al., 2011), which are strikingly similar to digitate stromatolites with putative biogenic origin found at the El Tatio geothermal field in Chile (Ruff and Farmer, 2016). Although not geothermal in nature, diagenetic silica enrichment is also observed in Marias Pass, Gale crater, by the Curiosity rover, which implies groundwater circulation in Gale crater long after lacustrine activity ceased (Frydenvang et al., 2017).
The preservation of physical cells and cellular organic matter in hot springs both constitutes biosignatures. Biosignatures are defined as an object, pattern, or substance whose origin specifically requires a biological agent (Hays et al., 2017). Extensive work has been performed, which documents the extraordinary preservation of microbial cell structure in sinter deposits by entombment in a very fine matrix of silica particles (Walter and Des Marais, 1993; Cady and Farmer, 1996; Walter, 1996; Renaut et al., 1998; Jones et al., 2000; Konhauser et al., 2001; Tobler et al., 2008; Guido et al., 2010; Brasier et al., 2011; Campbell et al., 2015; Westall et al., 2015; McCollom et al., 2016; Munoz-Saez et al., 2016; Tarhan et al., 2016). Few studies, however, have focused on the preservation of organic matter in modern and relict (up to 900 year old) siliceous sinter deposits (Pancost et al., 2005, 2006; Kaur et al., 2008, 2011, 2015; Gibson et al., 2014; Chaves Torres et al., 2019; Teece et al., 2019), and even fewer studies have explored organic preservation potential in Mars-analog hot-spring environments (e.g., Williams et al., 2019).
1.1. Martian in situ organics detection
The in situ search for organic molecules on Mars has been a priority of the scientific community for decades, and several missions have carried payloads with varying organics detection capabilities. The approaches that have been deployed and/or planned to detect organics on Mars missions include Raman spectroscopy (Hays et al., 2017 and references therein), deep ultraviolet laser excitation (Hays et al., 2015 and references therein), and gas chromatography–mass spectrometry (GC-MS). GC-MS has the highest heritage, having flown in modified forms on the Mars Viking landers (Biemann et al., 1977), the Mars Phoenix polar lander (Boynton et al., 2001), and the Sample Analysis at Mars (SAM) instrument on the Mars Curiosity rover (Mahaffy et al., 2012). A GC-MS is also included as part of the Mars Organic Molecule Analyzer (MOMA) instrument on the upcoming European Space Agency's Rosalind Franklin (ExoMars) rover (Goetz et al., 2016; Goesmann et al., 2017).
The fundamentals of the GC-MS system for all Mars flight instruments are essentially the same: the goal of the approach is to detect and identify various molecular species and classes at low concentrations with high analytic specificity. Volatile compounds thermally evolved from solid samples in an oven may be directly heated (pyrolyzed) or subjected to a derivatization procedure. The evolved species are separated by the GC into a time sequence, then separated by mass and analyzed with the MS. Although pyrolysis-GC-MS has the potential to liberate organics bound within mineral matrices, the detection of those organics can be compromised by the presence of perchlorate or other oxidizing salts. Perchlorate is present on the martian surface and is known to combust organics during pyrolysis due to the evolution of O2 from the perchlorate, effectively destroying the organic molecule in the process (Guzman et al., 2018). However, certain derivatization or thermochemolysis reactions can liberate and volatilize select organics in the presence of perchlorates without the organics combusting.
A derivatization or thermochemolysis reaction—referred to here, as in the literature, as a “wet chemistry experiment”—is a “one-step” reaction with a reagent that improves organic matter yield and liberates organics bound within macromolecules. Wet chemistry experiments aim to transform polar organic molecules (e.g., carboxylic acids) into volatile derivatives that are amenable and detectable by GC-MS (Metcalffe and Wang, 1981; del Rio et al., 1996), as well as freeing bound components of larger macromolecules otherwise undetectable with GC-MS (Grasset et al., 2002).
Different wet chemistry reagents can be used based on the chemistry of the targeted labile group and the required yield of the reaction, such as N,N-methyltert-butyl-dimethylsilyltrifluoroacetamide (MTBSTFA; Mahaffy et al., 2012; Goesmann et al., 2017), N,N-dimethylformamide dimethyl acetal (Goesmann et al., 2017), trimethylsulfonium hydroxide (Ishida et al., 1999), and tetramethylammonium hydroxide (TMAH; Grasset et al., 2002; Mahaffy et al., 2012), to name just a few. Thermochemolysis with TMAH in methanol (MeOH, a solvent) allows access to refractory or insoluble organic material, including macromolecular organics such as kerogen-like compounds. Simply pyrolyzing a sample at high temperature (>650°C) results in fragmentation of the parent molecule due to thermal stress, which limits molecule identification. To cleave these refractory molecules at a milder temperature (e.g., ≤600°C), thermochemolysis with TMAH allows the analysis of refractory matter in complex matrices containing low amounts of organic material with minimal destruction of the organic material (Geffroy-Rodier et al., 2009; He et al., 2019; Williams et al., 2019).
TMAH thermochemolysis selectively cleaves ester and ether bonds, replacing a labile hydrogen atom with a methyl group (Fig. 2), rather than random and uncontrolled thermal decomposition and fragmentation of the organic material (as during pyrolysis). TMAH has been used for several decades in a variety of terrestrial fields of study, including petroleum geochemistry (Larter and Horsfield, 1993), soil science (Schulten, 1996; Chefetz et al., 2000; Deport et al., 2006), and sedimentology (Pulchan et al., 1997; Guignard et al., 2005). TMAH is currently available on the Curiosity rover SAM instrument (Mahaffy et al., 2012; Williams et al., 2019) and will also be used on the ExoMars rover MOMA instrument (Goesmann et al., 2017). Fortunately, perchlorate does not substantially affect the recovery of fatty acids liberated with TMAH thermochemolysis (He et al., 2019), making TMAH an ideal reagent to use in the search for select organics on Mars.

Example TMAH thermochemolysis reaction with a carboxylic acid to form a methylated carboxylic acid. After the reaction, the molecule is more volatile and detectable to gas chromatography–mass spectrometry. TMAH in methanol is available on the SAM and Mars Organic Molecule Analyzer instruments. TMAH, tetramethylammonium hydroxide; SAM, Sample Analysis at Mars. Color images are available online.
Although the SAM instrument TMAH experiment on the Curiosity rover has yet to be performed (as of the time of this writing), several other lines of evidence already indicate that organics are present on the surface and near-surface (<6 cm) of Mars. Chlorinated hydrocarbons such as chlorobenzene and C2 to C4 dichloroalkanes were detected in the Sheepbed mudstone in Yellowknife Bay, Gale crater (Freissinet et al., 2015), and thiophenic, aromatic, and aliphatic organic compounds were found in the 3.5 Gy Murray Formation lacustrine mudstones of Pahrump Hills, Gale crater (Eigenbrode et al., 2018). In more recent SAM experiments, preliminary analyses suggest the presence of select medium-chain alkanes (Freissinet et al., 2019) and medium-to-high molecular masses, including derivatized molecules (Millan et al., 2019), in samples exposed to SAM's other wet chemistry experiment, derivatization with MTBSTFA (Buch et al., 2006).
These findings represent the first confirmed in situ organics detection on Mars, but the source of these organic molecules is uncertain. Abiotic martian sources (e.g., from atmospheric, hydrothermal, or igneous processes), abiotic exogenous sources such as comets, interplanetary dust particles, or meteorites (Freissinet et al., 2015; Eigenbrode et al., 2018), and life all produce organic molecules. TMAH thermochemolysis, however, provides an unprecedented opportunity to assess the biogenicity of martian organics, specifically carboxylic and fatty acids. Fatty acids bound to phospholipids and glycolipids are abundant and ubiquitous components of eukaryotic and bacterial cellular membranes (Vestal and White, 1989). Carboxylic acids have also been identified in exogenous carbonaceous material, such as in the Murchison meteorite (Cronin et al., 1993).
Biotic and abiotic-generated carboxylic acids produce distinctly different profiles in GC-MS analyses. Carboxylic and fatty acids derived from metabolic processes can range from C2 to >C30 in length with an even-over-odd carbon chain length preference due to enzymatically formed acetyl (C2) units derived from glucose (Volkman, 2006). Abiotic fatty acid patterns will favor shorter carbon chain lengths with no carbon preference (Bray and Evans, 1961; McCollom et al., 1999), although it is important to note that very short-chain carboxylic acids can represent metabolic by-products that were not incorporated into cellular membranes. Therefore, within a certain level of uncertainty, characteristics of a fatty acid methyl ester (FAME) profile could be used to deconvolve the origin of fatty acids detected on Mars (Williams et al., 2019).
The work presented here investigates the preservation and detection of organic molecules, specifically fatty acids, in modern inactive and relict Icelandic hydrothermal spring systems that precipitate siliceous sinter and travertine. Organics preservation in these Mars-analog hot-spring environments is compared between mostly inactive and relict spring deposits, and includes samples from spring vent, mid-apron, and distal apron facies, and within a depth range comparable with the sampling depth of the Curiosity and future rover drill systems. We present data to assess the limits of space-flight-like analyses and how returned in situ data from extraterrestrial hot springs may be interpreted.
2. Methods
2.1. Sample locations
Samples were collected at three Icelandic hydrothermal sinter sites, at the surface and subsurface ranging from 2 to 18 cm in depth from spring vent to distal apron, to explore the coupling of system activity with potential preservation of biosignatures. In hydrothermal hot-spring systems, there are three main regions: near-vent, the location of higher temperature fluid discharge from the subsurface; the middle-apron, a moderate temperature region often with pooling water; and the distal apron, areas of cooler fluid further from the vent (e.g., Fig. 1).
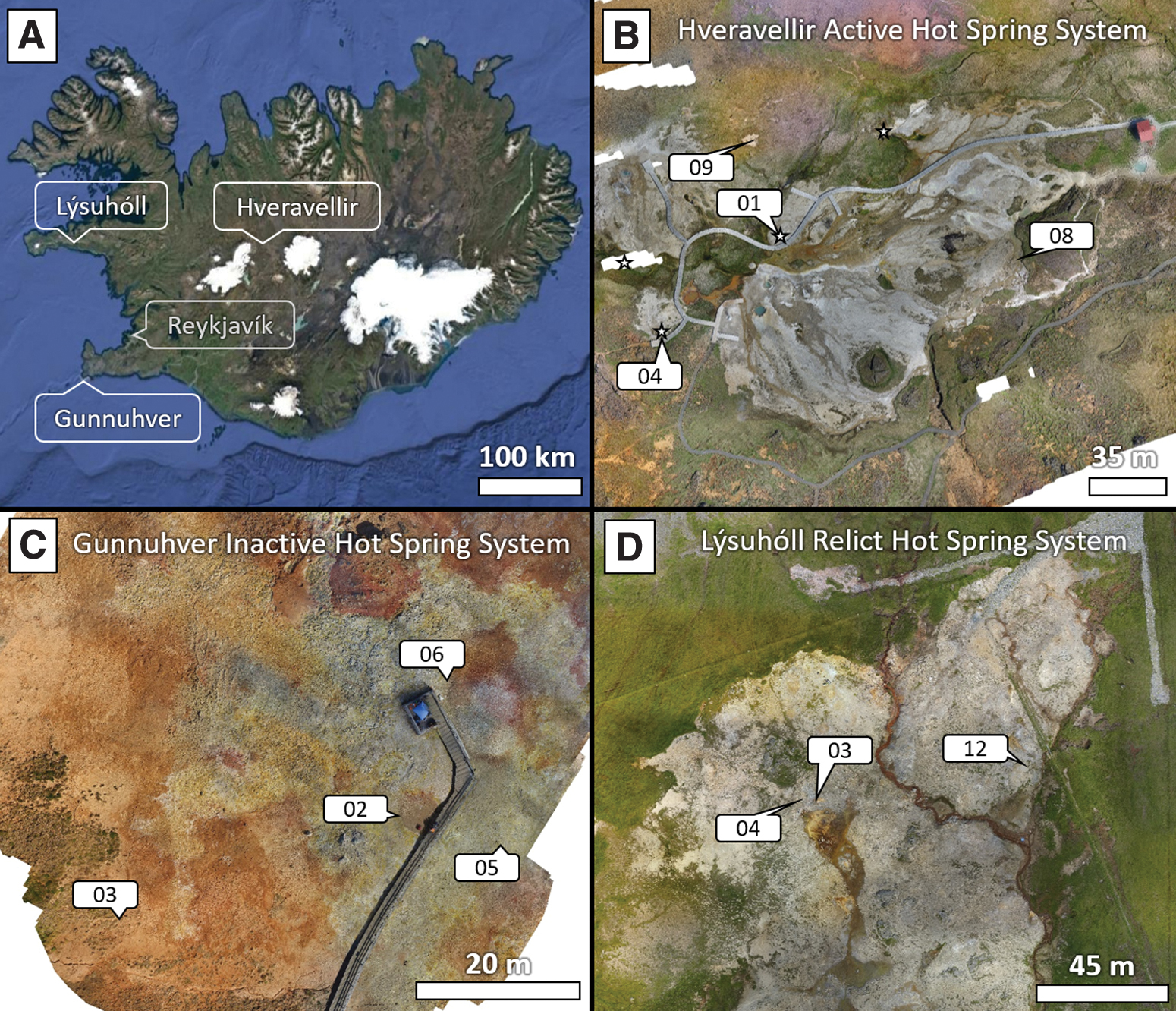
A currently active region, Hveravellir, sits in the central highlands area of Iceland (Fig. 3B), and contains many hot springs and fumaroles. This area is protected as a monument in Iceland, with boardwalks and restricted sites that preserve the formations and minimize human alteration. At Hveravellir, we collected sample 160730.01 (mid-apron) near an active spring vent, but sampling site was only moist and not within the main flow region (Fig. 4). We also sampled at a recently inactive vent (160730.09) and the distal apron (160730.08) of an inactive vent. The pisolith sample (160730.04) had formed in pools at an actively flowing vent. Given the variety of samples from active and recently inactive sites, the Hveravellir system will be referred to as “inactive” in this work. The geothermally active Hveravellir region has been geochemically characterized, with the spring vents in the area generally emerging at ca. 90°C, with a pH of 8.60 and [SiO2] of 599 ppm (Villaroel Camacho, 2017). The actively flowing spring vents proximal to Hveravellir vent sample “160730.09” ranged from 91.0°C to 97.1°C, pH 8.15–8.65, [SiO2] of 321–641 ppm, and [Cl] of 53.1–63.6 ppm. Hveravellir spring waters are considered part of a population of high-temperature geothermal waters, along with Geysir, Hveragerdi, and Reykjadalir, which contain elevated silica concentrations relative to the lower temperature geothermal regions on the island.

Field sample site images and corresponding scanning electron microscope images of sinter from
A second site, Gunnuhver, lies on the Reykjanes peninsula in SW Iceland (Fig. 3C). The sampling site is recently inactive (no spring activity), but with a large active fumarole ∼10 m away and geothermal heat still warming the surface. A downstream (∼50 m) channel from the active vent in the Gunnuhver region was 24.1°C with a pH = 7.34. Samples collected at Gunnuhver included those near-vents, mid- (15–23 m), and distal-apron (33–49 m) locations from deduced vent locations (Fig. 4).
The third site visited was Lýsuhóll, a farmland area with relict sinter deposits and few active vent systems (Fig. 3D) with lower temperatures (ca. 41°C) and pH = 6.3. Other low-temperature (ca. 50°C) thermal springs are present nearby in the Snӕfellsnes region (Villaroel Camacho, 2017). Silica sinter (e.g., Opal-A) deposits constitute the mineralogy of older regions in this deposit, overprinted by younger travertine deposits (Jones et al., 2005; Jones and Renaut, 2017). Vent and near-vent samples were collected at Lýsuhóll. The determination of vent locations was more difficult at this relict site as erosion had disrupted many of the deposits; therefore only three well-preserved sites were sampled (Fig. 4).
All samples were collected with the appropriate permissions. Hveravellir is a protected natural monument for which we were permitted to sample by the Environment Agency of Iceland. We were permitted to sample the Gunnuhver site by the National Energy Authority. Lýsuhóll is private land for which we had owner permission to sample. Finally, we also had permission from the Icelandic Institute of Natural History to transport samples out of Iceland.
2.2. Sampling procedure
To minimize contamination, all sinter samples were collected with solvent-washed and ashed tools, while wearing gloves, facemasks, and hairnets. Samples were collected with solvent cleaned (MeOH/DCM) and ashed (at 500°C) chisels, spoons, and tweezers. Rock hammers were covered in a new piece of ashed (at 500°C) foil before use. Samples were placed in solvent-washed and ashed (at 500°C) glass jars and stored on ice for transport back to the laboratory. Samples were collected both from the surface (denoted with an “S”) and from several centimeters into the subsurface (denoted with an “I”). Fresh solvent-washed tools were used for each surface and subsurface sample to limit crosscontamination between sample depths. Samples were transported to the laboratory on ice and stored in a −20°C freezer upon arrival. Samples for scanning electron microscope (SEM) analysis were either collected in the field in sterile whirlpak bags with solvent-washed tools, or subsampled in the laboratory from the solvent-washed and ashed glass jars with solvent-washed tools.
In the laboratory, sinter from the glass jars was subsampled by breaking off smaller pieces with solvent-washed and ashed tweezers. These subsamples were powdered with a solvent-washed and ashed ceramic mortar and pestle, and ground for 3 min to a homogeneous powder. As samples were carefully collected with organically clean tools and techniques in the field, samples were not treated to remove potential natural external contamination before powdering. This follows the approach of Wilhelm et al. (2017).
2.3. Imaging and mineralogy
The aerial drone maps for Fig. 3 were acquired by DJI Mavic Pro with the onboard camera at 12 MP resolution. Images were recorded at 2 s intervals to ensure that most surface points would be captured in >5 images. The images were stitched together by using the commercially available Pix4D Mapping Software to combine the images into the mosaic. Due to the size of the field sites, map images were acquired at different flight altitudes resulting in the following pixel resolutions: Gunnuhver (5 m, 0.32 cm/pixel), Hveravellir (33 m, 1.41 cm/pixel), and Lýsuhóll (60 m, 2.76 cm/pixel).
SEM images of sinter were collected with a Phenom ProX G5 tabletop environmental SEM. Qualitative analyses were conducted by using a 3 μm spot size, a beam current of 2.25–2.33 nA, and an accelerating voltage of 15 kV under low vacuum conditions consistent with environment SEM operation.
Mineralogy was determined through X-ray diffraction with either a Bruker D8 Discover X-Ray Diffractometer (Cu Kα radiation, λ = 1.54059 Å); patterns were acquired from 2° to 70° 2θ, or an Olympus Terra X-Ray Diffractometer (Co Kα radiation, λ = 1.78897 Å); patterns were acquired from 5° to 55° 2θ.
2.4. Pyrolysis-GC-MS conditions for analog samples
Aliquots of each powdered sample underwent pyrolysis (with a commercial Frontier pyrolyzer 3030D) GC-MS analysis to measure the distribution and abundance of FAMEs. Ground rock or sediment samples were weighed into nonreactive metal cups (sample mass was ca. 5–10 mg, depending on the sample). Just before sample analysis, TMAH in the ratio of 1 mg sample to 1 μL TMAH was added to the cup. The n-C19:0 internal standard was injected into each sample immediately before analysis to determine the efficiency of FAME transfer. The sample was then immediately loaded and dropped into the pyrolyzer oven to begin the run. The GC-MS analysis was run on either an Agilent 7890A-5975C inertXL GC-MS or a Thermo Trace 1310 GC-MS equipped with a 30 m Restek capillary column (MXT-5) with a 0.25 mm internal diameter and 0.25 μm thick Crossbond® 5% diphenyl/95% dimethyl polysiloxane stationary phase, and He as the carrier gas at a 3 mL/min flow rate and 10:1 split. Blank (without TMAH) and “TMAH-blank” (with TMAH) cleanup analyses were run between each sample to determine and subtract any background level of residual FAMEs in the column.
Chromatograms and the mass spectra for FAMEs were analyzed with ChemStation software (Agilent Technologies). Identifications were based on comparison with known FAME retention times of a Supelco 37 component FAME mixture that contains saturated FAMEs, monounsaturated fatty acid (MUFA) methyl esters, and polyunsaturated fatty acid (PUFA) methyl esters (Sigma-Aldrich). In addition, mass spectra were compared with the National Institute of Standards and Technology Spectral Library. Quantifications were based on integration under peaks and scaled to the known amount of the n-C19:0 internal standard.
The pyrolyzer and GC programs used are detailed below: For the SAM-like pyrolysis ramp and GC column method, the pyrolyzer program started at 50°C and was ramped at 35°C/min to 400°C with no hold. The inlet program and Tenax® TA trap started at 15°C with a 2 min hold. The inlet was ramped at 68°C/min to 150°C and held for 2 min, then ramped at 80°C/min to 305°C and held for 3 min. The GC column program started at 35°C and was held for 10.9 min, then ramped at 15°C/min to 175°C with no hold, then ramped at 10°C/min to 305°C and held for 1 min. The auxiliary transfer line was held at 135°C.
For the 500°C flash pyrolysis method, the sample was dropped into a 500°C pyrolyzer oven and held for 1 min. The inlet program and Tenax TA trap started at 15°C with a 2 min hold. The inlet was ramped at 900°C/min to 300°C and held for 2 min. The GC column program started at 35°C and was held for 2.5 min, then ramped at 5°C/min to 300°C and held for 5 min. The auxiliary transfer line was held at 270°C.
3. Results
3.1. Mineralogy
The mineralogy of the three sample locations was generally dominated by opal-A, opal-CT, and amorphous silica (Table 1). The modern inactive hot-spring vent at Hveravellir was dominantly opal-A with lesser percentages of clinopyroxene, plagioclase, gypsum, and magnesite. Several Hveravellir locations were composed entirely of amorphous silica. The modern inactive hot spring at Gunnuhver was predominantly opal-CT, with the distal apron being dominantly amorphous silica. This system also contained lesser percentages of halite and quartz. The older relict hot spring at Lýsuhóll is composed almost exclusively of amorphous silica with some minor calcite, and one sample site was composed entirely of calcite.
Sample Location, ID, Depth, Mineralogy, Latitude, Longitude, Percentage Total Carbon, and Percentage Total Organic Carbon

TC, total carbon; TOC, total organic carbon.
3.2. FAMEs detected with 500° C instantaneous flash pyrolysis
Samples were initially analyzed by using a 500°C flash pyrolysis technique to enable transesterification of fatty acids with TMAH thermochemolysis. By using this technique, there was relatively high diversity and abundance of FAMEs in the Hveravellir spring site, a lower diversity and abundance of FAMEs at the inactive Gunnuhver spring site, and diminished diversity and abundance of FAMEs at the Lýsuhóll relict site (Table 2).
Flash pyrolysis FAME Data in Nanograms FAME/Milligrams TOC
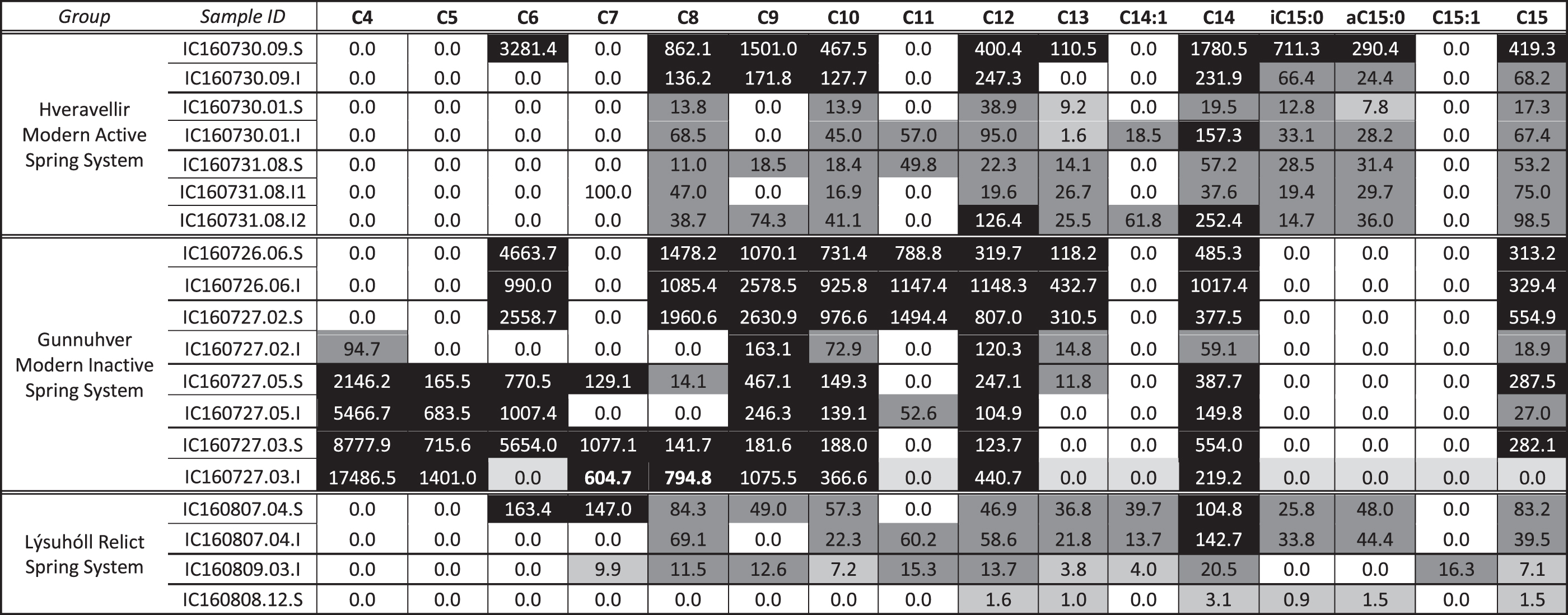
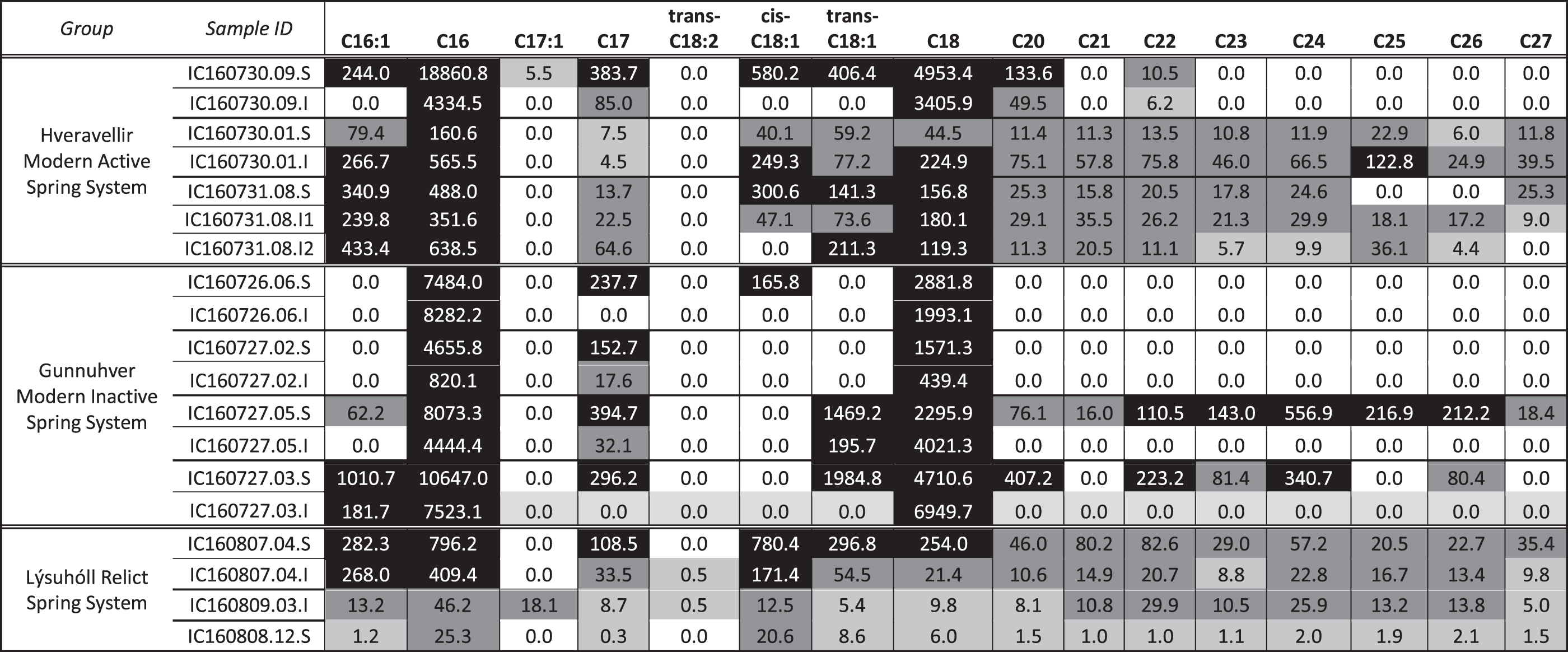
Black cells = concentrations >100 ng/mg TOC. Dark gray cells = concentrations 10–100 ng/mg TOC. Light gray cells = 0.01–10 ng/mg TOC. White cells = no detection (0.0 ng/mg TOC).
FAME, fatty acid methyl ester; TOC, total organic carbon.
At the Hveravellir site, samples were collected from the spring vent, mid-apron, and distal apron, as well as a pisolith-bearing layer near an active vent. FAMEs detected ranged from n-C6:0 to n-C27:0, and included a few MUFAs and branched iso- and antiso-C15:0 (Fig. 5; Table 2). n-C16:0 and n-C18:0 were the most abundant FAMEs in all samples, and FAMEs longer than n-C18:0 were identified at all sites except the vent. In general, the abundance and diversity of FAMEs were increased in the surface samples relative to the subsurface samples, which ranged from 3 to 18 cm in depth. The distal-apron sample demonstrated an unexpected general trend of greater FAME concentrations in the interior samples than the surface samples.
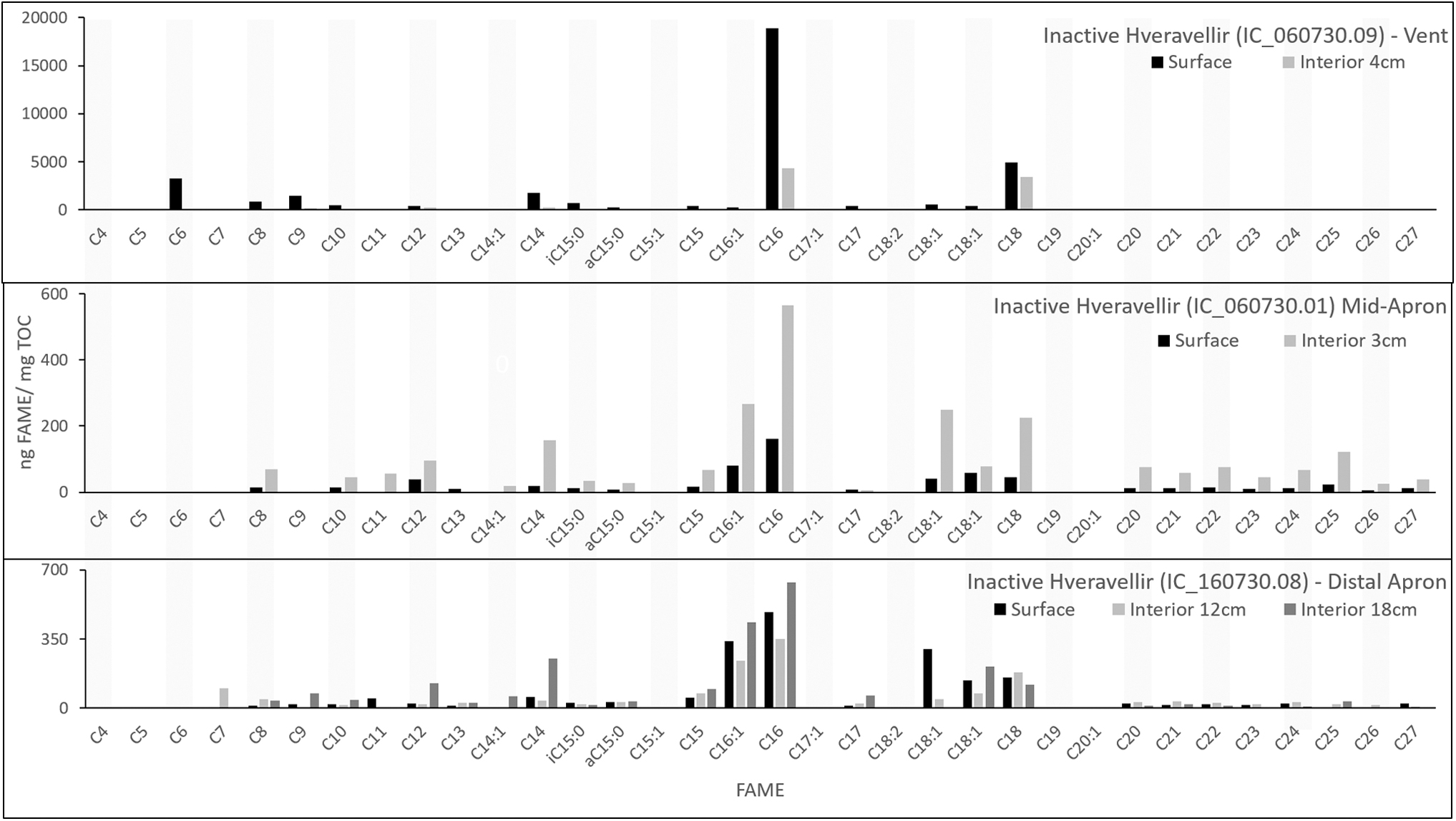
FAME profile for the active Hveravellir hot-spring system analyzed with a 500°C flash pyrolysis step.
At the inactive Gunnuhver site, samples were collected from two spring vents, the mid-apron, and the distal apron. FAMEs detected ranged from n-C4:0 to n-C27:0 and included few MUFAs (Fig. 6; Table 2). No methyl-branched fatty acids were detected. Again n-C16:0 and n-C18:0 were the most abundant FAMEs in all samples. FAMEs longer than n-C18:0 were only identified at the mid- and distal-apron sites. In general, the abundance and diversity of FAMEs were increased in the surface samples relative to the subsurface samples, which ranged from 5 to 7 cm in depth.
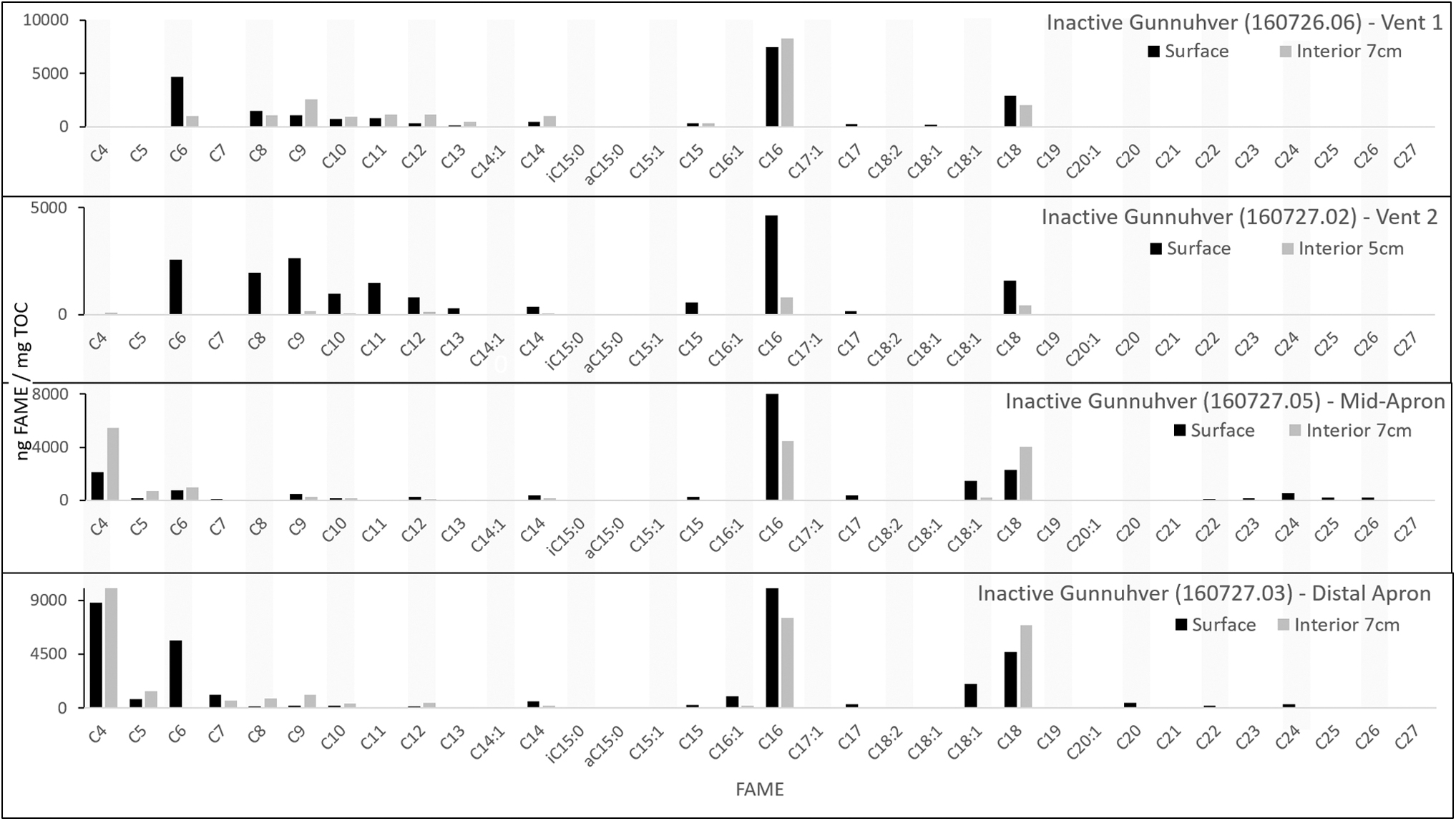
FAME profile for the inactive Gunnuhver hot-spring system analyzed with a 500°C flash pyrolysis step.
At the relict Lýsuhóll site, much of the characteristic features of the proximal slope, mid- and distal aprons were too degraded to assess, so samples were collected from three (near-)vent sites, which could be identified in field observations. FAMEs detected ranged from n-C6:0 to n-C27:0, and included a few MUFAs and one PUFA (Fig. 7; Table 2). Methyl-branched iso- and antiso-C15:0 were also detected in the surface and subsurface. Again n-C16:0 and n-C18:0 were the most abundant FAMEs in most samples, although this is not the case for Vent #1 (160809.03). FAMEs longer than n-C18:0 were identified at all sites. The abundance and diversity of FAMEs were increased in the surface samples relative to the subsurface samples, which ranged from 2 to 3 cm in depth.

FAME profile for the relict Lýsuhóll hot-spring system analyzed with a 500°C flash pyrolysis step.
3.3. FAMEs detected with SAM-instrument-like pyrolysis ramp
Samples were analyzed using a 35°C/min pyrolysis ramp to approximate the pyrolysis ramp used by the SAM instrument on the Curiosity rover (Mahaffy et al., 2012; Williams et al., 2019). This technique was utilized to explore how the FAME detection would change if sinter-bearing hot springs were analyzed on Mars with a SAM comparable TMAH experiment. In general, with this technique there was lower diversity and abundance of FAMEs in all three spring systems (Table 3).
Comparison of Flash Pyrolysis and Sample Analysis at Mars-Like Ramp Pyrolysis of Sinter Samples
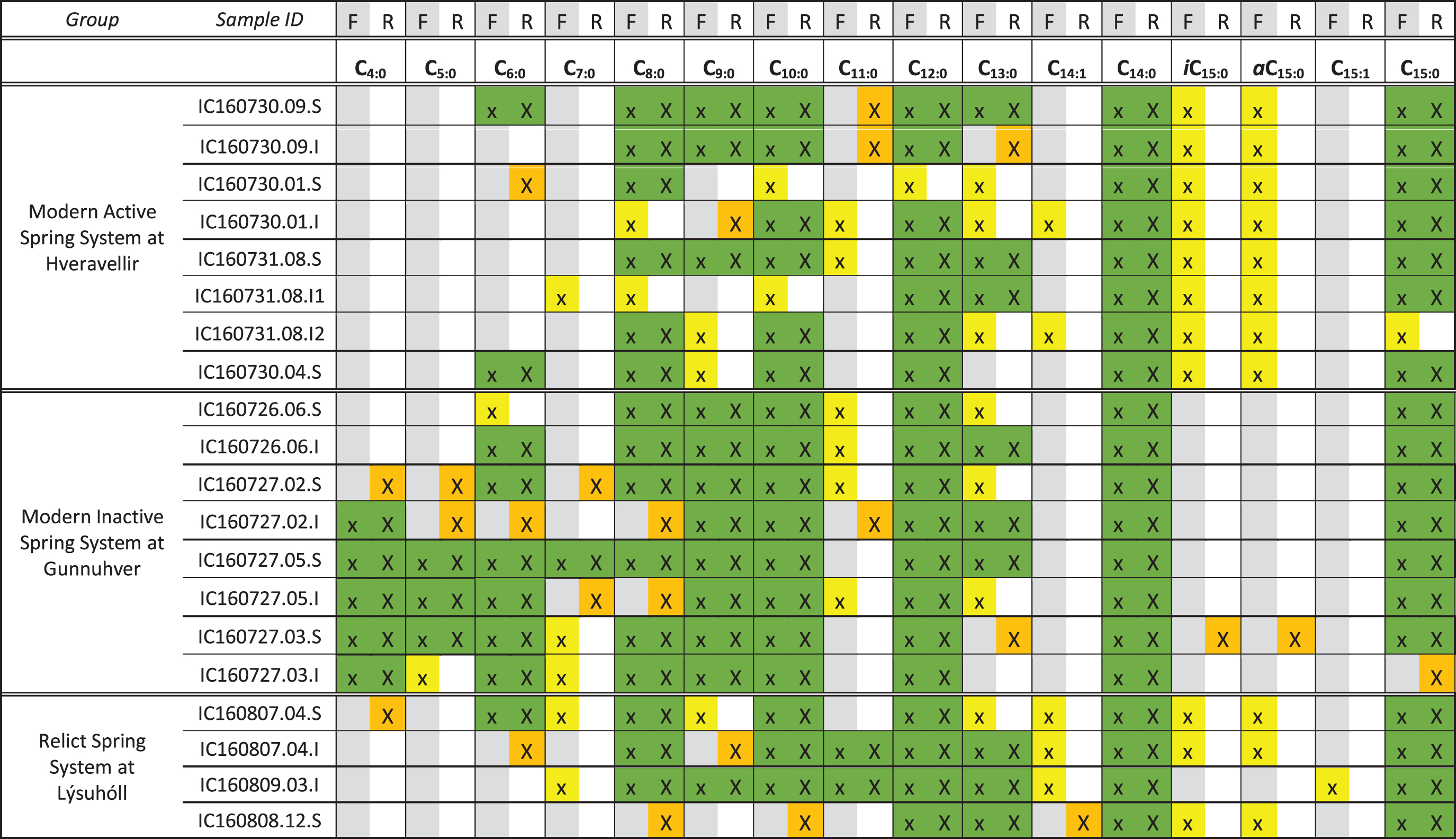
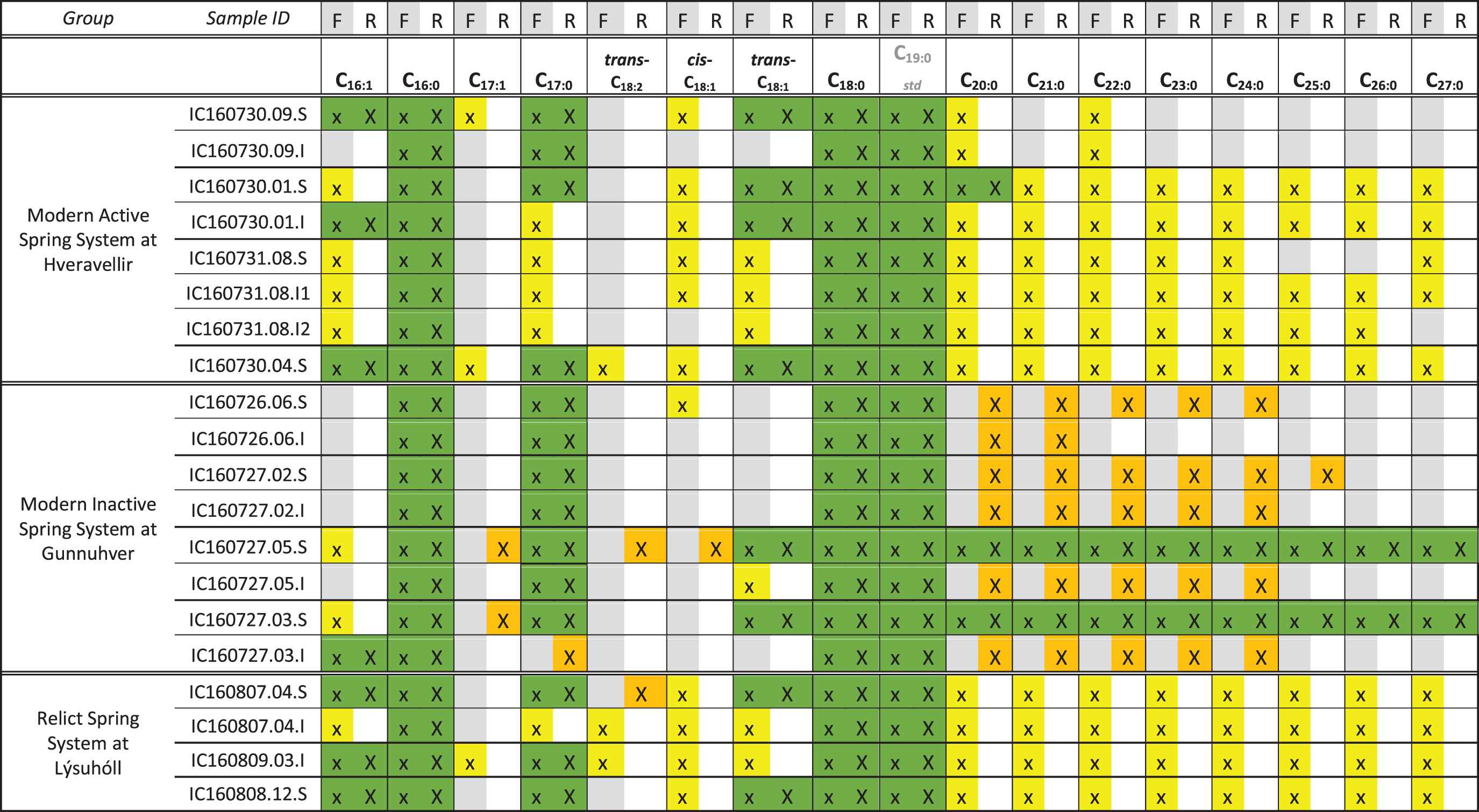
Green = FAME detection in both experiments. Yellow = FAME detection in flash pyrolysis experiment only. Orange = FAME detection in ramp pyrolysis experiment only. Blank cells = no FAME detection. Detection of the C19 standard is included for completeness. Gray columns correspond to flash pyrolysis, white columns correspond to ramped pyrolysis. Graphics are in color in the online version.
At the Hveravellir site, FAMEs detected ranged from n-C6:0 to n-C20:0 and included two MUFAs. n-C16:0 and n-C18:0 were the most abundant FAMEs in all samples, and only one FAME longer than n-C18:0 was identified at the surface mid-apron site. In general, the abundance and diversity of FAMEs were equal to or increased in the surface samples relative to the subsurface samples.
At the inactive Gunnuhver site, FAMEs detected ranged from n-C4:0 to n-C27:0, and included several MUFAs and PUFAs. n-C16:0 and n-C18:0 were generally the most abundant FAMEs in the mid- and distal-apron samples, and FAMEs longer than n-C18:0 were detected in all Gunnuhver samples. The abundance and diversity of FAMEs were higher in the surface vent samples relative to the subsurface samples, but lower than the subsurface samples from the mid- and distal-apron environments.
At the relict Lýsuhóll site, FAMEs detected ranged from n-C4:0 to n-C18:0, and included three MUFAs and one PUFA. n-C16:0 and n-C18:0 were the most abundant FAMEs in all samples, and no FAMEs longer than n-C18:0 were detected. The only site with both a surface and subsurface sample was the 160807.04 Vent #2 location, and the abundance and diversity of FAMEs were higher in the surface sample relative to the subsurface sample.
4. Discussion
The samples studied here comprise silica sinter and calcite hot-spring deposits ranging from modern and partially inactive to older, relict spring systems. Thus, they represent a range in geochemistry, mineralogy, age, and diagenetic history, as well as spanning a diversity of subenvironments present in hot-spring deposits: vents, mid-aprons, and distal aprons. Despite these differences, several trends in FAME abundance and diversity are shared between the hot-spring locations.
A high abundance of n-C16:0 and n-C18:0 is observed in all of the surface and subsurface samples relative to other FAMEs in all three locations, which is consistent with these two fatty acids being the most common in both bacteria and Eukarya (Kaur et al., 2015; Wilhelm et al., 2017; Johnson et al., 2020; Williams et al., 2019). Elevated n-C16:0 and n-C18:0 FAME abundances relative to other FAMEs are expected in younger sinters versus much older, inactive sinters (Kaur et al., 2011), reflecting more extant diverse microbial communities.
All samples contain FAMEs longer than n-C18:0, which most often represent a terrigenous plant or algal origin (Volkman et al., 1989; Kaur et al., 2011; Wilhelm et al., 2017; Johnson et al., 2020), although these fatty acids may be derived from bacterial-free, long-chain fatty acids (Summons et al., 2013) or intact polar lipids. Based on the abundance of surface vegetation near the spring sites, the FAMEs here are assumed to represent waxy longer chain FAMEs that are often more stable than short-chain FAMEs and may be more resistant to diagenetic processes and preserved longer after entombment (Wilhelm et al., 2017).
Long-chain FAMEs were identified in all surface and subsurface samples. At Hveravellir, they are present in greater abundance at the mid- and distal-apron sites than the vent site. At Gunnuhver, long-chain FAMEs are present in the mid- and distal-apron sites, and present at a lower abundance in the vent sites. This increase in abundance is consistent with models of lower temperatures (<40°C) downstream from vents, which support plant growth and the potential for their entombment in the sinter (Walter, 1976). At Lýsuhóll, these FAMEs are present in the relict vent sites both on the surface and in the subsurface.
Much higher concentrations of long-chain FAMEs in the surface samples suggest a more modern source. Observation of the long-chain FAMEs at depth suggests that plant matter grew and was entrained in the sinter as flow decreased and the spring system cooled and became dormant. The lower concentrations in the subsurface samples indicate some higher plants were also incorporated into the rock record, with some degradation of the FAMEs due to burial processes.
A strong even-over-odd carbon number preference is consistent with the production of the fatty acids due to enzymatic effects (e.g., the monomer addition of a 2-C subunit (Summons et al., 2008; Georgiou and Deamer, 2014) and of microbial cellular metabolic processes (Nishimura and Baker, 1986; Grimalt and Albaiges, 1987; Elias et al., 1997; Volkman, 2006). All surface and subsurface samples from the Hveravellir site exhibit this trend, as do all surface samples from Gunnuhver.
4.1. Organics load in the Hveravellir hot-spring system
The surface FAME load in the Hveravellir spring system varied greatly between the vent, mid-apron, and distal-apron facies. The abundance of FAMEs detected with flash pyrolysis at the opal-A vent facies is an order of magnitude (>35,000 ng total FAME/mg total organic carbon) greater than the FAME abundance at the amorphous silica distal apron (ca. 1875 ng total FAME/mg TOC). The amorphous silica mid-apron FAME abundance was lower still, at ca. 600 ng total FAME/mg TOC. The elevated FAME abundance at the Hveravellir vent sample indicates that the change in water flow and/or temperature at this recently ceased vent has likely led to colonization by bacteria with a lower temperature tolerance. Water flow and temperature are major factors in determining microbial diversity and biofilm texture in hydrothermal systems. When flow shifts, as it has at this vent and outflow system, change to abundance and diversity of the fatty acids should be expected. At the vent, the presence of the C20 and C22 fatty acids indicates that thermophilic bacterial growth has occurred; however, the relatively high fatty acid abundance, together with an increase in the cyanobacterial C18:ω9, indicates that a reduction in water temperature probably resulted in increased growth of a thermotolerant cyanobacterium such as the high-temperature form of Mastigocladus isolated from an Icelandic hot spring (Castenholz, 1969) or perhaps diatoms (e.g., Fig. 4), or both. This is consistent with the relatively low abundance and lack of C18:ω9 FA in the subsurface sample.
As we report on and discuss our results for the sinter samples below, it is important to keep in mind that these hydrothermal siliceous springs are dynamic systems suited for the production and preservation of fatty acid, and that under conditions ranging from recent to ancient, fatty acids survive for extended time entombed in siliceous sinter.
The opal-A pisolith layer was collected further away from the other three Hveravellir samples but exhibits a similar trend in even-over-odd carbon number predominance, with n-C16:0 and n-C18:0 dominating the FAME profile. Pisoliths are composed of >2 mm diameter spherical concretionary grains called pisolites, which are often formed from calcite or amorphous silica. Due to the concretionary nature of the grains, pisolites and related ooids often contain bound organic matter between cortices or submillimeter concretion layers (Davies et al., 1978; Folk and Lynch, 2001; O'Reilly et al., 2017). Therefore, the trends in presence and preservation of FAMEs in the pisolith layer are expected.
The FAME load at Hveravellir changed with depth between the vent, mid-apron, and distal apron facies as well. Most subsurface FAMEs decreased in abundance or were not detected relative to their complementary surface samples (e.g., Fig. 8, where the total organic carbon load is generally lower in subsurface samples relative to surface samples). A few exceptions exist in this data set, in particular in the mid-apron sample, in which all of the subsurface FAMEs were elevated in concentration relative to the surface (Fig. 5). In the distal-apron samples, several individual subsurface FAMEs were elevated relative to their complementary surface samples. This trend was distinct at the distal-apron site, where FAME concentration often increased stepwise from the surface to progressively deeper sampling depths (e.g., Fig. 5). We interpret this trend to represent either a temporary increase in organic preservation potential at spring sites that experience lower temperatures and diagenetic alteration, or a modern and robust subsurface microbial community that is generating the elevated FAME abundance.

Comparison of total FAME abundance (in nanogram FAME/gram TOC) detected with the 500°C flash pyrolysis and the SAM-like 35°C/min ramp pyrolysis methods at each sinter sample location. The number of unique FAMEs detected with each pyrolysis method is included on the secondary (right) y-axis. Flash pyrolysis nearly always yielded comparable numbers of FAMEs relative to ramp pyrolysis. TOC, total organic carbon.
This trend could also be an indication of preservational bias related to differences in mineralization rates, where early mineralization minimizes the degradation of organics and body fossils (Alleon et al., 2016). If a diversion of water flow precipitated a localized temperature shift, the supply of dissolved silica would also decrease leaving the microbial community more vulnerable to biotic and abiotic degradation processes since it was not mineralizing. These observations reinforce the complexity of these spring systems. While general trends can be preserved, changes in the local physical conditions (like diverted water flow) can have cascading impacts on the microbial composition and their ecology, that together end up biasing their fossil record.
Although our sampling techniques were optimized to decrease any chance of anthropogenic contamination, we cannot rule out the possibility that some of these samples reflect an anthropogenic influence, as this hot spring is a tourist destination and the mid-apron sample was collected below a visitor boardwalk. Elevated FAMEs from eukaryotic (e.g., human) sources may have been entrained in the sinter. Contamination of this type should yield a much higher FAME concentration, especially in the C16:0 and C18:0 FAMEs, in the surface relative to the subsurface samples. Surface samples did contain higher concentrations of FAMEs than the subsurface, but C16:0 and C18:0 FAMEs were not uniformly elevated. Therefore, we interpret the Hveravellir FAME concentration trends as reflecting a diverse and active microbial community.
In general, branched and unsaturated fatty acids were either not detected in subsurface samples or were decreased in abundance relative to the surface samples. This degradation is expected as the unsaturation in the fatty acid is susceptible to cleavage by microbial heterotrophy or diagenesis (Sun et al., 1997; Killops and Killops, 2005). A decrease in unsaturated fatty acids and branching is documented in older sinter samples (Kaur et al., 2011), and is consistent with the expectation for preferential degradation of unsaturated fatty acids. However, this trend was reverse in the mid-apron sample, in which all of the subsurface FAMEs, including branched and MUFAs, were elevated in concentration relative to the surface.
4.2. Organics load in the Gunnuhver hot-spring system
The surface FAME load in the inactive Gunnuhver spring system also varied between the vent, mid-apron, and distal-apron facies, although to a lesser degree than in the Hveravellir spring system. The abundance of FAMEs detected with flash pyrolysis is comparable between the opal-CT vent and opal-CT mid-apron facies (ca. 18,000–20,000 ng total FAME/mg TOC), and is elevated in the amorphous silica distal-apron facies (ca. 37,000 ng total FAME/mg TOC). This trend in abundance is expected, as a greater diversity of organisms with higher growth rates tend to colonize lower temperature geothermal regions such as the distal apron.
The abundance and diversity of lower molecular weight FAMEs increase to the mid-apron and distal-apron facies relative to the vent facies, suggesting that higher molecular weight (longer) FAMEs are not less well preserved further from the vent and are likely degraded to shorter chains, potentially due to microbial heterotrophy.
The FAME load in this system changed with depth more dramatically than in the Hveravellir system. Subsurface FAME abundance decreases substantially or to no detection relative to their complementary surface samples, indicating continued degradation of the FAMEs in the subsurface due to diagenesis and microbial recycling. Only four FAMEs (n-C8:0, n-C10:0, n-C15:0, and trans-C18:1) are detected in the subsurface distal-apron site with flash pyrolysis. The subsurface FAME profiles exhibit strong-to-weak even-over-odd trends in carbon number. The subsurface distal-apron sample does not exhibit any trends in carbon number or molecular weight, which likely reflect whole-scale degradation of all FAMEs in the subsurface. MUFAs and PUFAs are already limited in detection at the surface, a dearth that continues into the subsurface and indicates preferential degradation of the unsaturated FAMEs. Branched fatty acids are not detected in either the surface or subsurface samples with flash pyrolysis.
4.3. Organics load in the Lýsuhóll hot-spring system
In contrast with the opal and amorphous silica-dominated Hveravellir and Gunnuhver sites, the Lýsuhóll relict spring system is composed of both amorphous silica and calcite, with Vent #3 composed entirely of calcite. The spring waters that precipitate these deposits, and the subsequent microbial communities that colonize the springs, may be different from those present in the siliceous sinter deposits (Zhang et al., 2004). In addition, several samples from the relict Lýsuhóll system were destroyed and/or contaminated during transport back to the United States, reducing the completeness of the surface versus subsurface samples originally collected at these sites. Regardless, several conclusions can be drawn from the remaining samples about the organics load in this relict system.
All sample sites at Lýsuhóll represented near-vent environments as these were the most readily identified in older, degraded hot-spring deposits. The surface FAME load in the relict Lýsuhóll system also varied between the vent facies, ranging from ca. 3700 ng total FAME/mg TOC in the amorphous silica- and calcite-dominated Vent #2, to ca. 350 ng total FAME/mg TOC in the amorphous silica Vent #1, to ca. 83 ng total FAME/mg TOC in the calcite Vent #3 (Table 2). As in the other spring systems, the surface FAME concentrations are higher than in subsurface samples. The total FAME abundance ranges from ca. 3700 ng total FAME/mg TOC in the Vent 160807.04 (Vent #2) surface sample to ca. 1500 ng total FAME/mg TOC in the complementary subsurface sample. This indicates continued degradation of the FAMEs in the subsurface due to diagenesis and microbial recycling.
The Lýsuhóll samples exhibit a moderate-to-weak even-over-odd preference, which may indicate degradation of fatty acids from the original microbial community, and limited input of organics from a modern community. The relict Lýsuhóll system is the only site where the PUFA trans-C18:2 algal biomarker is detected (Campbell et al., 2015), and then only in two subsurface samples. Branched and unsaturated fatty acids are already limited in detection at the surface, a dearth that continues into the subsurface and indicates preferential degradation of these fatty acids.
4.4. Comparison between flash pyrolysis and flight-like ramp pyrolysis
The 500°C flash pyrolysis method, combined with TMAH thermochemolysis, represents a laboratory-optimum method to release organics bound within macromolecules and sorbed to minerals. The SAM instrument on the Curiosity rover, while able to perform similar experiments, is limited to a 35°C/min pyrolysis ramp. To assess how this method variation might affect the detection of FAMEs in these Mars-analog siliceous sinters, samples were analyzed with the SAM-like 35°C/min pyrolysis ramp.
The flash pyrolysis method improved FAME detection uniformly across the sample suite (Fig. 8), and with few exceptions, FAMEs >n-C18:0 were only detected with the flash method. The ramp method released between 30% and 86% of the FAMEs detected in the flash method. Although the flash method is more efficient at FAME release, it is promising for the search for organics on Mars that FAMEs are also detectable with the SAM instrument pyrolysis ramp rate.
Correlations vary between the number of FAMEs detected and the concentration of FAMEs in the sample suite. In the Hveravellir system, the greatest diversity of FAMEs detected was generally correlated with low percentage TOC. This diversity is invaluable in assessing microbial community composition, even at low concentrations. In the inactive spring system, the greatest diversity of FAMEs correlates with higher percentage TOC, which represents an anticipated trend with regard to FAME preservation (and degradation), FAME diversity and concentration, and the total organic carbon load in the sample (Table 1). The greatest diversity of FAMEs was detected in the relict spring system, as were some of the highest percentage TOC values. In particular, n-C16:0 concentrations were predictably low in the subsurface samples and high in the surface samples. These lines of evidence indicate that the relict system is experiencing higher delivery of organic load now than it perhaps did in the past. The organics that constitute these higher percentage TOC values may represent a far greater diversity of organic molecules than just fatty acids, which are preserved but in low concentrations in the relict system.
4.5. Mineralogic evolution and the preservation of organic matter
It is important to note that the primary controls on lipid preservation in siliceous sinter deposits are thermal alteration, high temperatures, and the pH of each spring (Pancost et al., 2006; Kaur et al., 2015), which can vary greatly between spring systems. A great variety of lipid biomarkers have been identified in modern to ca. 900 years old geothermal environments (Shiea et al., 1991; Jahnke et al., 2001; Pancost et al., 2005, 2006; Zhang et al., 2007; Kaur et al., 2011). Often branched or unsaturated fatty acids are prevalent in younger deposits (Kaur et al., 2011), as was also demonstrated in the work presented here. In an actively flowing spring system, temperature in particular tends to strongly influence the length and therefore abundance of hydrocarbons, such as bacterial diethers in active geothermal systems (Kaur et al., 2015). Thermophilic organisms tend to have higher molecular weight diethers with higher thermostabilities, longer fatty acid chains (Weerkamp and Heinen, 1972; Oshima and Miyagawa, 1974; Russell, 1984), increased ratios of iso- to anteiso-branched fatty acids (Shen et al., 1970; Oshima and Miyagawa, 1974), and fewer unsaturations and branches (Daron, 1970; Ray et al., 1971). All these adaptations serve to maintain optimal membrane fluidity at higher temperatures.
Although the Hveravellir vent sampled here was no longer actively flowing, several of these trends were observed (Fig. 9), consistent with the recent cessation of flowing spring water. The average chain length ranges from 15.1 to 16.4 for all surface and subsurface Hveravellir samples, with the exception of the surface vent sample with an average chain length of 13.6. These values suggest that the surface vent microbial community and/or FAME preservation is different from the other surface and subsurface samples at Hveravellir. The low ACL value of the surface vent suggests that this spring vent never experienced very high fluid temperatures and even if thermophilic organisms were present, their organic biosignatures are not well preserved. An increase in heterotrophy may have occurred concurrent with the cessation of water activity that hosted the thermophiles. The ratio of i- to a-C15 is elevated near the vent where elevated temperatures affect community composition and the iso- to anteiso ratio trend was consistent at this site between the surface and subsurface samples. In addition, the ratio of branched to total fatty acids increases with distance from the higher temperature vent, consistent with a decrease in the dominance of any thermophilic organisms.

Fatty acid (FA) distribution as determined with flash pyrolysis across hot-spring facies, with locations “vent,” “mid-apron,” and “distal apron” representing environmental facies as determined by textural characterization.
In contrast, the inactive Gunnuhver site had a relatively consistent, elevated total FAME concentration at the vent and mid-apron facies, and a much higher FAME concentration at the distal apron facies, consistent with secondary recolonization of the sinter by more diverse, lower temperature mesophiles after spring flow ceased. Average chain length also decreases from the vent facies (ACL = 10.7–13.4 at vents) to the distal-apron facies (ACL = 0–8.9), suggesting the degradation of fatty acids to shorter chains due to heterotrophy or diagenesis.
Only relict inactive vents were sampled at the Lýsuhóll spring site, which allows a comparison with the modern inactive vent systems. The total FAME concentration at the relict site was within the range of the inactive spring sites, with higher abundances in the surface samples and lower abundances in the subsurface samples. Average chain length at the relict site (ACL = 14.9–17.4) is within the range of the Hveravellir spring site, suggesting that these Lýsuhóll vents were higher temperature when flowing and the thermophile FAMEs are well preserved.
The iso- to anteiso ratio was lower than that at the modern inactive spring sites, suggesting degradation of the branched fatty acids from an original, higher temperature population. The ratio of branched to total fatty acids, however, was comparable with or higher than the Hveravellir vent site, indicating that more branched fatty acids relative to the total fatty acid load are preserved. This trend may be due to a greater initial branched fatty acid load that, along with the total fatty acids, has experienced degradation. The branched to total fatty acid ratio at Vent #3 (160807.12) is higher than that obtained for all other samples and contains the longest ACL of all samples from this study. This vent is composed solely of travertine, which may introduce a preservation bias between this younger calcite deposit and the other vents that are older and silica dominated. The high branched to total fatty acid ratio and ACL value of Vent #3 may also correspond to an active mesophilic microbial community at this site.
Low-temperature degradative processes such as heterotrophy and diagenesis control the fatty acid distribution observed in this study. The primary fatty acid distribution from thermophilic versus mesophilic communities is discernable in some environments (e.g., Hveravellir vent) but not diagnostic of more specific temperature and pH conditions of each subenvironment at each spring system. The vents from the three systems studied here are compared to assess the preservation of organics as a concentration of FAMEs in the surface and subsurface (Fig. 10). Additional work can be completed to assess the preserved lipids separate from modern lipids in all three systems, including solvent washing sinter samples to remove surface lipids before analysis.

Comparison of FAME concentrations determined with flash pyrolysis experiments from the surface and subsurface vent sites from the Hveravellir, Gunnuhver, and Lýsuhóll spring systems. The even-over-odd carbon number preference is most apparent in the Hveravellir and Gunnuhver locations. Color images are available online.
At the recently inactive Hveravellir vent (sample 09), the C16 and C18 FAMEs are likely elevated due to these being the dominant FAMEs in both bacteria and eukarya, although anthropogenic influences, especially in the surface sample, cannot be completely dismissed. The lower concentrations of the other FAMEs in the surface and subsurface reflect a nominal FAME profile from an active microbial community, with the expected even-over-odd carbon number preference. The inactive Gunnuhver vent generally had the lowest FAME concentrations relative to the Hveravellir and Lýsuhóll systems, but still exhibited the even-over-odd carbon number preference characteristic of an active microbial community. The low FAME concentrations reflect FAME degradation leading to low preservation potential and a lack of extensive recolonization of the sinter by secondary microbial communities. The higher FAME concentrations at the relict Lýsuhóll Vent #2 (especially relative to the inactive Gunnuhver site) likely reflect microbial (re)colonization by a community distinct from that found in active/recent silica-precipitating hot springs. The mixed amorphous silica and calcite mineralogy of this vent likely also contributes to this marked difference in FAME abundance.
The mineralogic evolution and diagenesis of siliceous hot-spring sinters are well studied (Herdianita et al., 2000). Initial silica precipitates are composed of amorphous silica or opal-A (Tobler et al., 2017), and the very fine size of these precipitates is documented to preserve microbial cells in extraordinary physical detail (Cady and Farmer, 1996; Renaut et al., 1998; Jones et al., 2000), although progressive silicification and preservation biases over long timescales can also destroy these physical biosignatures (Jones and Renaut, 1996; Jones et al., 1997). As siliceous sinter deposits experience diagenesis, opal-A incrementally recrystallizes to Opal-C (containing disordered α-cristobalite) or opal-CT (containing cristobalite and tridymite). This transition may occur on the order of ca. 50 years up to tens of thousands of years. Over longer timescales (ca. 10,000–50,000 years), opal-C/-CT will reorder to microcrystalline quartz. Although sinter age can sometimes be used as an estimate for crystallinity, variables such as fluid mobilization and composition can also greatly affect crystallinity (Herdianita et al., 2000). This recrystallization process, specifically the transition to microquartz, reduces porosity and decreases the preservation of organic matter in siliceous sinter (Teece et al., 2020).
These trends in mineralogy and organics preservation were upheld in this study. In the Hveravellir system, which is composed of amorphous silica and opal-A, the diversity and abundance of FAMEs were elevated in both the surface and subsurface. In contrast, the inactive Gunnuhver system, which is composed primarily of opal-CT, had a much lower diversity and abundance of preserved FAMEs. Lynne et al. (2006) suggest that silica diagenesis is accelerated in hot-spring systems that have transformed from flowing water to steam dominated. The transformation to opal-CT in this relatively young system was likely controlled by fluid composition and fluid or steam mobilization. This mobilization led to accelerated diagenesis and fumarolic degradation of organics in the Gunnuhver system.
The surprisingly elevated diversity and abundance of FAMEs in the relict Lýsuhóll system were controlled in part by the mineralogy, which is dominated by calcite and amorphous silica, as well as a modern microbial community that had colonized the surface of this deposit. These results further reinforce the importance of ascertaining sample mineralogy before sampling for organics in hydrothermal systems, especially on life detection missions beyond Earth.
4.6. Biomarker preservation and implications for organics detection on Mars
The size and distribution of hot-spring facies vary across systems, yet trends in preservation styles, from the outcrop- to the submicrometer scale, are present and can be used to provide depositional context as we continue to explore life in other planets. On Earth, preservation at the vent is often less favorable with microfossils usually in the form of smaller diameter molds that can be readily infilled and obscured (Cady and Farmer, 1996). Vents may be identified by finely laminated deposits in a circular form that are often associated with geyserite. The directed water flow in channel facies produces bacterial streamer fabrics, long filaments parallel to water direction, sometimes composed of cyanobacterial filaments (Parenteau et al., 2014). This streamer morphology is readily identified in outcrop (e.g., Walter et al., 1998; Parenteau et al., 2014). The lower temperatures of the distal apron are usually accompanied by thick microbial mats creating palisade fabrics with abundant microfossils and organics that can be preserved (Cady and Farmer, 1996; Campbell et al., 2015). Apron facies are one of the best preserved lithofacies in degraded spring systems, with the mid-to-distal apron composing the largest aerial extent, followed by the channel facies and then vent facies. Based solely on the aerial extent, martian hot-spring apron facies may be most easily detected from orbital imaging and targeted for in situ exploration.
Biosignature preservation within all facies is complicated by preservation bias and diagenesis over geological timescales. Even the proximal slopes and aerially extensive mid- and distal aprons may become challenging to identify as the fine scale microtextures that define these spring subenvironments can be lost to diagenesis and preservation biases (Cady and Farmer, 1996). Although diagenesis can and does obscure some features, the apron facies contain structures such as pools and terracettes that are identifiable in surface deposits and layered deposits with palisade fabric identifiable in outcrop cross-section. Vents are just as prone to erosion and infilling (this was observed at Lýsuhóll and Gunnuhver), and with a much smaller footprint, can be challenging to identify from orbital images. However, the primary macroscale textural characteristics of spring vents can enable their identification after some geological reworking (Cady and Farmer, 1996, and references therein).
Given these complexities, it is therefore important to develop a facies-oriented understanding of biosignature preservation potential in terrestrial hot springs to better predict the ideal locations to search for organics in martian relict hydrothermal spring systems. The temperature of fluids at subaerial hydrothermal vent systems can vary greatly, up to or above surface boiling temperatures. Nonphotosynthetic microbes can live in these temperatures, with hyperthermophiles preferring >80°C up to a maximum of 110°C where elevated hydrostatic pressure allows waters to reach higher temperature without boiling (Segerer et al., 1993). For reference, the upper temperature limit for archaeal life is 121°C in submarine black smoker environments (Kashefi and Lovley, 2003); in contrast, photosynthetic microbes are present in waters <73°C (Ward et al., 2012). Although these organisms do not directly contribute to silica precipitation in these spring systems, they can be entombed and preserved in the rapidly precipitating silica, contributing to the organic load and microstructures found in near-vent environments (Cady and Farmer, 1996).
In our study, silicified traces of microorganisms are readily identified in SEM images from the Hveravellir and Lýsuhóll sample sites, but no distinct biogenic features were identified from the Gunnuhver sinter samples (Fig. 4). These observations appear to correlate with the relative abundances of organics detected at these three locations.
The search for (biogenic) organic molecules on Mars' near-surface requires new and novel techniques to detect these molecules with miniaturized space-flight payloads. In the terrestrial realm, many biomarkers are preserved and detectable in siliceous sinter environments with laboratory instrumentation such as offline lipid fraction extraction, derivatization, and/or saponification, followed by multiple GC-MS and/or liquid chromatography-mass spectrometry analyses. However, suites of these laboratory techniques are often too complex to be successfully miniaturized for space-flight and remote operation. Therefore, “one-step” thermochemolysis or derivatization pyrolysis-GC-MS is an optimized technique that is miniaturized to work on space-flight missions such as with the SAM and MOMA instruments.
To be more “SAM-like” in this study, samples that may contain low-temperature surface biofilms were collected and ground as whole-rock samples in a manner similar to Curiosity's sample acquisition system. Therefore, the results from this study likely reflect a greater abundance of organics than may be expected to be preserved in a martian hot-spring deposit with a hypothetical microbial community. Although more diagnostic biomolecules are present in Mars-analog hydrothermal spring environments, this work demonstrates that FAMEs are preserved and detectable with space-flight-like GC-MS techniques in modern inactive and relict siliceous sinter and calcite deposits from Iceland. The MOMA instrument on the Rosalind Franklin rover will be able to ramp the pyrolysis oven at >200°C/min (from thermal vacuum), achieving a much faster ramp than the SAM instrument and perhaps increasing the likelihood of detecting martian FAMEs, if present.
The rate of siliceous sinter formation, hydrothermal conditions, and those effects on biomass growth and potential for silicification in each individual Icelandic sinter deposit reflect differences in the diversity and abundance of biomolecules detected in this work. The geological process of a hot spring becoming quiescent should be similar on Earth and Mars. The evolution of a putative microbial community within that hot spring can be approximated in specific ways by assessing microbial community evolution in hot springs on Earth. The approximations require assumptions that do not account for Mars-specific conditions such as atmospheric composition, radiation flux, and energy sources. By assessing the microbial communities and preserved organics from lower temperature inactive and relict spring systems, this work approximates the organics load in martian hot springs with a hypothetical microbial community that would have evolved as the spring system cooled to dormancy. Understanding the distribution of organics in terrestrial systems is vital for choosing an ideal martian hydrothermal target for organic biosignature exploration.
A final consideration in the search for organics on Mars is the deleterious effects of oxidants and radiation on organics. Although organic matter has been identified in Mars near-surface substrates with the SAM instrument (Freissinet et al., 2015; Eigenbrode et al., 2018), fewer molecules detected by SAM represent reduced carbon, with many of these organics being oxidized. For example, polycyclic aromatic hydrocarbons and kerogen from exogenous delivery through meteoritic infall may undergo several oxidation reactions to form stable simple organics such as phthalic, mellitic, or oxalic acids (Benner et al., 2000; Freissinet et al., 2015) on the martian surface, which are then detectable by the SAM instrument [e.g., possible source organics for benzene in the Sheepbed mudstone, Freissinet et al. (2015)]. These oxidation processes are expected to occur both on the surface and in the subsurface of Mars.
Ionizing radiation is another serious concern for the long-term preservation of martian organics. The ionizing radiation dose on the surface of Mars is estimated at 0.54–0.85 Gy/year (Dartnell et al., 2007). Ionizing radiation destroys biomolecules (e.g., proteins or DNA), and most bacteria cannot survive doses >200 Gy (Daly, 2009). Recent studies indicate that within 300 million years, preservation of functionalized organic molecules >500 amu in the top ∼5 cm of rock drops 1000 × due to ionizing radiation (Pavlov et al., 2012). There is some indication that rapid burial of organics, short exposure ages due to relatively slow martian weathering rates, and the characteristics of some entombing mineralogies may assist in organics preservation in the martian near-surface (Hays et al., 2017 and references therein), but the need remains to sample the martian subsurface to reduce the impact of degradation through radiation effects.
The Curiosity rover can drill to a maximum of 6 cm, and the Rosalind Franklin rover will be able to drill to 2 m depth. The work presented here indicates that even with diagenetic effects and heterotrophic cycling of organics, organic molecules are preserved and detectable over short timescales in the near-subsurface (2–18 cm in depth) of modern and relict terrestrial hot springs. Organics may therefore be detectable on Mars in relatively recent hot-spring deposits, but preservation likely deteriorates over geological timescales. These depths are within the drill sampling range of the Curiosity and Rosalind Franklin rovers (partial to complete depths, respectively), and support the search for near-surface organics preserved in martian hydrothermal systems.
5. Conclusions
Hydrothermal spring deposits host unique microbial ecosystems, and have the capacity to physically preserve both microbial communities and their associated organic matter as biosignatures within siliceous sinter. These terrestrial geothermal springs serve as Mars-analog environments to prepare future Mars missions to target their search for biosignatures in martian hot-spring deposits. This study quantified the preservation and detectability of fatty acids in three Icelandic hot-spring deposits ranging from modern and inactive to relict, and samples were collected both at the surface and at depth (ranging from 2 to 18 cm in depth). Analyses performed with TMAH thermochemolysis pyrolysis-GC-MS indicated that fatty acids were preserved and detectable in all three spring deposits, with the subsurface samples degraded relative to the surface samples. TMAH thermochemolysis pyrolysis-GC-MS analyses using a SAM-instrument-like procedure were also successful in liberating preserved fatty acids from these sinter samples, although generally in lower abundances and with less diversity. These results build confidence in our ability to detect fatty acids and distinguish biogenicity when using the GC-MS instrumentation available on current and future Mars missions.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Financial support for sample collection was provided by the NASA Planetary Science and Technology through Analog Research (PSTAR) program grant #NNX15AJ38G to J.R.S. and the American Philosophical Society’s Lewis and Clark Fund for Exploration and Field Research in Astrobiology to K.L.C. S.S.J. acknowledges support from NASA grant #NNX17AJ68G (“Using Organic Molecule Detections in Mars Analog Environments to Interpret the Results of the SAM Investigation on the Mars Science Laboratory Mission”) as well as a Georgetown University Pilot Research Grant.
Abbreviations Used
Associate Editor: Christopher McKay