
Abstract
The role of minerals in the events that led to the origin of life is discussed with regard to (1) their catalytic role for the formation of RNA-like oligomers from their monomers and (2) their protective role for organic molecules formed in space that were delivered to planetary surfaces.
Results obtained in the laboratory demonstrate that minerals do catalyze the oligomerization of ribonucleic acid (RNA) monomers to produce short RNA chains. Furthermore, and more importantly, these synthetic RNA chains formed by mineral catalysis serve as a template for the formation of complementary RNA chains, which is a significant finding that demonstrates the role of minerals in the origin of life.
Simulation experiments run under Mars-like conditions have also shown that Mars analog minerals can shield the precursors of RNA and proteins against the harmful effects of UV and gamma radiation at the martian surface and 5 cm below the surface.
Dedicated to the memory of Professor George W. Brindley
1. Introduction
T
Organic compounds form in the interstellar medium and are delivered to the surface of Mars and other planetary surfaces from space via meteorites, comets, and interplanetary dust particles. The average mass of organic molecules delivered over millions of years by just carbonaceous meteorites is estimated to be on the order of 2.4 × 108 g of organic carbon/year (Love and Brownlee, 1993; Flynn, 1996), which is augmented by organic matter in interplanetary dust particles (Schramm et al., 1989; Flynn, 1996), meteorites (Botta and Bada, 2002; Pizzarello et al., 2006, 2013), and comets (Llorca, 2005).
In a lecture delivered in 1947, John D. Bernal suggested that minerals and soils might have protected these organic molecules from the destructive effects of radiation. He also suggested that they might have served as catalysts in chemical evolution by adsorbing the monomers of biologically important molecules, that is, RNA and proteins, on their surfaces, thereby bringing them into a favorable orientation for reactions to occur (Bernal, 1949).
The “RNA world” model proposed that the first life on primitive Earth consisted of mainly RNA (Crick, 1968; Orgel, 1968). In Gilbert's RNA world hypothesis, RNA may have formed by the oligomerization of mononucleotides present on primitive Earth (1986).
Simulation experiments performed in the laboratory by irradiation of simple molecules such as H2O, CH3OH, CO, and NH3 in astrophysically relevant proportions demonstrated the formation of the building blocks of life (Allamandola et al., 1988; Briggs et al., 1992; Bernstein et al., 1995, 1999, 2002; Nuevo et al., 2009, 2012, 2014; Materese et al., 2013; Sandford et al., 2015; Smith et al., 2015).
The present study is a review of the possible role of minerals as catalysts for the formation of building blocks of life and their protective role for biomolecules against the effects of radiation in the events that led to the origin of life. A comparison of the theories on the origins of life, namely the Oparin-Haldane theory and the panspermia theories, has been discussed in detail in a previous work (Davis and McKay, 1996).
2. Formation of RNA Oligomers by Mineral Catalysis
In our origin-of-life studies, we have demonstrated in the laboratory that montmorillonite, a member of the phyllosilicate group minerals, catalyzes the oligomerization of monomers of ribonucleic acids, RNA, to form short RNA-like oligomer chains. Formation of the phosphodiester bond between RNA monomers requires the removal of a water molecule (or molecules) to shift the equilibrium in favor of oligonucleotide formation, which would be impossible for reactions that take place in dilute aqueous solutions as demonstrated in the following equation:
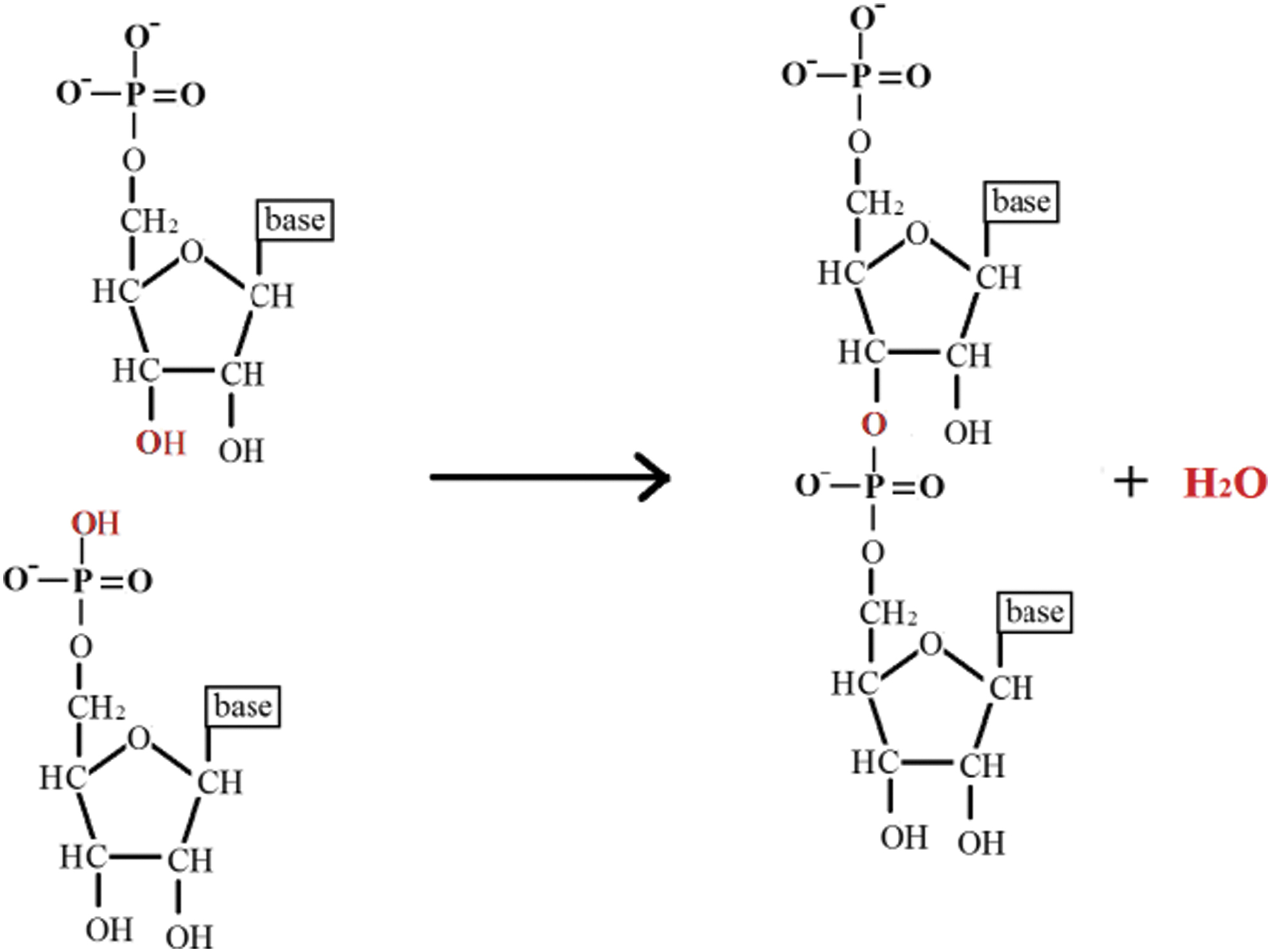
Phosphodiester bond forms by nucleophilic attack of the unpaired electron pair of the oxygen of one monomer on the phosphorus of another monomer.
It is important to note that montmorillonite-catalyzed oligomerization reactions, which led to the formation of short RNA-like chains, were run in dilute aqueous solutions, thus demonstrating the importance of the catalytic role of montmorillonite for the formation of the phosphodiester bond (Ertem, 2004).
Phyllosilicates, along with gypsum (CaSO4) and hematite (Fe2O3), are thought to have formed early in the planet's history, 4.6 to 4 billion years ago, when Mars was believed to have been warmer and wet (Bibring et al., 2006; Lowe et al., 2019; Weitz and Bishop, 2019).
The possible role of borate minerals, namely, colemanite (Ca2B6O · 5H2O) or (CaB3O4(OH) H2O), ulexite (NaCaB5O6(OH)6·5H2O), and kernite (Na2B4O7·4H2O), in the events that led to the origin of the RNA world has also been suggested in terms of the stability of the complexes that can form between the sugar moiety of RNA with borate minerals, though it can be concluded that there was not a sufficient amount of borate minerals present on the early Earth (Grew et al., 2011).
Here, we review our research on the catalytic role of montmorillonites for the formation of RNA-like oligomers, the formation of the complementary RNA chains on these synthetic RNA oligomers by template-directed synthesis, and the protective role of Mars analog minerals for bioorganic molecules against the destructive effects of radiation. This is the first literature report to review the template properties of a synthetic oligomer chain formed in the laboratory by mineral catalysis, which serves as a template for the formation of the complementary oligomer chain.
2.1. Structure of montmorillonite
Montmorillonite is a clay mineral with a layered structure; each layer is composed mainly of one octahedrally coordinated alumina sheet, Al2O3, which is sandwiched between two tetrahedrally coordinated silica sheets, SiO2, as shown in the work of Ertem (2004).
During the formation of montmorillonites by weathering, some of the aluminum ions in the octahedral sheet and silicons in the tetrahedral sheets may have been replaced by cations of lower valence. Charge deficiency that results from these isomorphic substitutions, namely, substitution of aluminum ions (Al3+) in the octahedral sheet by magnesium ions (Mg2+) and substitution of silicons (Si4+) in the tetrahedral sheet by aluminum ions (Al3+), is compensated by the interlayer cations held between the layers. In fact, montmorillonite owes its catalytic activity to these isomorphic substitutions.
Structural formula of the montmorillonite, commercially known as Volclay, used as a catalyst in our research for the formation of RNA oligomers was derived from its elemental analysis (Brindley and Ertem, 1971; Ertem et al., 2007) as shown below along with structure of pyrophyllite for comparison:
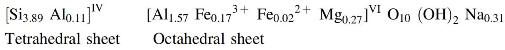
where IV and VI indicate the tetrahedrally and octahedrally coordinated cations, respectively.
Theoretically, the structural formula of montmorillonite can be derived from pyrophyllite, which has the structure shown below:
Without the presence of charge-deficient sites, montmorillonites would not have the capacity to expand and hold the exchangeable cations and organic molecules in their interlayer. The significance of isomorphic substitutions in montmorillonites is discussed in detail in the works of Grim and Kulbicki (1961), Schultz (1969), and Wolters et al. (2009).
The number of charge-deficient sites per 100 g of clay is called cation exchange capacity, CEC. As data presented in Table 1 demonstrate, there is a strong relationship between the extent of catalytic activity of montmorillonites to form RNA-like oligomer chains, the length of these oligomer chains, and number of charge-deficient sites, that is, CEC. While montmorillonites with a low charge density, 0.29, produce oligomer chains 10–20 monomer units long (Ferris and Ertem, 1992a; Ertem and Ferris, 1996), montmorillonites with a high layer charge density, 0.37, catalyze the formation of RNA oligomers that are only 4 monomer units long. Lower charge deficiency leads to longer separation distances between the charge-deficient sites on clay surfaces, which permits the elongation of nucleotide chains to a greater length. For montmorillonites with a higher charge deficiency, on the other hand, charge-deficient sites are closely positioned on the surface and prevent the elongation of oligonucleotide chains.
Mean Layer Charge of Montmorillonites per Formula Unit χ, Number of Monomer Units in the Longest Oligomer Chain Formed, and Their % Yields
χ was calculated per formula unit = 0.5 unit cell following the method developed by Lagaly.
Modified from Ertem et al. (2010), Table 3.
2.1.1. Preparation of controlled-charge montmorillonites
In dioctahedral montmorillonites, interlayer cations migrate into vacant octahedral sites upon heating the montmorillonite. The migration temperature depends on the nature and diameter of the interlayer cation. With montmorillonites saturated with Li+ cations, Li+ has a diameter of 134 pm and migration takes place at 220°C (Hoffmann and Klemen, 1950; Greene-Kelly, 1953, 1955; Glaeser and Méring, 1967, 1971). Due to this property, montmorillonites with desired expansion properties can be prepared in the laboratory by saturating the montmorillonite with a mixture of Li+-Na+ ions at calculated cation ratios (Brindley and Ertem, 1971).
2.2. Mineral-organic interactions
George W. Brindley is the pioneer in investigating the interactions of clay mineral montmorillonite with organic compounds using X-ray diffractometry, XRD. The presence of organic compounds in the interlayer region changes, that is, increases the interlayer spacing of montmorillonite (Brindley and Ertem, 1971; Ertem and Lagaly, 1978). The increase in the interlayer spacing is a function of the size of organic compounds that occupy the interlayer region (Lagaly and Weiss, 1970, 1971, 1976).
Max M. Mortland investigated the nature of interactions between montmorillonite and the organic compounds using mainly infrared spectroscopy (Mortland et al. 1963, 1966; Mortland and Boyd, 1989; Boyd and Mortland, 1990). These studies were followed by several publications by Pinnavaia and his coworkers (de Faria et al., 1998; Wang and Pinnavaia, 1998; Kim et al., 2000; Liu and Pinnavaia, 2002).
2.3. Binding of monomers onto montmorillonite and other minerals
We investigated the adsorption of nucleotides, that is, 5′-NMPs (building blocks of ribonucleic acids, RNA), nucleosides, and polynucleotides (short RNA chains) onto montmorillonite (Ferris et al., 1989a). The extent of adsorption of nucleotides and nucleosides onto montmorillonite varies with the structure and concentration of the organic compound, the pH of the solution, the presence and concentration of salts, and the nature of the mineral used (Ferris et al., 1989a).
In a recent work, Pedreira-Segade et al. (2018) investigated the interaction of building blocks of RNA and DNA with phyllosilicates under various physicochemical conditions such as temperature, pH, and salinity under ambient conditions and concluded that adsorption takes place via ligand exchange, where phosphate groups of the nucleotides interact with the hydroxyl groups of the phyllosilicate.
As the map taken by the OMEGA instrument on ESA's Mars Express demonstrates, iron oxide/hydroxide minerals are widely present on the martian surface (Bell et al., 1990). It plots the distribution of ferric oxides across the surface of Mars. 1
Because of the presence of iron oxide/hydroxide minerals on martian soil, we also investigated the possible catalytic role of iron oxide-hydroxide minerals, namely, akaganéite, which is an iron(III)oxide-hydroxide/chloride mineral with a structural formula of β-Fe3+O(OH-,Cl-), goethite, α-FeO(OH-), and hematite (Fe2O3) for phosphodiester bond formation. Neither goethite, akaganéite, nor hematite catalyzed the oligomerization of the phosphorimidazolide of adenosine (ImpA) (Holm et al., 1993).
3. Montmorillonite-Catalyzed Reaction of RNA Monomers to Form Short RNA-like Oligomer Chains
3.1. Reactions run in the presence of a condensing agent
We studied the montmorillonite-catalyzed oligomerization of an RNA monomer, namely, 5′-adenosine monophosphate, 5′-AMP, in aqueous solution in the presence of a condensing agent, 1-ethyl-3-(3-dimethylaminopyropil) carbodiimide (EDAC).
3.1.1. Experimental
Aqueous 5′-AMP solution containing EDAC was added to Na-montmorillonite, mixed, and left at room temperature for 3–5 days. Analysis of reaction products by high-performance liquid chromatography (HPLC) using a C-18 μ-Bondapak reverse-phase column demonstrated the formation of 3′,5′-linked dimers (RNA-like linkages) [3′,5′-(pA)2] at 11% and 2′,5′-linked dimers [2′,5′-(pA)2] at 19%, along with formation of about 5% of a dimer with pyrophosphate linkage, Ap5′pA. In the absence of montmorillonite, the reaction yielded only 1% of 2′,5′-dimer and 30% of Ap5′pA, clearly demonstrating the catalytic role of montmorillonite in the oligomerization reaction (Ferris et al. 1989b), Fig. 1.
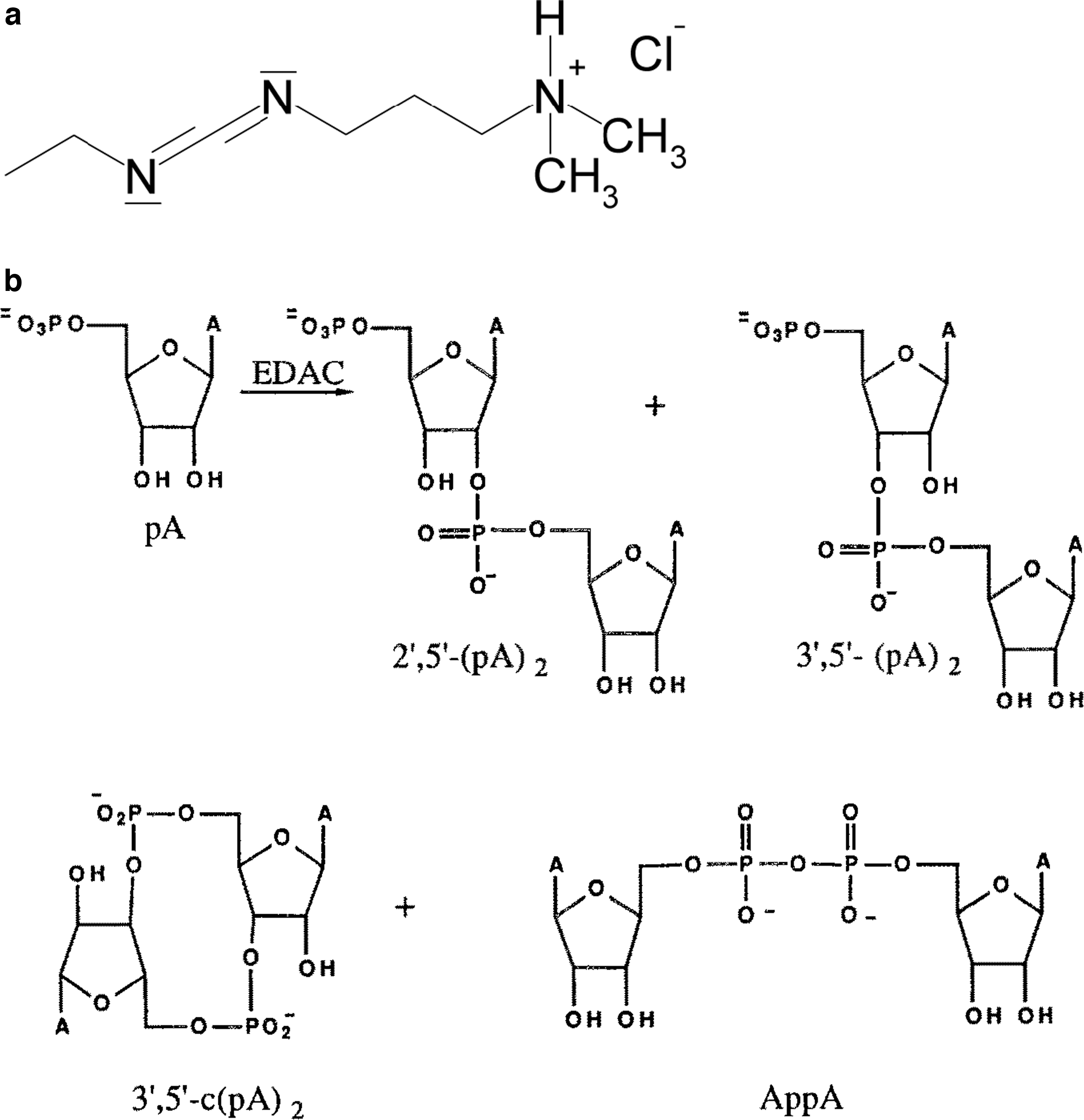
(
3.2. Reactions run with activated RNA monomers, ImpN
We extended our research by replacing the 5′-AMP with activated monomers of 5′-nucleotides, namely, 5′-imidazolide derivatives of 5′-AMP and 5′-CMP, ImpA and ImpC, respectively.
3.2.1. Formation of 5′-imidazolide derivative of 5′-AMP (ImpA)
Formation in the laboratory under prebiotic conditions was first demonstrated by Lohrmann (1977) with a yield of 10–50%, depending on the nature of the nucleotide. We synthesized the activated monomers by a modification of the procedure described in the work of Joyce et al. (1984a). Removal of strongly bound H2O from 5′-AMP was achieved by dissolution of 5′-AMP·H2O and imidazole in 20 mL of dimethylformamide and evaporation to near dryness in a rotary evaporator under vacuum at 40–45°C (Kay, 1990). The ImpNs obtained following this procedure were of 92–96% purity (Ferris and Ertem, 1992a). Figure 2 shows the structures of ImpC and the structure of a short oligocytidylate chain formed by montmorillonite catalysis.
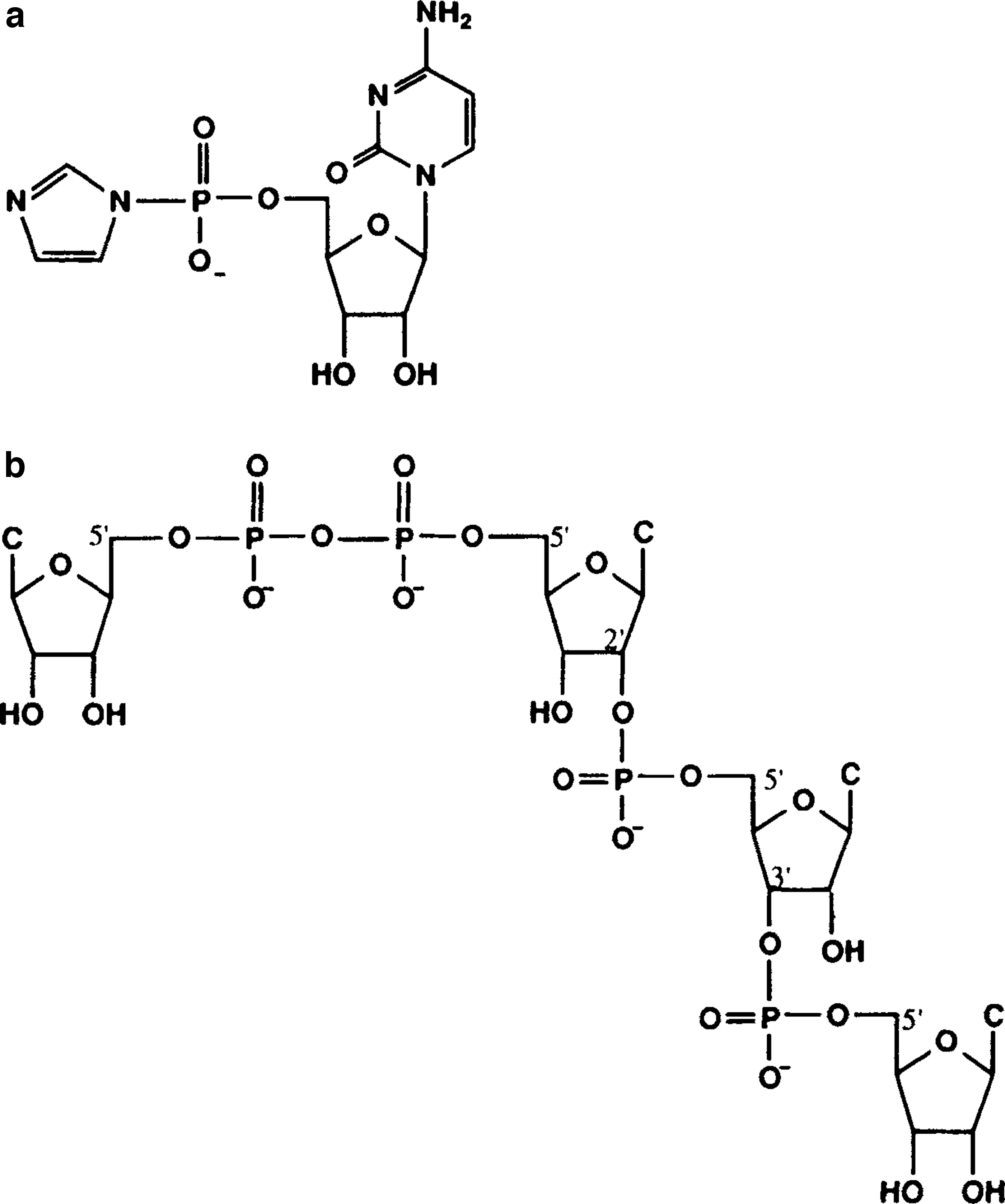
Structures of (
The montmorillonite-catalyzed reaction of activated RNA monomers ImpA and ImpC in aqueous solution at pH 8 produces oligomers containing 10–20 monomer units (Ferris and Ertem, 1992b).
Analysis of oligomer chains was performed by HPLC with a HEMA IEC anion exchange column (Stribling, 1991) and HPLC conditions described previously (Kanavarioti, 1997). While ImpA produces oligoadenylate chains, (pA) n , containing up to 10 monomer units (Ferris and Ertem, 1992a), montmorillonite-catalyzed oligomerization of 5′-imidazolide derivative of 5′-CMP (5′-cytidine monophosphate) ImpC produces oligomer chains containing up to 20 monomer units, Fig. 3. Furthermore, oligo(A)s containing a 3′,5′-linkage at the end of their chain elongate more rapidly compared to 2′,5′-terminus (Ferris and Ertem, 1993a). The course of montmorillonite-catalyzed ImpC reaction is shown in Fig. 4.
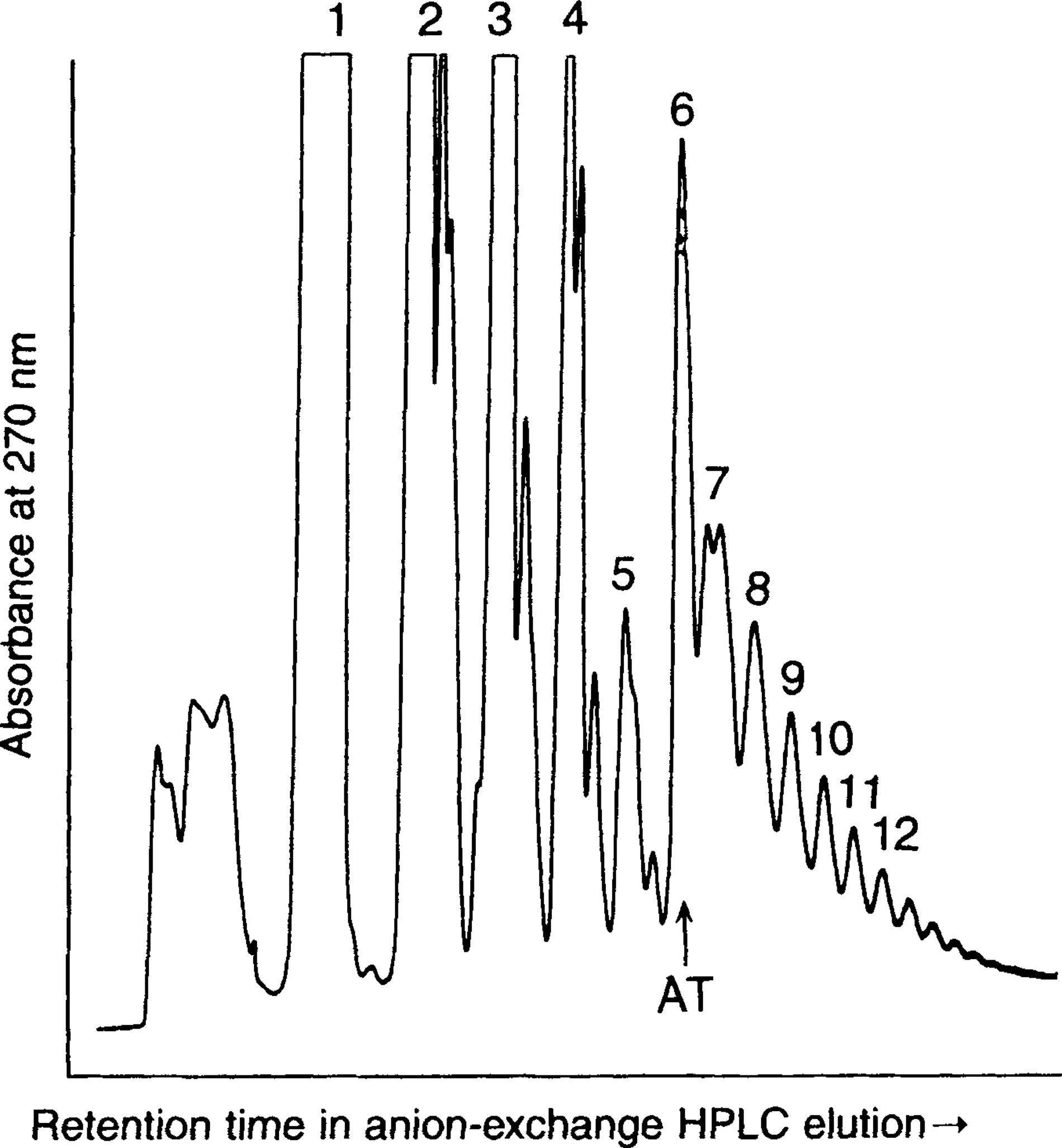
Anion-exchange HPLC elution profile of oligocytidylates formed in montmorillonite-catalyzed oligomerization reaction of ImpC. Oligomers formed in aqueous solution containing 15 mM ImpC in 0.2 M NaCl and 0.075 M MgCl2 within 5 days. Numbers 1, 2, 3 … 10, 11, 12 indicate the number of monomer units in the oligomer chain. AT = change in attentuation. (Data from Ertem and Ferris, 1996.)
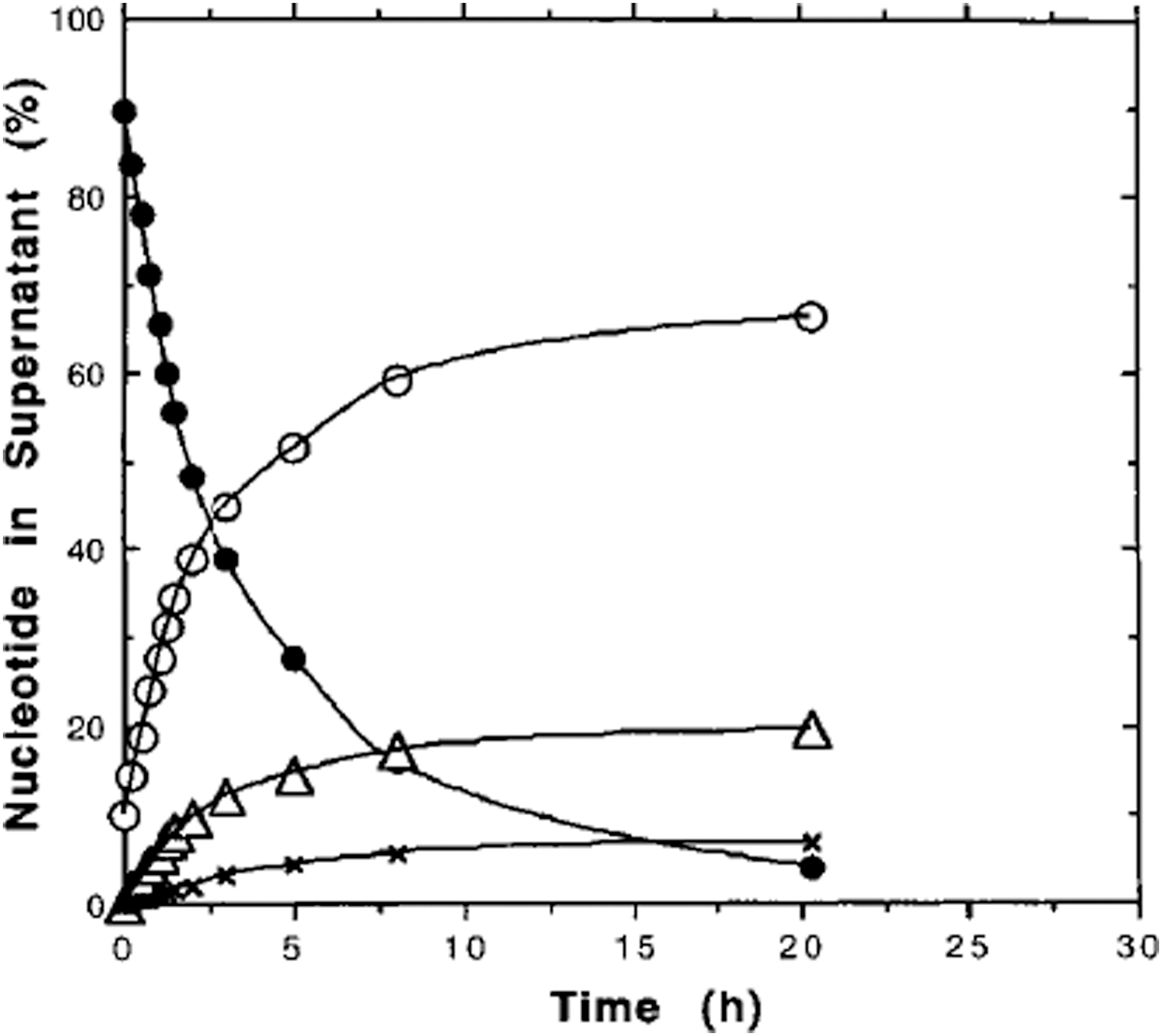
Course of ImpC oligomerization reaction was followed by taking a certain volume of reaction supernatant at regular intervals and analyzing it by HPLC. • = ImpC; ○ = monomers; ▵ = dimers; × = trimers. (Data from Ertem and Ferris, 1997.)
What is most significant is that oligocytidylates formed by montmorillonite catalysis serve as a template for the formation of complementary oligoguanylate chains from activated 2-methyl-ImpG, namely, 2-methyl imidazolide derivative of 5′-guanosine monophosphate (2-MeImpG; Ertem and Ferris, 1996). Synthetic oligocytidylates containing exclusively 2′,5′-linkages also serve as a template producing both 2′,5′- and 3′,5′-linked oligoguanylates. This is the first report in literature demonstrating that a synthetic oligomer chain formed in the laboratory by mineral catalysis can serve as a template for the formation of the complementary RNA oligomer chains.
Mineral catalyzed self-condensation of activated monomer of 5′-UMP, namely, ImpU in the presence of Na-montmorillonite produced oligomer chains 7 monomer units long (Ding et al., 1996).
Ferris et al. (1996) demonstrated the elongation of an oligonucleotide chain containing 10 monomer units, namely, 32pdA(pdA)8pA, to oligo(A)s containing 40–50 monomer units by daily addition of ImpA to the reaction mixture for 14 days in the presence of montmorillonite. In a similar manner, daily addition of ImpU to oligo(U) chains containing 10 monomer units produced oligomer chains 25–30 monomer units long (Ferris et al., 1996; Ferris 2002). They also demonstrated that non-expanding clay minerals illite (a dioctahedral, non-expanding smectite group mineral) and hydroxylapatite [Ca5(PO4)3(OH)] catalyzed the formation of long amino acid chains by daily addition of monomers to the reaction mixture.
Kinetic analysis of montmorillonite-catalyzed formation of dinucleotides and oligonucleotides from ImpA using the SIMFIT computer program demonstrated that the rate constants increased in the order of 2-mer <3-mer <4-mer …7-mer. More importantly, montmorillonite catalysis increased the rate constants for dimer and trimer formation more than 1000 times (Kawamura and Ferris, 1994).
3.3. Factors effecting the yields and length of the longest oligomers formed by montmorillonite catalysis
We studied the correlation between the extent of catalytic activity, layer charge deficiency, and charge distribution of montmorillonite(s) in a variety of phyllosilicates employing the procedures described previously (Lagaly and Weiss, 1970, 1971, 1976; Ertem and Lagaly, 1978; Ruehlicke and Kohler, 1981). Layer charge deficiency arises from the substitution of Al3+ and Si4+ in the octahedral and tetrahedral sheets, respectively, with the cations of lower valency. Results demonstrate that there exists a correlation between the extent of layer charge deficiency of montmorillonites and the extent of their ability to serve as catalyst for the formation of oligonucleotides, namely, short RNA-like chains. In montmorillonites with a lower charge deficiency (lower CEC, such as Vol montmorillonite and Swy-2 montmorillonite), charge-deficient sites on a clay surface that facilitate the elongation of nucleotide chains are farther apart from each other and result in the formation of longer oligomer chains. As a result, they serve as better catalysts that produce longer oligomers with higher yields. The distribution of charge-deficient sites within the layers, on the other hand, does not seem to play a crucial role for the extent of catalytic activity, Table 1.
A comparison of the extent of catalytic role of montmorillonites collected from different sources showed that a montmorillonite from Wyoming, commercially known as Volclay, produces oligomers with the highest yields and longest oligomer chains, while montmorillonite from Otay, California (a montmorillonite with high charge density) catalyzes the formation of oligomer chains with only 4 monomer units, Table 1. Lower charge deficiency leads to longer separation distances between the charge-deficient sites on a clay surface, which permits the elongation of nucleotide chains to a greater length. In montmorillonites with higher charge deficiency, on the other hand, charge-deficient sites are closely positioned on the surface, preventing the elongation of oligonucleotide chains.
The role of the nature of an interlayer cation of montmorillonite, base moiety of the mononucleotide, and purity of the activated monomer on the yield and length of the longest oligomers formed by montmorillonite catalysis was discussed in detail in the work of Ertem (2004, pp 560–563).
In the laboratory, charge density (ξ or χ, kappa) is determined by saturating the montmorillonites with n-alkyl-ammonium cations that contain increasing lengths of alkyl chains, [CH3-(CH2) n -NH3]+, where n = 13–16 and 18, then measuring the d(001) interlayer spacing of the resulting montmorillonite-alkyl ammonium cation-montmorillonite complexes by XRD. Data presented in Table 1 demonstrate that length and yield of the longest oligomer is a function of the mean layer charge density (χ) of montmorillonites. Layer charge density of montmorillonites is calculated by measuring the d(001), namely, interlayer spacing of the montmorillonite-alkyl ammonium-montmorillonite complexes by XRD (Lagaly, 1994; Ertem et al., 2010).
The Mars Science Laboratory rover has an instrument on board, CheMin, that is capable of performing remote robotic analysis of the martian surface rocks. It can provide information on the mineralogical and elemental analysis of the samples by combined application of XRD and X-ray fluorescence (XRF), which makes it possible to calculate the charge density and, hence, the extent of catalytic activity of martian montmorillonites for oligomer formation.
3.4. Role of preparation method of homoionic montmorillonites on the length of the oligomer chains' form
Montmorillonites converted into homoionic form by the titration method (Banin et al., 1985) produce longer oligomers compared to the saturation method (Brindley and Ertem, 1971). In the course of preparation of homoionic montmorillonites by the saturation method, montmorillonite is treated with 1 N NaCl solution to exchange the interlayer cations with Na+ ions. In the process, the OH- groups located at the edges of clay particles are also exchanged with Cl- ions. Since the reaction of 5′-AMP that leads to the formation of oligomers is run at pH 8.0, 5′-AMP monomers present in the solution become negatively charged and are repelled by the negatively charged clay particles resulting in low dimer yields. On the other hand, in the preparation of homoionic montmorillonites by the titration method, montmorillonite suspension is titrated with the cation solution at a controlled pH. As a result, OH- groups of the clay particles that occupy the edge sites remain intact, that is, without being exchanged by the highly electronegative Cl- ions.
35Cl solid state nuclear magnetic resonance (NMR) and XRD measurements of montmorillonites prepared by saturation and titration methods have shown that they have identical characteristics, Fig. 5.
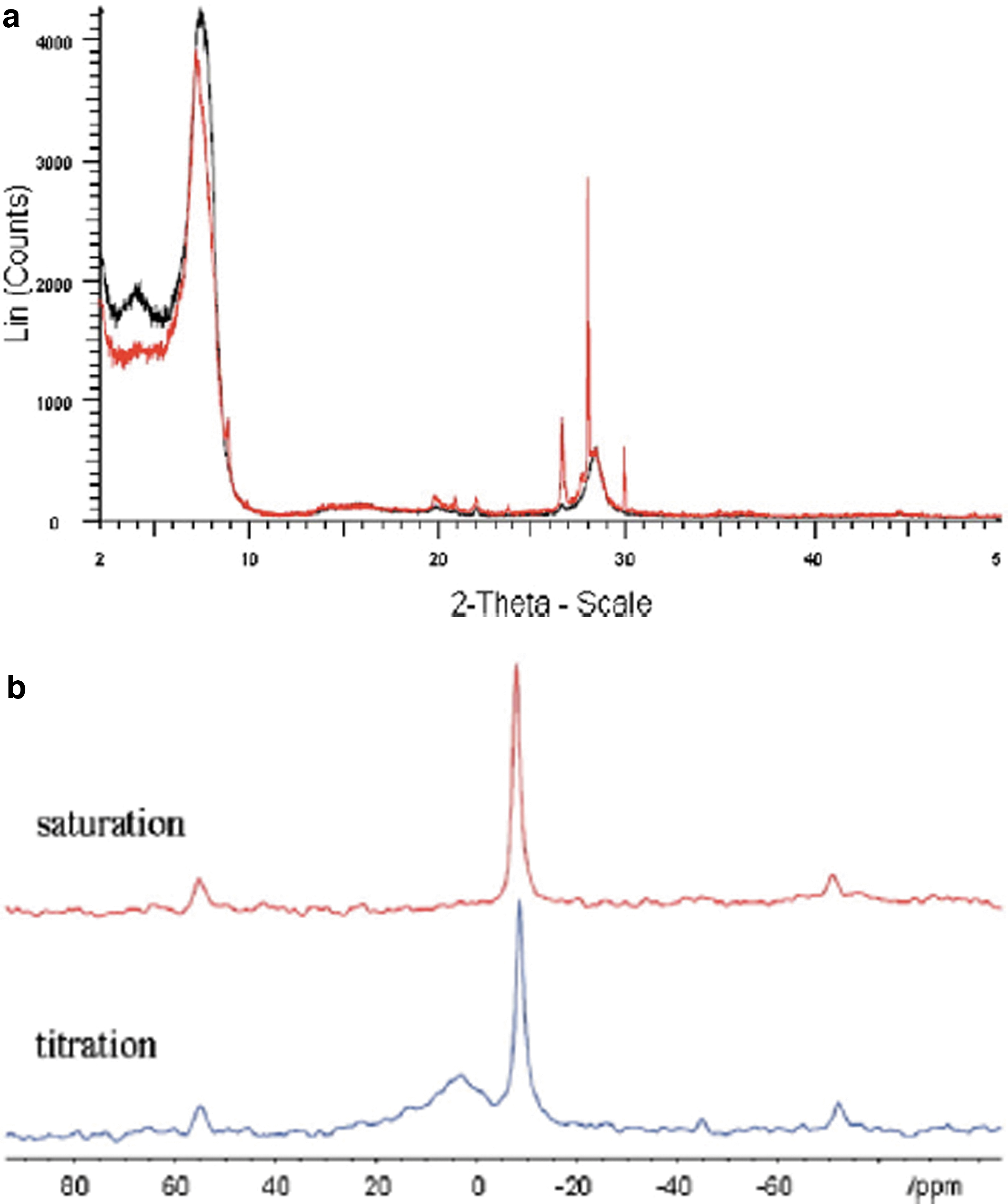
(
3.5. Formation of hetero-RNA oligomers by montmorillonite catalysis
Montmorillonite-catalyzed reaction of a mixture of ImpA and ImpC at a mixing ratio of 1:1 produces hetero-oligomers that contain 56% A-monomers and 44% C-monomers incorporated in their chain structure. In the hetero-oligomers thus formed, 56% of the phosphodiester bonds are RNA-like 3′,5′-linkages (Ertem et al., 2008). Sequence analysis of trimer isomers formed by a montmorillonite-catalyzed reaction of ImpA and ImpU mixture demonstrates that reaction produced oligomers that contain 61% RNA-like 3′,5′-linkages. Furthermore, 88% of the linear trimers were hetero-isomers; that is, they contained both A and U monomers incorporated in their chain structure (Ertem et al., 2007).
Regioselectivity and the length of the longest oligomers formed in the oligomerization of mixtures of activated monomers vary with the structure of the base unit attached to the ribose ring. Purine nucleotides produce mainly RNA-like 3′,5′-linked oligomers (Ferris and Ertem, 1992a, 1992b), while pyrimidine nucleotides form oligomers containing mainly 2′,5′-linkages (Ertem and Ferris, 1996).
We have extended our research to montmorillonite-catalyzed reaction of a mixture of four monomers that are building blocks of RNA. This reaction produced only a limited number of dimer isomers, which suggests that only a limited population of RNA would have formed in the mineral-catalyzed prebiotic reactions (Ertem and Ferris, 2000). Figure 6 shows the HPLC analysis of dimer isomers formed in montmorillonite-catalyzed reaction of ImpA, ImpC, ImpG, and ImpU mixture.

C-18-Reverse HPLC elution profile of NpN isomers formed in the montmorillonite-catalyzed reaction of ImpA-ImpC-ImpG-ImpU mixture within 2h/25°C. (1) Unknown. (2) U2′pU. (3) C2′pU. (4) U2′pC and G2′pU. (5) C2′pC. (6) A2′pU, U3′pU, and G2′pG. (7) C3′pU. (8) G2′pC. (9) G2′pA. (10) A2′pG. (11) C3′pC and A2′pC. (12) C2′pA and A3′pG. (13) A2′pA. (14) A3′pU and G3′pC. (15) C2′pG. (16) A3′pC. (17) G3′pA and G3′pG. (18) A3′pA. (Data from Ertem and Ferris, 2000.)
Kinetic studies of montmorillonite-catalyzed oligomerization of phosphorimidazolides of purine and pyrimidine nucleosides, namely, ImpA, ImpC, ImpG, and ImpU, demonstrated that reaction rates for oligomerization are the same, which suggests that RNA formed on early Earth could have contained a mixture of four of the heterocyclic bases (Kawamura and Ferris, 1999).
These studies clearly demonstrate the crucial role of mineral catalysis in the events that led to the origin of life. Montmorillonite-catalyzed formation of a phosphodiester bond between two RNA monomers, which requires the removal of a water molecule from two monomers, takes place in aqueous solution! This exactly resembles the analogy that, after doing the laundry, instead of putting the washed clothes into a drier, one throws them into the ocean and picks them up as dry due to the role of mineral catalysis!
The role of minerals in the events that led to the origin of life has also been discussed in two recent works. Kawamura and Maurel (2017) studied the chemical evolution of RNA from inorganic components through mineral-mediated RNA formation compatible with the early Earth environment. Fornaro et al. (2018) reviewed the catalytic and protective properties of martian minerals and their implications for the possible origin of life on Mars. Pedreira-Segade et al. (2018) investigated the interaction of building blocks of RNA and DNA with phyllosilicates under various physicochemical conditions such as temperature, pH, and salinity at ambient conditions and concluded that adsorption takes place via ligand exchange, where phosphate groups of the nucleotides interact with the hydroxyl groups of the phyllosilicates.
4. The RNA World Hypothesis and Template-Directed Synthesis of Oligoguanylates on Synthetic Oligo(C) Templates Formed by Montmorillonite Catalysis
In contemporary living systems, genetic information is transferred from one generation to the next by an enzyme called “RNA replicase,” which is an enzyme that catalyzes the replication of RNA from an RNA template. This is in contrast to a typical DNA-dependent RNA polymerase, which catalyzes the transcription of the RNA chain from a DNA template.
The RNA world hypothesis, as put forth in chemical evolution/origin-of-life studies, proposes that RNA can both store genetic information and act as template for the formation of complementary RNA chains (Joyce, 1989). Template-directed synthesis of complementary oligonucleotide chains on 3′,5′-linked oligonucleotides was first demonstrated in Leslie E. Orgel's laboratory, where commercially available oligonucleotides, namely, polycytidylic acid, poly(C), were used as a template. Oligomerization products were 6–10 monomer units long (Inoue and Orgel, 1983).
4.1. Template properties of oligocytidylates
We investigated the template properties of oligocytidylates formed by mineral-catalyzed reaction of ImpC for the formation of oligoguanylates from 2-MeImpG, Fig. 7. In two sets of experiments, oligocytidylates used as a template contained (a) either both 2′,5′- and 3′,5′-linkages or (b) exclusively 2′,5′-linkages (Ertem and Ferris, 1996).

4.1.1. Experimental
Montmorillonite-catalyzed oligomerization of ImpC produces oligocytidylates containing both 2′,5′- and 3′,5′-linked oligomers 20 monomer units long (Ertem and Ferris, 1996). Oligocytidylates containing exclusively 2′,5′-linkages have been isolated from the oligocytidylates formed from ImpC by mineral catalysis following the procedure described previously (Ertem and Ferris, 1996). This mixture has been treated with Ribonuclease T2, an enzyme that exclusively hydrolyzes the 3′,5′-linkages but is inactive toward the 2′,5′-linkages (unless a considerable excess of it is used!).
We have shown that oligocytidylates that contain a mixture of 2′,5′- and 3′,5′-linkages, and exclusively 2′,5′-linkages, can serve as a template for the formation of complementary oligoguanylates. More importantly, oligoguanylates formed on exclusively 2′,5′-linked oligocytidylates that contained both 2′,5′- and RNA-like 3′-5′-linkages. This is the first report in the literature that demonstrates the template capability of a synthetic oligomer chain formed by mineral catalysis in the laboratory and containing exclusively 2′,5′-linkages that serves as a template for the formation of complementary oligomer chains containing both 2′,5′- and 3′,5′-linked oligomers (Ertem and Ferris, 1996, 1997).
5. Significance of Montmorillonite-Catalyzed Formation of RNA Oligomers in the Events Leading to the Origin of Life
Mineral-catalyzed formation of RNA-like oligomers would have no real significance if these oligonucleotides did not serve as a template, that is, they did not catalyze the formation of complementary oligomer chains as shown in Fig. 7. Leslie Orgel is the pioneer in the study of a template-directed synthesis of complementary RNA oligomers. In his experiments, he used the commercially available polycytidylic acid, Poly(C), as a template for the formation of oligoguanylates from activated G-monomers, namely, guanosine 5′-phospho-2-methyl-imidazolide, 2-MeImpG (Sulston et al., 1968a, 1968b, 1969; Ninio and Orgel, 1978; Lohrmann and Orgel, 1980; Inoue and Orgel, 1981; Inoue et al. 1984; Joyce, 1984; Joyce et al., 1984a, 1984b; Schwartz and Orgel, 1984; Grzeskowiak and Orgel, 1986; Acevedo and Orgel, 1987; Orgel, 1992; Wu and Orgel, 1992a, 1992b; Hill et al., 1993; Koppitz et al., 1998).
Orgel also demonstrated that an oligo(C) chain only 4 monomer units long was capable of serving as a template for the formation of complementary oligo(G)s from 2-Me-ImpG (Grzeskowiak and Orgel, 1986).
The research reviewed above clearly demonstrates the role of mineral catalysis for the formation of RNA-like oligomers in the events that led to chemical evolution, and eventually Origin of Life as was first suggested by John D. Bernal in a lecture delivered in 1947 and published in 1949!
6. Protective Role of Minerals for Bioorganic Compounds Against the Effects of Radiation
We also investigated the protective role of Mars analog minerals for bioorganic compounds against the effects of UV (ultraviolet) and cosmic radiation (Ertem et al., 2020).
6.1. Protective role of Mars analog minerals for bioorganic molecules against the effects of UV radiation under simulated martian conditions
Phyllosilicates along with gypsum and hematite are thought to have formed on the martian surface 4.6 to 4 billion years ago, when Mars was believed to have been warmer and wetter (Bibring et al., 2006). Data obtained by OMEGA (Visible and Infrared Mineralogical Mapping Spectrometer on the Mars Express Orbiter) reveal the presence of iron-bearing silicates along with hydrated phyllosilicates and sulfates in the southern hemisphere (Bibring et al., 2005; Bishop et al., 2013).
6.2. Mars simulation experiments
We studied the protective role of Mars analog minerals for the bioorganic molecules purine, pyrimidine, and uracil, which are crucial components of biochemical processes in extant living systems, with regard to the effects of UV radiation. Irradiation was performed in a Mars Simulation Chamber under CO2 atmosphere and low temperature (Fig. 1a and 1b in Ertem et al., 2017).
6.2.1. Preparation of mineral-organic mixtures for UV irradiation
Organic compounds purine, pyrimidine, and uracil, Fig. 8, used in UV irradiation experiments are crucial biomolecules in scenarios of prebiotic molecular organization.
Structures of purine, uracil, and pyrimidine.
Aqueous solutions of purine, uracil, pyrimidine, and sodium perchlorate were prepared to contain the organic compound at a known concentration. Calculated volumes of these solutions were added to each 400 mg of a Mars analog mineral to give a final organic concentration of 25 ppm, that is, 25 mg organic/kg mineral, and 0.6% sodium perchlorate. Mixtures were freeze-dried at -85°C and 20–25 mbar for 24 h and UV irradiated in a Mars Simulation Chamber. Following irradiation, organics were extracted from minerals by water. For each mineral-organic mixture, extraction was repeated two more times. Combined extracts were freeze-dried, dissolved in 2.0 mL of water, and analyzed by UV spectrophotometry (Ertem et al., 2017).
Ertem et al. (2017) provided a photograph of the chamber, the parameters used in the chamber, and conditions present on the martian surface. The dose of UV radiation received by mineral-organic mixtures was 0.028 W m−2 nm−1, which corresponds to 5 martian sol.
Analysis of UV-irradiated organic compounds in the presence and absence of Mars analog minerals demonstrated that minerals do protect the organics from the destructive effects of UV radiation. In the absence of minerals, organic compounds completely decomposed as the result of UV radiation (Ertem et al., 2017).
The penetration depth of UV radiation into soil is about 1 mm. We also investigated the protective role of Mars analog minerals for biomolecules with regard to the effects of more energetic gamma radiation with a dose that corresponds to 500,000 years both on the martian surface and 5 cm below the surface (Ertem et al., 2019, 2020). While the energy of UV radiation is in the range of a few electron volts to about 100 eV, energy of gamma-ray photons is greater than 100 keV. Results of gamma irradiation research that demonstrates the protective role of Mars analog minerals on the martian surface and 5 cm below the surface are presented in separate publications (Ertem et al., 2019, 2020).
The possible role of UV light in the events that led to the origin of life on Earth, and on exoplanets, was discussed in the work of Szostak (2019).
7. Conclusions
Our investigations, as indicated above, into the catalytic role of minerals in the events that led to the origin of life, namely, for the formation of RNA-like oligomer chains, the capability of these oligomer chains to serve as a template for the formation of complementary oligonucleotide chains, and the protective role of minerals for biomolecules against the harmful effects of radiation demonstrate that minerals might have played a significant role in the events that led to the origin of life—as was first suggested by John D. Bernal as early as 1947—and in the protection of organic compounds from the effects of radiation on Mars and other planets.
Montmorillonite catalysis facilitated the formation of RNA-like oligomers in aqueous solution, where hydrolysis is a competing reaction. Furthermore, and equally important, montmorillonite catalysis limited the number of isomers that may have formed in the oligomerization reaction of a mixture of four activated monomers, namely, ImpA, ImpC, ImpG, and ImpU as shown in Fig. 6 above.
Also indicated is the protective role of minerals for building blocks of RNA against the harmful effects of radiation.
Footnotes
Acknowledgments
This review paper has been prepared as a response to the vast interest shown by my colleagues to the session I had the pleasure of organizing with the title “Role of Minerals in the Events Leading to the Origin of Life” during the 55th Annual Meeting of the Clay Minerals Society held at the University of Illinois at Urbana-Champaign on June 11–14, 2018.
Most of the montmorillonite catalysis research has been supported by several funding awards to the late Professor James P. Ferris during my stay in his laboratory 1986–1998! I am very grateful to Professor Ferris, the hardest-working scientist I have ever known, for the opportunity to work on this incredibly interesting research.
Ultraviolet irradiation research was supported by a grant from NASA NNX10AT27G-EXOB 2009. Mars Simulation Chamber was constructed by funds awarded by NAI DDF. Gamma irradiation research was supported by NASA Astrobiology Institute NNA15BB01A. I am very grateful to both NASA and NAI for their generous support.
Abbreviations Used
Associate Editor: Nita Sahai