
Abstract
Homochirality is a feature of life, but its origin is still disputed. Recent theories indicate that the origin of homochirality coincided with that of the RNA world, but proteins have not yet been incorporated into the story. Ribosome is considered a living fossil that survived the RNA world and records the oldest interaction between RNA and proteins. Inspired by several ribosome-related findings, we propose a hypothesis as follows: the substrate chirality preference of some primitive peptide synthesis ribozymes can mediate the chirality transmission from RNA to protein. In return, the chiral preference of protective peptide-RNA interaction can bring these ribozymes an evolutionary advantage and facilitate the expansion of enantiomeric excess in peptides. Monte Carlo simulation results show that this system's chemistry model is plausible. This model can be further tested through investigation of the chirality preference for the interactions between
1. Introduction
Homochirality is a significant feature of life. Only one of two enantiomers of chiral sugars and amino acids was used to build biological macromolecules such as RNA, DNA, and proteins. The origin of homochirality has attracted scientists' attention for more than a century. Many theories probing this issue attribute it to asymmetrical physical and chemical factors that may cause the symmetry break in the chiral small molecules. However, these theories are not faultless due to the weak effects or harsh preconditions for the physical and chemical factors (Blackmond, 2019). Other theories proposed that homochirality was achieved during the formation of biological macromolecules. RNA has always been a research focus of prebiotic chemistry, because of its catalysis and replication capabilities. The RNA world hypothesis (Crick, 1968; Orgel, 1968; Gilbert, 1986; Higgs and Lehman, 2015) could be one of widely accepted scenarios for the origin of life. The chirality of RNA is derived from its component sugar, that is, ribose. Meteorites/impactors may have brought ribose to Earth (Furukawa et al., 2019); excesses of the
Compared with RNA, proteins are difficult to self-replicate, and there is no enantiomeric cross-inhibition in the nonenzymatic polymerization of amino acids. Therefore, although the meteorites also brought excess
Some earlier studies found that
Is there a simpler system that also can fulfil both critical points in Sandars' theory? Some recent findings with regard to ribosome function have shown that the chirality transmission from RNA to protein may have occurred earlier than the origin of genetic codons and been carried out by the primitive simple protein synthesis system. In this paper, we review these studies and propose a systems chemistry model in which the chiral preference of peptide synthesis and protective peptide binding mediate the chirality transmission from RNA to protein.
2. Ribosome and Its Chiral Preference of Substrates
Ribosome, as the core of protein translation, could be a molecular fossil that reveals the early interactions between RNA and proteins (Harish and Caetano-Anollés, 2012; Petrov et al., 2015) and may also contain clues about the chirality transmission between them. The modern ribosome is composed of a small subunit (SSU) that mainly carries out codon recognition that cooperates with tRNA's bottom half; the ribosome's large subunit (LSU), however, catalyzes peptide bond formation and cooperates with tRNA's top half. The early evolution of LSU and SSU is deduced to have been a separate process, and so were the two parts of the tRNA (Fox, 2010; Petrov et al., 2015). The most ancient part of the ribosome was supposed to be the rRNA in the peptidyl transferase center (PTC) locating at LSU where the peptide bond synthesis occurs (Fox, 2010; Petrov et al., 2015). The tRNA top half can form a minihelix structure alone that can attach amino acids, and its CCA tail that directly attaches to amino acids is extremely conserved in the phylogenetic tree. Therefore, the tRNA top half is considered to be older than the bottom half (Fox, 2010). These findings depict a scenario for the primitive enzymatic peptide synthesis system (protein may not be available at the time) in which PTC-RNA-like ribozymes catalyze the condensation of amino acids with the help of the CCA-tailed proto-tRNA, while the coding and decoding components of modern translate system (mRNA, SSU, and tRNA bottom half) were not yet integrated into the system (Petrov et al., 2015).
Modern biological peptide synthesis is a two-step process. Amino acids are first attached to the CCA tails of the tRNA top half through the aminoacylation reaction. Then, the generated aminoacyl-tRNA enters the ribosome, and the condensation of amino acids occurs in the PTC. Both aminoacylation and PTC-catalyzed condensation have preferences for the chirality of the substrate. Tamura and Schimmel (2006) performed a nonenzymatic aminoacylation reaction between a phosphate oligonucleotide-modified alanine and an RNA minihelix that imitates the ancient tRNA top half. In the experiment, the reaction efficiency of
Recently, PTC-catalyzed peptide bond formation of
3. A Systems Chemistry Model for Protein Homochirality Origin
The tRNA CCA tail and PTC rRNA originated very early, so the above-mentioned two kinds of chiral preferences of substrates may have also existed in the primitive peptide synthesis system and enabled chirality transmission from RNA to peptides. Inspired by these findings, we propose a systems chemistry model for the origin of protein homochirality (Fig. 1) as follows: the primitive peptide synthesis system consisting of CCA-tailed proto-tRNA and PTC RNA-like ribozymes prefers
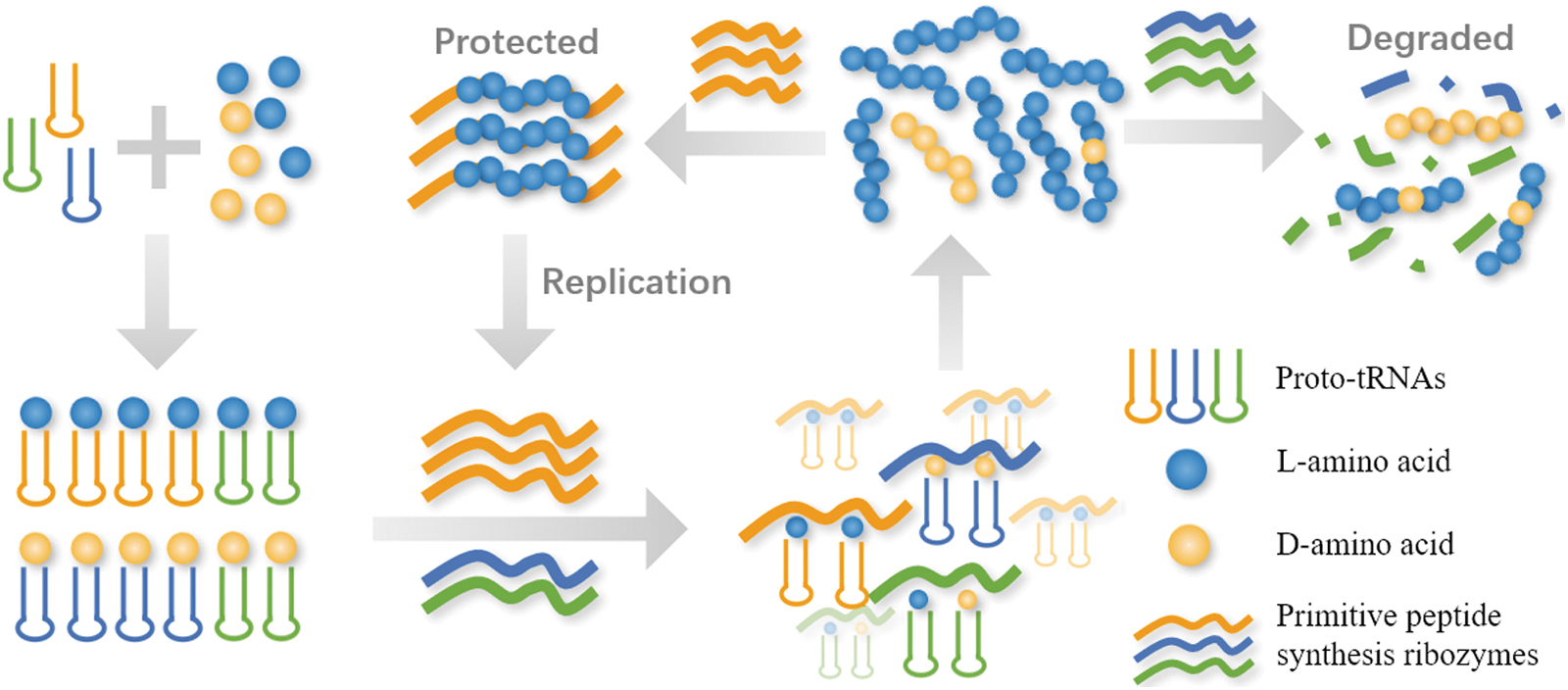
A systems chemistry model for the origin of protein homochirality. In a preexisting
In our model, RNA is the bearer of chirality information in a different way from Sandars' theory. RNA implements this function through substrate selectivity instead of working as codons. RNA's self-replicate ability can enable the nonlinear expansion of enantiomeric excess. It is noteworthy that the early evolution of codon recognition and peptide synthesis could be separate. The integration of the two functions may have occurred at a later stage of peptide synthesis system evolution (Petrov et al., 2015). The independence of the coding and decoding functions means that our model can make chirality transmission occur in an earlier and simpler peptide synthesis system.
It has been found that introducing mutations in the PTC and/or its adjacent regions can elevate the use of
A reasonable answer to this question is that chiral preference of substrates can bring an evolutionary advantage to the peptide synthesis system. In Sandars' theory, proto-cells with a protein synthesis system that produces homochiral proteins survived the competition because homochirality improved the protein functions by facilitating the formation of secondary structures (2005). As discussed before, this theory relies on complex systems that were unlikely available before the achievement of homochirality. Fox (2010) also supposed that the exclusive requirements for substrate chirality of the modern ribosome are the result of selection, but did not explain how producing homochirality can bring advantages in evolution.
Here, we suggest that an evolutionary advantage may be achieved by the protection of peptides on RNA with a chiral preference. The concept that peptides can protect RNA was presented in the work of Szostak, who suggested that short peptides containing acidic residues play a very important role in protecting RNA molecules against Mg2+-induced degradation by chelating these ions (2012). Mg2+ is ubiquitous in the environment and has been found to help the LSU rRNA collapse into a native structure (Van der Gulik and Speijer, 2015; Lenz et al., 2017). The modern ribosome's peptidyl transferase activity is also based on Mg2+ (Miskin et al., 1970). Therefore, it is reasonable to assume that peptides were employed by the ancient PTC RNA-like ribozymes to maintain their stability. Moreover, binding with peptides can reduce the chance of being attacked by water molecules, which is a major cause of biomolecule degradation (Bains et al., 2015). Interestingly, rRNA was proposed to function prebiotically as mRNA and code its protector proteins (Root-Bernstein and Root-Bernstein, 2015, 2016). If so, the rRNA-coded proteins may replace the protective peptides, which were used by the primitive synthesis system after the integration of coding and decoding components.
We further assume that the peptide-RNA interaction (PRI) has a chiral preference. No reports have been found to directly support this assumption, but some observations have shown that
For the primitive PTC RNA-like ribozymes, peptides synthesized by themselves were naturally the first choice for protectors. If the interaction between them also had a chiral preference as we assume, ribozymes that synthesize peptides with
4. Monte Carlo Simulations
To verify whether the chiral preference of protective PRI can bring an evolutionary advantage to the peptide synthesis systems that produce peptides with
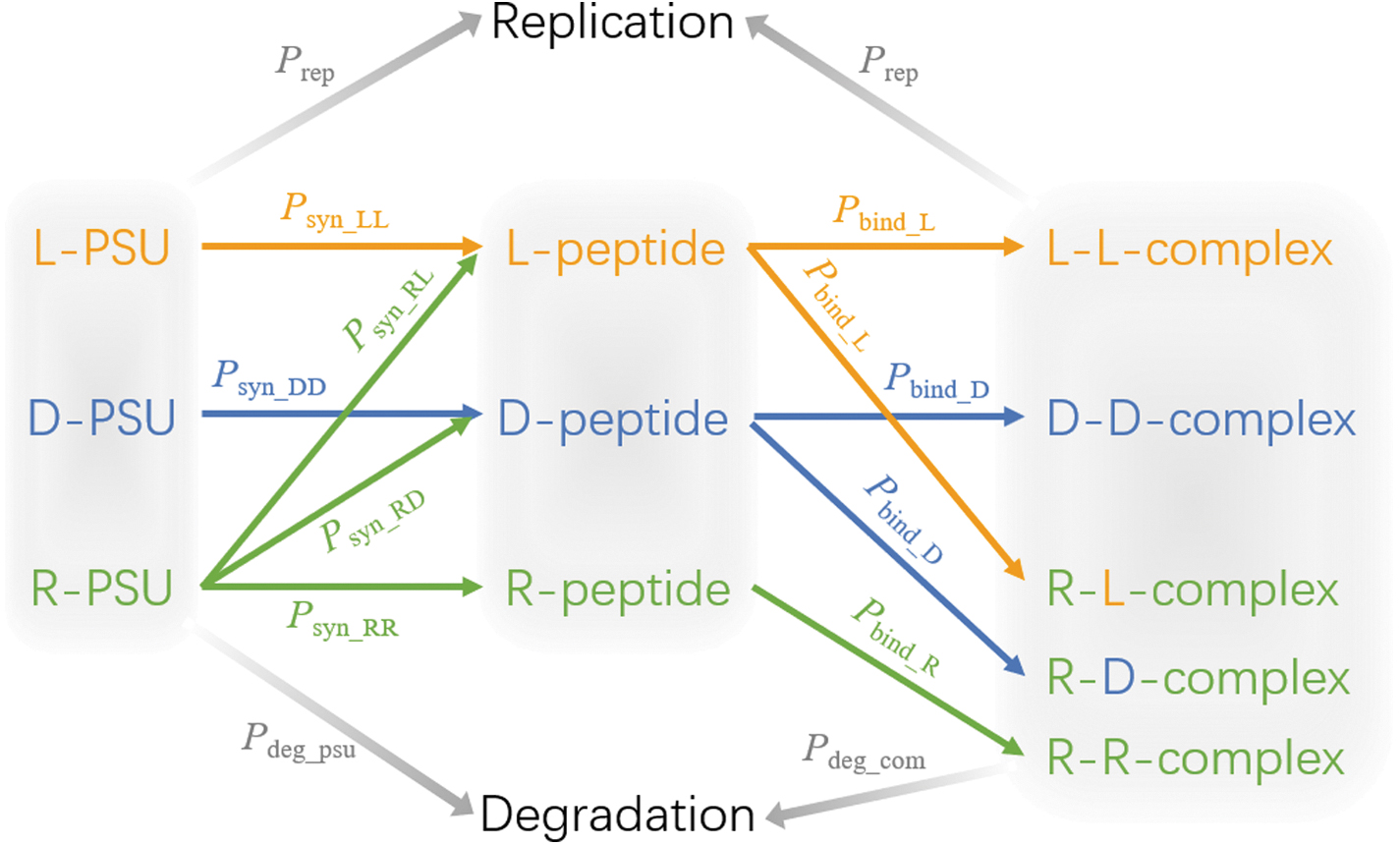
Monte Carlo simulation procedure. The simulated system is composed of peptide synthesis units (PSU), peptides, and the complex of PSU and peptides. These components are linked by the events that occurred in the simulation (arrows). Descriptions of these components and events are listed in Table 1. In one simulation step, these events go through in the following order: peptide synthesis, complex formation, degradation, and replication. The simulation is implemented in Python 3.6, and the corresponding script can be found at
Description of Monte Carlo Simulation System
Because most peptides synthesized by
We first assumed that the three PSUs accounted for the same proportion in the initial population and compared the changes of PSU population composition under different PRI chiral preference degrees and protective effects. When the protective effect does not exist, that is, the peptide-PSU complex degrades as readily as PSU (P
deg_psu = P
deg_com = 0.5), the population and composition of PSUs remain the initial values during the 1000-step simulation, no matter how the chiral preference of PRI changes (Fig. 3A). However, when the peptide can protect PSU (P
deg_psu = 0.5, P
deg_com = 0.05), and

Monte Carlo simulation results. The horizontal axis represents Monte Carlo simulation steps. Lines represent the ratios of
According to the opinions of Sandars (2005) and Fox (2010), most of the early peptide synthesis ribozymes may not distinguish the chirality of amino acids. To simulate this situation, the initial ratio of
5. Discussion
In summary, the present model provides a plausible scenario of chirality transmission from RNA to peptides. This scenario assumes the preexistence of a homochiral
Traces of the ancient protective peptides may remain in the modern ribosome. Most universal ribosome proteins (rProteins) are composed of globular domains arranged on the ribosomal surface and random coil tails extended into the core near the PTC (Kovacs et al., 2017). The tails may have had an earlier origin than the rest of the rProteins. The rProtein L2 in LSU is one of the oldest rProteins, and its tail is the protein component closest to PTC (Kovacs et al. 2017). L2 tail was found to interact with a magnesium microcluster, while the latter also interacts with PTC (Petrov et al., 2012). These findings suggest that rProtein tails may root in the ancient protective peptides that bind PTC RNA homologs, and the ancient RNA protector peptides may also lack fixed structures. Fox (2010) suggested that, in the primitive ribosome, the corresponding components of the rProtein tails could have been partially chiral peptides (peptides containing more
In the present study, we assume that protein synthesis relies on RNA, which follows the opinion of RNA world theory and is consistent with what happens in all existing organisms, but also means that chirality is directionally transferred from RNA to protein. However, RNA may not have been necessary for small protein synthesis in the primitive world (Canavelli et al., 2019). Recently, Skolnick et al. (2019) proposed a scenario of protein homochirality origin in which RNA was not involved. They found that homochiral proteins have longer secondary structures and are thermodynamically more stable than the proteins built from amino acids of mixed chirality, and thus homochiral proteins can be retained in natural selection. Interestingly, some of the most ancient protein structures such as P-loop can bind RNA (Romero et al., 2018), which suggests an ancient origin of PRI that may have been earlier than the establishment of the homochirality of the RNA world. Combined with the protection of peptides on RNA, these findings imply a possibility that the chirality of protein may also have been transferred to RNA. The abundance of different enantiomers of peptides may also have shaped the chirality preference of the primitive peptide synthesis system and even affected the RNA world's choice of chirality. This bidirectional influence between RNA and peptides can form a positive feedback loop, which amplifies the initial slight chirality imbalance and eventually achieves homochirality. Root-Bernstein (2007, 2010) suggested a similar concept, in which the main players are RNA trinucleotide codons and amino acids. Nevertheless, the chiral preference in RNA-amino acid interaction has not been demonstrated to be prevalent in all amino acids, nor has it been demonstrated as to whether it can affect the RNA-peptide interactions. To test our model, the chirality preference of RNA-peptide interactions will require further investigation, in particular examination of those interactions among
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31870837).
Author Disclosure Statement
The authors declare that they have no competing financial interests.
Abbreviations Used
Associate Editor: Norman Sleep