
Abstract
The search for life beyond Earth has focused on Mars and the icy moons Europa and Enceladus, all of which are considered a safe haven for life due to evidence of current or past water. The surface of Venus, on the other hand, has extreme conditions that make it a nonhabitable environment to life as we know it. This is in contrast, however, to its cloud layer, which, while still an extreme environment, may prove to be a safe haven for some extreme forms of life similar to extremophiles on Earth. We consider the venusian clouds a habitable environment based on the presence of (1) a solvent for biochemical reactions, (2) appropriate physicochemical conditions, (3) available energy, and (4) biologically relevant elements. The diversity of extreme microbial ecosystems on Earth has allowed us to identify terrestrial chemolithoautotrophic microorganisms that may be analogs to putative venusian organisms. Here, we hypothesize and describe biological processes that may be performed by such organisms in the venusian clouds. To detect putative venusian organisms, we describe potential biosignature detection methods, which include metal-microbial interactions and optical methods. Finally, we describe currently available technology that can potentially be used for modeling and simulation experiments.
1. Introduction
Among the Solar System bodies most often studied under the focus of astrobiology, Venus is not a preferred one to search for life as we know it. The surface is extremely hot (471°C), and the atmospheric pressure averages 90 atm (Avduevsky et al., 1971), which renders the surface uninhabitable (National Research Council, 2007). There are hypotheses that life could exist on the surface in another organic form, given that polymeric organic molecules are stable in temperatures as high as 460°C (Ksanfomality et al., 2019). Additionally, some studies have considered the potential for surface biochemistries that use supercritical CO2 as an alternate solvent for life (Budisa and Schulze-Makuch, 2014). However, such a hypothetical chemistry of life has not been thoroughly investigated and is purely speculative.
A more relevant ecosystem for study, in relation to search for life at Venus, is its cloud layer. Some conditions of the clouds (such as temperature and pH) are harsh but within the range that some terrestrial microorganisms could reproduce and survive. The search for environments that could support life in the Solar System is guided by the “follow the water” approach—searching in locations with evidence of current or past liquid water—because life as we know it requires it. The water activity of Venus, which is hypothesized to be suitable for the survival and proliferation of life, is further discussed below. While present Venus is a parched world (Donahue and Hodges, 1992), hypothetical life may have found safe haven in the clouds, where some water vapor remains.
The hypothesis of Venus' habitability was first introduced in the 1960s by biophysicist Harold Morowitz and astronomer Carl Sagan (Morowitz and Sagan, 1967) and was resurrected in the late 1990s by David Grinspoon and Charles Cockell (Grinspoon, 1997; Cockell, 1999). It was also recently reintroduced in the work of Limaye et al. (2018a) with the possibility of life-forms being the unknown absorbers, which has fueled a new, modern interest in the possibility of life on Venus; in the past, others have speculated that the unknown absorbers may be organisms (Hapke and Nelson, 1975; Shimizu, 1977; Boyer, 1986), though they have done so without a great deal of elaboration. This hypothesis is motivated by Earth- and satellite-based observations of differences in albedo across various wavelengths, including in the UV range, where it appears significant solar absorption is taking place (Limaye et al., 2018a). Though many chemical species have been proposed as candidates since the first absorbance measurements, these absorbers have yet to be definitively identified (Pollack et al., 1979; Krasnopolsky, 1986; Pérez-Hoyos et al., 2018). A detailed list of proposed chemical species can be found in the work of Limaye et al. (2021a). Despite having been proposed several times, a biological explanation for this absorbance has yet to be explored in detail.
In the present study, we use the habitability concept explored in the work of Cockell et al. (2016) to highlight the main characteristics of the venusian cloud ecosystem that make it hypothetically habitable. This concept states that “a series of physicochemical requirements must come together at the spatial scale of an organism to allow organism to be active in a given environment” (Cockell et al., 2016). These physicochemical requirements, discussed below, include the following: (1) a solvent for biochemical reactions, (2) appropriate temperatures and physicochemical conditions, (3) available energy, and (4) biologically relevant elements (CHNOPS and others).
2. Venus under a Habitable Lens
2.1. Solvent: Water
All known forms of life require liquid water as a universal solvent for biochemical reactions, and therefore this guides the search for life elsewhere (Mottl et al., 2007). Water does not currently exist on the surface of Venus, as the high temperatures make it impossible to exist in a liquid state; however, this may not have been the case during Venus' early years. The present deuterium-to-hydrogen ratio (D/H) of Venus suggests that it may have had a shallow liquid water ocean during the first 2 billion years of its existence (Way et al., 2016), which could have contributed to an origin of life. All the inner solar system planets are likely to have had the same initial D/H; so when increased levels of deuterium were detected on Venus, it was inferred that a significant amount of water was lost, and after dissociation the heavier deuterium was lost at a slower rate. However, further surface rock analysis must be conducted to confirm the presence of water; for further discussion see the works of Kane et al. (2019) and Way and Del Genio (2020). There are multiple hypotheses as to why Venus lost its water, including the runaway greenhouse effect or atmospheric escape (Salvador et al., 2017). While most of the water on Venus is gone, some is still present as water vapor in the atmosphere, with current estimates of global mixing ratios of 40–200 ppm (Donahue and Hodges, 1992). While these levels are severely low, sulfuric acid droplets are also abundant in the atmosphere (Pollack et al., 1974). A characteristic property of sulfuric acid is its hygroscopic nature, which means these droplets in the clouds likely contain some liquid water that would potentially be accessible to putative life (Limaye et al., 2018a). The currently known limit of water activity (αw) for terrestrial microorganisms is 0.605 (Stevenson et al., 2015a, 2015b). Additionally, there is evidence of some activity as low as 0.585 (Stevenson et al., 2017). The concentration of H2SO4 corresponding to such water activity is 6.3 M. This concentration, however, is much lower than the estimated concentration of H2SO4 at Venus, based on radio occultation measurements across the middle and lower clouds, of 12.6–17.8 M (75–95% w/w) at 25°C to 12.5–17.4 M (75–95% w/w) at 50°C (Kliore et al., 1979; Jenkins et al., 1994; Gubenko et al., 2001; Imamura et al., 2017). According to the method used in the work of Grinspoon and Bullock (2007), with corrections for high activities (Nordstrom et al., 2000), the above concentration of sulfuric acid corresponds to a pH of -1.45 and, hence, in the pH range predicted for all layers of the venusian clouds (Grinspoon and Bullock, 2007). This value can be slightly corrected by taking into account any other chemical components dissolved in aerosols. However, the water activity (and by proxy pH) estimated with this method informs us that the pH of the clouds is most likely not a limiting factor for extremophilic organisms of the terrestrial type.
Nevertheless, another approach to the estimation of environmental pH in venusian clouds gives values of the actual pH as low as -11.5, as cited in the work of Seager et al. (2020). This makes such venusian clouds incomparable to any terrestrial ecosystems in relation to acidity (Seager et al., 2020). In this case, the possibility for living organisms can be correlated with any mechanism of water concentration in small microenvironments, in which organisms could still develop.
Another limiting factor related to the concentrations of various compounds in water solutions is chaotropicity. Chaotropic solutes weaken electrostatic interactions and destabilize biological macromolecules (Hallsworth et al., 2007; Yakimov et al., 2015). Sulfates, on the other hand, act as kosmotropes and strengthen electrostatic interactions and stabilize macromolecules (Yakimov et al., 2015).
2.2. Appropriate physicochemical conditions
Given that we hypothesize that some water is available to putative life, we must also consider the physicochemical conditions of the venusian clouds; that is, are they appropriate for life? Based on terrestrial environments, the conditions in the clouds are extreme; however, terrestrial extremophiles have been known to survive and even reproduce in some of these conditions. The temperature in the lower cloud layer (47–50.5 km) ranges from approximately 100°C to 60°C (Moroz, 1981; Limaye et al., 2017, 2018b), which is well within the currently understood temperature limits of life (National Research Council, 2007). Several microorganisms survive and reproduce in temperatures as high as 121–122°C, including the archaea Geogemma barossii (known as “Strain 121”) (Kashefi and Lovley, 2003) and Methanopyrus kandleri strain 116 (Takai et al., 2008). Unlike the high atmospheric pressure at the surface, the pressure in the lower clouds is not a limiting factor for habitability—varying from approximately 1.6 to 1 atm and decreasing with increasing altitude (Moroz, 1981). Additionally, it has been shown that microorganisms can even survive in the high vacuum of space (Imshenetsky and Lysenko, 1965; Horneck et al., 2010; Cheptsov et al., 2018; Yamagishi et al., 2018a).
While the conditions of the clouds are more favorable for life compared to the surface, any hypothetical life still faces challenges, including the environment's acidity. The clouds are composed of large percentages of sulfuric acid, which makes the environment highly acidic, with a pH range of -1.5 to 0.5 (47–65 km) (Grinspoon and Bullock, 2007). There are no organisms that are able to survive at such a low pH of -1.5, however some acidophiles are able to survive in a pH of -0.06, such as the genus of aerobic archaea Picrophilus (Schleper et al., 1996); this is the currently known lower pH limit of life (National Research Council, 2007). We have provided some examples of terrestrial organisms that can potentially survive these respective hazards in Table 2.
It is important to note, however, that the putative venusian organisms would need to survive these conditions simultaneously, as do terrestrial polyextremophiles. We discuss this in Sections 4 and 6 below.
Nevertheless, potentially habitable zones in the form of aerosol microniches may extend throughout the cloud layer, up to approximately 70 km, and may be suitable for microorganisms. In the upper layer, temperatures drop to about -45°C (Moroz, 1981), a temperature where microbial cells are known to remain viable in a dormant state (Clarke et al., 2013). At 62 km, the temperature is approximately -25°C (Moroz, 1981), the lowest temperature limit for prokaryotic cells to metabolize as proposed for the icy Antarctic environment (Clarke et al., 2013). Thus, the widest suitable temperature range can be -25°C to 100°C, the highest temperature measured in the lowest level of the clouds (Limaye et al., 2018a), assuming the viability of both thermophilic and psychrophilic microorganisms. In addition to being an area of relatively habitable physicochemical conditions, the altitude range ∼47–70 km is where the aforementioned unknown absorbers have been observed via in situ measurements at ∼47–64 km (Berteaux et al., 1996) and via spacecraft images, which gives credence to the Venus astrobiology hypothesis. Taking this evidence into account, the altitude range of 47–70 km seems to be the most suitable for putative venusian microorganisms that would need to survive multiple stressors cumulatively.
2.3. Energy
The energy transformation process proposed to be available to hypothetical life is a combination of chemosynthesis and photosynthesis where an iron- and sulfur-centered metabolism is coupled with the reduction of CO2 by photosynthesis (Limaye et al., 2018b; Mogul et al., 2021b). The UV flux in the high venusian atmosphere (above the clouds) is extremely high, approximately 190% that of present Earth (Cockell, 1999). However, this flux decreases while decreasing in altitude as it is absorbed by the cloud layer. At the proposed habitable altitudes, the UV flux is depleted to levels lower than at the surface of Archean Earth, when life arose (Cockell, 1999). In addition to UV, the upper atmosphere of Venus also receives a higher amount of cosmic ionizing radiation compared to Earth due to its lack of a magnetic field and closer orbit to the Sun (Nordheim et al., 2015). Increased intensity of cosmic rays can cause damage to microbial cells, such as DNA damage. However, modeling of radiation, both background and from extreme solar events in the context of the venusian atmosphere, has shown that doses at altitudes within the proposed venusian habitable zone would likely not be lethal to life (Dartnell et al., 2015; Herbst et al., 2020). Outside of ionizing radiation, non-ionizing (solar) radiation is an additional hazard for photosynthetic life. In terrestrial organisms, continuous exposure to photosynthetically active radiation has been shown to damage or inactivate photosynthesis-related structures, such as in cyanobacteria via the production of reactive oxygen species (Latifi et al., 2009; Pathak et al., 2019; Soriano et al., 2019). Nevertheless, photosynthesis serves as the dominant source for carbon input on Earth despite the potential radiation hazards. Similarly, non-ionizing radiation may not be a major constraint for photosynthetic microbial metabolism in Venus' clouds. Measurements obtained by the Pioneer Venus Large Solar Flux Radiometer show a solar irradiance of ∼531 W/m2 between 400 and 1000 nm at 50 km and a solar elevation of 45° (Tomasko et al., 1980; Moroz et al., 1985). While solar irradiance is dependent on the solar zenith angle, the values are holistically comparable to the irradiance that reaches the surface of Earth of ∼619 W/m2 (as calculated from ASTM G173-0 reference data across 400–1000 nm). When considering the potential for photosynthesis, comparisons of the venusian spectra to the absorption spectra of photosynthesis-related molecules (e.g., chlorophylls and carotenoids) show considerable overlap (Limaye et al., 2018a). Accordingly, calculations for photosynthetic photon flux density (PPFD) at an altitude of 49.5 km in Venus' clouds provide a value of ∼3600 μmol m−2 s−1 at the 350–950 nm range, which is similar to the value on Earth's surface of ∼3300 μmol m−2 s−1; the range of wavelengths chosen for these calculations include those associated with terrestrial oxygenic and anoxygenic photosynthesis. PPFD was calculated from irradiance spectra, where irradiances (W m−2 μm−1) for Earth (ASTM G173-0) and Venus (calculated spectra; Mogul et al., 2021b) were converted to photon flux (m−2 s−1 nm−1), integrated across 350–950 nm, and expressed as photosynthetic photon flux density (μmol m−2 s−1). Extending to 65.5 km, which is near the upper limit of our defined habitable zone, PPFD is calculated to be ∼4642 μmol m−2 s−1; however, due to the likely dormant status of putative life at that altitude, we focus on the PPFD at 49.5 km.
The makeup of the clouds also allows for energy to be obtained chemically. In situ studies of the clouds have measured multiple inorganic compounds, such as sulfur compounds, iron compounds, and others (see Table 1). These inorganic compounds, with the addition of carbon from fixing carbon dioxide, may be used by chemolithoautotrophs that perform various types of metabolic reactions for their energy source. Some of the most likely terrestrial analogs, like extremely acidophilic Acidthiobacillus ferrooxidans, are chemolithoautotrophs (Valdés et al., 2008).
Life-Critical Elements and Compounds Detected in Venus' Atmosphere
We therefore define our habitability zone broadly as the altitude range of 47–70 km with the following conditions: T: 100°C to -45°C; pH: -1.3 to ∼0.5; P: 1.18 to 0.0336 atm (Moroz, 1981; Grinspoon and Bullock, 2007). We note that our habitability zone is broader than those described by others (Limaye et al., 2021b) as we expand up to 70 km, where temperatures are near the lower temperature limit of life (Clarke et al., 2013). Our defined habitable zone can be divided into two subzones (1) an active habitable zone at altitudes of 47–57 km and (2) a dormant habitable zone up to an altitude of 70 km. This dormant zone can be defined as an area where microorganisms remain temporarily in a dormant or extremely low activity state, like terrestrial organisms in icy ecosystems on Earth. This could also be a source for additional biomass for the active habitable zone below through a cycling atmosphere process. However, we focus on the active habitable zone when discussing metabolic cycles and analog terrestrial organisms.
2.4. CHNOPS + other important elements
The basic elements for all known life on Earth—carbon, hydrogen, nitrogen, oxygen, phosphorus, and sulfur (CHNOPS)—are the guiding elements for astrobiologists in the search for life. These elements can be found in DNA, RNA, amino acids, sugars, and various other biologically relevant molecules. In addition to CHNOPS, other elements can be important to organisms for specific biological functions in different ecosystems. For example, terrestrial iron-oxidizing bacteria require Fe2+ to survive and reproduce (Weber et al., 2006). In the venusian clouds, iron could potentially be in different oxidation states. Of course, it must be taken into account that a variety of possible intermediates could be present that are a part of elemental redox cycles and their trace (or transient) presence in the reaction sites. The hypothetical biologically dependent cycling of sulfur and iron for survival in the venusian cloud environment has been previously proposed (Limaye et al., 2018a). Possible microbial metabolic processes involved are discussed below.
The CHNOPS elements and other biologically relevant elements/chemical species, such as iron and other transition metals, have been detected in the atmosphere of Venus through in situ measurements from the Venera, VeGa, and Pioneer missions (Table 1). It has been proposed that topography, storms, and weathering forces could result in the cycling and movement of these elements up to the clouds (Limaye et al., 2018a); similar mechanisms are known to occur on Earth (Kirshbaum et al., 2018). The bioavailability of these elements is unspecified, and further in situ measurements are required to determine their concentration and bioavailability; for further speculations in this article, we assume they are bioavailable. The possible bioavailability of phosphorus is discussed in an accompanying article in this issue (Milojevic et al., 2021a).
Based on the known availability of carbon and energy sources, as well as the presence of various electron acceptors, we can hypothesize the metabolic mode of a putative venusian microorganism as “thermoacidophilic chemolithoautotrophy.” However, based on the various environmental conditions in the clouds, psychrophilic (for the “dormant habitable zone”) and phototrophic organisms could also be considered (Fig. 1).

Habitability zone in the venusian clouds and the hypothesized metabolic mode of a relevant microbial form.
3. Microbial Life in Earth's Clouds
Clouds are a key component of Earth's atmosphere, covering over 60% of the surface (King et al., 2013), and they may be the only potentially habitable terrestrial region that may provide clues as to how life could be organized in a cloud environment on Venus (Sattler et al., 2001). Earth's clouds are made up of microscopic droplets of liquid water, crystals of ice, or a combination of both phases; droplets have a mean diameter range of 5–50 μm (Miles et al., 2000), which is partially comparable to aerosols on Venus (Knollenberg et al., 1980). The cloud aqueous phase is a reactive and dynamic system composed of inorganic and organic chemical species, potentially available for use by organisms (Delort et al., 2017).
The major inorganic ions found in the Earth cloud water are sulfate (SO4 2-), chloride (Cl-), and nitrate (NO3 -) anions. These are present in addition to alkali and alkaline Earth metal cations (Na+, K+, Mg2+, Ca2+) and ammonium (NH4 +) (Delort et al., 2017). Organic matter also makes up a large fraction of the soluble matter in the cloud droplets and is able to interact directly or indirectly with the aqueous chemistry of radicals, oxidants, and transition metal ions like iron (Herckes et al., 2013). Carboxylic acids and aldehydes with short carbonaceous chains represent approximately 10% of the dissolved organic carbon in the cloud droplets, and their concentration can reach levels greater than 1 mg L−1 (Marinoni et al., 2004). Carboxylic acids, as well as nitrate and sulfate, contribute to the acidification of the aqueous phase, which can range from pH 2.2 to 7 depending on the air mass origin (Delort et al., 2017).
Viruses, bacteria, and fungi are present in almost all atmospheric layers of Earth, from indoor air to altitudes of 77 km (Imshenetsky et al., 1978; Delort et al., 2010). Some seem to be involved in various biochemical cycles in the clouds, as metabolic activity has been seen in cloud water samples via the detection of ATP (Amato et al., 2007) and RNA (Krumins et al., 2014). These microorganisms are exposed to a dynamic medium that changes extremely rapidly (quick temperature and pressure changes, freeze/thaw cycles), strong physical stresses such as solar radiation (especially UV) and desiccation, various chemical stresses such as strong oxidants (H2O2, O3, HO•, etc.), toxins, and an acidic pH (Amato et al., 2017). Such microorganisms can successfully resist these stress factors by specific physiological characteristics such as the production of protecting metabolites like osmo- and thermo-protectants, antioxidants or specific enzymes, and the synthesis of lipopolysaccharides or the production of exopolymeric substances that enhance the hydrophilicity of the cell surface (Vaïtilingom et al., 2013; Amato et al., 2017). These resistance mechanisms could also be used by putative venusian microorganisms and even be considered potential biomarkers. Additionally, sporulation can be an important process that would allow venusian microorganisms to survive during possible atmospheric overturning; we note that we use “sporulation” broadly as any form of dormant, metabolically inactive protective state. The life cycle of putative microorganisms can, in particular, significantly depend on a hypothetical process of formation and coagulation of aerosols, descending under gravitational force and evaporating under higher temperatures (i.e., wet/dry cycle) (Seager et al., 2020). These putative microorganisms could thus potentially survive unfavorable conditions through “sporulation” such as desiccation in the lower layer of the clouds, from which they could rise again to higher layers by convective flows to continue their life cycle. A survival cycle of sporulation and germination would require sufficient time and energy to be realized by venusian microorganisms.
On Earth, airborne microorganisms originating from the surface can act as cloud condensation nuclei and be incorporated into the clouds to be in a more hospitable environment with water and nutrients present (Delort et al., 2017). However, the microbial composition of various tropospheric environments is quite different and depends on whether microbes are associated with dry air masses, clouds, or precipitation (such as rain, hail, or snow) (Els et al., 2019). Airborne microorganisms associated with aerosols are suspended in small water droplets, with the total number of bacteria in such bioaerosols in the troposphere ranging from 1 × 103 to 1 × 105 cells mL−1, with a decrease in concentration with increasing altitude (Delort et al., 2010).
Terrestrial clouds are a transient habitat that can last anywhere from a few hours to as long as a few days, and therefore the amount of time a microorganism would spend in the atmosphere would likely follow a similar time span. Terrestrial microbes only cope with environmental stressors transiently, whereas putative venusian organisms would have to persistently endure the stresses. However, airborne terrestrial microorganisms are good analogs for putative venusian life in relation to our understanding its physicochemical organization. Due to the evaporation-condensation mechanisms in the clouds, an exchange of microbial metabolites between droplets can occur. In addition, airborne microorganisms can perform biotransformation of organic compounds in cloud water by adapting to the cloud conditions, including any existing stressors (Amato et al., 2017).
4. Terrestrial Microorganisms That Might Survive Venusian Clouds
The venusian clouds (including our defined “habitable zone”) can only conditionally be compared to the clouds of Earth. They consist of aerosols and can also be classified as an extreme environment, like Earth's clouds, but they exhibit quite different physicochemical conditions (including pH and temperature). The venusian clouds are characterized by conditions that are close to the limits of life for terrestrial microorganisms. By some estimations (Seager et al., 2020), however, venusian cloud conditions significantly exceed known limits for terrestrial life, for example extreme acidity. Redox cycling reactions that involve ferrous and sulfur compounds are proposed to be predominant in the venusian cloud layer chemistry (Limaye et al., 2018a) and can be an energy source for chemolithotrophic microorganisms.
Table 2 shows a sample of known terrestrial extremophilic microorganisms. These microorganisms have been selected based on their ability to survive at low pH and grow at high temperatures, characteristic of the active habitable zone and, drive chemosynthetic processes with iron and sulfur as sources of energy and CO2 as the sole source of carbon. Aside from Stygiolobus azoricus, all the microorganisms listed perform both aerobic and anaerobic metabolism. Furthermore, some of them can drive reductive and oxidative reactions with Fe- and S-ions, depending on the electron acceptor available. In the venusian clouds, nitrates (rather than oxygen) can potentially be involved in redox cycling as the strongest electron acceptor. Discussion of the availability of nitrates and some possible metabolic reactions in the venusian cloud layer of interest is presented in the following section.
Extremophilic Chemolithoautotrophic Microorganisms Hypothesized to Withstand the Conditions in Venus' Clouds
Kelly and Wood (2000). 2Ohmura et al. (2002). 3Valdés et al. (2008). 4Giaveno et al. (2009). 5Segerer et al. (1991). 6Segerer et al. (1986). 7Two more species of gen. Acidianus perform the same metabolic reactions and have similar temperature optima for growth: A. ambivalens (Fuchs et al., 1996) and Acidianus tengchongensis (Zhong et al., 2004). 8Yoshida et al. (2006). 9Plumb et al. (2007).
The microorganisms presented in Table 2 have a pH limit for growth within the range 1.0–1.5 (with the exception of Acidianus sulfidivorans at 0.35), which is slightly higher than the estimated upper pH value of 0.5 for venusian clouds at 65 km (Grinspoon and Bullock, 2007). However, all other physiological characteristics of these organisms fall within the predicted physiological parameters of the active habitable zone. Therefore, the organisms presented in Table 2 are some of the closest known terrestrial candidates for survival in the venusian cloud environment. We note, however, that the physicochemical characteristics of an environment interact and may result in a different survivability. Future bacterial discoveries will likely result in the isolation of new terrestrial extremophiles that may serve as improved analog venusian organisms, as an estimated 99% of terrestrial microbial species have yet to be discovered (Locey and Lennon, 2016). For example, taking into account that the aerobic organisms Picrophilus oschimae and P. torridus can grow at a pH below 0 (Schleper et al., 1996), it is possible that new microorganisms will be found in an extremely acidic terrestrial environment that has a lower limiting pH for growth and even better corresponds to the proposed physicochemical conditions of our venusian habitable zone. Such microorganisms may even reproduce under these kinds of conditions. However, it should be noted that there are terrestrial locations with physicochemical characteristics similar to those of the venusian clouds where active life has not been found (Oren, 2013; Belilla et al., 2019; Moors et al., 2020). In addition to Venus-like temperatures and pH values, these locations also contain high concentrations of ions, such as magnesium, that increase the chaotropicity and thus decrease the limit of life (Hallsworth et al., 2007; Yakimov et al., 2015). Life-limiting factors and their cumulative effect is an active research area with many yet-unanswered questions.
5. Hypothetical Cycles of Elements in Venusian Clouds and Relevant Terrestrial Biological Processes
Chemolithotrophy is considered the most possible metabolic mode for hypothesized microbial forms in the venusian clouds. Chemolithotrophic microorganisms employ an astonishing number of metabolic pathways to extract energy from diverse inorganic electron donors and acceptors, which can have significant consequences for global biogeochemical cycles (Hooper and DiSpirito, 2013). The predicted presence of iron and sulfur at different oxidation states in the venusian clouds makes it possible for putative microorganisms to gain energy through redox reactions similar to those used by various terrestrial microorganisms, as has been previously proposed (Limaye et al., 2018a). These energy-gaining processes often occur in different ecosystems of secondary volcanic activity such as terrestrial thermal systems, for example, solfataras, fumaroles, geysers, and deep sea hot springs (Merino et al., 2019).
Taking into account the potential survivability of terrestrial extremophiles in the venusian cloud conditions, the presence of biologically relevant elements, and relevant biological processes characteristic of extreme terrestrial ecosystems, a hypothetical biological cycle based on redox reactions with ferrous iron and sulfur compounds can be proposed (Fig. 2). Sulfur and iron are considered to have played an important role for the metabolisms of the most ancient microorganisms on Earth. These compounds were likely directly involved in primitive metabolic reactions performed by the last universal common ancestor and might be crucial for life's origin (Weiss et al., 2016).

Hypothetical scheme of metabolic sulfur- and iron-centered cycles in which some terrestrial extremophiles can be involved in venusian cloud layers. DNRA = dissimilatory nitrate reduction to ammonium; NRFO = NO3 - reducing Fe(II) oxidation.
There are several known metabolic processes performed by terrestrial extremophiles that may be relevant to the hypothesized venusian microorganisms including: sulfate reduction, sulfur disproportionation, dissimilatory nitrate reduction to ammonium (DNRA), nitrate-reducing Fe(II) oxidation (NRFO), and anoxygenic photosynthesis. Species that belong to the nitrogen cycle, including nitrate, nitrite, and ammonia, have been detected in situ by the Pioneer Venus Large Probe Neutral Mass Spectrometer (Mogul et al., 2021a); however, these detections have not been confirmed by other measurements. Nevertheless, due to the abundance of nitrogen on Venus (Oyama et al., 1980) and terrestrial organisms known to metabolically convert nitrogen into nitrate (Latysheva et al., 2012), we speculate that nitrate-based metabolisms could be used by putative venusian microorganisms.
By elucidating the possible metabolism (or metabolisms) of the putative venusian microorganism (or microorganisms), we can develop an understanding of how much bioavailable energy would be needed to support life and, further, which cloud strata are likely able to support the biomass of putative venusian microorganisms. Additionally, an understanding of a possible metabolism (or metabolisms) is important for the development of future astrobiological missions by helping to determine the most favorable places for the presence of key metabolites and thermodynamic feasibility of appropriate biological processes.
5.1. Sulfate reduction
Sulfate reducers are microorganisms that belong to the Bacteria and Archaea domains that use sulfate (SO4 2-) as a terminal electron acceptor to perform anaerobic respiration with the production of hydrogen sulfide (H2S) (Muyzer and Stams, 2008; Barton and Fauque, 2009). They are considered to be among the most ancient forms of microbes and are found widely in environments rich in decaying organic material, as well as in extreme environments such as hydrothermal vents and acid-mine drainage sites, among others. In the venusian clouds, sulfate reduction could be important as a source of reduced sulfur (S2-) required in some metabolic reactions.
5.2. Sulfur disproportionation
Sulfur can exist in a number of different oxidation states that vary between S2- and S6+. The disproportionation of inorganic sulfur compounds (with intermediate oxidation states such as elemental sulfur, thiosulfate, and sulfite) is a microbially catalyzed chemolithotrophic process in which these compounds serve as both electron donors and acceptors and hence are simultaneously reduced and oxidized, generating hydrogen sulfide and sulfate.
Along with sulfate reduction, this type of metabolism is considered to be one the oldest biological processes on Earth (Canfield and Thamdrup, 1994) and occurs in different freshwater, hypersaline, and thermal ecosystems (Jorgensen, 1990; Finster, 2008; Slobodkin and Slobodkina, 2019).
The disproportionation of thiosulfate and sulfite is thermodynamically favorable under standard conditions, whereas sulfur disproportionation is endergonic and can be feasible only if the produced sulfide is removed from the reaction site. However, in the presence of ferric iron (acting as an electron acceptor), the disproportionation of elemental sulfur becomes exergonic, even under standard conditions (Slobodkin and Slobodkina, 2019).
At higher temperatures, both the energy gained from sulfur disproportionation and the solubility of elemental sulfur increase, making the conditions for this type of metabolism more favorable. Sulfur disproportionation has also been reported to occur at high temperatures (Slobodkin et al., 2011) and low pH (Boyd and Druschel, 2013; Hardisty et al., 2013). Moreover, the extremely acidophilic bacterium Acidithiobacillus ferrooxidans likely drives sulfur disproportionation in anaerobic metabolism in the presence of Fe(III) (Osorio et al., 2013).
In the process of disproportionation, microorganisms use only one compound as both an electron donor and acceptor to gain energy; this therefore minimizes the requirements of the chemical composition of their environments. This can be important for understanding the survivability and function of hypothetical organisms in the venusian clouds.
5.3. Dissimilatory nitrate reduction to ammonium
In this type of anaerobic respiration, microorganisms use nitrate (NO3 -) as an electron acceptor and produce NH3 + (Shao et al., 2010; Lam and Kuypers, 2011) that can also be important for the acidic cloud conditions as an alkaline neutralizing metabolite that produces bioavailable soluble NH4 +. In many terrestrial environments, these microorganisms can couple the biogeochemical cycles of nitrogen and sulfur to oxidize either sulfide or sulfur (Slobodkina et al., 2017). They have also been found in deep-sea hydrothermal vents. It is important to note that nitrogen species, including nitrate and ammonia, have been detected in Venus' atmosphere (Oyama et al., 1980; Mogul et al., 2021a).
5.4. Nitrate-dependent microbial Fe(II) oxidation (NDFO)
Microbes that perform this metabolism reduce nitrate to nitrite (or dinitrogen gas) and oxidize iron under anoxic conditions. This reaction is highly exergonic due to the large difference in redox potential between the couples Fe2+/Fe3+ and NO3 -/NO2 -. This type of metabolism supports microbial growth in various freshwater and marine sediments such as paddy soil, brackish lagoon, and deep-sea hydrothermal vents (Schaedler et al., 2018; Liu et al., 2019).
Nitrate-dependent microbial Fe(II) oxidation (NDFO) has been proposed to be a possible important mechanism of gaining energy for microorganisms on early or even present-day Mars in subsurface aquifers (Price et al., 2018) and can be considered as a potential metabolism for putative venusian organisms.
5.5. Anoxygenic photosynthesis
Sufficient solar energy is available in the photosynthetic range in the lower clouds, which could be available for anaerobic phototrophic organisms (Hanada, 2016; see also Limaye et al., 2021, Mogul et al., 2021b, and Section 2.3 above). At least two groups of photosynthetic microorganisms that couple their metabolism with iron and sulfur cycles could be responsible for a light-dependent type of life in the venusian clouds rich in CO2, that is, purple and green sulfur bacteria. These organisms are able to trap light energy and store it as chemical energy via the oxidation of sulfide to sulfur and use Fe2+ as electron donors and CO2 as a carbon source (Schulze-Makuch et al., 2004).
5.6. Other potential processes
While the origin of methane detected in the clouds is unknown (Donahue and Hodges, 1993), it can be hypothesized to be of biological origin. Methane can potentially be oxidized under anaerobic conditions with sulfate, nitrate, or ferric compounds (Kallistova et al., 2017) that act as electron acceptors.
Additionally, phosphine was recently detected in the venusian clouds from Earth-based telescopes (Greaves et al., 2020a) and potentially much earlier by the Pioneer Venus Large Probe Neutral Mass Spectrometer (Mogul et al., 2021a). The origin of the detected phosphine is unknown, with undiscovered chemical processes or lightning activity (Glindemann et al., 2004) being possible explanations. However, a potential origin of phosphine, including a biological one, is now being actively discussed (Bains et al., 2021). While this gas is detected in terrestrial microbial anaerobic ecosystems (see Sousa-Silva et al., 2019, for examples), the biochemical mechanisms of its formation are still unknown. It is important to note, however, that the presence of phosphine has been questioned by the community, and further studies are needed to confirm its detection (Cockell et al., 2020; Encrenaz et al., 2020; Greaves et al., 2020b, 2020c; Snellen et al., 2020; Villanueva et al., 2020; Akins et al., 2021; Mogul et al., 2021a; Thompson, 2021). Additionally, confirmation of a biological origin of phosphine is critical for its classification as a biosignature. We note that, while there have been phosphine detections near present microbial life in anaerobic terrestrial environments, animal intestinal tracts, feces, and even in laboratory bacterial cultures, this does not provide evidence of its microbial production (see Sousa-Silva et al., 2019, for known detections).
In addition to the metabolic processes involving sulfur compounds that are already known, a new process of sulfur comproportionation has been proposed as thermodynamically feasible in terrestrial sulfur-rich ecosystems (Amend et al., 2020). Here, sulfur compounds of different oxidation states, such as sulfide and sulfate, can be combined into one product, that is, sulfur, sulfite, or thiosulfate; this is the opposite of sulfur disproportionation (Amend et al., 2020). Such a reaction is exergonic under standard conditions and can be even more favorably thermodynamic in the venusian clouds at high temperature and low pH.
Finally, complex biological redox cycles can be integrated into abiogenic cycling reactions. Elemental sulfur has been demonstrated to be chemically hydrolyzed into sulfate and sulfide at temperatures ranging from 50°C to 200°C with enhancement by the presence of sulfide-binding cations such as copper (Smith, 2000). Such reactions can in principle occur abiotically in the layers close to the surface and be sources of sulfur compounds to venusian cloud microorganisms.
Microbial systems that function under conditions of limited energy sources and in extreme environmental parameters should not exhibit high biomass and are closely connected to abiogenic geochemical processes in their surroundings to most effectively gain energy. Functioning under limited energy sources, but taking advantage of abiogenic geochemical processes, was evidently the case for ancient Earth ecosystems and is at the boundary of biology and geology that could also be true for putative venusian microbial systems (Martin and Sousa, 2016; Weiss et al., 2016).
6. Multiple Protective Measures of Microbial Systems
To withstand the relatively harsh environment of the venusian clouds, putative microorganisms need to exhibit polyextremophilic properties, meaning that they must have mechanisms of resistance to multiple stress factors.
To date, a sufficient amount of data has been gathered on various natural habitats that are the most extreme in terms of the combination of physicochemical factors, and the stress resistance systems of the microbes that inhabit them (Gilichinsky et al., 1992; Abyzov, 1993; Vorobyova et al., 1997; Dib et al., 2008; Bowers et al., 2009; Harrison et al., 2013; Schuerger et al., 2013; Takai, 2019). The results of model laboratory experiments indicate an extremely high resistance in microbial communities to physical and chemical stress impacts, which may exceed the possible combinations of stressors known for terrestrial ecosystems (Schuerger et al., 2013; Cheptsov et al., 2017, 2018, 2019; Merino et al., 2019). The important conclusion from these studies is that the natural environment, especially the heterophasic environment, has a protective function in relation to the microorganisms inhabiting it (Cheptsov et al., 2018). Additionally, it helps maintain the mechanisms of microbial cell adaptation that allow for an increasing resistance to stress (Cheptsov et al., 2018). This contributed to the evolution and biodiversity of organisms in parallel to the evolution of Earth.
Life on our planet arose in the absence of a protective ozone layer, experiencing radiation of the young Sun (about 1 billion years old) that was 10–30 times higher than that of the present—in the interval of 200–350 Å (Giardino et al., 2008). Additionally, X-ray radiation is hypothesized to have been 3–4 orders of magnitude greater than its modern intensity level (Giardino et al., 2008). According to the current concept of cell viability, such conditions would inevitably lead to the suppression and death of emerging life. However, life did originate and thrived with increased biodiversity, eventually forming the present biosphere (Lever et al., 2013; Noffke et al., 2013; Grosch and McLoughlin, 2014; Allwood, 2016; Nutman et al., 2016). Similar processes could occur on other Solar System objects, in particular on Venus, taking into account the specific features of its evolution. Specific conditions in the venusian clouds and the high adaptive abilities of life as we know it can lead to the evolution of biosystems different from terrestrial ones.
Typically, stress exposure occurs as a combination of interconnected physicochemical factors, and the resistance of microorganisms in situ or ex situ to one stress factor often correlates with resistance to others: drying, salinity, temperature fluctuations, and so on (Chattopadhyay, 2002, 2008; Soina and Vorobyova, 2004; Rainey et al., 2005; Harrison et al., 2013; Beblo-Vranesevic et al., 2016; Fagliarone et al., 2017). For example, a correlative relationship between resistance to UVB and resistance to arsenic compounds has been shown in microorganisms, many of which were also resistant to antibiotics (Dib et al., 2008); the antibiotic resistance was not associated with the mutagenic effect of UVB but corresponded to the arsenic resistance. Additionally, studies conducted with samples of extreme arid soils showed that a significant percentage of the isolated strains of heterotrophic bacteria present demonstrated resistance to various physicochemical factors, including low temperatures, and a wide range of pH values and salt concentrations (Belov et al., 2018, 2019). About 30% of desiccation-resistant strains grew at a temperature of 2°C and 4°C. The growth of the bacterial community was observed in the pH range of 3–12, where 20–60% and 60–80% of isolates grew at pH 3–4 and 11–12, respectively (Fig. 3). Another example would be the resistance to ionizing radiation at doses significantly higher than the natural radioactive background, which can presumably be explained by adaptation to the extreme effects of drying, temperature, salinization, and other physicochemical factors (Jolivet et al., 2003; Shukla et al., 2007; Schuerger et al., 2013; Musilova et al., 2015). It has been shown that microorganisms have increased resistance to radiation when in their natural environment. Microorganisms, including radiosensitive ones, withstood much higher doses of ionizing radiation under low pressure and low temperature conditions when they were in their native microbial communities in the soil, compared to when isolated in pure cultures (Cheptsov et al., 2017, 2018; Kryuchkova et al., 2018). These and other data on the responses of microorganisms and natural microbial systems to the simultaneous exposure to multiple stress factors comparable to the space-like environment provide compelling reasons for an astrobiological search for Earth-like microbial life on other Solar System objects, including Venus (Harrison et al., 2013; Schuerger et al., 2013; Cheptsov et al., 2018; Merino et al., 2019). Thus, a variety of stressors result in a spontaneous or cumulative response of cells, populations, communities, or biosystems. This indicates that a common system of adaptive mechanisms is responding to the exposure to extreme factors by activating certain protective processes; this has the potential for adaptive development. The microbial mechanisms of resistance to multiple stressors are not quite clear and are being intensively studied and discussed (Shukla et al., 2007; Sghaier, 2011; Harrison et al., 2013; Mirete et al., 2016; Fagliarone et al., 2017). On Venus, novel stress response mechanisms or mechanisms similar to those here on Earth can be an important feature of living systems.
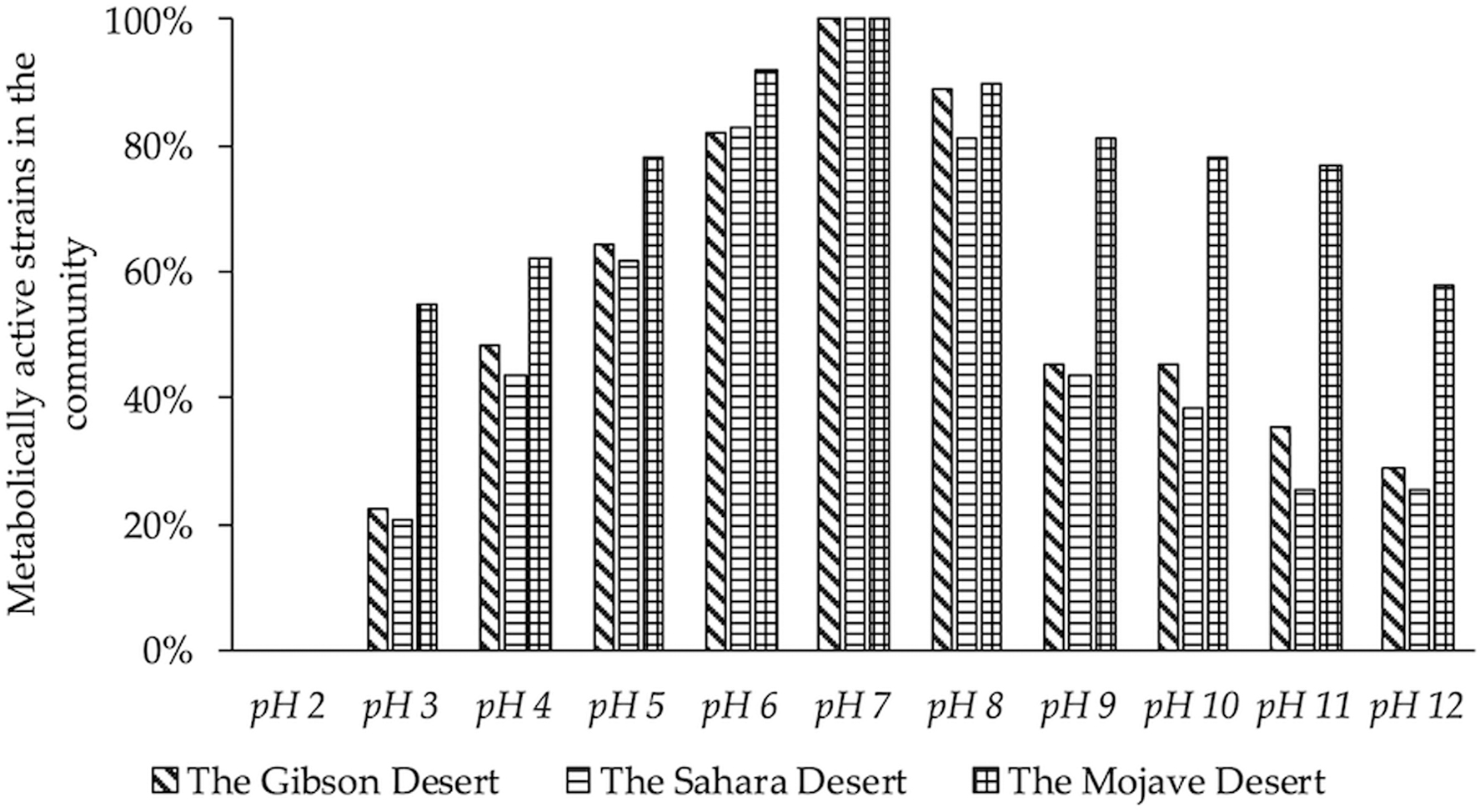
The percentage of grown bacterial strains isolated from arid desert soils at different pH.
7. Potential Biosignatures in the Venusian Clouds
No direct terrestrial cloud-based analogs exist that can represent the cloud environment of Venus. The proper methodology for detection of biological activity in such a unique environment can be developed through the combination of different techniques used for terrestrial microbial systems and modified to the specific characteristics of the venusian cloud environment. Putative venusian microorganisms likely live in an aerosolized form, exhibit chemolithotrophy, and should survive in extreme acidity and a range of temperatures. Promising methods for the detection of venusian organisms include those based on the interactions of microorganisms with particles, including (i) metals and (ii) biomarkers associated with the unique metabolic machinery of extremophiles.
7.1. Metal-microbial interactions of metallophilic extreme thermoacidophiles
The hypothetical microbial aerial biosphere in the venusian clouds contains Fe and S particles (Andreichikov et al., 1987). Metal-transforming thermoacidophiles represent an exciting field of research for the study of microbe–Fe/S interactions in order to find true biosignatures of life in the acidic venusian clouds. Deciphering the metal-microbial interfaces of metallophilic extreme thermoacidophiles is of special interest.
Metallophilic extreme thermoacidophiles of the archaeal order Sulfolobales (Metallosphaera spp., Sulfolobus metallicus [Fig. 4], Sulfolobus acidocaldarius, and Acidianus spp.) are metal-mobilizing microorganisms, which thrive in hot acidic environments and utilize energy from the redox transformations of metal inorganic sources (Kölbl et al., 2017; Milojevic et al., 2019a, 2019b, 2021b). While the use of morphological assessment and carbon-content evaluation may not be fully useful as a criterion for distinguishing biotic from abiotic chemistries, organometallic interfaces can provide evidence to support a biological origin of examined samples. In some particular cases, it is possible to assess the metal-microbial interface of extreme thermoacidophiles down to the nanometer scale and derive fine structural information of organometallic complexes (Blazevic et al., 2019; Milojevic et al., 2019a, 2019b, 2021b).
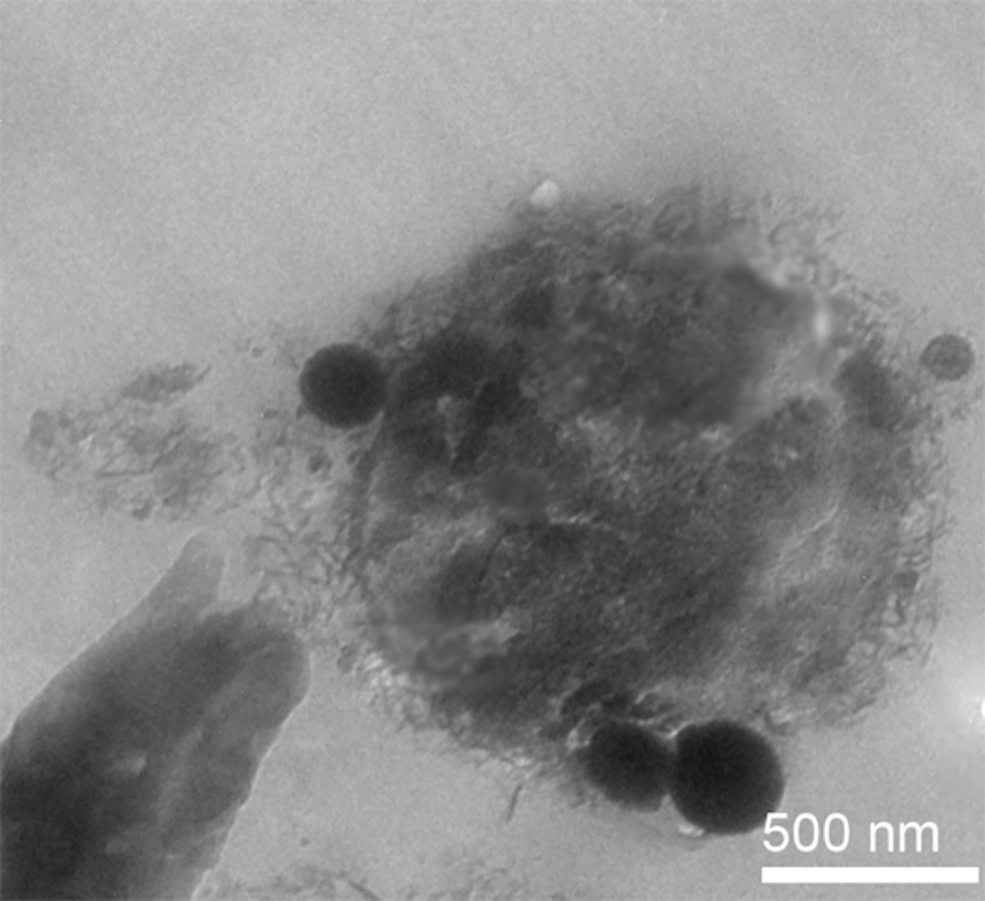
Transmission electron microscope image showing a single cell of the extreme thermoacidophile Sulfolobus metallicus with attached iron-containing nanoparticles.
The spatial distribution and fine structure of iron species on the cell surface of M. sedula grown on genuine (not synthetic) meteorite material has been investigated (Milojevic et al., 2019b). Nanoanalytical spectroscopy measurements acquired locally at the meteorite-microbial interface (point analysis with a beam diameter of 1 Å) by scanning transmission electron microscopy demonstrated that the cell surface of M. sedula bears a mixed valence iron distribution with predominant Fe2+ species (Milojevic et al., 2019b). Fe2+ detected on the cell surface may primarily serve as an energy source. The accumulation of redox heterogeneous Fe species occurs at the microbe-metal interface due to M. sedula undergoing oxidative metabolism; this is analogous to microbial-mediated sulfur oxidation with the wide variety of redox heterogeneous intermediate sulfur compounds and serves as a fingerprint of chemolithoautotrophic life. The organometallic interfaces described in the referenced study and their fine structural details may constitute relevant biosignatures to search for in environmental samples, if they are not destroyed by harsh environmental conditions. Today, advanced nanoanalytical techniques allow for the detection and analysis of single atoms incorporated into a single microbial cell, thus virtually providing no limit to biosignature detection (Blazevic et al., 2019; Milojevic et al., 2019a, 2021). With regard to the preservation potential of the organometallic interfaces, many questions remain to be answered, including the influence of complex environmental parameters (e.g., physicochemical constraints and the atmospheric chemistry of venusian clouds) on metal-bearing complexes attached to the cell wall. Such investigations may help establish chemical and morphological criteria for confirmation of the biogenicity of organometallic assemblages.
7.2. Sulfolobales quinones as unique microbial fingerprints
Sulfur-thiophene-bearing quinones are unique biomarkers for metallophilic Sulfolobales. The S-containing molecules of benzodithiophenoquinones play an important role in electron transport and metal respiration. S/Fe-redox transforming members of the genera Sulfolobus, Acidianus, and Metallospharea have these unique Sulfolobales quinones, which occur specifically only in these archaea (Thurl et al., 1986; Elling et al., 2015). Respiratory quinones are usually identified based on exact molecular mass and fragmentation patterns in mass spectrometry (MS). These quinones comprise 0.28–0.38% of cell dry weight and represent ancestral biomarkers in terrestrial marine environments (Elling et al., 2015). Considering the high abundances of these respiratory quinones, their lower limit of detection was determined as <1 pg by using MS-assisted analysis (Elling et al., 2015). Moreover, these sulfur-thiophene-bearing signals can be examined by spectroscopy techniques, which could guide further in situ measurements to analyze collected venusian aerosol samples and possibly future returned samples. Experimental in situ MS measurements are desirable to accurately probe the venusian clouds for the presence of sulfur-thiophene-bearing quinones. Furthermore, promising new technologies of in situ transmission electron microscopy have been recently developed that provide extremely high magnification and resolution, allowing for sample study at the atomic level in real-time and under real environmental conditions (Dukes et al., 2013; Deng et al., 2019). Such in situ transmission electron microscopy measurements should be further developed in order to be employed for studying fine structural features of venusian aerosols, thus permitting a comprehensive analysis of possible metal-microbial interfaces
7.3. Microscopy of biosignatures in cloud layer particles
Life-signature detection instruments can be useful for astrobiological missions when using aerial platforms, as the presence of microorganisms in the clouds could be contributing to spectroscopic characteristics. The Life-Signature Detection Microscope (LDM) was conceptualized to obtain visible images of particles and search for signatures of life including possible “cells” in venusian cloud samples (Yamagishi et al., 2016; Sasaki et al., 2020). The resolution of 1 micrometer/pixel is sufficient for the observation of almost all terrestrial microbes and is likely sufficient for putative venusian microorganisms also.
In the LDM concept, a stain system is employed to distinguish biotic organic compounds from abiotic ones. The use of stains (fluorescence dyes) allows (1) detection of organic compounds including those of abiotic origin such as polyaromatic hydrocarbon (PAH), (2) highlighting of biotic organic compounds observed inside cells, (3) discernment of biotic organic compounds surrounded by a membrane, and (4) detection of the product of a catalytic reaction (Yamagishi et al., 2018b). The LDM system first stains a sample with the stain SYPRO Red (Rane et al., 2019); previous experiments have shown that SYPRO Red can stain not only biological cells but also PAH which is produced by both biotic and abiotic processes (such as being found in meteorites). Due to this characteristic of SYPRO Red, its use can potentially result in a false positive; however, this is ameliorated by different morphological features in biological cells compared to abiotic PAH. Cells are generally found in groups and show similar shape and size. Since cell membranes, and their contents, consist mostly of organic compounds, SYPRO Red fluorescence images of cells are full of similar round shapes with similar sizes, if they are cocci; abiotic sources of PAH do not have a similar appearance. In addition to SYPRO Red, the hydrophilic fluorescent dye propidium iodide (PI) can also be used; PI can only penetrate damaged bacterial membranes. It stains nucleic acids such as DNA and RNA inside injured or dead cells, emitting a red fluorescence. In combination with hydrophobic fluorescence dyes such as SYTO 24 or SYTO 9, PI can be used to determine the live/dead viability of cells (Taverniti et al., 2019). After the addition of PI-SYTO 24 mixture to cells, the latter can enter all cells, with or without damaged cell membranes, and bind to DNA and RNA, emitting a green fluorescence. These dyes are used together by choosing a suitable concentration that allows PI to replace SYTO 24, since both fluorophores approach the same DNA, resulting in a red (PI) fluorescent signal. Such co-staining results in dead cells emitting red fluorescence while live cells emit green fluorescence.
A reaction catalyzed by the enzyme esterase is found in most terrestrial microorganisms. The result of this metabolic reaction can be detected by 5-carboxyfluorescein diacetate acetoxymethyl ester (Calado et al., 2018; Marqueño et al., 2019). 5-Carboxyfluorescein diacetate (CFDA) is normally a nonfluorescent, nonpolar compound and can penetrate cell membranes like SYTO 24. When hydrolyzed CFDA is incorporated into a live cell, it will change into a carboxy fluorescein, and emit green fluorescence when irradiated with blue light (Yamaguchi and Nasu, 1997). The cytoplasm of putative venusian cloud microorganisms may maintain a more neutral pH compared to a rather acidic cloud environment, as many terrestrial acidophiles do; therefore hydrolysis of CFDA can occur within the cell. It has been previously shown that some of the abovementioned fluorophore solutions can stain cells in the presence of 2% hyperchloride or after irradiation by 80 Gy gamma rays or after heat treatment at 70°C, providing further potential opportunities to use this technology.
Some terrestrial acidophiles live in acidic conditions that partially overlap with the predicted acidity for some layers of the venusian clouds (pH -1.3 to ∼0.5) (Grinspoon and Bullock, 2007); and their size (1 μm or larger in diameter), biogenic materials (DNA for heredity, and phospholipids of the membrane), and metabolism can be detected with fluorescence microscopy. Preliminary experiments show that at a positive suspension pH, DNA stain emits fluorescence from the cell of A. ferrooxidans, presumably because of the relatively higher intracellular pH. Even for bacterial cells suspended in a 75% sulfuric acid solution, a mixture of stain and sodium hydroxide (NaOH) resulted in fluorescent cells that were discriminated from autofluorescent particles. CFDA in acidic conditions also showed fluorescence, and the methods for false-positive prevention have now been examined (Sasaki et al., 2020). So far, SYTO 24, PI, DAPI, and CFDA have shown fluorescence in Acidithiobacillus ferrooxidans cells in 75% sulfuric acid after pH neutralization using NaOH (Sasaki et al., 2020). Other stains are planned to be tested.
8. Simulating Experiments and Modeling
Finally, to understand whether terrestrial polyextremophiles could survive in venusian cloud-like conditions and if the methodology proposed above would work, we must conduct experiments under such conditions. Similar experiments have been conducted for Mars, Europa, and other planetary bodies (Abrevaya et al., 2011; Pavlov et al., 2018; Taubner et al., 2018; Warren-Rhodes et al., 2019).
While no cloud-based analogs exist in nature that fully represent the cloud environment of Venus (temperature, acidity, chemical composition, etc.), artificial analog chambers have been constructed that are able to partially reproduce the environment of interest. One such chamber is the Glenn Extreme Environments Rig (GEER) located at NASA's Glenn Research Center (Cleveland, Ohio, USA). The GEER has the capability to reproduce the chemical composition (8 gases +1 liquid), temperature (including 60°C, up to higher than 500°C), and atmospheric pressures (up to 92 atm) of Venus (Kremic et al., 2019). The GEER conditions can be customized for the physicochemical conditions of the clouds as described above and thus can simulate the necessary parameters throughout the potentially habitable layer of interest. Other similar chambers may also prove to be useful in these studies.
While it is critical to reproduce the physicochemical conditions of the clouds, we must also take into consideration that this hypothesized life is likely floating in the clouds as an aerosol. There is a lack of analog sites that offer long-lived aerosols, and we are unaware of studies that have successfully aerosolized bacteria in these conditions. However, despite these analog challenges, it is well known that bacteria can indeed survive at high altitudes in the terrestrial atmosphere, providing some evidence that life could exist in the clouds of Venus (see section 3 above) (Smith et al., 2013).
One of the best candidate extremophilic organisms for simulating experiments is A. ferrooxidans, a polyextremophile capable of performing Fe- and S-dependent metabolism under anaerobic conditions (Mirete et al., 2017). To explore its survival potential, the conditions in the lower layer of the venusian clouds in analog chambers should be reproduced.
Such analog experiments are proposed to include the following conditions: (a) composition of gases and their abundance (Taylor et al., 2018): carbon dioxide (96%; percent abundance), nitrogen (3.5%), argon (0.007%), carbon monoxide (0.004%), neon (0.0005%), sulfur dioxide (150 ppm), water vapor (30 ppm), carbonyl sulfide COS (4 ppm), heavy water HDO (3 ppm), hydrogen chloride (0.5 ppm), hydrogen fluoride (0.005 ppm), and atomic oxygen, hydroxyl, atomic hydrogen (traces) (b) particle densities ∼50 particles/cm3 (∼2–8 μm diameter) and 600 particles/cm3 (∼0.4 μm diameter) (c) mass loading ∼0.1–100 mg3
(d) temperature and pressure of Venus altitude range of 47.5–50.5 km (with values from approximately 90–60°C and 1.5–1 atm, respectively) (Krasnopolsky, 2019) (e) pH of -1.5 to 0.5 (Grinspoon and Bullock, 2007)
Of note for these proposed experiments is the chosen pH range, which is outside the current known pH limit of life (pH of -0.06). This pH refers to extremophilic organisms of terrestrial origin. However, putative venusian microorganisms have potentially been evolving for millions of years, initially inhabiting the surface of Venus before moving up to the clouds, and thus may have extended their bio-adaptability to this particular environment with extremely low pH values. Choosing a pH range outside of the known limits of terrestrial life will allow us to conclude the adaptiveness of the studied terrestrial extremophilic organisms through an experiment, such as a long-term evolution experiment in the environmental conditions of the venusian clouds. This of course assumes that the longer the survival time of a given organism, the greater the degree of its adaptation to the given environmental conditions. However, pH is just one of many factors limiting the survival of microorganisms on Earth, and it is impossible to isolate only this factor of the environment to carry out a correct analysis of adaptive constraints. Other physicochemical factors that will affect the microorganisms of interest shall also be taken into account.
These proposed experiments should examine
metabolites of the tested bacteria such as Fe2+ (in moles per minute per cell) (Ohmura et al., 2002)
the ability to reproduce, measured with bacterial mass gain (e.g., in the form of a biofilm on a carbon electrode)
The results from these simulation experiments on survival and proliferation of A. ferrooxidans (and other relevant microorganisms) could provide an understanding of how putative venusian microorganisms may operate under in situ conditions and can be used as the basis for metabolic modeling for further study.
9. Discussion
The exobiology concept in relation to Venus is the existence of a hypothetical habitable zone located in its cloud layer. This habitable zone is surrounded by aggressive environments that are extreme for known terrestrial life. It contributes to the modern definition of the habitable zone that is not only limited by the range of orbits around a star within which a planetary surface has suitable physicochemical conditions for life as we know it (circumstellar habitable zone) but also includes some locations within and out of the aforementioned zone where appropriate conditions can exist such as beneath the surface (e.g., deep biosphere) or in the atmosphere where organisms are capable of gaining energy from various redox reactions. Chemolithoautotrophic life based mostly on sulfur and iron compounds has been hypothesized to be present in the clouds of Venus (Cockell, 1999; Schulze-Makuch et al., 2004; Limaye et al., 2018a). Chemolithotrophy is the most ancient metabolic form of life, which enables the transition of energy from nonliving inorganic matter to energy for a living entity. The abundance of CO2 in the cloud layer creates appropriate environmental conditions to sustain chemolithotrophic life.
On Earth, some microorganisms are able to spend at least part of their life cycle in the clouds in spite of different hazards such as fast temperature changes, increased acidity and salinity, desiccation, free radicals, and high radiation (Amato et al., 2017; Delort et al., 2017). They survive in the clouds within small aerosols containing water. Venusian clouds, on the other hand, are hypothesized to consist of sulfuric acid–water droplets; therefore, putative venusian aerosolized microorganisms would have access to this water.
Chemosynthesis, through the use of sulfur and ferrous compounds, and photosynthesis are considered to be possible sources of energy for the putative venusian organisms. Overall, venusian microorganisms could be involved in complex redox cycles and maintain their biomass at a level necessary for survival. This may, in particular, be favored by the greater stability of the venusian cloud layer as an ecosystem, compared to the clouds on Earth.
The venusian clouds are characterized by their wide range of physicochemical parameters which include high temperatures and low pH. There are a few known terrestrial microorganisms capable of survival and reproduction in conditions of 50–80°C and pH 0–0.5, as estimated for the low cloud layer (Cockell, 1999; Grinspoon and Bullock, 2007). Additionally, some microorganisms are known to survive in the low temperatures of our upper habitable zone (up to 70 km) (Clarke et al., 2013). Acidithiobacillus ferrooxidans, Stygiolobus azoricus, Acidianus spp., Sulfolobus spp., and Metallosphaera spp. are bacteria and archaea that may serve as analog organisms due to their ability to reduce and/or oxidize various sulfur compounds and use ferric compounds as electron acceptors. Some of these microorganisms can also oxidize Fe2+ under aerobic conditions and hence perform opposite reactions important for iron cycling. In the venusian clouds, similar putative organisms can hypothetically use such a process with nitrogen instead of oxygen.
Dissimilatory nitrate reduction to ammonium (DNRA) and nitrate-reducing Fe(II) oxidation (NRFO) can be proposed as sources of oxidized forms of sulfur and iron in the venusian clouds. In terrestrial ecosystems, including extreme environments, such metabolic reactions contribute to the cycling of biogenic elements. A peculiarity of sulfur, one of the main elements of venusian clouds, is the existence of its various oxidation states. Sulfur compounds such as elemental sulfur (S0), sulfide (S2-), sulfate (SO4 2-), as well as sulfite (SO3 2-) and thiosulfate (S2O3 2-) are actively used by terrestrial microorganisms performing sulfur-centered metabolism. This makes it possible to consider various anoxic redox reactions using sulfur compounds (including reduction, oxidation, and disproportionation) as sources of energy for putative venusian organisms. Phototrophic organisms can also use elemental sulfur to couple its oxidation with the anoxygenic phototrophic reduction of CO2 for gaining energy.
As is the case for many extreme terrestrial ecosystems, a strong dependence on geochemistry may be relevant to the venusian cloud biosystem. Microorganisms in the cloud layer can be incorporated into an aerogeochemical process gaining energy in various redox reactions and in turn affecting their microenvironments by producing metabolites, such as ammonium, that to a certain extent can compensate for the acidity of the environment and increase the potential for survival. Additionally, physical organization is likely crucial for these putative microorganisms, as aerosols containing water are considered to be the environment most suitable for them to proliferate. All these features make the actual conditions in microenvironments especially important for putative venusian organisms. Therefore, such organisms likely exhibit an aerogeochemically dependent lifestyle.
In addition to having microbial mechanisms for survival in aggressive environments, it is also crucial that the metabolic reactions suggested for the clouds are thermodynamically feasible. High acidity and temperature can be thermodynamically favorable for some reactions like sulfur comproportionation; while it has not been discovered yet in terrestrial ecosystems, it can be taken into consideration. In principle, due to the unique set of physicochemical parameters of the venusian clouds, which do not have direct terrestrial analogs, astrobiologists may be facing unusual biochemistry and metabolisms in putative venusian microbes. Finally, the biogenic cycles of elements in the venusian clouds can be closely coupled with abiogenic cycles including various surface processes that can be sources of different compounds (see Limaye et al., 2018a, for detailed discussion).
Nevertheless, some terrestrial polyextremophiles are good models for selecting potential biomarkers to be used in life detection. The identification of relevant biosignatures and biomarkers and development of methods for their reliable detection should be included in the main goals for astrobiology missions to Venus. Future directions to help develop these models can be the detection of crucial metabolic intermediates and specific compounds, such as those used in response to extreme factors of the environment. Such detections should be made in aerosols, as these are microenvironments where microbial activity can be expected, as first described in the work of Limaye et al. (2018a) and further elaborated on in the work of Seager et al. (2020). A list of the most relevant biomarkers can be created based on our knowledge about compounds involved in stress-resistance mechanisms and metabolites characteristically produced by extremophiles. Here we make some suggestions that we believe will be well complemented in the future. Additionally, understanding the isotopic fractionation of key elements related to life can be an effective indicator of biological processes. Since living organisms prefer to use light isotopes for their metabolic reactions, stable isotope signatures of biogenic elements are useful biomarkers. These signatures provide key information not only on the biological, living or non-living, origin of organic remains but also on the pertinent metabolic pathways of formation and degradation of organic matter and related biominerals (Van Zuilen, 2008; Thomazo and Strauss, 2014; Kring et al., 2021).
The existence of putative microorganisms in the venusian cloud layer can be explained by the following: (1) their local emergence and evolution in the clouds, (2) emergence on the surface based on the possible existence of liquid water in the past and transport to the clouds via active volcanoes (Shalygin et al., 2015), or (3) the interplanetary transport of microorganisms that originated from Earth. The hypothesis of bacteria in the clouds is supported by several terrestrial analogs. On Earth, bacteria were found to be present at high altitudes as early as 1861 (Pasteur, 1861). Since then, metabolically active microbes have been detected in the atmosphere at altitudes of 20–70 km (Imshenetsky et al., 1978; Griffin, 2004; Wainwright et al., 2004; Bowers et al., 2009; Womack et al., 2010). Additionally, bacteria have been shown to survive long-distance transport, in addition to living and reproducing in vitro in water collected from aerosols (Dimmick et al., 1979). Even raindrops help bioaerosol generation (Joung et al., 2017). It should be noted, however, that there is no evidence of a complete microbe life-cycle taking place while airborne (Smith et al., 2013). Such bacteria likely originate from high-altitude habitats, such as alpine sites, or are transported to high altitudes via air currents. Rapid upward transport of bacteria can result from natural processes such as storm activity, volcanic activity, and impact events, as well as human activity such as weapons testing and aviation and spacecraft launches (Hall and Bruch, 1965; Bucker and Horneck, 1969; Simkin and Siebert, 1994; Kring, 2000; Griffin et al., 2002; Griffin, 2004). Generally, terrestrial microbes have adapted to every ecological niche on Earth. Keeping in mind that ancient Venus potentially harbored a shallow liquid ocean and had a similar climate as ancient Earth, the chance of life emergence and its transport into the atmosphere cannot be denied.
The basic survival strategy needed by putative microorganisms to survive the environmental conditions of the venusian clouds is polyextremophily. Evolution in such an environment can potentially result in microbial forms exhibiting a combination of properties found in terrestrial extremophiles not found in any single known organism. In fact, hot springs and deep-sea hydrothermal vents can be model systems for studying putative venusian organisms in relation to survival at high temperatures (characteristic of the active habitable zone defined above), low pH, and basic metabolic processes with sulfur and ferrous compounds. Meanwhile, microbial life in terrestrial clouds is an analog of the hypothetical venusian microbial system with respect to physical organization and stress factors such as desiccation and radiation. We have to take into account the characteristics of these two terrestrial microbial systems while creating a habitability concept for the cloud layers of Venus, since we do not have direct analogs of such a system on Earth. Acidithiobacillus ferrooxidans may be the most useful model organism for studying viability in simulating experiments. It thrives at extremely low pH values, tolerates elevated temperatures, fixes both CO2 and N2 from the atmosphere, and gains energy performing three different forms of respiration, including H2/Fe3+, H2/S0, and S0/Fe3+ under anaerobic conditions. Regardless of the model organism, it is important to develop a technique of aerosolization of the organism to properly simulate the aerial environment of venusian clouds to study the specific ecology of this microhabitat.
Finally, modification and development of novel methodology for the detection of biosignatures under very aggressive environmental conditions should also be of focus for the proper detection of life in the venusian clouds. One possible biomarker is the detection of organometallic signatures such as nanoscale iron/sulfur microbial interfaces and sulfur-bearing specific organic biomarkers (thiophenes from Sulfolobus quinones). Efficient life-detection microscopy and nanoanalytical spectroscopy should be developed in order to assess the biogenicity of samples collected at Venus. Other methods may include the search for chemical species potentially produced in biological ecosystems, such as methane, hydrogen sulfide, nitrous oxide, and probably phosphine, should a biological origin in terrestrial ecosystems be confirmed in addition to its presence in the venusian atmosphere.
The collected knowledge will give insights into the hypothetical ecosystem of venusian clouds and motivate further experimental and theoretical work to build up a detailed understanding of the full functional setup of life based on inorganic sources elsewhere.
Footnotes
Acknowledgments
The work presented here was motivated by fruitful dialogue at the 2019 Venus Cloud Layer Habitability and Landing Site Selection workshop organized by the Roscosmos-IKI/NASA Venera-D Joint Science Definition Team and supported by NASA HQ Planetary Science (A. Ocampo, Lead Venus Scientist) and Astrobiology programs (M. Voytek, Senior Scientist for Astrobiology) and the Space Research Institute of the Russian Academy of Sciences (IKI RAN). JAC acknowledges the support of the Genome Sciences Training Program at University of Wisconsin–Madison. TM is grateful to the Austrian Science Fund (FWF) for providing support through the Elise-Richter Research fellowship (V333). We thank Sanjay Limaye for his support, including of this publication, and for resparking the conversation on Venus astrobiology.
Author Disclosure Statement
No competing financial interests exist.
Associate Editor: Lewis Dartnell