
Abstract
Ancient veins of calcium sulfate minerals (anhydrite, bassanite, and gypsum) deposited by subsurface aqueous fluids crosscut fluviolacustrine sedimentary rocks at multiple localities on Mars. Although these veins have been considered an attractive target for astrobiological investigation, their potential to preserve biosignatures is poorly understood. Here, we report the presence of biogenic authigenic pyrite in a fibrous gypsum vein of probable Cenozoic emplacement age from Permian lacustrine rocks in Northwest England. Pyrite occurs at the vein margins and displays a complex interfingering boundary with the surrounding gypsum suggestive of replacive authigenic growth. Gypsum-entombed carbonaceous material of probable organic origin was also identified by Raman spectroscopic microscopy in close proximity to the pyrite. Spatially resolved ion microprobe (SIMS) measurements reveal that the pyrite sulfur isotope composition is consistently very light (δ34SVCDT = -30.7‰). Comparison with the sulfate in the vein gypsum (δ34SVCDT = +8.5‰) indicates a fractionation too large to be explained by nonbiological (thermochemical) sulfate reduction. We infer that the pyrite was precipitated by microorganisms coupling the reduction of vein-derived sulfate with the oxidation of wall-derived organic matter. This is the first evidence that such veins can incorporate biosignatures that remain stable over geological time, which could be detected in samples returned from Mars.
1. Introduction
Despite inhospitable conditions today and for the past few billion years, there is now overwhelming evidence that Mars once hosted stable bodies of surface water, which were probably habitable when life arose on Earth (Fassett and Head, 2011; Grotzinger et al., 2014). NASA's Mars 2020 rover (Perseverance) and the ESA–Roscosmos ExoMars rover (Rosalind Franklin) will both seek evidence of ancient life preserved in rocks and minerals associated with these water bodies (Westall et al., 2015; McMahon et al., 2018a; Lakdawalla, 2019). Possible targets for in situ (bio)geochemical analysis and/or the extraction of samples for analysis on Earth include veins (mineralized fractures) composed of calcium sulfate minerals such as gypsum (CaSO4·2H2O), bassanite (CaSO4·½H2O), and anhydrite (CaSO4). Calcium sulfate veins have frequently been encountered crosscutting Noachian–Hesperian sedimentary rocks in the craters explored by NASA's Mars Exploration Rovers (MER) and Mars Science Laboratory (MSL) (Squyres et al., 2012; Arvidson et al., 2014; Nachon et al., 2014; Kronyak et al., 2019). These minerals have also been detected in other forms elsewhere on Mars: the OMEGA imaging spectrometer aboard ESA's Mars Express orbiter detected gypsum in dune fields at ∼80°N (Langevin et al., 2005); and the CRISM spectrometer aboard NASA's Mars Reconnaissance Orbiter observed bassanite at 23°N in the Mawrth Vallis region (Wray et al., 2010) and stratabound gypsum in Noachian terrain at 29°S (Wray et al., 2009).
Should calcium sulfate mineral veins be prioritized in the search for biosignatures on Mars? It is generally agreed that geological materials on Mars must meet two criteria to be attractive targets from an astrobiological point of view (e.g., Westall et al., 2015; Hays et al., 2017; McMahon et al., 2018a; Beaty et al., 2019). First, they must have formed in habitable paleo-environments. Second, they must have a strong potential to record biosignatures—morphological, compositional, molecular, mineralogical, or isotopic—and to protect them from degradation over geological time.
Calcium sulfate minerals are likely to meet the first criterion. On Earth, these minerals are familiar as evaporites, that is, minerals that precipitate from saline waters where evaporation outpaces fluid resupply, typically in restricted basins and arid regions where halophiles commonly thrive. Evaporite minerals precipitate in order of increasing solubility, beginning with calcite or aragonite (CaCO3) followed by calcium sulfate, usually in the form of gypsum, which dominates many primary evaporite assemblages (Dean, 2014). Gypsum and anhydrite also occur as diagenetic nodules and as veins formed by the dissolution, redistribution, and reprecipitation of calcium sulfate in the subsurface. Hydrated calcium sulfates are of particular astrobiological significance since they typically record habitable conditions, that is, water activities, temperatures, and other physicochemical conditions well within the limits of microbial tolerance (Tosca et al., 2008; Nachon et al., 2014). Variably hydrated calcium sulfate veins crosscutting Noachian–Hesperian fluviolacustrine sedimentary rocks on Mars have thus been interpreted, together with other evidence, as evidence that the paleoenvironment was habitable (Grotzinger et al., 2014).
It is less clear that calcium sulfate veins on Mars meet the second criterion noted above, that is, the capacity to preserve ancient biosignatures (McMahon et al., 2018a). On the one hand, there is a growing body of evidence that beds, crusts, and cements formed of gypsum and other evaporite minerals can preserve a variety of biosignatures on Earth, including permineralized microfossils (Schopf et al., 2012; Dela Pierre et al., 2015), stromatolites (Allwood et al., 2013), organic compounds (Aubrey et al., 2006), and even viable microbial colonies enclosed by pockets of water on crystallographic cleavage planes (Parnell et al., 2004). On the other hand, sulfate veins formed in the subsurface after the lithification of the host rock may be less likely to entomb biomass or preserve such biosignatures than bedded sulfates formed by nucleation onto the sediment-water interface, or even shallow diagenetic nodules. Veins form after the dissolution, remobilization, and re-precipitation of nodular or bedded sulfates, processes which may exclude, degrade, or destroy any biosignatures originally present. Geologically ancient biosignatures have not hitherto been reported from calcium sulfate occurring as veins, although veins of other minerals have yielded fossils of subsurface-dwelling organisms (McMahon and Ivarsson, 2019). Weng et al. (2018) reported that gypsum veins exposed in the Atacama Desert contain organic compounds of possible biological origin, but the source and age of these compounds is unclear. The lack of evidence for biosignature preservation in ancient calcium sulfate veins motivated our study of fibrous gypsum veins of probable Cenozoic age hosted by late Permian lacustrine mudstones (∼0.25 Ga) from Kirkby Thore, Cumbria, NW England.
2. Materials and Methods
2.1. Gypsum vein sample
The gypsum vein analyzed here was collected in hand samples underground in situ from the Kirkby Thore gypsum mine, which accesses the Eden Shales Formation of the Cumbrian Coast Group in the Vale of Eden Basin, Cumbria, NW England. Gypsum has been mined here for over 100 years (Hughes, 2003). It occurs in numerous veins parallel to bedding in dark grey mudstone and derives from gypsum originally deposited in an evaporitic lacustrine basin or coastal sabkha in the late Permian. The gypsum is white and fibrous (“satin spar”), with fibers oriented normal to the vein walls, which are themselves also impregnated with gypsum cement. The veins are displacive and grew antitaxially, that is, by successive additions at the reopening and resealing margin between the vein and the host rock, causing fragments of the wall rock to be included along irregular median planes within the veins.
The age of the veins and their emplacement mechanism are poorly constrained, although the fibrous habit suggests a key role for fluid overpressure (i.e., fluid pressure in excess of hydrostatic pressure) and hydraulic fracturing (Parnell et al., 2000). Burgess and Holliday (1974) interpreted the veins in the Eden Shales Formation as mineralized hydraulic fractures formed by compaction-related pore-fluid overpressure during the burial of the host sediment. However, the wall rock fragments included within the veins are as fully compacted as the outside matrix, suggesting that veining postdated compaction. Moreover, it is now widely thought that gypsum veins can form during uplift as a consequence of fluid overpressure during tectonic compression and/or the rehydration (dissolution and reprecipitation) of anhydrite beds, which causes a volume expansion of >60% and associated hydraulic fracturing by sulfate-rich fluids. This mechanism has been invoked to explain the occurrence of gypsum veins in other Permo-Triassic evaporite basins in the west of England, which experienced Cretaceous and Cenozoic compression and uplift (Murray, 1964; Philipp, 2008; Meng et al., 2017). In the Vale of Eden, uplift began in the Cretaceous (Fig. 1). Fluid-inclusion microthermometry and palaeomagnetic data from the Penrith Sandstone Formation conformably underlying the Eden Shales Formation indicate a cooling trend beginning at that time from a maximum temperature of ∼120°C (equivalent to a depth of about 3–4 km; Turner et al., 1995). The Eden Shales Formation was uplifted through the 50°C isotherm and to depths of <1 km approximately 50 million years ago. Since anhydrite typically rehydrates to gypsum under these conditions (Zanbak and Arthur, 1986), it is likely that vein emplacement occurred at this time (and at this temperature), although an earlier or later origin cannot be ruled out.
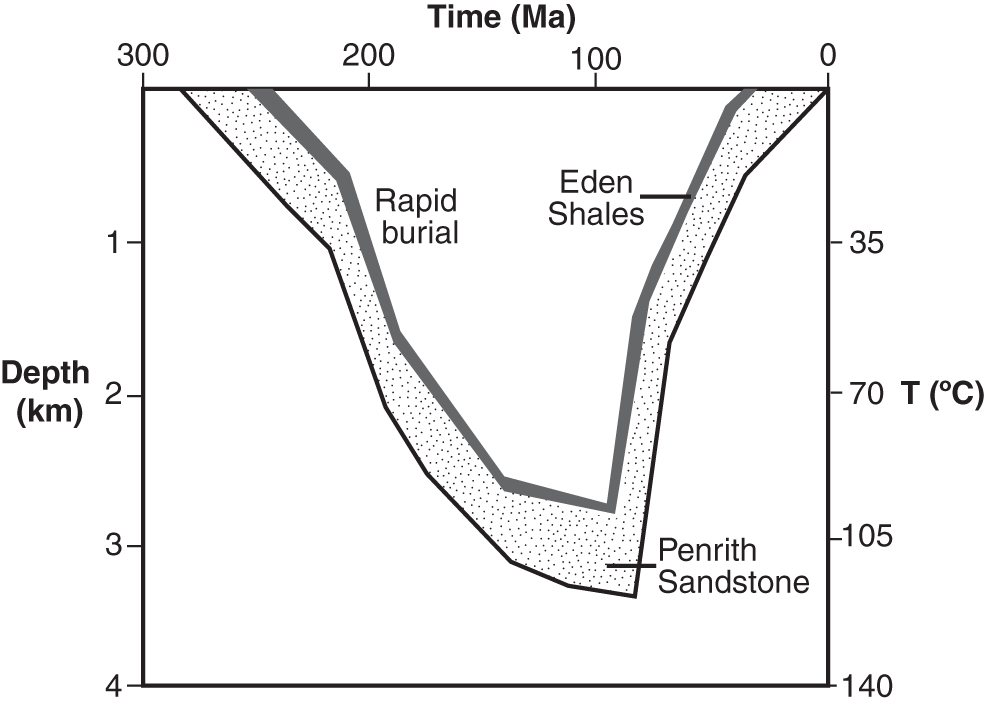
Sketch of the burial history of the Eden Shales and Penrith Sandstone Formations. Gypsum veins in the Eden Shales Formation could have formed either during burial or during uplift as a consequence of hydraulic fracturing. Adapted from Turner et al. (1995).
2.2. Optical microscopy
Optical microscopy was undertaken with a Leica DMLP reflected/transmitted light polarizing microscope with DFC 420C camera and Leica Application Suite v 4.00.
2.3. Scanning electron microscopy and energy-dispersive X-ray spectroscopy
Scanning electron microscopy (SEM) and energy-dispersive X-ray spectroscopy (EDS) of carbon-coated polished, uncovered thin sections were undertaken at the Imaging Spectroscopy and Analysis Centre (ISAAC) at the University of Glasgow using a Zeiss Sigma analytical scanning electron microscope equipped with an Oxford Instruments X-Max silicon drift energy dispersive X-ray detector and an operating voltage of 20 kV.
2.4. Total organic carbon
The carbon contents of the host mudstone were measured by a LECO CS225 elemental analyzer, after decarbonatization with hydrochloric acid.
2.5. Raman spectroscopy
Raman spectra were acquired from an uncoated chip of the gypsum vein and host rock by using a Renishaw inVia Raman microscope at the University of Edinburgh. The 514 nm, 2 mW laser beam was focused onto the samples with a 100 × objective lens, providing an excitation spot of about 2 μm diameter. Raman point spectra were taken at different positions on the samples over the range 100–3200 cm−1. The spectra were acquired with a 30 s exposure time at 50% laser power. Renishaw Wire 2.0 software was used for data acquisition.
2.6. Ion microprobe sulfur isotope analysis
Spatially resolved pyrite sulfur isotope compositions were measured on a sample of the Kirkby Thore gypsum vein with the Cameca IMS-1270 (#309) instrument at the School of GeoSciences at the University of Edinburgh, UK (the NERC Ion Microprobe Facility). Sulfur isotopes (32S and 34S) were analyzed as S- ions produced by bombardment of the target by a ∼1 nA, 133Cs+ primary beam accelerated at +10 kV resulting in a net impact energy of 20 keV at the sample surface. To eliminate charging during the analysis, the sample was coated with a ∼30 nm layer of gold, and the exact position of the primary cesium beam was flooded with low-energy electrons produced by the normal incidence electron gun. Secondary ions were accelerated at -10 kV and analyzed at a mass resolution sufficient to resolve molecular interference by 33S1H- and 34S1H2 - on the 34S- peak. An energy window of 40 eV was used. After pre-sputtering for 30 s, automated secondary ion beam alignment was performed by using the DTxy deflection plates to center the beam in the mass spectrometer field aperture (2,500 μm) and entrance slit (90 μm) positions.
Measurements of sulfur isotopes were made simultaneously in multicollection mode using two off-axis Faraday cups (L′2 for 32S and H′2 for 34S), which were calibrated at the start of the analytical session with in-built Cameca hardware and software. Count rates were typically ∼2 × 108 cps of 32S and ∼9 × 106 cps of 34S. The total acquisition time was 100 s comprising 20 cycles, split into two blocks of 10, with each cycle comprising a 5 s counting period. A single ∼4 min analysis (including 30 s pre-sputtering time and beam centering routines) resulted in an internal error of <0.25‰. Sulfur isotope compositions are reported as δ34SVCDT, calculated as
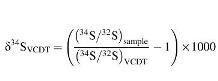
(VCDT = Vienna Canyon Diablo Troilite).
The Balmat pyrite standard (δ34SVCDT = +14.6‰; Crowe et al., 1990) from the University of Wisconsin was mounted in the same epoxy block as the sample and was analyzed five times at the start and five times at the end of the session so that any change in instrumental mass fractionation could be monitored and corrected for. This gave mean δ34SVCDT values of +14.73‰ and +14.97‰ before and after the sample analysis respectively, with standard deviations of 0.12 and 0.24. The average difference in the standard values before and after measuring the sample was used to apply a small instrumental drift correction to the sample values.
3. Results
3.1. Occurrence of pyrite and dolomite
Study of polished hand samples and thin sections of fibrous, bedding-parallel gypsum (“satin spar”) veins from Kirkby Thore revealed the presence of brassy, reflective mineral linings <1 mm thick embedded in gypsum at the interfaces between the gypsum veins and the wall rock (mudstone) above and below (Fig. 2). These linings do not occur either in the vein or in the host mudstone except near this interface. The mineral linings displayed the optical characteristics expected of a metal sulfide (opacity, brassy reflective luster; Fig. 3A–3C) but an unusually complex morphology consisting of disseminated patches of micrometer-scale crystals, rounded ∼5–25 μm globules, and growth structures 100 μm in scale consistently oriented outward from the vein margin, showing complex, micro-digitate, interfingering boundaries with the surrounding gypsum (e.g., top of Fig. 3C; right of center of Fig. 3D). The mineral was identified as pyrite (FeS2) by EDS analysis (Fig. 3E). Subhedral rhombohedral crystals of dolomite (CaMg(CO3)2; identified by EDS; Fig. 3F) were also observed close to the pyrite. In one instance a dolomite crystal appeared to enclose a pyrite grain (Fig. 3D, upper right).
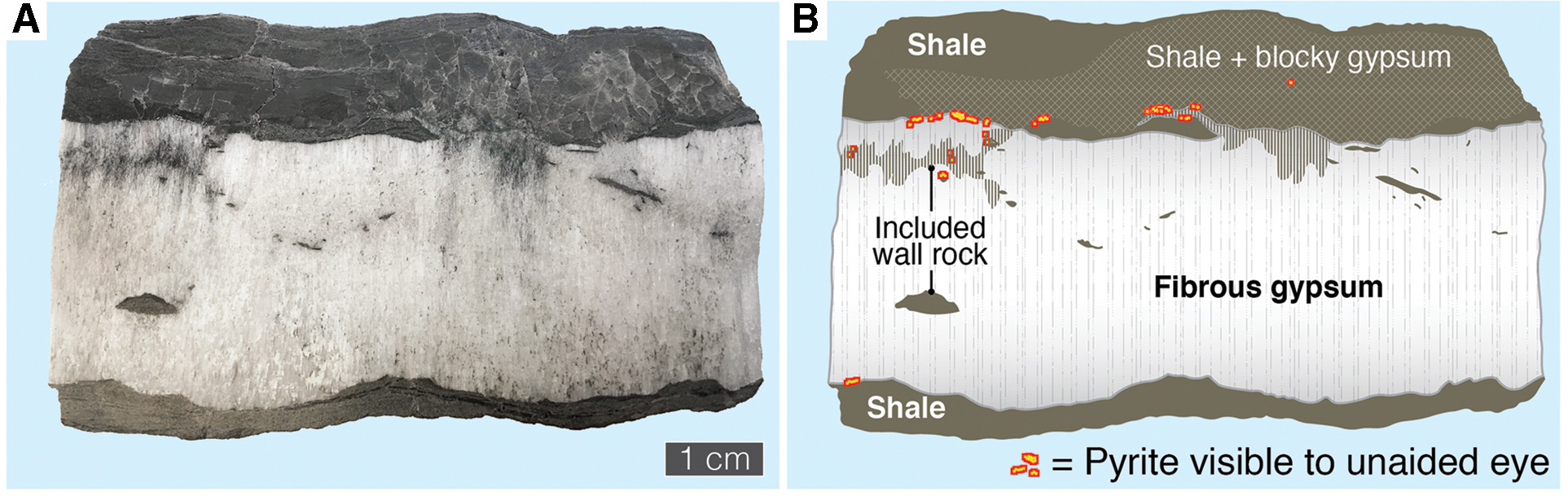
Macroscopic distribution of pyrite across a polished vertical cross section through a gypsum vein. (
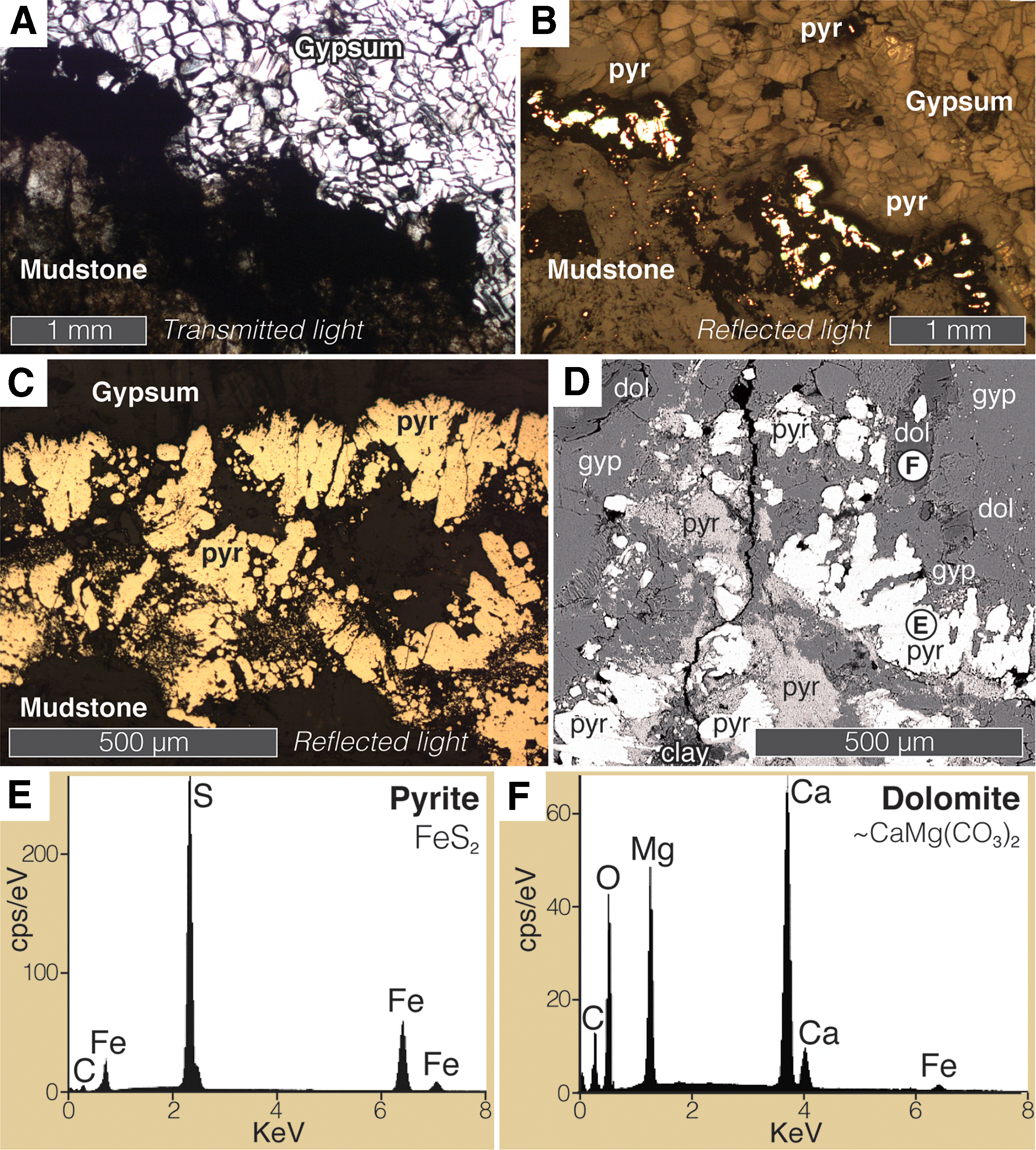
Pyrite and dolomite at the margins of a gypsum vein. (
3.2. Carbonaceous and organic matter
Samples of the mudstone hosting the gypsum veins were analyzed for total organic carbon content, yielding values of 0.22, 0.64, and 0.74 wt %; palynomorphs and phytoclasts were also observed within the mudstone using optical microscopy. The reflective pyrite exposed by polishing extends into the gypsum vein beneath the polished surface of the thin section, forming an opaque region bordered by diffuse dark material embedded within the gypsum (Fig. 4A, 4B). This material was investigated with Raman spectroscopy, which consistently detected bands at c. 1350 and c. 1600 cm−1, both in thin section and in rock chips, in addition to bands attributable to pyrite and gypsum (Fig. 4C). As discussed below, these observations suggest that the dark material included disordered carbonaceous material of probable organic origin. Clear gypsum within the veins was also analyzed, yielding only the expected peaks for gypsum.
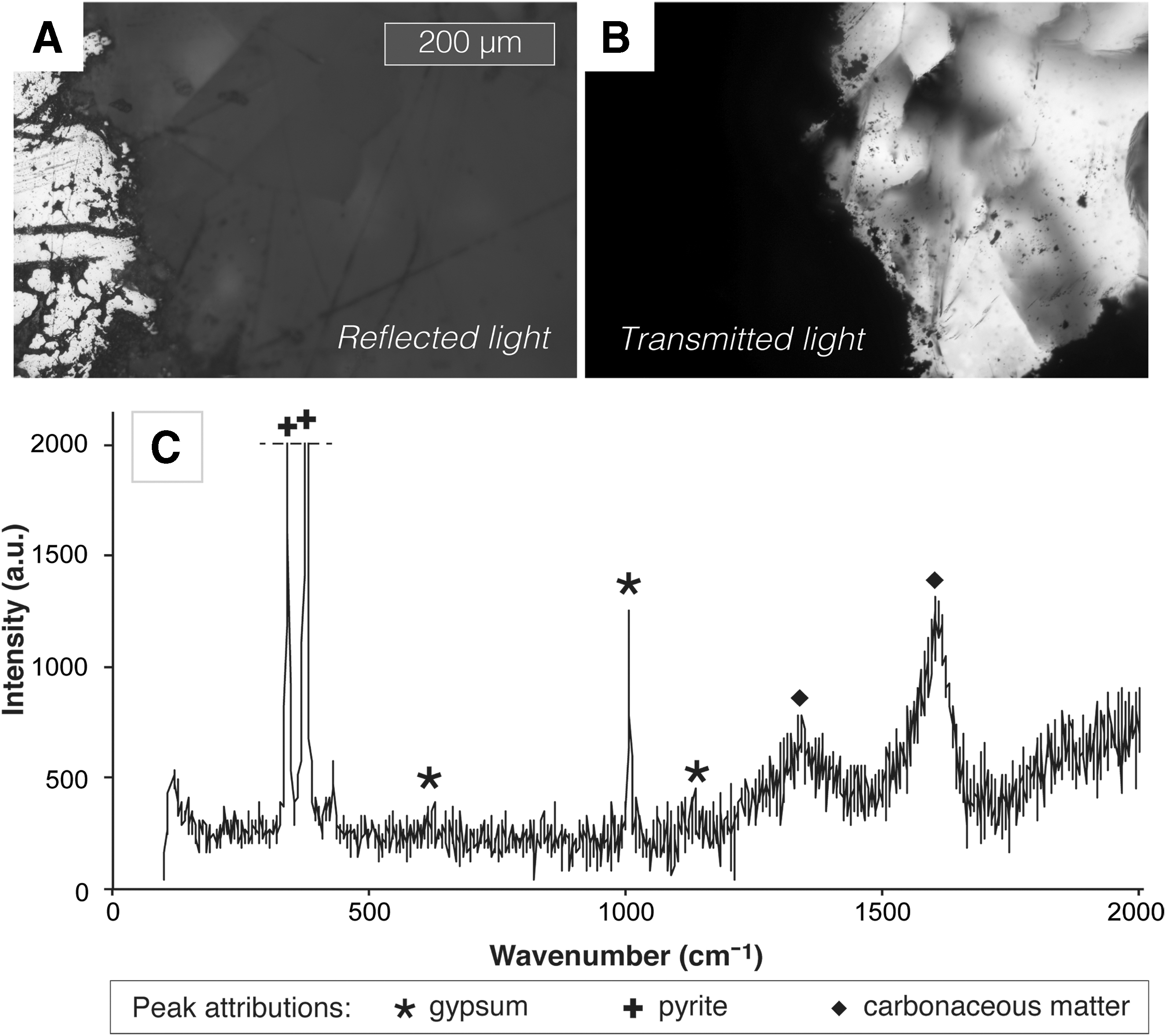
Pyrite-associated carbonaceous material. (
3.3. Pyrite sulfur isotope composition
Sulfur isotope compositions (δ34SVCDT) were measured at 13 points on a sample of the vein-margin pyrite, yielding values from -36.47‰ to -28.50‰ with a mean of -30.70‰ and a standard deviation of 2.19‰ (Fig. 5).

Reflected-light photomicrograph indicating sulfur isotope compositions (δ34SVCDT) measured at individual points on pyrite from a Kirkby Thore gypsum vein. The sample is a gold-coated, epoxy-mounted wafer with a pyrite grain from the Balmat standard mounted at its center (“Standard”). The edge of the vein is toward the bottom of the field of view in this image and is encrusted with pyrite (high relief). A well-preserved dolomite crystal also occurs in the lower right (low relief, “dol”). The dark marks associated with each measurement are pits made in the sample surface by the ion beam. The inset image shows a wider field of view; note the scale bar.
4. Discussion
4.1. Evidence for ancient subsurface gypsum-hosted life
To our knowledge, the present study is the first to report pyrite lining the walls of gypsum veins. This disseminated, microcrystalline pyrite occurs only in close proximity to the vein–wall interface, where it is largely embedded in the gypsum; it does not appear to be a preexisting constituent of the wall rock passively entrained into the vein by antitaxial growth. Rather, the pyrite appears to have precipitated authigenically in situ and thus retains its original growth habit, including complex interfingering with the surrounding gypsum. Although the two minerals appear to be intergrown, it is unlikely that pyrite and gypsum formed competitively at the same time, since they require reducing and oxidizing conditions respectively. We suggest instead that pyrite precipitated as a replacement of preexisting gypsum, which dissolved into reducing aqueous fluids percolating through microporosity along the vein margin, perhaps long after the vein formed.
Pyrite formation in low-temperature (<80°C) depositional and postdepositional (diagenetic) settings on Earth is typically a consequence microbial sulfate reduction (MSR) coupled to the respiration of organic matter or the oxidation of H2 (Machel, 2001; Seal, 2006). MSR produces H2S, which reacts with iron minerals to form the metastable precursors of pyrite (Raiswell and Canfield, 1998). Sulfate-reducing microorganisms are globally abundant in subsurface porewaters, where sulfate is commonly supplied by the dissolution of calcium sulfate minerals (Machel, 2001). Hence, we infer that pyrite in the Kirkby Thore veins derives from the microbial reduction of sulfate dissolved out of the gypsum veins. Microbial consumption of sulfate ions may in turn have promoted further gypsum dissolution, creating additional microporosity into which pyrite precipitated. Such a feedback process may explain the unusual dendritic morphology of the pyrite–sulfate interface, although this texture should not be regarded as a biosignature in itself (e.g., Fig. 3C, 3D). Sulfate reduction was probably coupled to the oxidation of organic matter in the pore-fluid or derived from the host mudrock, which contains ∼0.5 wt % total organic carbon and visible palynomorphs and phytoclasts. Exposure to water when this rock was hydraulically fractured would likely have facilitated diffusion of low-molecular-weight organic acids from the vein walls and entrained fragments into the sulfate-bearing fluid (Fredrickson and Balkwill, 2007). There is no evidence for the migration of exogenous hydrocarbons through these veins.
The authigenic, subhedral dolomite intimately associated with the pyrite may also be a by-product of sulfate reduction (Baldermann et al., 2015); speculatively, the necessary Mg2+ and CO3 may have derived from dissolution of dolostone elsewhere in the stratigraphic sequence (Hughes, 2003). Sulfate anions in solution can inhibit the precipitation of dolomite by forming complexes with Mg2+, but this effect requires medium-to-high sulfate concentrations and would therefore be suppressed by sulfate reduction, which may also favor dolomite precipitation by increasing alkalinity (Brady et al., 1996; Sánchez-Román et al., 2009). Indeed, Mg-sulfate complexes can also promote dolomite precipitation (Brady et al., 1996). Other factors favorable to dolomite formation include microbially induced local supersaturation of Ca2+ and Mg2+ and the presence of carboxylated organic matter (Petrash et al., 2017). The dolomite in the Kirkby Thore gypsum veins generally does not contain inclusions of pyrite, but there is one possible exception (Fig. 3D), which implies that dolomite may have precipitated (at least somewhat) later than the pyrite. In principle this could have occurred at any time in the later alteration history of the veins.
Sulfide minerals can also form abiotically via thermochemical sulfate reduction (TSR) at temperatures of ∼100–140°C and above (Machel, 2001; Watanabe et al., 2009; Meshoulam et al., 2016). Given that they most likely formed by anhydrite rehydration during uplift, the Kirkby Thore veins have probably not experienced such high temperatures, and our spatially resolved sulfur isotope measurements confirm that the pyrite is microbial in origin. MSR is associated with a kinetic sulfur isotope fractionation such that sulfur-34 is preferentially excluded from the reduced (i.e., sulfide) product. Where the supply of sulfate is not limiting, this leads to the formation of sulfides with markedly higher 32S/34S ratios than the original sulfate (usually expressed relative to a standard using the δ34S notation; see the Materials and Methods section). This effect can lead to sulfide δ34S values negatively offset by ∼20–45‰ (and occasionally more) compared to the original sulfate (Machel, 2001; Sim et al., 2011). TSR is also isotopically selective but induces a maximum sulfur isotope fractionation of only about 20‰ at ∼100°C (Thomazo et al., 2009; Watanabe et al., 2009; Meshoulam et al., 2016). Thus, highly negative δ34S values in sulfide minerals, especially where demonstrably >20‰ more negative than the precursor sulfate, are commonly considered diagnostic of MSR, that is, a biosignature (e.g., Thomazo et al., 2009). The Kirkby Thore pyrite yielded sulfur isotope values (δ34SVCDT) consistently around -30‰. These compare with δ34SVCDT values for the host gypsum veins of +7.6‰, +7.5‰, and +10.4‰ (mean 8.5‰), which were measured on samples of the same gypsum vein for a previous study (Armstrong et al., 2020; our Fig. 6) and are broadly in line with the expected sulfur isotope composition of Permian marine evaporites (Fig. 6; Claypool et al., 1980). Armstrong et al. (2020) reported similar values (δ34SVCDT = +8.8‰ and +9.6‰) from coeval veins at the Barrowmouth gypsum mine about 70 km west of Kirkby Thore, suggesting that regional variation is small. The sulfide–sulfate offset of about -38‰ at Kirkby Thore is incompatible with TSR and is interpreted here as a robust signature of ancient microbial activity. By contrast, sulfide veins of known hydrothermal origin in Northwest England have published δ34SVCDT values mostly between 0 and +10 (Fig. 6), much heavier than the pyrite analyzed here and consistent with TSR (Solomon et al., 1971).
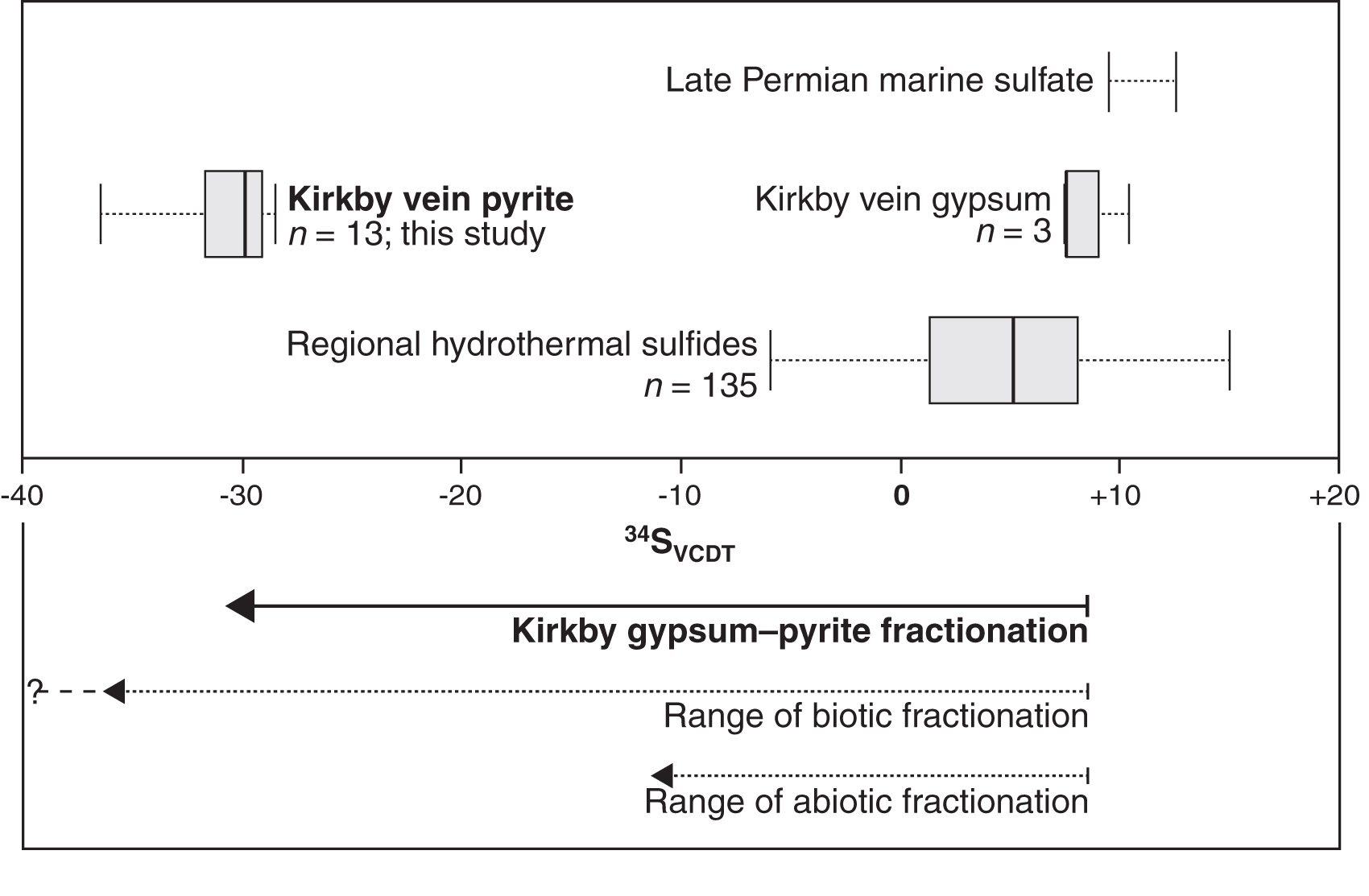
Box plot showing the sulfur isotope composition of pyrite and gypsum in the veins from Kirkby Thore, Cumbria. The gypsum data are reported in Armstrong et al. (2020). Sulfide mineral veins of known hydrothermal origin in the same region of England are shown for comparison (Solomon et al., 1971). Typical values for late Permian marine sulfate deposits are also indicated (Claypool et al., 1980). The lower panel illustrates the magnitude of the offset between gypsum and pyrite in the Kirkby Thore veins (bold arrow) compared to magnitudes known to be achievable by biotic and abiotic sulfate reduction (dotted arrows).
An apparent association between the pyrite and preserved carbonaceous material adds further weight to our interpretation of the sulfide as biotic in origin. Pyrite-rich opaque regions within the margins of the gypsum veins showed broad Raman bands at c. 1350 and c. 1600 cm−1 in addition to other bands attributable to pyrite and gypsum (Fig. 4). We identify the feature at 1350 cm−1 as the D band characteristic of disordered carbonaceous material, including organic matter (Beyssac et al., 2002). The feature at about 1600 cm−1 peaked between 1580 and 1620 cm−1 (e.g., Fig. 4C) and is likely to represent the graphitic G band produced by polyaromatic carbon (Beyssac et al., 2002), with a variable contribution from the D2 band. The carbonaceous material indicated by the Raman data is likely to be of ancient biological origin and appears to have been preserved in close association with the pyrite.
4.2. Comparison with calcium sulfate veins on Mars
Calcium sulfate veins observed crosscutting fine-grained siliciclastic rocks on Mars have important contextual, textural, and inferred genetic features in common with those from Kirkby Thore. For example, veins encountered by MER Opportunity in Endeavour Crater appear to be antitaxial and composed of fibrous gypsum, with fibers oriented normal to the vein walls (Squyres et al., 2012; Arvidson et al., 2014). White veins of calcium sulfate—anhydrite, bassanite, and possibly gypsum—also crosscut hundreds of meters of fluviolacustrine/eolian stratigraphy traversed by MSL Curiosity in Gale Crater, including the Yellowknife Bay and Murray formations (Nachon et al., 2014; Vaniman et al., 2014; Kronyak et al., 2019; Minitti et al., 2019). Most of these veins are oriented perpendicular to bedding (like those at Endeavour Crater), but at higher stratigraphic levels bedding-parallel and subparallel vein sets also occur (Nachon et al., 2014; Fedo et al., 2018; Kronyak et al., 2019). Like those in Kirkby Thore, the veins in the Yellowknife Bay formation are thought to postdate lithification and probably formed at depths of at least 1 km in rocks that underwent fairly rapid exhumation (Caswell and Milliken, 2017). In the stratigraphically higher Murray Formation, diverse mutually crosscutting vein sets record multiple generations of fracture opening and subsurface fluid circulation through a long window of geological time (Kronyak et al., 2019). Many of these veins are seemingly fibrous and are interpreted as generally syntaxial rather than antitaxial, although some appear to show entrained fragments of older material at boundaries between consecutive generations of sulfate, an indication of crack-seal growth shared by veins from Kirkby Thore (Fig. 7).

Photograph of the “Palmwag” calcium sulfate vein in the Murray formation in Gale Crater. This image was taken by the Mars Hand Lens Imager (MAHLI) aboard NASA's rover Curiosity on Sol 1275 of the MSL mission (Hurowitz et al., 2017). In common with gypsum veins from Kirkby Thore, England, this vein displays an apparent crystallographic preferred orientation normal to the vein walls, a variably distinct and asymmetric median line, and entrained fragments of older material. MAHLI image 1275MH0005550010404415C00. Scale bar (lower left) = 2 cm (after Kronyak et al., 2019).
4.3. Implications for biosignature detection on Mars
Discoveries in Earth's deep biosphere and its fossil record have inspired widespread speculation that life on Mars could also have colonized the subsurface, perhaps surviving there for billions of years after the surface became inhospitable (e.g., Boston et al., 1992; Fisk and Giovannoni 1999; Cockell, 2014). This line of research has been encouraged by multiple detections of methane as a trace gas (at ppbv level) in the martian atmosphere, where it is strongly out of equilibrium (Krasnopolsky et al., 2004; Mumma et al., 2009; Webster et al., 2015; Giuranna et al., 2019). It is unknown whether this methane is biological in origin, but methane-producing microorganisms are known to be globally abundant in Earth's subsurface. However, because Mars is a predominantly basaltic planet, studies of terrestrial analogs to a possible martian deep biosphere have mostly focused on porous basalt, where subsurface habitability could be maintained by water–rock reactions (serpentinization) that liberate hydrogen and thereby fuel both biotic and abiotic methanogenesis (e.g., McKinley et al., 2000; Ivarsson et al., 2008; McMahon et al., 2013). Although there is abundant evidence for a sediment- and sedimentary-rock-hosted deep biosphere on Earth, only a few studies have reported signatures of ancient subsurface life in sedimentary rocks or considered them relevant to Mars (Kretzschmar, 1982; Trewin and Knoll, 1999; Parnell et al., 2013; McMahon et al., 2018b). Our results imply that the porous margins of calcium sulfate veins in the subsurface can serve as conduits for the flow of sulfate-rich groundwater and therefore as potential habitats for prokaryotes capable of utilizing this sulfate to oxidize organic carbon (including methane) and/or molecular hydrogen. Such habitats may disappear and reappear several times over long spans of geological time as veins reopen under changing stress regimes and re-seal as further sulfate precipitates. Although in our samples MSR was probably stimulated by the organic content of the host rock, other sources of carbon and electrons may be available in organic-poor rocks. On Mars, pulses of groundwater circulation through sedimentary rocks could introduce methane and hydrogen produced by deeper serpentinization, and could perhaps also facilitate the production of hydrogen in situ through radiolysis or fracture-driven processes (e.g., McMahon et al., 2016; Dzaugis et al., 2018). Lithoautotrophic methanogens are found in sedimentary as well as basaltic rocks on Earth (e.g., Kimura et al., 2005). Thus, MSR coupled to hydrogen or methane oxidation could potentially have occurred in sulfate-bearing fracture or vein-margin fluids on Mars even if the host rock was relatively organic-poor, although organic-rich rocks are clearly preferable as sampling targets.
The veins from Kirkby Thore are younger than 0.25 Ga; by the time they formed, habitable conditions on Mars were already severely restricted, perhaps to depths of several kilometers on average; any early biosphere may have been extinguished by this time. However, the well-preserved calcium sulfate mineral veins observed on Mars formed close to 3.5 Ga ago when habitable conditions were probably widespread much nearer the surface (Westall et al., 2013; Grotzinger et al., 2014). If the vein-hosted martian habitats we envisage were actually inhabited, could they have produced durable, detectable biosignatures analogous to those described in the present study? Encouragingly, iron sulfide minerals have now been detected on Mars, although they are not yet well characterized. The Mössbauer instrument aboard MER Spirit made a tentative detection of FeS2 (as either pyrite or marcasite) in a hydrothermally altered basaltic rock at the Home Plate site in Gusev Crater (Morris et al., 2008). More recently, and more significantly for the present study, the CheMin XRD instrument aboard MSL Curiosity has suggested the presence of both pyrrhotite (Fe(1-x)S where 0 ≤ x ≤ 0.17) and pyrite in the sulfate-bearing Sheepbed mudstone (Yellowknife Bay formation) in Gale Crater; pyrrhotite was detected at 1.0 wt % in the “Cumberland” and “John Klein” drill samples, and pyrite was found at ∼0.3% (close to detection limits) in the latter sample, which is crosscut by hairline calcium sulfate veins (Vaniman et al., 2014). The sources of these sulfide minerals are unknown and may include igneous detritus and meteoritic material as well as sulfate reduction in the crater lake or its underlying sediment column (McAdam et al., 2014; Franz et al., 2017). Importantly, the Sheepbed mudstone is not pervasively oxidized and also preserves ancient organic matter (Ming et al., 2014; Freissinet et al., 2015). Thus, despite mineralogical evidence for sulfide oxidation in Gale Crater and elsewhere on Mars (i.e., the presence of jarosite; Zolotov and Shock, 2005; Rampe et al., 2017) and widespread oxidation of the martian surface in general, a small quantity of pyrite has survived locally alongside calcium sulfate and organic matter in at least some martian mudstones. The results of the present study recommend such materials for further in situ analysis and indeed return to Earth should they be encountered by future missions. We would additionally recommend that veins should be sampled where still encased in their host rocks rather than where they form free-standing “raised ridges,” since the vein–wall-rock interface is of key astrobiological interest. Drilling below the surface or sampling recently exposed materials may improve the probability of obtaining unoxidized martian sulfides in future missions.
On Earth, the δ34S biosignature has been identified in rocks as old as 3.47 Ga, although it did not become widespread until after 2.5 Ga (Shen et al., 2009; Thomazo et al., 2009). It must be conceded that sulfur isotope systematics are not well understood in martian contexts, and the isotopic fingerprints of any indigenous martian organisms may well have differed from life on Earth. Nevertheless, the Curiosity rover has already detected isotopically light sulfide (by evolved gas analysis) in Gale Crater sediments, where it could ultimately have originated either from abiotic or from biotic processes (Franz et al., 2017). The best available abiotic interpretation invokes the slow equilibration between aqueous sulfide and sulfate in an impact-driven subsurface hydrothermal system followed by the upward migration of fluids and the precipitation of sulfide minerals in the overlying sediment (Franz et al., 2017). Such a process would induce 34S-depleted sulfide mineralization throughout the accessible porosity of the host rock and not exclusively within microenvironments localized to the margins of preexisting calcium sulfate veins, as seen in our samples. Hence, multiscale analysis of the distribution of martian pyrite through its host rock could be essential for determining its origin. The analysis of 33S and 36S alongside δ34S could also constrain the biogenicity of martian sulfide and its relationship to adjacent sulfate minerals versus exogenous fluids (Ueno et al., 2008); these signatures would also discriminate between sulfide produced by sulfate reduction and that produced by sulfur disproportionation (Johnston et al., 2005; Ueno et al., 2008). Finally, evidence for ancient biological activity may also be obtained through the analysis of carbonaceous material associated with metal sulfides, as in the present study; both the Perseverance and Rosalind Franklin rovers will be equipped with Raman laser spectrometers as well as other instruments to investigate organic matter.
5. Conclusion
This study has described authigenic microbial pyrite and associated carbonaceous matter preserved in calcium sulfate veins crosscutting mudrocks. Our interpretation of these results suggests that calcium sulfate veins can create conditions conducive to subsurface microbial habitability, and also provides the first direct evidence that such veins can incorporate biosignatures capable of remaining stable over geological time. Such evidence may be detectable in samples returned from Mars, particularly if samples are taken from vein–wall-rock interfaces (rather than, e.g., the free-standing veins known as “raised ridges”). Calcium sulfate veins commonly postdate their host rocks by hundreds of millions of years (e.g., Meng et al., 2017). In Gale Crater, some are thought to have formed at depths of over 1 km (Caswell and Milliken, 2017). It is unclear whether mudrocks at the Perseverance or Rosalind Franklin landing sites have been exhumed from such extreme depths. Nevertheless, any biosignatures present in calcium sulfate veins crosscutting these rocks would have originated in a subsurface habitat markedly different from the depositional environment of the host mudrock. These rocks may therefore offer insights into two ancient martian habitats—surface and subsurface—for the price of one.
Footnotes
Acknowledgments
This work was funded by the European Union's Horizon 2020 Research and Innovation Programme under Marie Skłodowska-Curie grant agreement 747877. We acknowledge the Natural Environment Research Council for supporting the NERC Ion Microprobe Facility in Edinburgh and for supporting P.B.R.R.'s PhD studentship. The authors sincerely thank Mike Hall and John Craven for generous assistance with sample preparation and the operation of the ion microprobe; Alison McDonald and Andrey Gromov for technical assistance with Raman microscopy and analysis; Connor Brolly for assistance with the TOC analysis; and Stephen Phillips for originally collecting the vein sample. We also thank Diana Marosi for bibliographic assistance. We thank Tim Lyons and an anonymous reviewer for helpful comments that improved the manuscript.