
Abstract
The question whether organic compounds occur on Mars remained unanswered for decades. However, the recent discovery of various classes of organic matter in martian sediments by the Curiosity rover seems to strongly suggest that indigenous organic compounds exist on Mars. One intriguing group of detected organic compounds were thiophenes, which typically occur on Earth in kerogen, coal, and crude oil as well as in stromatolites and microfossils. Here we provide a brief synopsis of conceivable pathways for the generation and degradation of thiophenes on Mars. We show that the origin of thiophene derivatives can either be biotic or abiotic, for example, through sulfur incorporation in organic matter during early diagenesis. The potential of thiophenes to represent martian biomarkers is discussed as well as a correlation between abundances of thiophenes and sulfate-bearing minerals. Finally, this study provides suggestions for future investigations on Mars and in Earth-based laboratories to answer the question whether the martian thiophenes are of biological origin.
1. Introduction
The first detection of organic compounds on Mars was done by gas chromatography-mass spectrometry (GC-MS) on the Viking landers, which revealed the presence of chloromethane and dichloromethane. At that time, this detection was interpreted as terrestrial contamination, but it is now realized that it derived most likely from the reaction of indigenous martian organic compounds with perchlorate (Navarro-González et al., 2010). A reanalysis of Viking mission data was also consistent with the detection of aromatic organic compounds such as chlorobenzene (Guzman et al., 2018). Recently, chlorobenzene (150–300 ppb) was also detected by the Curiosity rover in the Sheepbed mudstone of Gale Crater (Freissinet et al., 2015). In addition, C2 to C4 dichloroalkanes (up to 70 ppb) have been found, but it could not be completely ruled out that these compounds resulted from instrument background sources (Freissinet et al., 2015).
A more convincing discovery of the existence of indigenous organic compounds on Mars was the detections reported by Eigenbrode et al. (2018) in lacustrine mudstone of the Murray formation in Gale Crater. They found several aliphatic, aromatic, and thiophenic compounds at nanomolar levels through evolved gas analysis (EGA) and pyrolysis-GC-MS. Many more organic compounds are likely present as the methodology of pyrolysis-GC-MS was recently been shown to inhibit the detection of organic compounds when perchlorates and oxyanion salts are present (Montgomery et al., 2019).
Nevertheless, the organic matter detected by Eigenbrode et al. (2018) contained 5% carbon as organic sulfur molecules. Interestingly, the sulfur-containing molecules detected on Mars include thiophene and some of its derivatives (2-methylthiophene, 3-methylthiophene, and presumably benzothiophene) as well as smaller fragments (methanethiol, dimethyl sulfide, carbonyl sulfide, carbon disulfide, hydrogen sulfide, and sulfur dioxide) (Eigenbrode et al., 2018). Since sulfur compounds are essential for life and thiophenes play an important role in the sulfur cycle on Earth (Kertesz, 2000), we present here a brief summary of conceivable biotic and abiotic production and degradation pathways of martian thiophenes and their potential in representing biomarkers.
2. Thiophenes on Earth
2.1. Thiophenes—formation in natural environments and significance as biomarkers
Thiophenes are aromatic five-membered rings containing four carbon atoms and one sulfur atom. They occur naturally in kerogens, bitumen, coals, crude oil (petroleum), and sediments at concentrations up to ∼10 wt % (Table 1).
Examples of Total Sulfur and Thiophenic Sulfur Contents in Various Sulfur-Rich Environments
Referred to total sulfur, unless otherwise stated.
Only organic sulfur.
Range of masses of dibenzothiophene in 1 g marine sediment soils.
n.d., not determined.
In these environments, the thiophenes are thought to be produced abiotically during diagenesis where inorganic sulfur nucleophiles such as sulfides (e.g., H2S) react with organic matter at elevated temperatures (Kohnen et al., 1991; Krein and Aizenshtat, 1994). However, the parent substances of these reactions can indeed be biological in origin. The production of sulfides by sulfate-reducing bacteria is driving the sulfurization of organic matter during diagenesis (Kenig and Huc, 1990), whereas the organic matter itself includes biomolecules such as lipids or terpenoids (Simoneit, 2005).
One proposed mechanism of the formation of isoprenoid thiophenes detected in many sediments is the incorporation of sulfur into chlorophyll-derived phytol or bacteriochlorophyll-derived phytenes to yield polysulfides followed by the thermal conversion into thiophenes (Brassell et al., 1986; Krein and Aizenshtat, 1994). It even has been argued that the ratios of different isoprenoid thiophenes can be used to distinguish between hypersaline and nonhypersaline paleoenvironments depending on the occurrence of certain bacteriochlorophylls in these environments (Leeuw and Sinninghe Damsté, 1990).
Thus, thiophenes can represent at least secondary biomarkers. In general, due to the formation of intermolecular sulfur bonding (polymerization), sulfur-rich biomarkers are more stable and preservable than biomarkers with a low sulfur/carbon ratio and are, therefore, prolonging the lifetime of these biomarkers in the geological record (Sinninghe Damsté et al., 1989; Lemelle et al., 2008). It has been shown that even at relatively high levels of thermal maturity, alkylthiophenes are stable thermal degradation products wherein the positions of the thiophene moiety coincides with the original positions of the functional groups of their precursors (Koopmans et al., 1995, 1996). This implies that alkylthiophenes are useful biomarkers because the information regarding the original positions of functionalities in the precursor molecules is retained (Koopmans et al., 1995).
Indeed, thiophenes were detected in the cell wall of 700–800 Ma-old Neoproterozoic microfossils (Lemelle et al., 2008) as well as in 2.72 Ga-old fossil stromatolites (Lepot et al., 2009). Although it remained unclear whether the thiophenic compounds are of endogenous origin or were produced through diagenesis of the microfossils, Lemelle et al. noted that the low sulfur content (10–300 ppm) within the cell-like structures points toward an endogenous origin because these quantities are more comparable with sulfur contents in living microorganisms (e.g., 4000 ppm for single marine cyanobacterial cells) than with the sulfur content in kerogens, which is usually at a percentage level (Table 1).
The thiophene synthesis during diagenesis is influenced by at least six major aspects: Thermal maturation: with increasing maturity (higher temperatures and pressures during diagenesis and early catagenesis), the total sulfur content of the environment (kerogen, oil, or coal) decreases, for example, by outgassing of H2S. Nevertheless, the relative abundance of thiophenic compounds (especially dibenzothiophenes) increases due to their enhanced thermal stability (Orr and Sinninghe Damsté, 1990). Content of total sulfur and organic carbon: the thiophene concentration in sediments and coals correlates with the content of organic carbon and total sulfur (Yang et al., 1998; Wawrzynkiewicz and Sablik, 2002). Clay content: sediments having higher clay content favor accumulation of dibenzothiophene (Yang et al., 1998), although, in general, the sulfur content in clay-rich sediments is low (see Point 4). Carbonate content: carbonate deposits contain larger amounts of organic sulfur compounds (including thiophenes) than clay-rich clastic environments (Kenig and Huc, 1990). The reason for this observation might be that clay-rich clastic deposits contain more iron than carbonate muds (see Point 5) (Krein, 1993). Iron concentration: iron competes successfully with organic matter as a sink for sulfur by the formation of pyrite (Berner et al., 1985). Thus, thiophene synthesis in iron-rich environments is limited to a certain degree. Sulfate-reducing bacteria: when the microbial H2S production exceeds the formation of pyrite, the rate of sulfur incorporation into organic matter during early diagenesis is increased (Kenig and Huc, 1990).
2.2. Biosynthesis of thiophenes
Besides the formation of thiophenic compounds as secondary biomarkers during early diagenesis, only a few examples of direct biosynthesis of thiophenes are described. Caldariellaquinone (CQ) is a benzo[b]thiophen-4,7-quinone (Fig. 1A) that first has been isolated from the extremely thermo- and acidophilic archaeon Caldariella acidophila (de Rosa et al., 1975, 1977). Today, it is known that CQ is the major component of the quinone fraction in all known Sulfolobus spp. (Zhou and White, 1989). In Sulfolobus solfataricus, a CQ analog—benzodithiopheno-quinone (BDTQ)—has been discovered that contains even two thiophene rings (Fig. 1B) (Lanzotti et al., 1986).
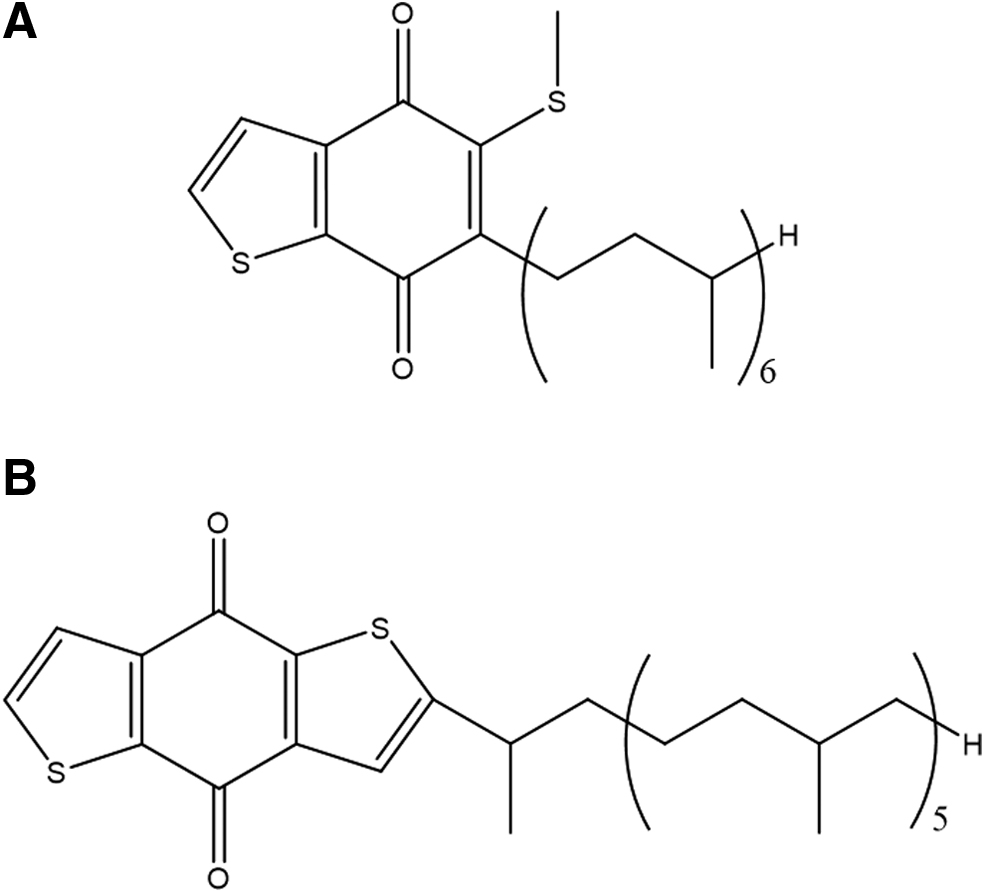
Chemical structures of Caldariellaquinone
CQ and BDTQ might fulfill respiratory functions like other quinones, for example, ubiquinone, and tyrosine might serve as a source material for their biosynthesis (Zhou and White, 1989). Thus, it can be hypothesized that the thiophenes found in natural environments (Table 1) could also be—at least in part—the metabolic fossils of archaebacterial metabolism (Juhl and Clark, 1990). It has been suggested that under severe reducing conditions of hypersaline sediments, CQ could degrade into smaller fragments, including thiophenes and benzothiophenes as detected on Mars (Eigenbrode et al., 2018), which could serve as biomarkers (Wang and Fu, 1997).
In addition, the biosynthesis of thiophene and some derivatives has been described for bacteria associated with fruiting bodies of the white truffle Tuber borchii (Splivallo et al., 2015). The truffle bacteria community was dominated by α- and β-Proteobacteria, whereas γ-Proteobacteria and Bacteroidetes were detected at much lower quantities. Interestingly, all of these phyla/classes—independent of their isolation source (truffle or other sources such as soil)—were able to produce thiophenes (Splivallo et al., 2015). Together with the biosynthesis of CQ and BDTQ, this indicates that microbial thiophene production might be a widespread metabolic pathway.
2.3. Biodegradation of thiophenes
Thiophenes can be degraded microbially by either sulfate-reducing or sulfur-oxidizing bacteria. One potential metabolic pathway for microbial sulfur oxidation is the formation of a sulfone followed by its degradation to sulfite (SO3 2−) and an alcohol (Fig. 2A). This pathway has been shown for various bacterial strains degrading, for example, dibenzothiophene (Kertesz, 2000; Gogoi and Bezbaruah, 2002). The sulfur atom in thiophene can also be reduced anaerobically by sulfate-reducing microbial communities to sulfide (Fig. 2B) (Marcelis et al., 2003). For example, dibenzothiophene can be degraded to biphenyl and H2S by Desulfovibrio desulfuricans (Kim et al., 1990).
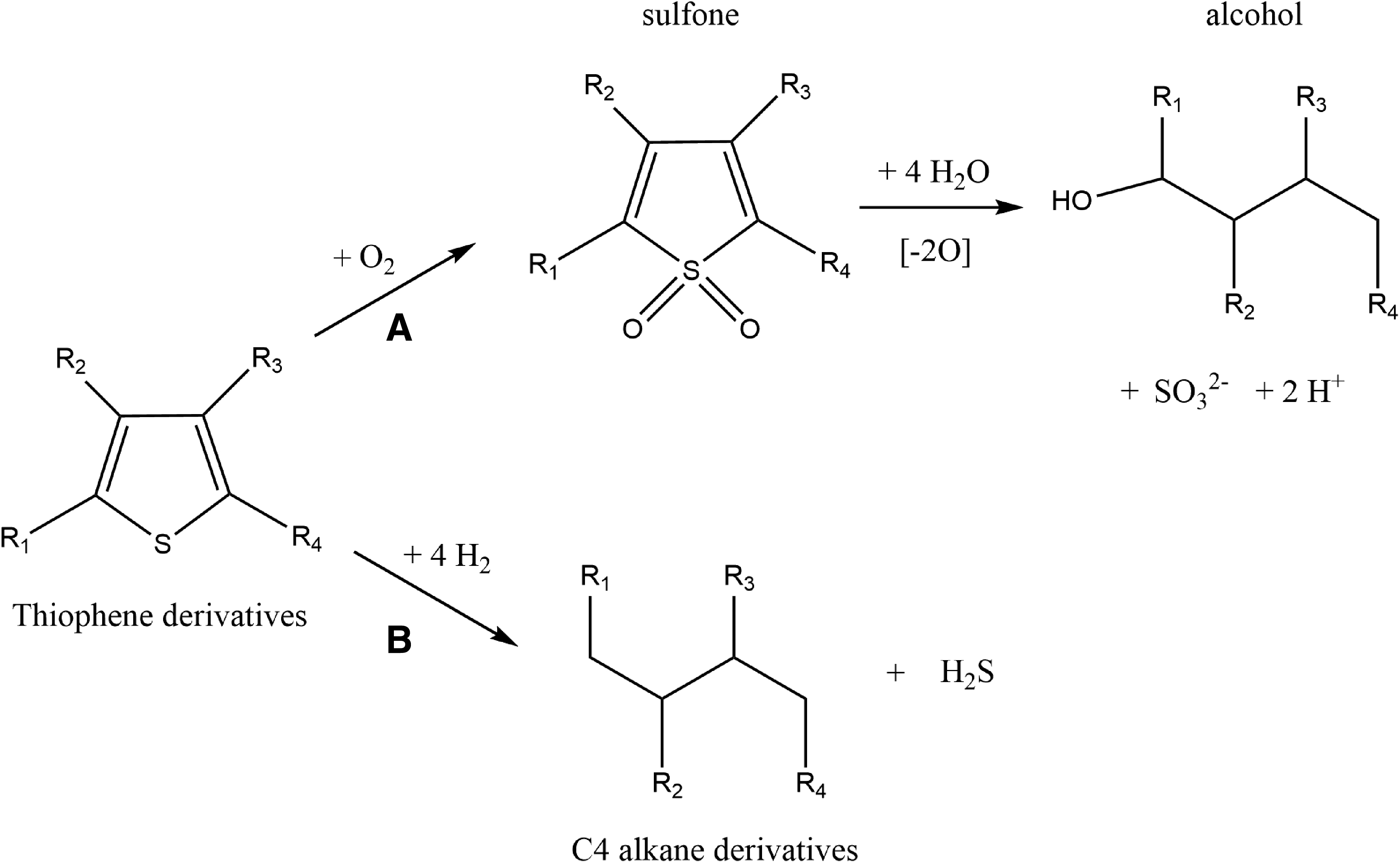
Biodegradation pathways through sulfur-oxidizing
3. Thiophenes on Mars
3.1. Detection of martian thiophenes and the correlation with sulfate occurrences
Thiophene and a few of its derivatives (2-methylthiophene, 3-methylthiophene, and presumably benzothiophene) (Fig. 3) have been detected on Mars together with smaller sulfur-containing substances (methanethiol, dimethyl sulfide, carbonyl sulfide, carbon disulfide, hydrogen sulfide, and sulfur dioxide) in lacustrine mudstone at Pahrump Hills in the lower Murray formation, Gale Crater, through EGA and pyrolysis-GC-MS onboard the Curiosity rover (Eigenbrode et al., 2018). The total sulfur content of the organic matter is with ∼5% similar to sulfur content in kerogens on Earth (Table 1). In addition, several sulfur-free aliphatic and aromatic compounds have been found. The concentrations of the detected components in the various drill holes reported by Eigenbrode et al. (2018) are summarized in Table 2.
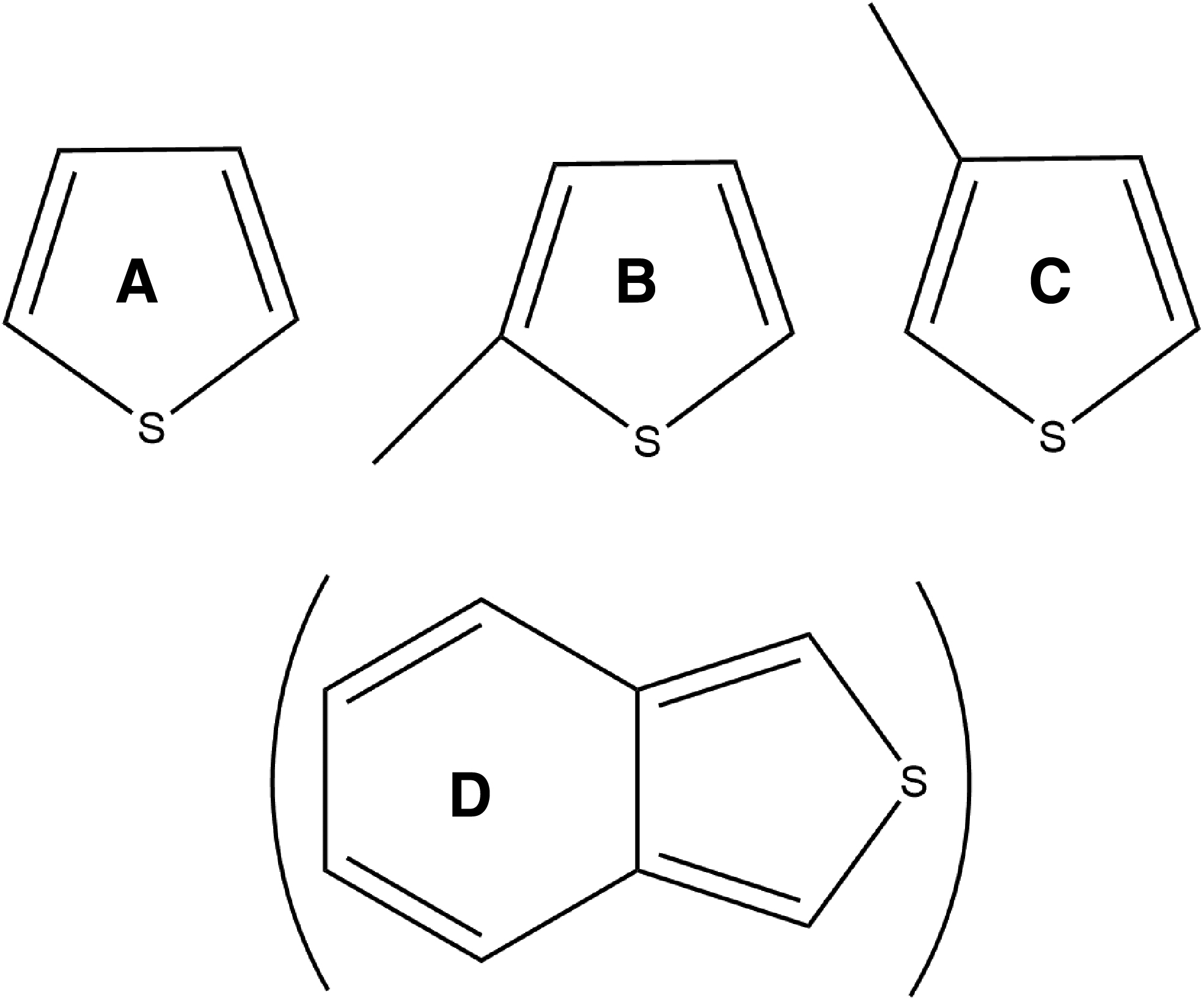
Thiophene derivatives detected on Mars (Eigenbrode et al., 2018): thiophene
Concentration of Chemical Compounds Detected Through Evolved Gas Analysis (in nmol of Carbon) (Eigenbrode et al., 2018)
Drill hole abbreviations: JK, John Klein; CB, Cumberland; CH, Confidence Hills; MJ, Mojave 2; TP, Telegraph Peak; BK, Buckskin. Blank values are printed in italic, total thiophene concentrations are printed in bold.
Average values of several measurements. For further information including error values, see Eigenbrode et al. (2018).
Table 2 reveals that compared with the blank measurements at Cumberland (CB blank, 0.46 nmol) and Confidence Hills (CH blank, 0.49 nmol), significant thiophene concentrations appeared only in the samples of CH (2.02 nmol) and Mojave 2 (MJ, 2.20 nmol). However, the slightly increased thiophene concentrations at John Klein (JK, 0.79 nmol), Telegraph Peak (TP, 0.70 nmol), and Buckskin (BK, 0.63 nmol) might represent positive thiophene detections close to the detection limit. In contrast, no thiophene seems to be present at CB because the measured thiophene concentration (0.26 nmol) is below those measured for the blanks.
The mineralogy of the different sites has already been described elsewhere (Grotzinger et al., 2014, 2015; Vaniman et al., 2014; Hurowitz et al., 2017; Rampe et al., 2017). Particularly interesting is a possible correlation between sulfate occurrences and thiophene detections (Fig. 4). MJ, the drill hole with the highest thiophene abundances (Table 2), is also the sample with the highest jarosite [KFe3(OH)6(SO4)2] concentration (3.1 wt %) yet discovered by the Curiosity rover (Martin et al., 2017). In addition, magnesium sulfate (MgSO4) has been detected at both thiophene-rich sites (MJ and CH) (Nachon et al., 2017), whereas the correlation between the MgO and SO3 content was significantly lower at TP and BK, indicating the absence or only low concentrations of MgSO4 (Hurowitz et al., 2017). Furthermore, it has been proposed that MJ and CH are situated in a section of the stratigraphic column of the former Gale Crater lake where sulfate precipitation from leached minerals occurred (Rampe et al., 2017), also correlating with the enhanced thiophene abundances in that section.
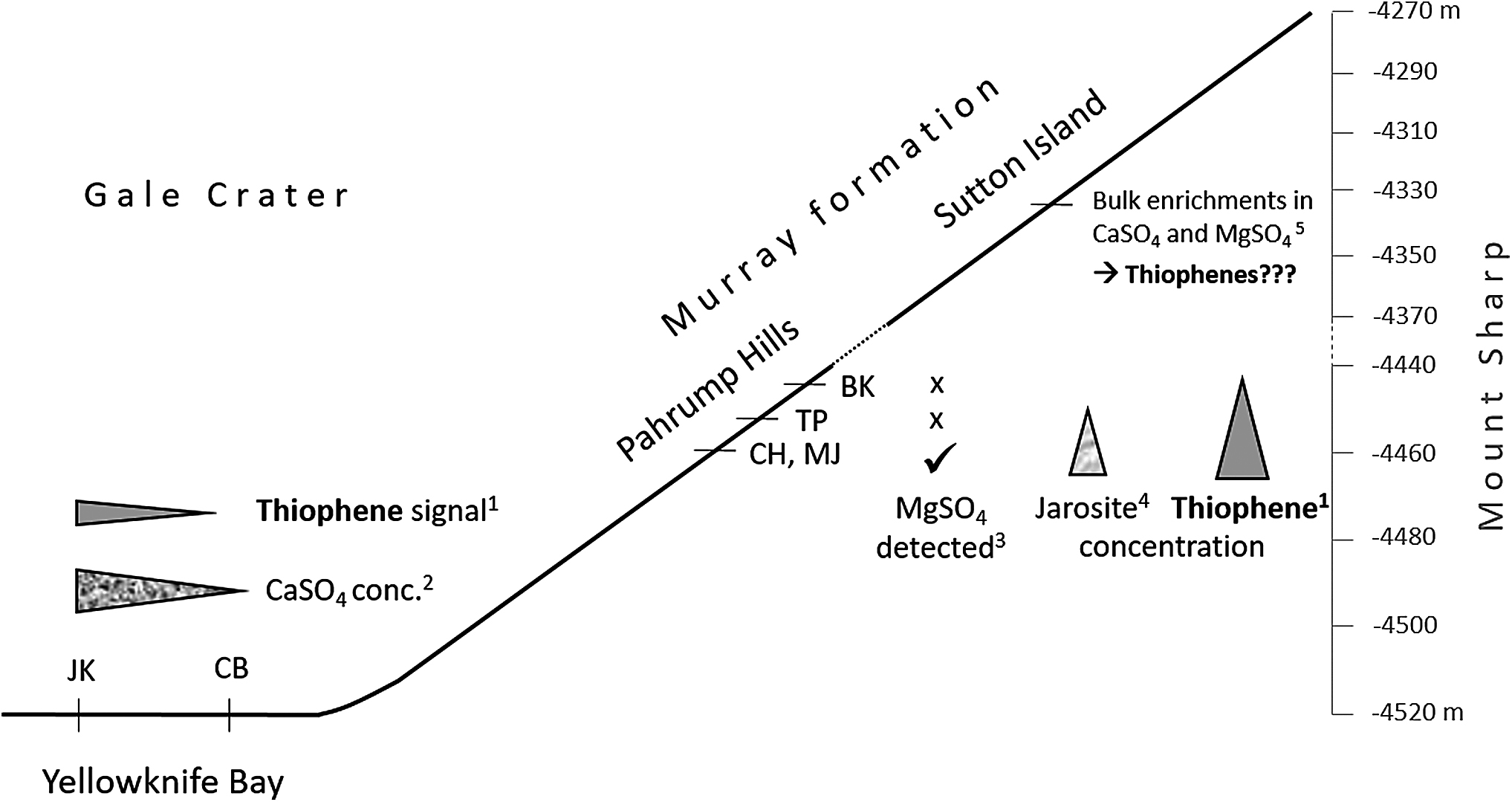
Stratigraphic context in the Gale Crater on the correlation between thiophene and sulfate abundances in mudstones (Yellowknife Bay and Pahrump Hills) and bedrock (Sutton Island). References: 1Eigenbrode et al. (2018), 2Vaniman et al. (2014), 3Hurowitz et al. (2017), 4Rampe et al. (2017), 5Rapin et al. (2019).
At CB and JK, neither jarosite nor MgSO4 [no MgO/SO3 correlation (Hurowitz et al., 2017)] has been detected, accompanying no or a low thiophene signal, respectively. However, CaSO4 veins appear in the mudstone of the Yellowknife Bay like on various other places along the Curiosity route (Nachon et al., 2017). Nevertheless, the CaSO4 concentration at CB is very low [0.8 wt % anhydrite, 0.7 wt % bassanite (Vaniman et al., 2014)], possibly correlating with the absence of thiophenes. The CaSO4 concentration at JK is slightly increased [2.6 wt % anhydrite, 1.9 wt % bassanite (Vaniman et al., 2014)] and CaSO4 veins are visible in the drill hole (Nachon et al., 2014), correlating with a slightly increased thiophene signal for JK compared with CB (Table 2).
There are several possible explanations for the observed correlation between thiophene and sulfate abundances:
The correlation might be simply the result of sulfur enrichments. An increased content of sulfur nucleophiles would enhance both the formation rate of thiophenes during diagenesis and the sulfate formation through oxidation processes (Eglinton et al., 1992; Sinninghe Damsté and Leeuw, 1992).
Sulfates are known to preserve organic matter (Aubrey et al., 2006). Hence, the detected sulfate minerals might increase the stability of the thiophenes that formed earlier in the martian history.
Sulfate might be a parent substance of the thiophene formation. A possible biological conversion of sulfates into thiophenes is described in Section 3.2.3.
Independent of the reason for the thiophene/sulfate correlation, we hypothesize that in sediments at Sutton Island (Fig. 4), where the Curiosity rover very recently discovered bulk enrichments of CaSO4 (30–50 wt %) and MgSO4 (26–36 wt %) (Rapin et al., 2019), the thiophene concentration might be even higher than at Pahrump Hills. It should be noted that the thiophene/sulfate correlation reported in this study is only semiquantitative, referring on several specific sulfate-bearing minerals, because the total sulfate concentration (including soluble salts such as MgSO4) is unknown. Correlations with other geochemical parameters have not been found.
3.2. Possible origins of thiophene derivatives on Mars
3.2.1. Meteoric input
Thiophene and several of its derivatives such as benzothiophene and dibenzothiophene have been detected in various meteorites, for example, the carbonaceous chondrites in Murchison (Shimoyama and Katsumata, 2001; Sephton, 2004), Murray (Hayes and Biemann, 1968; Sephton, 2002), and Orgueil (Bandurski and Nagy, 1976). Thus, it is plausible to assume that the thiophene derivatives found on Mars are at least partly the result of meteoric impacts. In addition, these cosmic events could provide organic compounds that can react with sulfurization agents to yield thiophene derivatives (Section 3.2.2.).
3.2.2. Abiotic thiophene synthesis on Mars
Similar to the formation of thiophenic compounds on Earth (Section 2.1.), it is conceivable that the sulfur-rich organic matter on Mars formed during early diagenesis by reaction of sulfur nucleophiles with organic matter. The sulfur nucleotides might have been transported in the form of sulfides through hydrothermal groundwater to the Gale crater lake (Franz et al., 2017) or might be biological in origin (see Section 3.2.3). The organic substances might be of cosmic origin (infall of extraterrestrial carbonaceous material), a product of abiotic organic synthesis on igneous rocks (Steele et al., 2016), or even of biological origin. Indeed, in the martian Sheepbed mudstone, organic molecules with a four-carbon chain (C4) have been detected (Freissinet et al. 2015). C4 compounds can react with sulfurizing agents such as elemental sulfur, hydrogen sulfide, sulfur oxides, or metal sulfides in a ring-closing condensation reaction to thiophene or its derivatives (Campaigne, 1984).
It is very likely that during diagenesis, the formed thiophenes are directly incorporated into kerogen-like macromolecular structures, which would be more stable than the single molecules. A hypothetical macromolecular structure representing the organic molecules detected at MJ and CH is presented in Fig. 5. These large structures would break up during pyrolysis and would provide the fragments detected through EGA and GC-MS (Eigenbrode et al., 2018).
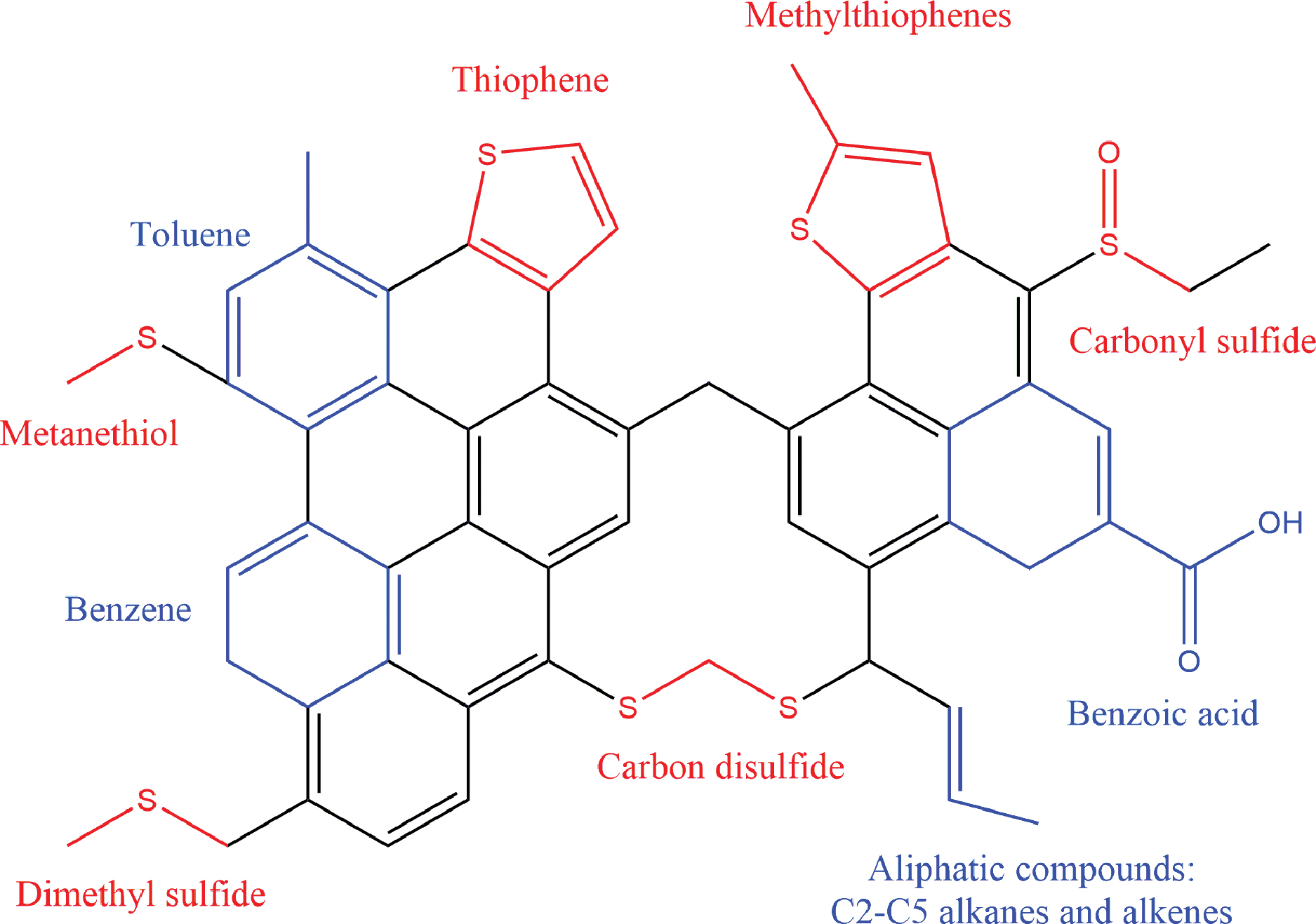
Hypothetical macromolecular structure of the organic material detected at Pahrump Hills. Sulfur-containing and sulfur-free structures detected through evolved gas analysis are shown in red and in blue, respectively. Color images are available online.
3.2.3. Biosynthesis and thiophenes as biomarkers on Mars
Nucleophilic sulfur species are needed for the diagenetic thiophene synthesis described previously. However, the majority of available sulfur on Mars exists in the form of non-nucleophilic sulfates (King and McLennan, 2010). Sulfate can be reduced into nucleophilic sulfide species by thermochemical sulfate reduction at temperatures >120°C (Goldstein and Aizenshtat, 1994). A possibility to transform sulfates into sulfides also at low temperatures is given by bacterial sulfate reduction (BSR).
For example, the mineral jarosite detected on Mars by the Curiosity rover (e.g., at CH and MJ, Fig. 4) as well as the Opportunity rover (Christensen et al., 2004) could have been converted to H2S through reductive dissolution by sulfate-reducing bacteria (Gao et al., 2019). The formation of jarosite requires acid environmental conditions (pH 2–6) and possibly occurred on Mars by postdepositional sediment reactions with an acidic oxidizing fluid flowing through the Pahrump Hills area (Rampe et al., 2017). Additional moderately acid conditions on Mars have been proposed accompanying the lack of large carbonate deposits (Fairén et al., 2004).
BSR under these acid conditions is possible, although it is more common in natural environments with pH >6 (Koschorreck, 2008). An example for the occurrence of BSR in a natural acidic environment is the Río Tinto in the Iberian Pyritic Belt (Amils et al., 2007; Sánchez-Andrea et al. 2011). Nevertheless, the environmental conditions at the thiophene-bearing sites could not have been extremely acid for a long period, because otherwise the detected organic compounds (and some pH-sensitive minerals such as olivine or apatite) would have been oxidized or destroyed (Eigenbrode et al., 2018).
Furthermore, BSR is feasible at subzero temperature prevailing on Mars (Knoblauch and Jorgensen, 1999). Therefore, BSR has been suggested as a plausible metabolism pathway on ancient Mars and in the subsurface of Mars today (Marnocha et al., 2010). The sulfide formed through BSR of martian sulfates could have been triggering the diagenetic thiophene production on ancient Mars and could help to explain the observed correlation between sulfate and thiophene abundances (Fig. 4).
Especially the detection of isoprenoid thiophenes would be of huge astrobiological interest because it would hint either toward the biotic synthesis of the precursors such as phytenes (Section 2.1.) or toward their direct biosynthesis similar to the formation of CQ or BDTQ (Section 2.2.). In both cases, they could be considered as martian biomarkers. If these isoprenoid thiophenes were present in the Pahrump Hill samples, the heating during pyrolysis (500–820°C for EGA) would have destroyed these more fragile compounds and revealed smaller fragments with a higher thermal stability, like the detected methyl thiophenes and alkanes and alkenes of various chain lengths.
3.2.4. Biodegradation of thiophenes on Mars
Theoretically, the thiophenic compounds—independent of their formation pathway—could also be biodegraded by putative heterotrophic martian microorganisms as described in Section 2.3. In this case, the anaerobic degradation of thiophenes by sulfate-reducing bacteria seems to be a more realistic scenario for the predominantly anaerobic conditions on Mars, especially since this type of bacteria could drive the thiophene production in the first place (Section 3.2.3.). It could even be speculated that the C4 dichloroalkanes detected on Mars (Freissinet et al. 2015) might be the result of anaerobic degradation of chlorinated thiophenes following the equation from Fig. 2B with two of the residues (R1–R4) substituted by chlorine.
4. Discussion and Recommendations
We briefly summarized conceivable production and degradation pathways of thiophenes detected on Mars by the Curiosity rover and evaluated their potential in serving as biomarkers. With the available data, none of the presented thiophene formation pathways can be ruled out. Certainly, the biosynthesis of thiophenes is the most fascinating option; however, more detailed analyses are needed to support or discard this possibility.
In future Mars missions, more structurally sensitive (less destructive) instrumentation is needed since the pyrolysis-based techniques used by the Curiosity rover provide only information on fragments of organic matter. The Mars Organic Molecule Analyzer onboard the Rosalind Franklin rover (expected to be launched in July 2020) will provide the necessary nondestructive analyzing method by using laser desorption mass spectrometry to characterize organic molecules and putative biomarkers (Goesmann et al., 2017).
Isotopic analyses of the martian organic matter would be helpful to distinguish an abiotic origin from a biotic origin. Biotically produced organic compounds are expected to be enriched in lighter isotopes, for example, lower 13C/12C ratios, compared with organic compounds formed abiotically (Schidlowski, 1992; Schulze-Makuch and Irwin, 2018). Although a decreased 13C/12C ratio alone cannot provide a definite proof for the presence of martian life, carbon isotopic data would nevertheless be very useful in determining prebiotic carbon transformations on Mars (Rothschild and DesMarais, 1989).
Sulfur isotopic ratios may be even more revealing, especially when comparing thiophenes produced by biota with abiotically deposited sulfur sources such as sulfate minerals (van Zuilen, 2008). In contrast to carbon isotopic analyses that are lacking for martian samples to date, in situ analyses of the sulfur isotopic compositions of SO2 volatilized from sediment samples have been carried out recently by the Curiosity rover (Franz et al., 2017). The experiments revealed large sulfur isotope fractionations (δ34S) ranging from −47 ± 14‰ to +28 ± 7‰. Interestingly, this range closely reflects observed isotopic fractionations between sulfide and sulfate (−42 to +28‰) of the Haughton impact structure in the Canadian High Arctic, which is interpreted to result from biological sulfate reduction in an impact-generated hydrothermal system (Parnell et al., 2010; Franz et al., 2017). Certainly, this is no proof that martian sulfur compounds (including thiophenes) are of biological origin but should prompt further investigations.
One of these investigations could be laboratory experiments to explore the possible biotic production of thiophenes under simulated martian conditions. Microbial growth and production of thiophenes may be possible under martian environmental niche conditions (near subsurface), if sufficient adaptation time is provided (de Vera et al., 2014). Furthermore, it can be assumed that microbial thiophene production occurred only on early Mars with its more habitable conditions. Thus, laboratory experiments should also be conducted for this early Mars scenario that might include increased pressures, warmer but still subzero temperatures, as well as saline and slightly acidic liquid water (Fairén et al., 2009). If microbial thiophene production could successfully be simulated under these conditions in the laboratory, also the isotopic fractionation in source materials, metabolic intermediates, and metabolic end products should be determined to evaluate experimental data obtained from Mars.
Footnotes
Acknowledgments
We thank the reviewers Alberto Fairén and Chris McKay for constructive reviews, which were detailed and included many helpful suggestions.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This project was funded by European Research Council Advanced Grant “Habitability of Martian Environments” (HOME, No. 339231).