
Abstract
The Iberian Pyrite Belt in southwest Spain hosts some of the largest and diverse extreme acidic environments with textural variation across rapidly changing biogeochemical gradients at multiple scales. After almost three decades of studies, mostly focused on molecular evolution and metagenomics, there is an increasing awareness of the multidisciplinary potential of these types of settings, especially for astrobiology. Since modern automatized exploration on extraterrestrial surfaces is essentially based on the morphological recognition of biosignatures, a macroscopic characterization of such sedimentary extreme environments and how they look is crucial to identify life properties, but it is a perspective that most molecular approaches frequently miss. Although acid mine drainage (AMD) systems are toxic and contaminated, they offer at the same time the bioengineering tools for natural remediation strategies. This work presents a biosedimentological characterization of the clastic iron stromatolites in the Tintillo river. They occur as laminated terraced iron formations that are the most distinctive sedimentary facies at the Tintillo river, which is polluted by AMD. Iron stromatolites originate from fluvial abiotic factors that interact with biological zonation. The authigenic precipitation of schwertmannite and jarosite results from microbial–mineral interactions between mineral and organic matrices. The Tintillo iron stromatolites are composed of bacterial filaments and diatoms as Nitzschia aurariae, Pinnularia aljustrelica, Stauroneis kriegeri, and Fragilaria sp. Furthermore, the active biosorption and bioleaching of sulfur are suggested by the black and white coloration of microbial filaments inside stromatolites. AMD systems are hazardous due to physical, chemical, and biological agents, but they also provide biogeochemical sources with which to infer past geochemical conditions on Earth and inform exploration efforts on extraterrestrial surfaces in the future.
1. Introduction
Throughout Earth's history, mineral-microbial interactions have resulted in a wide variety of macroscopic organic sediments. Stromatolites, distinguished by their alternate lamination, stand out as the oldest and most conspicuous evidences in the fossil record of life (Buick et al., 1981; Grotzinger and Knoll, 1999; Westall, 2005). The alternate lamination of stromatolites can preserve the interplay between periodic sediment supply and biological changes in mat communities, including pauses in the sedimentation rate (Golubic et al., 2000). Although the vast majority of modern and fossil stromatolites are composed of calcium carbonate, fossil stromatolites of siliceous, phosphate, iron, manganese, and sulfate also occur, either as primary precipitates or, more commonly, as diagenetic replacement (Chauhan, 1979; Lundberg and McFarlane, 2011; Riding, 1999; Riding, 2011; Lozano and Rossi, 2012; Sallstedt et al., 2018). Today, iron stromatolites are formed in unique modern settings with complex biogeochemical interactions that serve as geochemical, mineral, biological, and environmental sources. The early record of life suggests that iron metabolism was not only ubiquitous, but it was also among the most ancestral biochemistries. For instance, the distribution of iron-oxidizing bacteria that are capable of switching from chemolithoautotrophy to photoautotrophy in deep-sea hydrothermal settings (Ishii et al., 2015) also indicates that dissimilatory Fe(II) oxidation is probably an ancient energy metabolism (Ilbert and Bonnefoy, 2013).
Iron is not only the most abundant metal on Earth, but also about 90% of all iron formations are derived from Precambrian banded iron formations (BIFs) (Pilchin and Eppelbaum, 2009). However, pyrite (FeS2) is the most abundant iron–sulfur mineral at Earths' surface, and it is present in igneous, metamorphic, and sedimentary rocks (Rickard and Luther, 2007). Because iron and sulfur were abundant and accessible, early life was probably fueled by multiple oxidation and reduction reactions (Knoll, 2003; Canfield et al., 2006; Planavsky et al., 2009; Lalonde and Konhauser, 2015).
Current polluted environments known as acid mine drainage (AMD) contain copious amounts of iron and sulfur, and therefore, they are excellent modern analogues of ancient scenarios. AMD settings, which are rich in Fe2+ ions, trace elements such as arsenic (As), and other heavy metals, represent the contaminated tailings of the mining industry. The oxidation of sulfide minerals (as pyrite) in AMD causes high acidity and high concentrations of soluble metals (Konhauser, 2007). Despite their toxicity with regard to health, environment, soil, and water resources, AMD fosters complex biogeochemical interactions that maintain a rich geomicrobiological diversity pool.
Recent trends in mineral–microbial interactions have led to the proliferation of studies in AMD as niches for microbial colonization (Bird et al., 2011; Caraballo et al., 2011; Nordstrom et al., 2015; Huang et al., 2016; Shuster et al., 2018; Gavrilov et al., 2019; Hasiotis and Brake, 2019; Lukhele et al., 2020; Rivera et al., 2019; Gao et al., 2020). Such extreme environments are also settings that sustain the formation of recent iron stromatolites that accrete as iron terraces. As documented by pioneer studies, the AMD in the Tintillo river is one of the few places worldwide where extensive terraced iron formations readily form (Sánchez-España et al. 2005b, 2006, 2007). Throughout this article, the term “TIFs” will be used to refer to terraced iron formations as used by Sánchez-España et al. (2007). We define terracettes as depositional facies that have a geomorphological attribute of narrow, short, and rarely parallel treads at a steeper angle than other depositional morphologies (Anderson, 1972). These features generally form in carbonate travertine systems. Although Fe-rich sediments are globally distributed, their occurrence as TIFs is rather scarce (Table 1).
Iron Stromatolites in Reported Acid Mine Drainage Sites with Terraced Iron Formations
AMD = acid mine drainage; masl = meters above sea level; TIF = terraced iron formation.
Stromatolites grow and accrete by trapping, binding, and precipitation of sedimentary particles by benthic microbial communities (Awramik, 1977). Yet, one of the greatest challenges in stromatolitic lamination is the distinction between biotic and abiotic processes, which are by far more complex in iron stromatolites (Brake et al., 2001, 2004, 2014). Previous studies at Tintillo suggest that iron stromatolites result from the alternation between abiotic factors (hydrological process) and biological factors where iron precipitation is mediated by bacterial metabolism (Sánchez-España et al., 2007). Studies have also shown that TIFs are mainly influenced by cyclic seasonal variations: cooler temperatures in winter correlate with higher flow conditions that increase pH and favor mineral precipitation. Concordantly, higher temperatures correlate with lower streamflow, iron precipitation, and higher bacterial populations.
What is less clear is the relation between photosynthetic eukaryotes and the mineral matrix, especially in the dry season. In the present study, our goal was to analyze the occurrence of photosynthetic eukaryotes in Tintillo TIFs in seasons when accretion is at minimum. Since TIFs display multiple scalar variations, a second goal for this study was to assess their astrobiological relevance from a morphological perspective. To characterize TIFs at the geochemical, mineral, and biological level, we employed a combination of qualitative and quantitative approaches.
Gradients generated by concurrent biogeochemical processes often develop into multiple-scale biosignatures. This study was designed to contribute to this growing research area by revising a morphological first approach in astrobiology. As long as modern space exploration depends on large-scale morphological recognition, AMD systems have the potential to provide good models with which to study the occurrence of structures, processes, and patterns.
2. Geological Setting
The Iberian Pyrite Belt (IPB) in southwest (SW) Spain contains unusual AMD deposits that have been studied over the course of decades. This area forms part of the IPB and the volcano-sedimentary complex, which are dominated by volcanic-siliceous rocks (rhyolitic todacitic tuffs, breccias and lava flows, tuffites) and graywackes and shales, where no carbonate or alkaline materials are present (Sánchez-España et al., 2005a, 2006). The area under study here corresponds to the Tintillo deposit, which is polluted by AMD and ∼10 km long, 40 m wide, covering an area of 57 km2. This stream is characterized by receiving exclusively the input of acid sulfate waters from the four main springs that emanate from the base of sulfide-bearing waste-rock piles. The larger spring is from a vast open-pit (∼1 km long and 40 m deep) that is situated in the surroundings of Corta Atalaya. The three other fluvial inputs are from the Barrizal, Gangosa, and Escorial tributary creeks of the Tintillo river (Sánchez-España et al., 2005b).
The present study was conducted in the Tintillo river contaminated by the Río Tinto area, located to the North of Huelva, a province of SW Spain (37°43′0″N; 6°38′37″O). The area under study is located 7.3 km north of Campillo (Fig. 1). The mean annual temperature is 16.1°C, with a mean minimum temperature of 9°C (between December and January) and a mean maximum temperature of 24°C (July and August), reaching maximum values of 35°C in these 2 months. The climate zone corresponds to Continental Mediterranean, with a mean annual precipitation of 849.1 mm and a mean minimum precipitation around 3.65 mm between July and August (Tavira, 2016). Previous works have reported Fe-rich microbial laminated muds and emphasized the profuse development of microbial mats under the influence of biotic and abiotic factors (Sánchez-España et al., 2005b, 2006, 2007). Caraballo et al. (2011) documented the seasonal variations in the formation of Al- and Si-rich Fe stromatolites in the highly polluted AMD of Agua Agria Creek (Tharsis). These authors characterized the chemical and mineralogical composition of Fe stromatolites and confirmed the prominent role of diatoms in their formation.
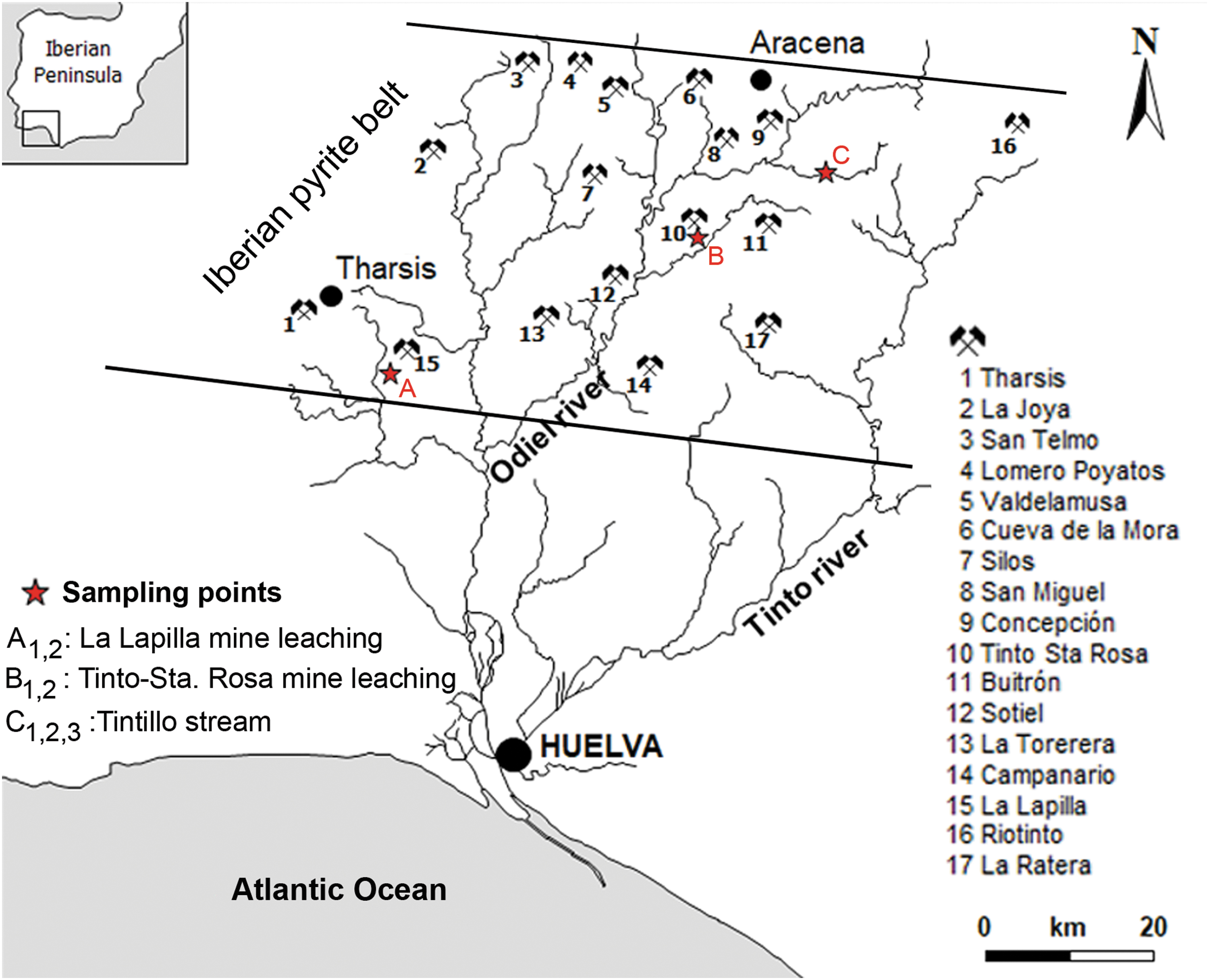
Location map of the Tintillo AMD river in southern Spain. The hydrological map shows main localities with AMD tails, waste rock dams, and sampling points (red stars). AMD, acid mine drainage.
The Tintillo is a meandering river with dendritic tributaries that runs on gently inclined floodplains and decreases in slope in the downcurrent direction. Tintillo stream waters run through shallow water subenvironments toward the distal parts of the system before it confluences with the Odiel river (Sánchez-España, 2007). The geomorphology in Tintillo AMD is dominated by gradational terraces that extend along the lateral stream of mine tailings (Sánchez-España et al., 2005a, 2007). In this context, the sulfurous waters of Tintillo river, which are polluted by AMD, can be defined as an extreme habitat in terms of their very low mean pH (near 2.5) and high concentration of heavy metals, especially ferric iron, copper, zinc, and sulfate (López-Archilla and Amils, 1999; Aguilera et al., 2006). The Tintillo shores are characterized by the presence of extremophiles vegetation such as Cistus ladanifer, the endemic heath Erica andevalensis, Pinus pinea, and Pinus pinaster (Santa-Bárbara and Valdés, 2008).
3. Materials and Methods
This study included both field and laboratory analyses. Fieldwork consisted of geochemical measurements of surface waters, mapping, facies description, photo documentation, and sampling of solid TIFs. Based on the work of Sánchez-España et al. (2007), selected sampling points along the main course of the Tintillo river were visited during the summer of 2018 (Fig. 1). Samples taken from Tintillo surface waters were tagged as T. (V 2018). To evaluate the evolution of water geochemistry, this work included a comparison with other localities with the formation of TIFs in Tintillo Santa Rosa [T.S. (P 2003) and T.S. (V 2003)] sampled in spring and summer of 2003, respectively. Other included nearby sites without the presence of stromatolites were from La Lapilla stream, tagged as [L. (I 2003) and L. (P 2004)], taken in winter of 2003 and spring of 2004, respectively. We also included TIFs previously sampled in summer of 2004 T. (V 2004) and summer of 2015 T. (V 2015).
3.1. Fieldwork: measurements on site and sampling
Temperature (T), pH, redox potential (Eh), and electrical conductivity (EC, μS/cm or mS/cm, at 25°C) were recorded in situ with a multiparametric portable device (CrisonMM40) and a measuring scale of up to −2 pH units. The pH meter range was between 0.00 and 14.00 pH with a 0.01 pH resolution and a ±0.02 pH accuracy, and it was calibrated with WTW standard solutions (pH 2.00, 4.01, and 7.00); the redox potential was checked with Hanna standard solutions (240 and 470 mV). For cations and metal analysis, two water samples were taken; to avoid metal precipitation, samples were acidified in the field to pH <2 with Suprapur HNO3 (2%). Samples were kept in the dark at 4°C in 75 mL sterilized polyethylene bottles during the transport to the laboratory.
For sedimentological analysis, two fresh cross-sectional TIFs samples were collected from Tintillo river, tagged as P282 and P295 (in blocks of 50 × 20 cm), and kept cold at 4°C until immediate laboratory analysis. For comparison and as a negative control, ∼100 g of sediment samples without (TIFs) was taken from the river bedrock. Algal biofilms attached to TIFS rims were sampled, fixed with a formaldehyde solution (4% v), and stored in a light-protected flask for further analysis.
3.2. Analytical methods
3.2.1. Water chemical analysis
The concentration of dissolved Al, As, Ca, Cr, Cu, Fe, Mg, Mn, Ni, Pb, Si, and Zn was determined by inductive coupling plasma atomic emission spectrometry (ICP-AES; Jobin-Ybon Ultima2). Calibrations were performed at the beginning and at the end of each analytical series by using a multielement standard solution prepared from certified standards supplied by SCP SCIENCE. The detection limits were calculated on average after three repeats, and the standard deviations of 10 blanks were also estimated. Certified Reference Material SRM-1640 NIST and interlaboratory standard IRMM-N3 wastewater test material were used according to the European Commission Institute for Reference Materials and Measurements. The detection limits for larger cations were: 200 μg/L for Al, Fe, Mn, Mg, Si; 500 μg/L for Ca; for trace elements, they were 50 μg/L for Zn, 5 μg/L for Cu, 2 μg/L for As, and 1 μg/L for the rest of the elements.
3.2.2. Sedimentology
A granulometry analysis was carried out to obtain in-depth information on TIF lamination. A subset of collected field samples of TIFs were air-dried at room temperature, ground, and sieved to <2 mm. TIF fractions and controls were processed by forcing the material through a 2-mm screen, by hand or with a large rubber stopper, and stored. Weight measurements were made and recorded on the 20- to 75-mm, 5- to 20-mm, and 2- to 5-mm fractions.
3.2.3. Petrography and compositional analysis of TIFs
Two petrographic-thin sections along the x and z axes were made, using a GTS polyester casting resin (Modulor GmbH), and analyzed by optical microscopy using a Nikon ECLIPSE TS100 inverted microscope coupled to a DS-Fi2 Camera with Digital Sight Unit DS-U3. To investigate whether lamination textures were correlated with chemical variations, dry TIF carbon-coated samples were examined under a JEOL JSM-5410 scanning electron microscope (SEM). For a quantitative compositional map of TIF samples, a fitted wavelength-dispersive spectrometer was used under the following conditions: accelerating voltage was 15 kV, the probe current was 20 nAmp, and beam diameter was 5 μm. Twelve TIF subsamples from P-281 (x axis) and P-281 (z axis) were analyzed, and 13 elements were determined (Si, Ca, Zn, Mg, Al, K, Cu, As, Cl, Fe, Pb, Mn, and S). One advantage with this analysis is that it avoided the problem of sample disruption, since TIFs are fragile and delicate. Several gold-coated TIF samples were studied under a SEM (JEOL JSM-5410) using an accelerating voltage of 15 kV and a beam current between 0.5 and 3 nAmp to observe the morphology and distribution of the constituent minerals.
3.2.4. Microbial analysis
TIF samples were analyzed under a NIKON Eclipse E200 microscope (Plan UW 20X) coupled to a DS-Fi2 Camera with Digital Sight Unit DS-U3. Also, some permanent slides, after gold coating, were analyzed under a SEM (JEOL JSM-5410) using an accelerating voltage of 15 kV and a beam current between 0.5 and 3 nAmp.
4. Results
4.1. Water geochemistry
The Tintillo water chemistries are presented in Table 2. Acid waters exhibit a pH that ranges between 2.6 and 2.9 in the selected sampling sites; the pH remains relatively constant along the stream course up to its confluence with the Odiel river (Fig. 1). Tintillo waters show a high oxidation potential (above 600 mV) and an electrochemical conductivity above 12 μS/cm. The variation in the As concentration is evident, with values ranging below the detection limit. Other measured elements show a high temporal stability, except for Si, whose lower concentration correlates with a decrease in As and Pb. The concentration of Fe and Ca is much lower in streams without stromatolites than in streams with stromatolites, with concentrations that do not exceed 20 mg/L of Fe and 57 mg/L of Ca (Table 2).
Comparisson of Physical–Chemical Parameters and Water Composition of the Different Localities (With and Without Stromatolites) Studied
Samples with the presence of stromatolites: T.S. (P 2003) = Tintillo Santa Rosa (Spring 2003); T.S. (V 2003) = Tintillo Santa Rosa (Summer 2003); T. (V 2015) = Tintillo (Summer 2015); T. (V 2018) = Tintillo (Summer 2018); T. (V 2004) = Tintillo (Summer 2004). Samples without the presence of stromatolites: S.T. (V 2004) = San Telmo (Summer 2004); S.T. (P 2006) = San Telmo (Spring 2006); L. (I 2003) = La Lapilla (Winter 2003); L. (P 2004) = La Lapilla (Spring 2004).
EC = electrical conductivity; Eh = redox potential.
4.2. Depositional facies and TIFs
The Tintillo AMD river discharges acidified reddish waters along a main channel bordered by a deciduous shrub forest and red shales. The marginal zonation of shales caused by sulfur precipitates suggests seasonal precipitation (Fig. 2a, b). The geomorphology of Tintillo produced by transport, erosion, and depositional processes includes aqueous subenvironments and their associated sedimentary structures, which can be grouped into four depositional facies: (1) gossans and stream deposits, such as pebbles and boulders, from proximal to distal facies; (2) marginal channel facies characterized by benthic and planktonic microbial mats with minerals precipitated around shales; (3) point bars streams with iron terracettes or TIFs; and (4) small oxbow lakes and creeks. TIFs are the most conspicuous facies in Tintillo (Fig. 2c, d), and they form by erosion and inundation episodes by circulating AMD waters. They occur as downstream low-relief crenulated ridges with 0.3–0.5 m steep slopes (0.5–1 m in diameter), which confine polygonal red pools (Fig. 2e, f).

Sedimentary facies in Tintillo AMD.
The polygonal geometry of pools breaks in those areas where channels are wider (more than 7 m across) and where sinuosity and turbulent flow are higher (Fig. 3a). Ridged TIFs exhibit a macroscale corrugated surface with intercalated layering (Fig. 3b, c). At the centimeter and millimeter scale, undulating white sulfur precipitates on siltstones are common along the mainstream course (Fig. 3d). In contrast to exposed TIFs, submerged TIFs show a green coloration due to the spotty presence of microbial mats.

Terraced-iron formations (TIFs) in the Tintillo river. (TIFs).
The thickness of TIFs varies from a few centimeters up to 50 cm and exhibit a surficial ocher coloration in areas of higher water depth, especially in zones where orange, yellow, and green floating filamentous mats develop. There is a clear yellow to green zonation along the mid-ridges of pools (Fig. 4a). In general, coarser filamentous populations coincide with coarser sediments, whereas finer filamentous mats develop in flat muddy areas. Filaments align in parallel in zones where sedimentary ridges show a regular pattern (Fig. 4b). The occurrence of monotypic yellow filaments in zones of higher turbulence (Fig. 4c) contrasts with pustular green mats developed around a steep relief or inside natural crevices (Fig. 4d).

Representative microbial mats in Tintinillo AMD.
4.3. Characterization of TIFs
TIFs exhibit a variable texture and variations among brownish, ochre, and dark red colorations. At 25°C, sampled TIFs show an internal pH of 1.36. The macrostructure of TIFs consists of corrugated surfaces and smooth wavy layering of variable thickness (Fig. 5a, b). A fine greenish-silty and porous lamination at the top is followed by a compacted zone with a millimeter-thick reddish lamination (white arrow in Fig. 5c). The TIFs mesostructure shows a crude granular red/ochre lamination (between 0.5 and 2 cm) that becomes evident upon desiccation. The spongy and porous zone exhibits a brownish lamination and a higher content of plant debris at the lower part, especially acicular pine needles (Fig. 5d). TIFs also reflect a preferential granulometry, consisting of coarse silt-fine-sands and, to a lesser extent, coarse and very coarse sand. TIFs incorporate medium to coarse sand in both analyzed samples, unlike nonstromatolitic iron sediments, which exhibits a more regular distribution of particle size (Fig. 6). Petrographic analysis results show a sharp transition between the ochre and brownish coloration (Fig. 7a), followed by a Fe-rich shrub-like porous texture that shows a dendritic fabric with dark red aggregates (Fig. 7a, b). A second TIF sample similar in color and texture shows differences in lamination thickness and dendritic aggregates (Fig. 7d, f). The dendritic fabric is clearly separated from a massive iron precipitation and followed by iron aggregates (Fig. 7f).

Mesostructure of sampled TIFs.
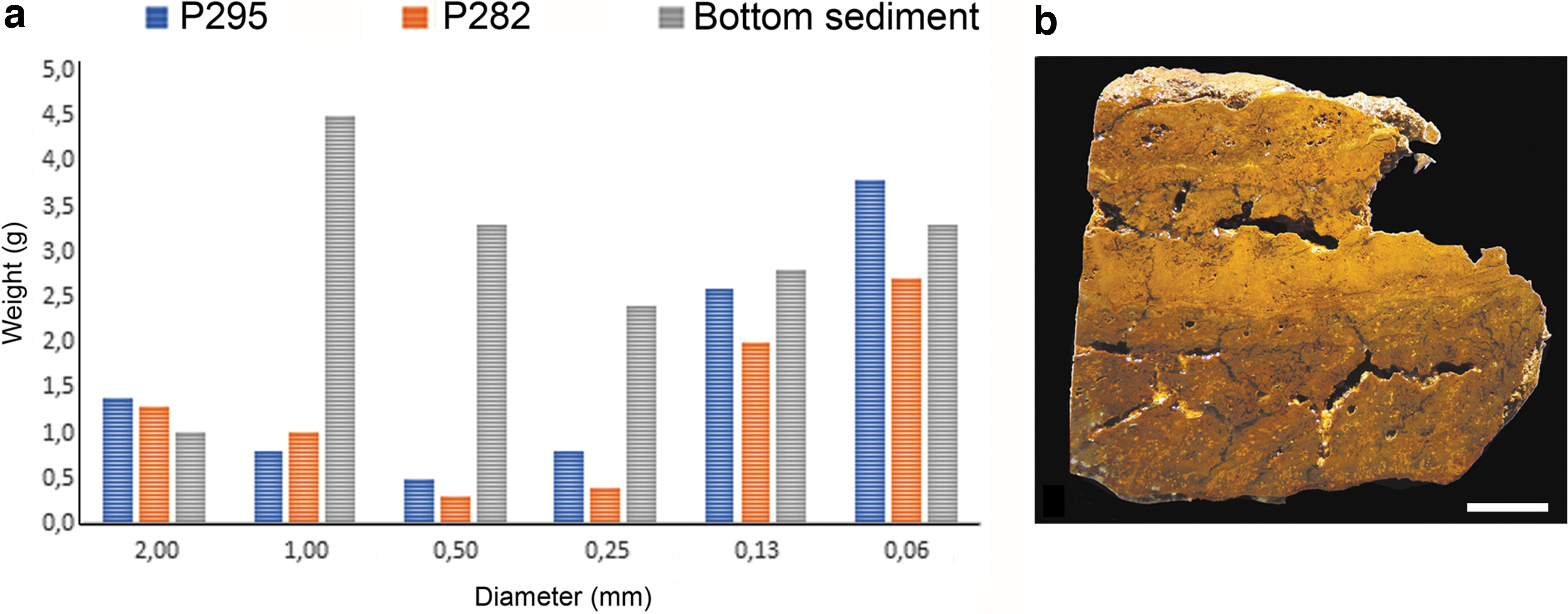
Grain size distribution in two TIFs (P282 and P295).

TIFs petrography.
At higher resolution, TIFs show three main textures: a porous and spongy zone, followed by a well-developed lamination with irregular margins that alternates with a spongy fabric, and massive and compacted white zones (Fig. 8a). Thrombolitic textures form a clear banding distinguished by their precipitate size, composition, and color. The main microtextures consist of light-gray millimeter-alternating-layers, with a central precipitation zone in lateral continuity, and an upper mosaic precipitation pattern (Fig. 8b, c). The lower area exhibits two distinctive zones, with heterogeneous iron minerals, and a coarser layering associated with a branching pattern of precipitates (Fig. 8d, e). The cyclic presence of gray and white banding with bundles of parallel filaments and precipitates is observed through the scanned profile. Some zones show a composite fabric, represented by an upper banding with a high structural organization, and sharp boundaries among light, gray, dark, and white zones (Fig. 9a). The internal geochemical profile clearly shows fluctuations mainly in SO3 and FeO (Fig. 9b). Interestingly, chloride, SiO2, and Al2O3 show intermittent variations, but other cations remain relatively constant. Similar textural variations can be observed along the z axis orientation (Fig. 9c).

TIFs microstructure.

Petrographic compositional map of TIFs.
4.4. Microbial assemblage in TIFs
TIFs show at least four discontinuous layers that contain filamentous microbial mats, mainly composed of long algal filaments and other minor uniseriate filaments (arrow in Fig. 10a). The unbranched filaments formed by short cells have been presumably assigned to Klebsormidium. Other long filaments, formed by barrel-shaped cells similar to preakinetes of Zignema sp., are frequent (Fig. 10b, c). Some cells show a terminal dichotomous branching (a Y-bifurcation) and a spheroidal protuberance (arrow in Fig. 10b). Filaments are closely associated with diatoms and with brownish precipitates (Fig. 10d). In contrast to the floating microbial mats formed by thick light orange and green filaments, the filaments in TIFs show yellow, white, and brown precipitates among filament intersections. Such precipitates occur as small, euhedral, or irregular yellow minerals that entomb filaments (Fig. 10e). Other precipitates occur as small globular sulfur on filaments, which may exhibit a black or white color (arrows in Fig. 10f). A closer look reveals that barrel-shaped cells (4 μm in diameter) form uniseriate trichomes surrounded by a thin mucilaginous material (Fig. 10g, h). The black entangled filaments with sulfur precipitates are associated with diatoms, which are abundant through the vertical profile of TIFs.

Microbial filaments in Tintillo TIFs.
4.5. Mineral–extracellular polymeric substances interaction
Although filamentous morphotypes are a dominant microbiota, pennate frustules are not only well represented, but they occur in close association with mineral precipitates and with extracellular polymeric substances (EPS). Diatoms are conspicuous throughout the vertical profile of TIFs, and they preferentially align on outlining interfaces of the mineral matrix (Fig. 11a). The mineral matrix is composed of radial spherules of schwertmannite [Fe+3 16O16(OH)12(SO4)2] with organic remains in which diatoms are embedded (Fig. 11b–f). It is clear that the density of diatom populations influences the geometry of open spaces inside the mineral matrix (Fig. 11a–c). The EPS seems to promote mineral cohesion and agglutination (Fig. 11e). Stauroneis kriegeri is commonly found inside a reticulated mineral matrix (Fig. 11h) and in massive amorphous surfaces (Fig. 11i).

TIFs 3D microstructure.
The diatom assemblage includes Nitzschia aurariae, Pinnularia aljustrelica, S. kriegeri, and Fragilaria sp. These pennate diatoms form an integral part of the mineral matrix, which contains remains of organic EPS and schwertmannite as the most common mineral (Fig. 12a). TIF upper layers contain interconnected spheroidal radial crystals of schwertmannite (Fig. 12b). P. aljustrelica, among others, is closely associated with schwertmannite (Fig. 12c–f). Other small crystals may be attached to cell walls as connecting pads, only on one side (Fig. 12f).

SEM micrographs of diatoms inside iron stromatolites.
5. Discussion
5.1. The stromatolitic texture of TIFs
The formation of terracettes as a geomorphological feature of riverine systems has been classically assigned to erosion and aggradation. The floodplain can be transformed into a terrace by tectonic, climatic, or human-induced changes that alter the regimen of the river (Gordon and Luna, 1967) where erosional edges are suitable substrates for microbial mats (Gerdes et al., 1994). The high diversity of geochemical gradients promotes the colonization of the water–sediment interface by microbial communities. These microecosystems, in turn, accrete and stabilize during erosion, turbulence, and inundation episodes. The increasingly AMD anthropogenic deposits, which unfortunately exhibit an extensive distribution worldwide, are propitious environments for the formation of iron stromatolites. These clastic iron stromatolites are the result of a synergistic effect among clastic input, microbial metabolisms, and fluvial processes that produce similar mineral facies in AMD systems.
A common problem associated with microbial sediments is their classification in genetic and descriptive terms. However, the depositional environment adds another criterion in microbialites. The Tintillo clastic iron stromatolites are unique in their genesis, and therefore, their classification as microbialites is not straightforward. In low-energy settings, TIFs could also be regarded as unlithified microbial mats where mineralogy may dictate the difference between lithified and unlithified mats. Their morphological attributes, such as a millimeter-scale flat-topped crests similar to ripples and a reticulate pattern on crenulated microstructures, are comparable to other ancient siliciclastic surfaces (Porada and Bouougri, 2007). Since TIFs are not discrete structures, but form part of a clastic interface environment formed by chemical iron precipitation (basically, a clastic deposit), they could also be regarded as a laminated example of MISS (microbially induced sedimentary structures), following the criteria given the works of Noffke et al. (2003, 2008) and Noffke and Awramik (2013). The debate about the nature of MISS (Davies et al., 2016) highlights the complex interplay between abiotic and biotic factors in the accretion of microbial structures with a well-defined pattern (lamination).
TIFs are characterized by a specific grain-size range formed in siliciclastic environments, under relatively stable hydrodynamic conditions, with seasonal variations within a contaminated river. Because of their lamination, they are reported as clastic iron stromatolites, with precipitation, union, baffling, and trapping processes, but no lithification. As pointed out by Gerdes (2007), many terms reflect similar features (see, for instance, the pioneer terminology in Logan et al., 1974). Under this umbrella, TIFs could be regarded as laminated microbial deposits with a high amount of detrital sediments. Given that TIFs represent modern, although not lithified, potential stromatolites (Krumbein, 1983) and form accretionary structures with a biogenic primary lamination, they could also be fitted into the stromatolites' definition given in the work of Hofmann et al. (1999). In modern environments, TIFs may be viewed as products of biologically mediated geochemical gradients that develop a “stromatolitic” lamination (sensu Kalkowsky, 1908), where nutrients, flow rates, and other critical parameters promote their development (Dodd et al., 2018).
5.2. TIFs and water geochemistry
As observed in Table 2, there is a trend toward a seasonal stability for measured parameters in the studied sampling sites. The concentrations of dissolved elements in Tintillo are very high. For instance, the sodium concentration averaged 29.8 mg/L, but previous results reported a sodium average value of 32.02 and 24.17 mg/L for 2003 and 2004, respectively (Sánchez-España et al., 2007). In the present study, potassium was not detected, but its average values were 1.61 and 0.98 mg/L in 2003 and 2004, respectively (Sánchez-España et al., 2007). The absence of Cl and Br in this and in previous studies of the same localities is clear.
The microtexture of TIFs shows that FeO is the most abundant ion, followed by SO3, that presents larger variations in abundance along the vertical fabric. SiO2 also shows large variations, but Al2O3 shows variations to a lesser extent. Potassium, magnesium, and calcium show smaller fluctuations in TIFs. As presented in Table 2, waters surrounding TIFs also contain a significantly higher content of most measured cations. This cation enrichment in TIFs probably elicits bacterial ion pumps and multiple biogeochemical pathways to cope with high metal concentrations. Such adaptive microbial responses further increase the complexity between abiotic and biotic factors in AMD.
The EC is clearly different in localities with and without TIFs development. TIFs show a relatively high EC (Table 2), which is due to elements such as Fe, Mg, Zn, Ca, Al, and sulfates. A higher EC may also favor the development of lamination due to a rapid current flow through the biofilm. However, other stromatolites samples, as shown in Table 2 [samples T.S.R. (P 2003) and T.S.R. (V 2003)], display lower EC values and a higher pH, but they also exhibit a relatively smaller size.
These results also show that stream waters at Tintillo AMD present anomalies in the distribution and sequestration of toxic metals. Apparently, TIFs grow under a high mineral selectivity, since surrounding waters show higher concentrations of metal in comparison with waters where TIFs are absent (Table 2). This finding may also suggest that floating microbial mats may capture and provide sequestration sites for toxic metals. Metal concentration in AMD systems changes seasonally but is strongly dependent on the proximity of tailings piles and abandoned mines (Campaner et al., 2014); it is also a function of depth (Wielinga et al., 1999). As observed in Fig. 6, TIFs preferentially accumulate either very large or very small grains in their lamination. This attribute may be related to the microbial population density and the quantity of organic matter, which may promote trapping and binding of some grains more than others. The accretion of mats by precipitation, trapping, and binding has been reported in similar TIFs structures (Hasiotis and Brake, 2019).
5.3. Diatoms EPS and schwertmannite
The present study provides direct evidence of the interaction among diatoms as P. aljustrelica and N. aurariae, EPS, and schwertmannite (Figs. 11 and 12). Recent literature has documented the role of microbial EPS on mineralization processes during accretion of stromatolites and a prominent role for diatoms (Brake et al., 2004, 2014; Hasiotis and Brake, 2019). The acidophilic diatom P. aljustrelica has been found to thrive in the conspicuous biofilms at the Río Tinto and other related rivers from the IPB (Amaral-Zettler, 2012; Luís et al., 2012, 2016). The close relationship between schwertmannite and EPS suggests an active role of diatoms in precipitation. These microbial–mineral interactions not only influence mineral transformations (Bao et al., 2018) but may also increase the porosity of sediments.
The precipitation of schwertmannite, a common metastable mineral found in extremely acidic environments, is the product of abiotic reactions (Sánchez-España et al., 2007, 2011), but field and experimental evidence suggests that microbial populations may enhance its nucleation and stability. Furthermore, the removal of toxic metals may be naturally enhanced by schwertmannite and ferrihydrite, depending on the pH (Carlson et al., 2002; Sánchez-España et al., 2006, 2011). The secretion of cohesive EPS by microbial biofilms also influences sediment entrainment, transport rate, and dimensions, even where thick surficial biofilms and microbial mats are not present (Garwood et al., 2013; Malarkey et al., 2015).
In general, AMD environments exhibit not only mineral–microbial interactions but also a notable diversity of morphologically diverse microbial mats promoted by steep geochemical gradients (Blackmore et al., 2018). Most organisms, except euryhaline species, are unable to cope with environmental extremes in AMD settings, and thus, bacterial proliferation increases. The occurrence of Acidithiobacillus ferrooxidans, Acidithiobacillus thiooxidans, Leptospirillum ferriphilum, Leptospirillum ferrooxidans, Gallionella ferruginea, Thermoplasmata, Ferrovum, Sulfobacillus, and Thiobacillus has been described from AMD around the world (Benner et al., 2000; Bruneel et al., 2006; Sánchez-España et al., 2007; Goltsman et al., 2009; Schippers et al., 2010; Korehi et al., 2013; Williams et al., 2017; Bomberg et al., 2019), including eubacteria from highly acidic environments (Benner et al., 2000; Korehi et al., 2013; Johnson and Aguilera, 2016; Johnson and Quatrini, 2016). Molecular studies from acid waters have shown that they are enriched in filamentous algae, cyanobacteria, diatoms, as well as protozoans and eukaryotic communities across all distinct microhabitats (Mesa et al., 2017). The most common phototrophs correspond to green microalgae (Dean et al., 2019). Among eukaryotes, green algae, diatoms, and euglenoids are among the most represented, but ciliates, cercozoans, rotifers, amoebas, stramenopiles, and fungi have also been reported (Packroff and Woelfl, 2000; Brake et al., 2001, 2004; Aguilera et al., 2007; Rawlings and Johnson, 2009; Aguilera, 2013; Massocato, 2018). Open-air water samples contain diverse eukaryotes from the fungal group Alveolata, Stramenopiles, and Opisthokonta.
Pine needles are another essential component of TIFs, not only in terms of size and textures but also as an additional source of organic matter and carbon, as corroborated in this work. The presence of an ochre colloidal mixture (mostly schwertmannite) and green algal mats, mainly composed of Euglena mutabilis and pine needle debris (LeBlanc et al., 1996; Sánchez-España et al., 2005c), is common in AMD environments but was not detected inside the sampled TIFs, probably due to marked seasonal changes in its population (Hasiotis and Brake, 2019).
5.4. Analogues of Archean Earth
Clastic iron stromatolites from AMD may be used as valuable modern analogues to infer sedimentary environments, before and after the Global Oxygenation Event (GOE). AMD systems may represent modern analogues of Paleoproterozoic continental surfaces during the Mesoproterozoic times, just when the pre-Cambrian BIFs were deposited (Brake et al., 2002, 2014; Parenteau and Cady, 2010; Westall et al., 2015; Kanellopoulos et al., 2019). Before the GOE, pyrite would accumulate as placer deposits on continental surfaces during the Archean (Johnson, 2014). Habitats that resembled AMD probably existed for billions of years. Once oxygen became available, the oxidation of pyrite promoted the eventual sedimentary riverine transport that delivered iron and sulfate to the oceans (Havig et al., 2017).
BIFs have also been interpreted as sedimentary files of transient atmospheric conditions, during the Archean. Paleoproterozoic oceans were rich in Fe(III) (Poulton and Canfield, 2011) under anoxic atmospheric conditions with widespread surficial oxygenation at archean ocean margins (Kendall et al., 2010; Konhauser et al., 2011). For instance, the Biwabik BIFs (1.85 My) are the most extensive and economically important iron deposits worldwide, and they were abundant at the end of the Archean. Other Precambrian low-pH environments generated by volcanism may have been crucial for the origin of life on Earth (Phoenix et al., 2006).
5.5. Natural bioremediation through geomicrobiology
Several years of mining in the IPB, one of the most extensive metal sulfide ore deposits regions in the world (Sáez et al., 1999; Johnson, 2014), has resulted in enormous metal-rich wastes and eventual ecosystem degradation by AMD of the main river systems that run over the IPB, which include the Tinto and the Odiel rivers (Grande et al., 1999; Nieto et al., 2007; Sarmiento et al., 2008, 2018; Shuster et al., 2017). When AMD sulfides are exposed to oxygen and water, they dissolve and produce H2SO4 and Fe(II) (Nordstrom and Southam, 1997). These reactive chemical species acidify as the stream flows down and the pH decreases. When the pH drops to 3.5 or even 2.5, the toxicity and metal mobility are higher (Sánchez-España et al., 2005a). This toxicity creates, in turn, new niches for acidophiles (Konhauser, 2007; Johnson and Quatrini, 2016), such as A. ferrooxidans or L. ferrooxidans (Rohwerder et al., 2003). Since mineral matrices provide suitable substrate for bacterial colonization, AMD hold great potential in bioremediation (Verb et al., 2001; Chockalingam and Subramanian, 2006; Florence et al., 2016; Byrne et al., 2018; Dean et al., 2019) and biopharmacy (Oberholster et al., 2013; Ayangbenro et al., 2018). Additionally, the bioleaching process of some toxic metals is a common phenomenon in AMD systems (Johnson, 2014). In fact, biosorption techniques employ the synergy between microalgae that supply organic matter and heterotrophs as CO2 suppliers (Rohwerder et al., 2003; Rawlings and Johnson, 2009; Florence et al., 2016; Abinandan et al., 2018). The biological removal of toxic metals by Ferrovum myxofaciens through biosorption (Hedrich et al., 2011) and the biosorption of Ni(II) and Pb(II) by fungal biomass (Aytar et al., 2014) are good examples. Biological strategies to cope with heavy metals found in AMD microorganisms include efflux-mediated metal transport, metal exclusion, intracellular and extracellular sequestration, and enzymatic detoxification (Denef et al., 2010). E. mutabilis shows a metal-fixing behavior in the formation of accretionary organic sedimentary structures in AMD settings (Brake et al., 2002, 2004, 2014). The transparency of the same filaments in conjunction with black and yellow colorations (Fig. 10) suggests an active process of bioleaching and biosorption for sulfur precipitates at the periplasmic membrane. The unidentified filament that presumably takes sulfur from the surroundings is similar in shape, size, and behavior to the small filaments reported in the work of Mori et al. (2017), who identified them as a novel Eubacterial genus (Acidithrix). This study also confirms a differential sulfur precipitation that is mainly concentrated at filament junctions and in specific locations (Fig. 10g–j).
5.6. Astrobiological implications
Modern microbial colonization of such extreme environments is a consequence of the biogeochemical gradients generated through the stream channel. The occurrence of iron stromatolites as TIFs in Tintillo is not only scarce but also peculiar in their precipitation and origin. Because they provide new sedimentary, mineralogical, and chemical biosignatures, microbial consortia inhabiting highly acidic extreme environments hold a great industrial potential in bioremediation and biopharmacology; more importantly, they are also recognized as excellent analogues of ancient Mars environments (Amils et al., 2007, Amils and Fernández-Remolar, 2014). The present study shows that a broad spectrum of textures and colors, enhanced by sediment topology, is also an empirical indication of sharp geochemical gradients.
TIFs are astrobiologically relevant because of their similarity with geological structures found on Mars (Brake et al., 2004; Fernández-Remolar and Knoll, 2008; Loiselle et al., 2018). For instance, the exposed rocks in the Meridiani Planum suggest arid, acidic, and oxidizing environmental conditions, which probably preserve mineralogical biosignatures (Fernández-Remolar et al., 2005; Knoll et al., 2005; Amils et al., 2007; Fernández-Remolar and Knoll, 2008; Loiselle et al., 2018). Furthermore, the relatively high amount of sulfate and hematite may indicate the presence of schwertmannite on the Red Planet (Des Marais et al., 2008; Bishop et al., 2015). Hydrated minerals and sulfates are also common on the Valles Marineris of Mars surface (Kaplan et al., 2016). Hence, Tintillo TIFs represent an excellent modern analogue of BIFs and a promising extreme environment to search for microbial biosignatures. Life in extreme environments enables us to understand the versatility of metabolisms in prokaryotes and explore for possible life beyond Earth (Westall et al., 2013; Preston et al., 2020).
The diversity of minerals, microorganisms, biofabrics, and geomorphological structures in the Tintillo AMD system provides multiple scales of observation with which to identify biosignatures, some of which are the result of pervasive interaction between abiotic and biotic factors (Fig. 13).

Multiscale comparison of morphological targets relevant in astrobiology (modified from McKay, 1992).
There is a continuous need to develop criteria and standards from microscopic to planetary scales in Astrobiology (Cady et al., 2003; Des Marais et al., 2008; Hays et al., 2017) to facilitate examination and identification of sedimentary contexts across multiple spatial and temporal scales, in particular to distinguish abiotic from biotic substances, objects, and patterns (Chang et al., 2019). The recognition of biogenic landforms and a lateral comparative assessment of diverse terrestrial environments provides guidance for future space exploration (Ibarra and Corsetti, 2016; Corenblit et al., 2019). Under the rationale that complexity arises from gradients (Spitzer et al., 2015), large-scale attributes such as morphology and complexity patterns are still needed to delineate the astrobiological quest, which may include extreme environmental interfaces as well as large-scale geological processes.
The main challenge in Astrobiology is the recognition of life as we know it, but also as we do not expect it. This implies that geological environments may abound in geochemical gradients that produce gradational colors and morphological patterns. Other challenges faced by astrobiology include the cost of high-resolution equipment and the robotic exploration time of targeted sites that store, retrieve, analyze, and visualize spatial data in Geographic Information System (GIS) applications and map services (Macfarlane et al., 2018; McKay, 2020). Such large-scale attributes as topography, geomorphology, and coloration that reflect geochemical gradients, temperature, or geochemical gradients can be assessed by GIS. Since a morphological recognition precedes topographic and cartographical studies, and this mega-scale recognition precedes the macro-, meso-, and microanalysis for characterization of biosignatures, a strong emphasis should be placed on morphological approaches in studies derived from extreme environments.
Footnotes
Acknowledgments
We thank the kind suggestions from Prof. Stepko Golubic and from one anonymous Reviewer. We are very thankful for all the professional help and kindness of the Astrobiology Editorial Team. We also thank Prof. Gloria Arjona (Caltech) for academic editing. The authors are very grateful to Prof. Eduardo Mayoral for fruitful discussions and field photography.
Author Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This paper was partially supported by Research Project CGL2015-66835-P of the Ministerio de Ciencia e Innovación of Spain. Publication costs were partially covered by UANL-PAICyT CN1246-20.
Abbreviations Used
Associate Editor: Russell Shapiro