
Abstract
The plant cell wall provides each cell with structural support and mechanical strength, and thus, it plays an important role in supporting the plant body against the gravitational force. We investigated the effects of microgravity on the composition of cell wall polysaccharides and on the expression levels of genes involved in cell wall metabolism using rice shoots cultivated under artificial 1 g and microgravity conditions on the International Space Station. The bulk amount of the cell wall obtained from microgravity-grown shoots was comparable with that from 1 g-grown shoots. However, the analysis of sugar constituents of matrix polysaccharides showed that microgravity specifically reduced the amount of glucose (Glc)-containing polysaccharides such as 1,3:1,4-β-glucans, in shoot cell walls. The expression level of a gene for endo-1,3:1,4-β-glucanase, which hydrolyzes 1,3:1,4-β-glucans, largely increased under microgravity conditions. However, the expression levels of genes involved in the biosynthesis of 1,3:1,4-β-glucans were almost the same under both gravity conditions. On the contrary, microgravity scarcely affected the level and the metabolism of arabinoxylans. These results suggest that a microgravity environment promotes the breakdown of 1,3:1,4-β-glucans, which, in turn, causes the reduced level of these polysaccharides in growing rice shoots. Changes in 1,3:1,4-β-glucan level may be involved in the modification of mechanical properties of cell walls under microgravity conditions in space.
1. Introduction
After ancestral plants emerged on land around 450 million years ago, they were directly exposed to a gravitational force of 1 g. So, terrestrial plants have evolved in the presence of gravity and developed the cell wall structure to support and stand their body upright against the gravitational force at 1 g, as do the bones and muscles in an animal body (Hoson and Soga, 2003; Volkmann and Baluska, 2006). Plant cell walls surround each protoplast and provide cells with structural rigidity. So they are a major source of mechanical strength for plant bodies. Furthermore, plant cell walls directly determine the size and shape of plant cells. Thus, the cell wall plays an important role in the regulation of growth and morphogenesis in plants (Cosgrove, 2000; Hoson, 2002).
Plant cell walls consist of cellulose, a variety of matrix polysaccharides, proteins, and phenolic substances, and they are divided into two groups, type I wall and type II wall, based on the differences in matrix polysaccharide composition (Carpita, 1996). Gramineous (cereal) plants such as rice, wheat, and maize have the type II wall, while other monocotyledons and dicotyledons have the type I wall (Carpita, 1996). In gramineous cell wall (type II wall), 1,3:1,4-β-glucans and (glucurono)arabinoxylans are the major matrix polysaccharides (Carpita, 1996). 1,3:1,4-β-Glucans consist of a backbone of unbranched (1,3)- and (1,4)-linked β-glucosyl residues with high-molecular mass and they may tether cellulose microfibrils (Carpita and Gibeaut, 1993). On the contrary, arabinoxylans consist of xylose (Xyl), arabinose (Ara), and a small quantity of glucuronic acid (GlcA) residues. In addition, arabinoxylans possess phenolic acid (hydroxycinnamate) monomers such as ferulic acid (FA) and p-coumaric acid, which are ester-linked to Ara residues of arabinoxylans (Smith and Hartley, 1983; Saulnier and Thibault, 1999). Of these phenolic acids, FA undergoes a peroxidase-catalyzed coupling reaction to produce diferulic acid (DFA), which crosslinks arabinoxylans (Fry, 1986; Grabber et al., 1995; Saulnier et al., 1999). Arabinoxylans are considered to form intranetwork structure via phenolic acid-mediated cross-linkages within type II wall. Therefore, quantities of 1,3:1,4-β-glucans and arabinoxylans may be involved in the regulation of mechanical strength of type II wall.
We carried out the space experiment termed Ferulate using the Cell Biology Experiment Facility (CBEF) in the “Kibo” module of the International Space Station (ISS) to clarify the effect of microgravity environment on the formation of phenolic acid-mediated intranetwork structure in type II wall. The Ferulate experiment showed that cell walls of microgravity-grown rice shoots were loosened compared with those of artificial 1 g-grown ones (Wakabayashi et al., 2015). Similarly, a former space experiment showed that the cell wall extensibility of coleoptiles obtained from microgravity-grown rice seedlings was higher than that from ground-grown ones (Hoson et al., 2002, 2004). Furthermore, we observed that the microgravity environment affected the metabolism of cell wall-bound phenolics, especially it suppressed the DFA formation in shoot cell walls (Wakabayashi et al., 2015). Therefore, the decrease in DFA-mediated cross-linkages within the cell wall architecture may contribute to keeping the cell wall loosened under microgravity conditions. On the contrary, the effect of microgravity environment on the levels of cell wall polysaccharides, particularly matrix polysaccharides, has not been analyzed in detail. The former space experiment showed that in cell walls of rice coleoptiles, the relative content of the high-molecular-mass matrix polysaccharides decreased under microgravity conditions (Hoson et al., 2002, 2004), suggesting a possibility that microgravity modifies the metabolism of certain matrix polysaccharides, in addition to the DFA formation, in gramineous plants. The present study aims to clarify the levels of matrix polysaccharides, in particular those of 1,3:1,4-β-glucans and arabinoxylans, and also the expression levels of genes involved in their metabolism using rice shoots that had been grown under microgravity and artificial 1 g conditions in space.
2. Materials and Methods
2.1. Plant materials and onboard procedures
Three days before the launch, rice (Oryza sativa L., cv. Koshihikari) seeds (caryopses) were sterilized in 5% (v/v) sodium hypochlorite solution for 1 h and then thoroughly washed with sterilized water. Twenty-five sterilized seeds were aligned along a groove on 11 mL of 1% (w/v) agar (Bacto Agar; BD) in a black polycarbonate culture dish (W × D × H = 40 × 40 × 30 mm, outer dimensions) that shaded seeds from light irradiation (Fig. 1A, B). After planting, culture dishes were stored in a measurement experiment unit (MEU) (Fig. 1C) and kept at 2–4°C before and during the flight, until the start of the growth experiment on the ISS. We prepared 4 MEUs, each of which contained 12 culture dishes (Fig. 1C).
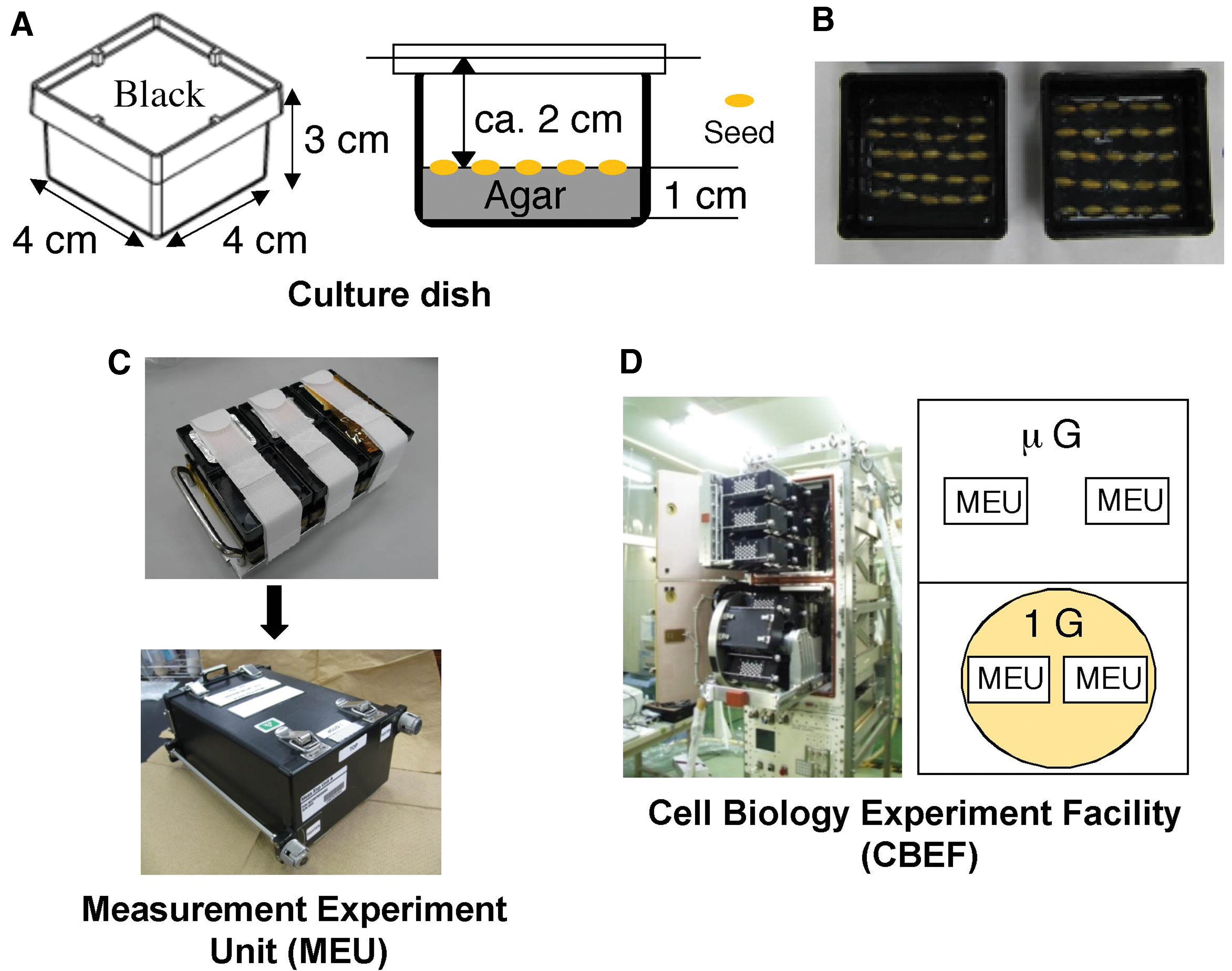
Experimental equipments used in Ferulate experiment.
The MEUs containing rice seeds were launched aboard the Space Shuttle STS-132 flight. In orbit, the MEUs were stored in the Minus-Eighty Laboratory Freezer for ISS (MELFI) for 12 days at 2°C. Then, the MEUs were transferred to the CBEF (Fig. 1D). The CBEF is an incubator with two compartments: an artificial gravity compartment has a centrifuge to produce artificial 1 g conditions and another compartment gives microgravity conditions (Yano et al., 2012). The CBEF has a merit to cultivate plants under microgravity and artificial 1 g conditions simultaneously in space. Two MEUs were placed in the microgravity compartment of the CBEF and two in the artificial gravity compartment. The temperature inside the CBEF was raised under microgravity conditions in both compartments and rotation in the artificial gravity compartment was started when the temperature reached 20°C. The temperature of culture dishes increased at the same rate in all MEUs, reaching 22.6°C 180 min after being transferred to the CBEF. The seeds were germinated and grown in darkness for 99 h (4 days), 127 h (5.3 days), or 136 h (5.6 days) at 22.6°C ± 0.1°C. At the end of each cultivation period, the culture dishes were immediately transferred to the MELFI and stored at −80°C. The samples were brought back to earth aboard the Space Shuttle STS-133 flight and transported to the laboratory at Osaka City University. Plant samples were kept frozen during the flight and transportation to the laboratory and were stored at −80°C until the analysis.
We initially planned to harvest plant materials at 1-day intervals from days 4 to 6, but operational limitations in orbit constrained sampling to days 4, 5.3, and 5.6. Because the gap between 5.3 and 5.6 days was short and the available samples were limited, we used 5.3-day shoots for the chemical analysis of cell wall polysaccharides, and 4- and 5.6-day shoots for the analysis of gene expression.
2.2. Fractionation of cell walls
Shoots consisting of the coleoptile and first leaf were excised from the frozen seedlings. Excised shoots were immediately dipped in 80% (v/v) ethanol and then boiled for 10 min. The ethanol-fixed shoots were rehydrated with water and their lengths were measured with a ruler. After that, the cell wall polysaccharides were fractionated according to the method of Wakabayashi et al. (1997). Rehydrated shoots were homogenized in water with a mortar and a pestle, washed with water, acetone, and a methanol:chloroform mixture (1:1, v/v), and then treated with 2 units/mL porcine pancreatic α-amylase (type I-A; Sigma, St. Louis, MO) in 50 mM sodium acetate buffer (pH 6.5) at 37°C for 3 h to remove starch. After the amylase treatment, the cell walls were extracted three times (8 h each) with 17.5% NaOH containing 0.02% NaBH4 at room temperature. The fraction extracted with 17.5% NaOH was neutralized with acetic acid and then dialyzed against water for 24 h at 4°C. This fraction was designated as the matrix polysaccharide fraction. The alkali-insoluble fraction (cellulose fraction) was washed successively with 0.03 M acetic acid and ethanol, and dried at 40°C. The cellulose fraction was dissolved in 72% sulfuric acid for 1 h at room temperature and then diluted with a 29-fold volume of water.
2.3. Determination of sugar content
The total sugar content of each cell wall fraction was determined by the phenol/sulfuric acid method (Dubois et al., 1956) using Glc as the standard. The sugar composition of matrix polysaccharide fraction was determined according to the method of Wakabayashi et al. (1997) and Kimpara et al. (2008). A portion of the matrix polysaccharide fraction was lyophilized and then hydrolyzed with 2 M trifluoroacetic acid for 1 h at 121°C. After the trifluoroacetic acid had been removed by evaporation under the stream air at 50°C, sugars derived from matrix polysaccharides were analyzed by high-performance anion-exchange chromatography with pulsed amperometric detection using a Dionex DX-500 liquid chromatography (Dionex, Sunnyvale, CA) fitted with a CarboPac PA-1 column (4 × 150 mm) and a pulsed amperometric detector. The amounts of sugars were determined using authentic monosaccharides (Sigma).
2.4. Quantifying gene expression levels
After excising, shoots were immediately frozen with liquid nitrogen and kept at −80°C until analysis. Total RNA was extracted from frozen shoots using the RNeasy Plant Mini Kit (Qiagen, Valencia, CA), including a DNA elimination step (RNase-Free DNase Set; Qiagen). Complementary DNA (cDNA) was then synthesized from the RNA sample, and the Cyanine 3 (Cy3)-labeled RNA was synthesized with Cy3-CTP using the Low Input Quick Amp Labeling Kit (Agilent Technologies, Santa Clara, CA). The Agilent Rice Oligo Microarray 4 × 44 K slides (Design ID 015058; Agilent Technologies) were used for microarray analysis. The labeled RNA was purified, fragmented, and then hybridized onto the microarray slides according to the manufacturer's instructions. Hybridization signals (fluorescence intensities) on the arrays were analyzed with a feature extraction software (Agilent Technologies). Microarray analysis was conducted on four independent samples prepared from four culture dishes. The expression levels of genes involved in cell wall metabolism were analyzed using the values of fluorescence intensities obtained through a microarray analysis, and the information was obtained mainly from the Oryzabase (
2.5. Statistical analysis
For each measurement, the means and the standard errors of the means were calculated. The significance of differences between artificial (on-orbit) 1 g and microgravity conditions was analyzed using Student's paired t test.
3. Results
Rice seedlings grown in orbit for 5.3 days under artificial 1 g and microgravity conditions are shown in Fig. 2. Roots grew in various directions with some emerging into the air under microgravity conditions, while they elongated downward along the gravity vector under artificial 1 g conditions, indicating that the artificial gravity in orbit mimics the 1 g condition on the ground. On the contrary, the length of rice shoots at 5.3 days was almost the same under both conditions (artificial 1 g conditions: 12.8 ± 0.2 mm; microgravity conditions: 12.9 ± 0.3 mm; n = 12–15).

Rice seedlings grown for 5.3 days under artificial (on-orbit) 1 g and microgravity conditions in orbit. Color images are available online.
Cell walls of young expanding tissues are mainly composed of cellulose and a variety of matrix polysaccharides. The cell wall polysaccharides of rice shoots were fractionated into the matrix polysaccharide and cellulose fractions using strong alkali solutions and the sugar contents of both fractions were determined. At 5.3 days, the contents in artificial 1 g and microgravity-grown shoots were 73.8 ± 2.5 and 78.5 ± 2.8 μg/shoot for cellulose (n = 3) and 95.1 ± 0.4 and 91.2 ± 2.5 μg/shoot for matrix polysaccharides (n = 3), indicating that the microgravity environment had little effect on the mass amounts of matrix polysaccharides and cellulose. Next, the effect of microgravity on the composition of matrix polysaccharide fraction was examined. The major sugar constituents in the fraction were Glc, Xyl, and Ara (Fig. 3). The galactose (Gal) content was smaller than the major constituents, and fucose (Fuc), rhamnose (Rha), galacturonic acid (GalA), and GlcA were minor constituents in the fraction. The amount of Glc per shoot in microgravity-grown shoots was significantly lower than that in artificial 1 g-grown ones, while other constituents did not show significant differences between two conditions (Fig. 3). These results suggest that the microgravity environment specifically reduced the amount of Glc-containing polysaccharides, such as 1,3:1,4-β-glucans, in the matrix polysaccharides.
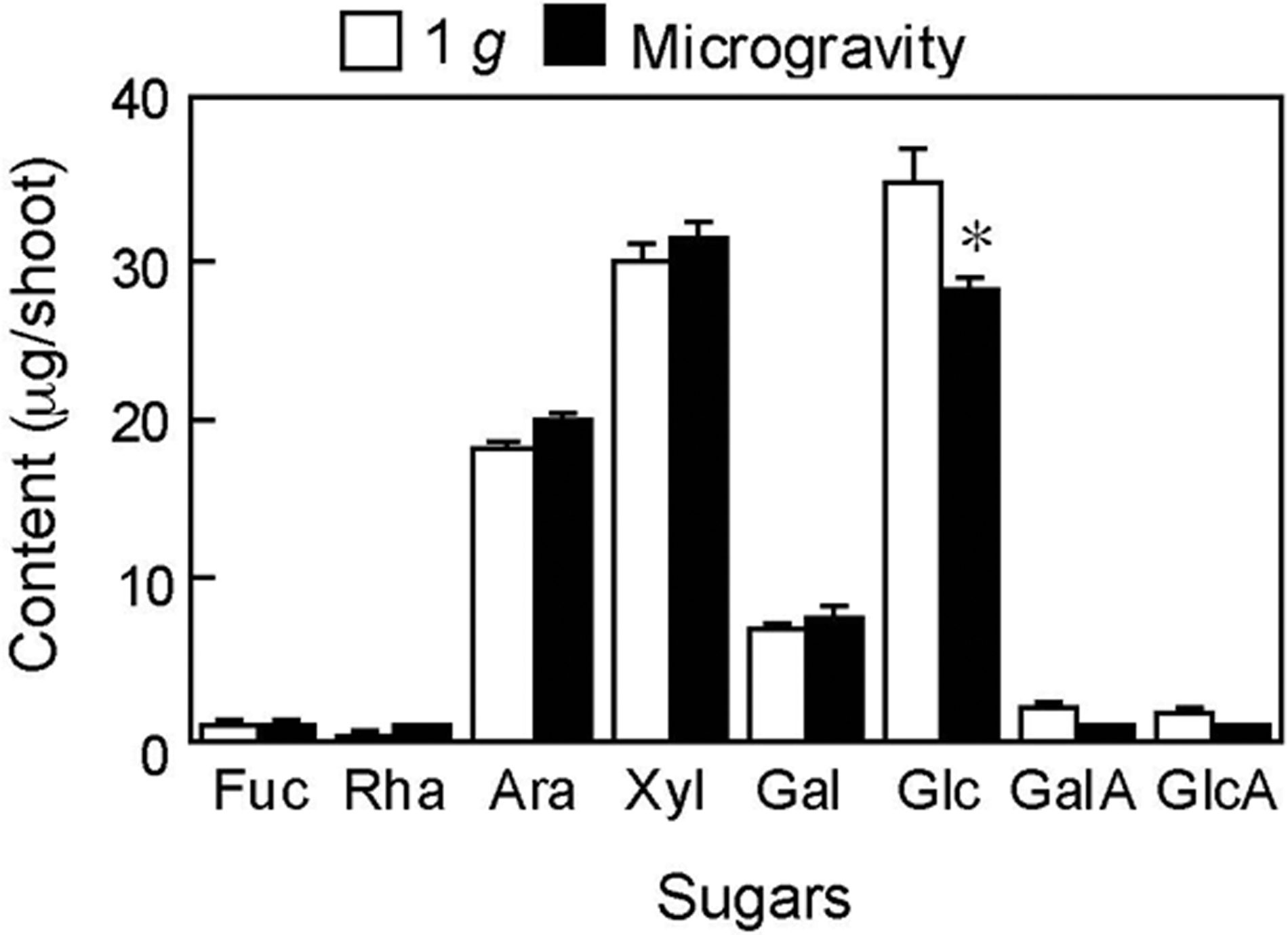
Amounts of sugar constituents in the matrix polysaccharide fraction extracted from the cell walls of rice shoots grown under artificial 1 g and microgravity conditions in orbit for 5.3 days. Sugar constituents were analyzed by the HPAEC-PAD on an anion-exchange column. Data are mean ± SE from three independent samples. *Mean values significantly different between artificial 1 g and microgravity conditions (Student's t test, p < 0.05). Ara, arabinose; Fuc, fucose; Gal, galactose; GalA, galacturonic acid; Glc, glucose; GlcA, glucuronic acid; HPAEC-PAD, high-performance anion-exchange chromatography with pulsed amperometric detection; Rha, rhamnose; SE, standard error; Xyl, xylose.
The processes of both biosynthesis and breakdown were considered as these may be involved in the regulation of the levels of cell wall constituents. We next analyzed expression levels of genes involved in the biosynthesis and breakdown of 1,3:1,4-β-glucans and arabinoxylans in rice shoots using microarray analysis data. In the present analysis, about 34,000 genes represented on the arrays were found to be expressed in shoots grown for 4 and 5.6 days under artificial 1 g and microgravity conditions. A total of 174 and 838 genes were upregulated more than twofold, while 97 and 206 genes were downregulated <0.5-fold under microgravity conditions at 4 and 5.6 days, respectively. Table 1 shows the expression levels of the Cellulose synthase-like (CslF and CslH) genes, which have been shown to mediate the biosynthesis of 1,3:1,4-β-glucans (Burton et al., 2006; Doblin et al., 2009). In the present experiment, genes with a fluorescence intensity of <100 were eliminated. Among the CslF and CslH genes, CslF6 and CslH2 showed strong signal intensity and their expression levels were almost constant during the cultivation period. Although microgravity slightly increased the expression levels of CslF6, CslH1, and CslH2 genes on 5.6 days, it barely affected those on 4 days and also the expression levels of other CslF genes on either 4 or 5.6 days (Table 1).
Expression Levels of Genes for 1,3:1,4-β–Glucan Synthesis (CslF and CslH) in Rice Shoots Grown Under Artificial 1 g and Microgravity Conditions in Orbit
The expression levels of genes were analyzed using the values of fluorescence intensities obtained through a microarray analysis. Genes with a fluorescence intensity of more than 100 are shown. Data are mean ± SE from four independent samples.
Mean values significantly different between artificial 1 g and microgravity conditions (Student's t test, p < 0.05). Figures in parentheses are values of the fold change in signal intensity of microgravity-grown shoots over artificial 1 g-grown ones.
SE = standard error.
Two genes (OsEGL1 and OsEGL2) encoding endo-1,3:1,4-β-glucanase (EC 3.2.1.73), which specifically hydrolyze 1,3:1,4-β-glucans, are present in rice plants (Akiyama et al., 2009). In the resent study, OsEGL1 and OsEGL2 were expressed in rice shoots and their levels increased during the cultivation period (Fig. 4). The expression levels of OsEGL1 in shoots grown under microgravity conditions were higher than those grown under artificial 1 g conditions on both 4 and 5.6 days, while those of OsEGL2 were almost the same between the two treatments (Fig. 4).
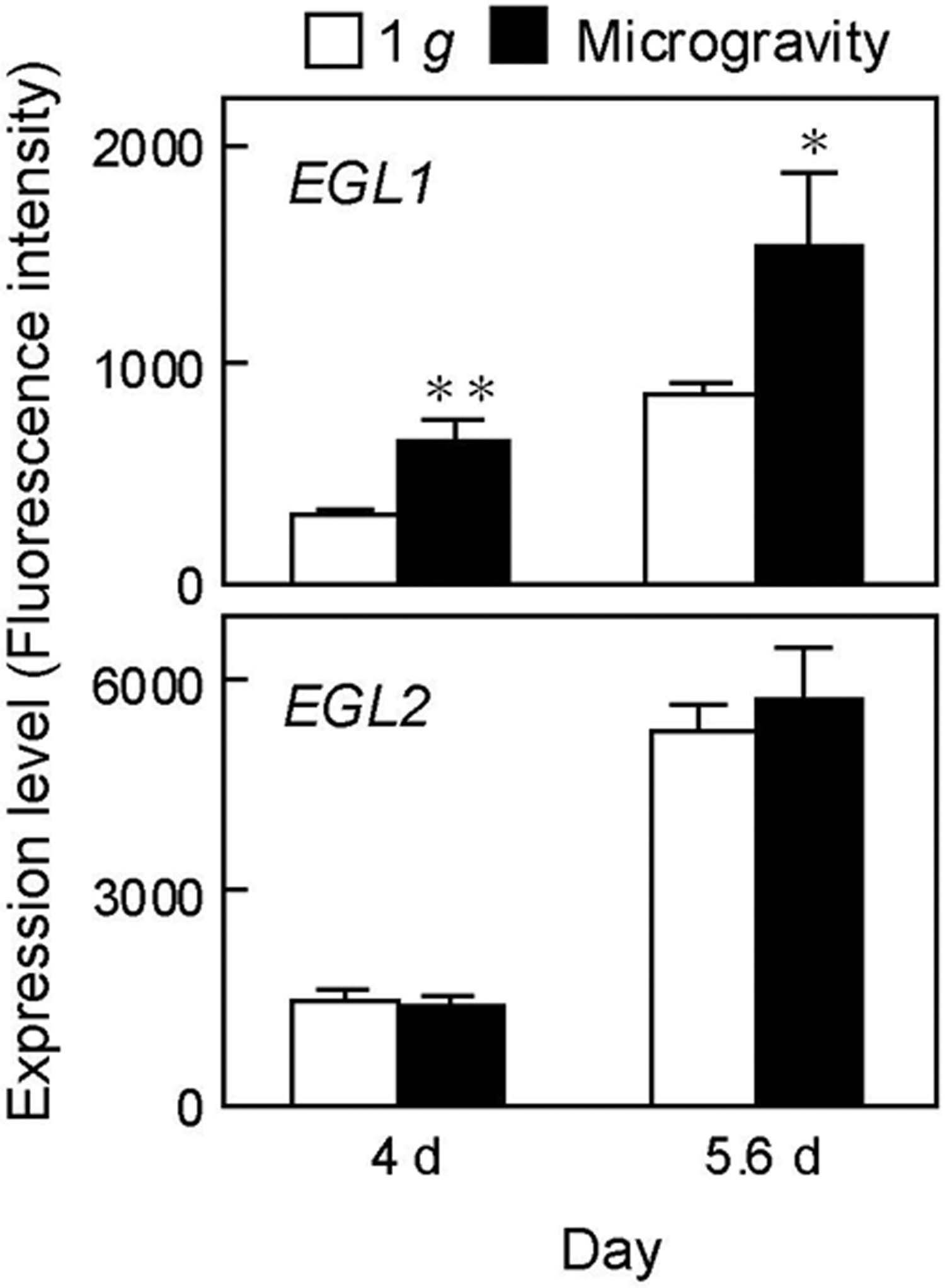
Expression levels of endo-1,3:1,4-β-glucanase genes (OsEGL1 and OsEGL2) in rice shoots grown under artificial 1 g and microgravity conditions in orbit. The expression levels of genes were analyzed using the values of fluorescence intensities obtained through a microarray analysis. Data are mean ± SE from four independent samples. Mean values significantly different between artificial 1 g and microgravity conditions at **p < 0.05 and *p < 0.1, respectively (Student's t test).
It has been shown that the synthesis of xylan backbone is a preferential process for the biosynthesis of arabinoxylans and glycosyl transferases (GT), particularly GT43 and GT47, involved in the xylan backbone synthesis (Hirano et al., 2013). Most of the genes in GT43 and GT47 families showed constant expression levels during the cultivation period, and microgravity slightly modified the expression levels of some GT43 and GT47 genes on 5.6 days (Table 2). On the contrary, glycosyl hydrolases (GH), especially xylanase (GH10) and α-L-arabiosidase or β–D-xylosidase (GH51), seem to be involved in the breakdown of arabinoxylans (Hirano et al., 2013). Similar to the results of GT43 and GT47 genes (Table 2), the expression levels of GH10 and GH51 genes were almost constant during the cultivation period and microgravity had little effect on these expression levels (Table 3).
Expression Levels of Genes for Xylan Backbone Synthesis (GT 43 and GT 47) in Rice Shoots Grown Under Artificial 1 g and Microgravity Conditions in Orbit
The expression levels of genes were analyzed using the values of fluorescence intensities obtained through a microarray analysis. As to the GT47 genes, genes homologous to OsIRX10 were investigated. Genes with a fluorescence intensity of more than 100 are shown. Data are mean ± SE from four independent samples.
Mean values significantly different between artificial 1 g and microgravity conditions (Student's t test, p < 0.05). Figures in parentheses are values of the fold change in signal intensity of microgravity-grown shoots over artificial 1 g-grown ones.
Expression Levels of Genes for Xylanase (GH 10) and α-L-Arabiosidase or β-D-Xylosidase (GH 51) in Rice Shoots Grown Under Artificial 1 g and Microgravity Conditions in Orbit
The expression levels of genes were analyzed using the values of fluorescence intensities obtained through a microarray analysis. Genes with a fluorescence intensity of more than 100 are shown. Data are mean ± SE from four independent samples.
Mean values significantly different between artificial 1 g and microgravity conditions (Student's t test, p < 0.05). Figures in parentheses are values of the fold change in signal intensity of microgravity-grown shoots over artificial 1 g-grown ones.
4. Discussion
1,3:1,4-β-Glucans and arabinoxylans are major matrix polysaccharides in gramineous cell wall (type II wall), and 1,3:1,4-β-glucans consist of Glc residues, while arabinoxylans consist mainly of Ara and Xyl residues (Carpita, 1996). Since the gramineous cell walls contain only small amounts of pectic polymers and xyloglucans (Carpita, 1996), the Glc content in the fraction represents mainly the 1,3:1,4-β-glucan content, and the arabinoxylan content can be estimated from the combined amounts of Ara and Xyl in the fraction (Obel et al., 2002). Therefore, the results of sugar contents shown in Fig. 3 suggest that microgravity decreased the amount of 1,3:1,4-β-glucans in cell walls of rice shoots, but it barely affected the amounts of arabinoxylans and other matrix polysaccharides. The molecular masses of 1,3:1,4-β-glucans are shown to be higher than one million (Sakurai, 1991; Hoson, 2002). The glucans are thought to interact with other matrix polysaccharides and cellulose microfibrils in cell walls of gramineous plants (Carpita and Gibeaut, 1993). Therefore, the metabolism of 1,3:1,4-β-glucans is involved in the regulation of the mechanical (physical) properties of gramineous cell walls (Sakurai, 1991; Hoson, 1993). For example, the breakdown of 1,3:1,4-β-glucans was closely associated with the cell wall loosening mediated by auxin, a plant hormone, treatment and inhibition of the degradation of such glucans with antibody raised 1,3:1,4-β-glucans and 1,3:1,4-β-glucan hydrolases nullified auxin-induced cell wall loosening (Hoson, 1993). Recent molecular genetic studies also showed that the decrease in level of 1,3:1,4-β-glucans reduced the mechanical strength of cell walls in vegetative tissues of rice (Vega-Sanchez et al., 2012; Kido et al., 2015). The present results (Fig. 3) suggest that the amount of 1,3:1,4-β-glucans in shoot cell walls was decreased under microgravity conditions. Our previous study showed that cell walls of rice shoots grown for 5.3 and 5.6 days under microgravity conditions were loosened compared with those grown under artificial 1 g conditions, and that microgravity suppressed the DFA formation in shoot cell walls (Wakabayashi et al., 2015). We assumed that the decrease in DFA-mediated cross-linkages within the cell wall architecture contributes to keeping the cell wall loosened under microgravity conditions. Taken together, the present results suggest that, in addition to the decreased DFA formation, the reduced level of 1,3:1,4-β-glucans under microgravity conditions is also involved in keeping the cell wall loosened.
Metabolic processes, such as breakdown and biosynthesis, are closely associated with the regulation of the amounts of cell wall constituents. Endo-1,3:1,4-β-glucanases (EC 3.2.1.73), members of GH family 17, have been shown to specifically hydrolyze high-molecular-mass 1,3:1,4-β-glucans and release oligosaccharide products, and rice plants possess two genes encoding endo-1,3:1,4-β-glucanase (OsEGL1 and OsEGL2) (Akiyama et al., 2009). The transgenic rice line in which the OsEGL1 gene is overexpressed showed a significant reduction in 1,3:1,4-β-glucan content in stem tissues compared with wild-type plants (Kido et al., 2015). Therefore, OsEGL1 may have a principal role in the breakdown of 1,3:1,4-β-glucans in rice cell walls. In the present study, OsEGL1 and OsEGL2 were expressed in shoots and their levels increased during the cultivation period (Fig. 4). The expression levels of OsEGL1 in microgravity-grown shoots were higher than those in artificial 1 g-grown ones at both 4 and 5.6 days, while those of OsEGL2 were almost the same between the two conditions (Fig. 4). The former space experiment showed that the 1,3:1,4-β-glucan degrading activity in rice (cv. Koshihikari) shoots grown for 5.5 days under microgravity conditions in space was higher than that under 1 g conditions on the ground (Hoson et al., 2000). The increase in the expression level of OsEGL1 gene under microgravity conditions coincides with the above observation. The former and present studies suggest that microgravity promotes the breakdown of 1,3:1,4-β-glucans in rice shoots.
On the contrary, two subfamilies (CslF and CslH) of the Cellulose synthase-like (Csl) superfamily of glycosyl transferases have been shown to mediate the biosynthesis of 1,3:1,4-β-glucans in gramineous plants (Burton et al., 2006; Doblin et al., 2009). Particularly CslF6 has a principal role in the 1,3:1,4-β-glucan biosynthesis in rice. The expression level of CslF6 gene was closely correlated with the 1,3:1,4-β-glucan synthase activity in growing rice seedlings (Kimpara et al., 2008), and rice CslF6 knockout mutants showed a drastic decrease in the 1,3:1,4-β-glucan content and in the 1,3:1,4-β-glucan synthase activity (Vega-Sanchez et al., 2012). In the present study, the CslF6 gene showed a strong signal intensity and its expression level was almost constant during the cultivation period. The expression levels of CslF6 gene in microgravity-grown shoots were comparable with those in artificial 1 g-grown ones (Table 1). Furthermore, the expression levels of other CslF genes and CslH genes were almost the same under both conditions (Table 1). These results suggest that microgravity barely influences the biosynthesis of 1,3:1,4-β-glucans. Taken together, we assume that microgravity promotes the breakdown of 1,3:1,4-β-glucans by enhancing the expression level of OsEGL1 gene, which, in turn, may cause the reduced level of such glucans in growing rice shoots.
At present, the regulation mechanism by which microgravity modifies the expression level of OsEGL1 gene has been unclear. Akiyama et al. (2009) showed that the expression level of OsEGL1 gene in rice seedlings was enhanced by the treatment with jasmonate, a kind of plant hormone. We previously reported that the levels of jasmonate in rice shoots grown under microgravity were higher than those grown under artificial 1 g conditions during the cultivation period (Wakabayashi et al., 2017). Therefore, it is probable that the jasmonate signaling may be involved in the regulation of OsEGL1 gene expression in rice shoots grown under microgravity conditions.
In gramineous cell walls, arabinoxylans, in addition to 1,3:1,4-β-glucans, are predominant matrix polysaccharides (Carpita and Gibeaut, 1993) and they also possess phenolic acid monomers such as FA and p-coumaric acid, which are ester-linked to Ara residues of arabinoxylans (Smith and Hartley, 1983; Saulnier and Thibault, 1999). According to the cell wall model, gramineous arabinoxylans continue to increase during cell wall maturation and they form a tight network via phenolic acid crosslinks within the cell wall architecture (Carpita and Gibeaut, 1993; Carpita, 1996). The amounts of Ara and Xyl in the matrix polysaccharide fraction obtained from microgravity-grown shoots were comparable with those from artificial 1 g-grown ones (Fig. 3), suggesting that microgravity barely affects the amount of arabinoxylans. Furthermore, the results of expression levels of genes involved in the biosynthesis and breakdown of arabinoxylans (Tables 2 and 3) show that the microgravity environment had minimal impact on arabinoxylan metabolism. As described above, we previously reported that microgravity decreased the arabinoxylan/phenolic acid network structures in rice shoot cell walls by decreasing the cell wall peroxidase-mediated coupling reaction to produce DFA, which crosslinks arabinoxylan molecules (Wakabayashi et al., 2015). Taken together, microgravity affects the formation of such intranetwork structure by modifying the metabolism of phenolic acid moiety, but not the arabinoxylan metabolism.
Modifications of mechanical properties of cell walls under microgravity conditions have been observed in dicotyledons having the type I wall. The cell wall extensibility, which represents the ability of cell wall to extend, in elongating regions of Arabidopsis inflorescence stems increased under microgravity in space compared with artificial (on-orbit) and ground 1 g conditions (Hoson et al., 2014). Similarly, Soga et al. (2002) showed that in seedlings of Arabidopsis, the cell wall extensibility of space-grown hypocotyls was higher than that of ground-grown ones. In cell wall constituents, space-grown hypocotyls had lower molecular mass of xyloglucans (Soga et al., 2002). Xyloglucans are predominant matrix polysaccharides in the type I wall and tether cellulose microfibrils (Carpita and Gibeaut, 1993). Therefore, the breakdown and thus the molecular mass downshift of xyloglucans are closely associated with the cell wall loosening of the type I wall (Sakurai, 1991; Hoson, 1993). Furthermore, the activity of xyloglucan-degrading enzymes extracted from cell walls of space-grown hypocotyls was higher than that of ground-grown ones (Soga et al., 2002), suggesting that microgravity reduces the molecular mass of xyloglucans by stimulating their breakdown, which leads to an increase in the cell wall extensibility of hypocotyls. These findings and the present results suggest that the preferential stimulation of metabolic turnover of certain wall polysaccharides possessing a load-bearing function underlies the modification of mechanical properties of both the type I and the type II walls under microgravity conditions.
Plant cultivation will be required to construct bioregenerative life-support systems on the long-term space missions (Ferl et al., 2002). Particularly, production of cereal grains under microgravity conditions contributes directly to the food supply in space environment. It has been shown that grains of some gramineous plants contain a significant amount of 1,3:1,4-β-glucans as a storage compound, and their levels are comparable with the endosperm cell walls (Fincher, 2009). Because microgravity affected the metabolism of 1,3:1,4-β-glucans in shoot cell walls, it is likely that microgravity affects the turnover of 1,3:1,4-β-glucans even in grains and thus it may cause aborted grain development as observed in the former space experiment (Levinskikh et al., 2000). Therefore, intensive studies on polysaccharide metabolism not only in vegetative but also in reproductive organs of crop plants grown under microgravity conditions should be undertaken in the near future to develop methods for efficient grain production in space.
In conclusion, microgravity reduces the cell wall 1,3:1,4-β-glucan content in rice shoots by increasing the expression level of an endo-1,3:1,4-β-glucanase gene. The decrease in 1,3:1,4-β-glucan content may be involved in keeping the cell wall loosened under microgravity conditions. On the contrary, microgravity scarcely affected the metabolism of arabinoxylans.
Footnotes
Acknowledgments
The Ferulate experiment was a close collaboration between the Japan Aerospace Exploration Agency (JAXA), the National Aeronautics and Space Administration (NASA), and related organizations and companies. We thank Dr. Akira Higashibata of JAXA for his great efforts during the course of the experiment.
Author Disclosure Statement
No competing financial interests exist for the authors.
Funding Information
No funding was received
Abbreviations Used
Associate Editor: Petra Rettberg