
Abstract
Martian lava tube caves resulting from a time when the planet was still volcanically active are proposed to contain deposits of water ice, a feature that may increase microbial habitability. In this study, we taxonomically characterized and directly measured metabolic activity of the microbial communities that inhabit lava tube ice from Lava Beds National Monument, an analogue environment to martian lava tubes. We investigated whether this environment was habitable to microorganisms by determining their taxonomic diversity, metabolic activity, and viability using both culture-dependent and culture-independent techniques. With 16S rRNA gene sequencing, we recovered 27 distinct phyla from both ice and ice–rock interface samples, primarily consisting of Actinobacteria, Proteobacteria, Bacteroidetes, Firmicutes, and Chloroflexi. Radiorespiration and Biolog EcoPlate assays found these microbial communities to be metabolically active at both 5°C and −5°C and able to metabolize diverse sets of heterotrophic carbon substrates at each temperature. Viable cells were predominantly cold adapted and capable of growth at 5°C (1.3 × 104 to 2.9 × 107 cells/mL), and 24 of 38 cultured isolates were capable of growth at −5°C. Furthermore, 14 of these cultured isolates, and 16 of the 20 most numerous amplicon sequences we recovered were most closely related to isolates and sequences obtained from other cryophilic environments. Given these results, lava tube ice appears to be a habitable environment, and considering the protections martian lava tubes offer to microbial communities from harsh surface conditions, similar martian caves containing ice may be capable of supporting extant, active microbial communities.
1. Introduction
Presently, the average temperature on the surface of Mars is −60°C, together with extremely dry conditions, and atmospheric pressures of ∼6 mbar (1% of that on Earth; Merino et al., 2019) create an environment inhospitable to life. In addition, surface soils are extremely oxidizing and receive large doses of ultraviolet (UV) and ionizing radiation due to Mars' thin atmosphere and lack of a magnetic field (Hassler et al., 2014; Merino et al., 2019). Given these harsh surface conditions, martian caves, specifically lava tubes, may be better environments to search for signs of extinct or extant life (Boston et al., 2001; Léveillé and Datta, 2010; Northup et al., 2011; Uckert et al., 2017).
Lava tube caves form in the shallow subsurface of volcanic regions after cooling of a basaltic lava flow's exterior, leaves hollow voids below (Léveillé and Datta, 2010). It is understood that lava tubes can form on any rocky celestial body, which is or was volcanically active, with strong evidence to suggest that these subsurface features are extensive in the volcanic provinces of Mars (Carr et al., 1977; Hodges and Moore 1994; Wyrick et al., 2004; Cushing et al., 2007; Léveillé and Datta, 2010; Farrand et al., 2011; Cushing 2012).
Like terrestrial lava tubes, martian lava tubes should theoretically provide protection from UV radiation, ionizing radiation, and surface dust storms; decrease temperature variation; and provide energy sources for microorganisms, such as Fe2+, Mn2+, H2S, H2, and CO2 from alteration of mafic rock (Léveillé and Datta, 2010). Most lava tubes on Mars are geologically old, although there is evidence to suggest that volcanism has occurred between 3.7 Gya and 100–200 Mya (Werner, 2009). Nevertheless, lava tubes are likely much longer lived on Mars than on Earth due to lower tectonic/seismic activity and rock weathering. In addition, modeling suggests that lava tubes located in the volcanic provinces of Mars may contain stable or accumulating water ice, even today (Williams et al., 2010), although this has not clearly been observed yet.
On Earth, water can remain liquid within ice by two mechanisms. The first is due to the insolubility of ionic impurities, which create micron-sized veins of liquid water to temperatures as low as −90°C (Price and Sowers, 2004), whereas the second forms when mineral grains contact ice forming a liquid film around the mineral surfaces, with both mechanisms accepted to occur on Mars (Price, 2007).
Despite subfreezing temperatures, metabolically active microorganisms have been observed within ice (Carpenter et al., 2000; Rivkina et al., 2000; Jakosky et al., 2003; Junge et al., 2004; Steven et al., 2008; Chan-Yam et al., 2019) and to temperatures as low as −33°C (Bakermans and Skidmore, 2011). Thus, the combination of shelter and bioavailable liquid water within ice could have made martian lava tubes a potential refuge to microbial life, especially after Mars' climate transitioned from wet and warm to cold and dry ∼3.7–3.0 Gya (Ramirez and Craddock, 2018). Therefore, terrestrial lava tubes that present similar characteristics are considered to be important analogues for studying habitability in the martian subsurface (Boston et al., 2001; Léveillé and Datta, 2010; Northup et al., 2011; Hathaway et al., 2014).
To date, microbial research of lava tubes has largely focused on microbial mats (Hathaway et al., 2014; Riquelme et al., 2015; Lavoie et al., 2017; Gonzalez-Pimentel et al., 2018) and microbe/mineral interactions (Boston et al., 2001; Northup et al., 2011; Ríos et al., 2011; Uckert et al., 2017). However, relatively few studies of lava tubes have focused on cryoenvironments, that is, those caves which contain ice or where freezing temperatures exist. Such environments are better analogues for martian lava tubes given the freezing temperatures likely encountered even in the shallow subsurface.
One culture-based study by Popa et al. (2012) recovered microbes from the basalt–ice interface within a terrestrial lava tube capable of metabolizing iron from the mineral olivine. Teehera et al. (2017) conducted a taxonomic characterization of the microbial communities found associated with secondary minerals and ice from two lava tubes in Hawaii. Despite these limited studies, a knowledge gap remains regarding the identity, viability, and metabolic activity of the microbial communities present within lava tube ice.
The goal of the present study was to determine the structure of the microbial community, together with the cold temperature adaptability and metabolic activity of microorganisms inhabiting lava tube ice from Lava Beds National Monument (LABE). These findings are important to understanding the microbial limits of life in Earth's subsurface, determining the applicability of using lava tube ice caves as terrestrial analogues for subsurface martian environments, and determining whether such environments could support life.
2. Materials and Methods
2.1. Sample site description and sample collection
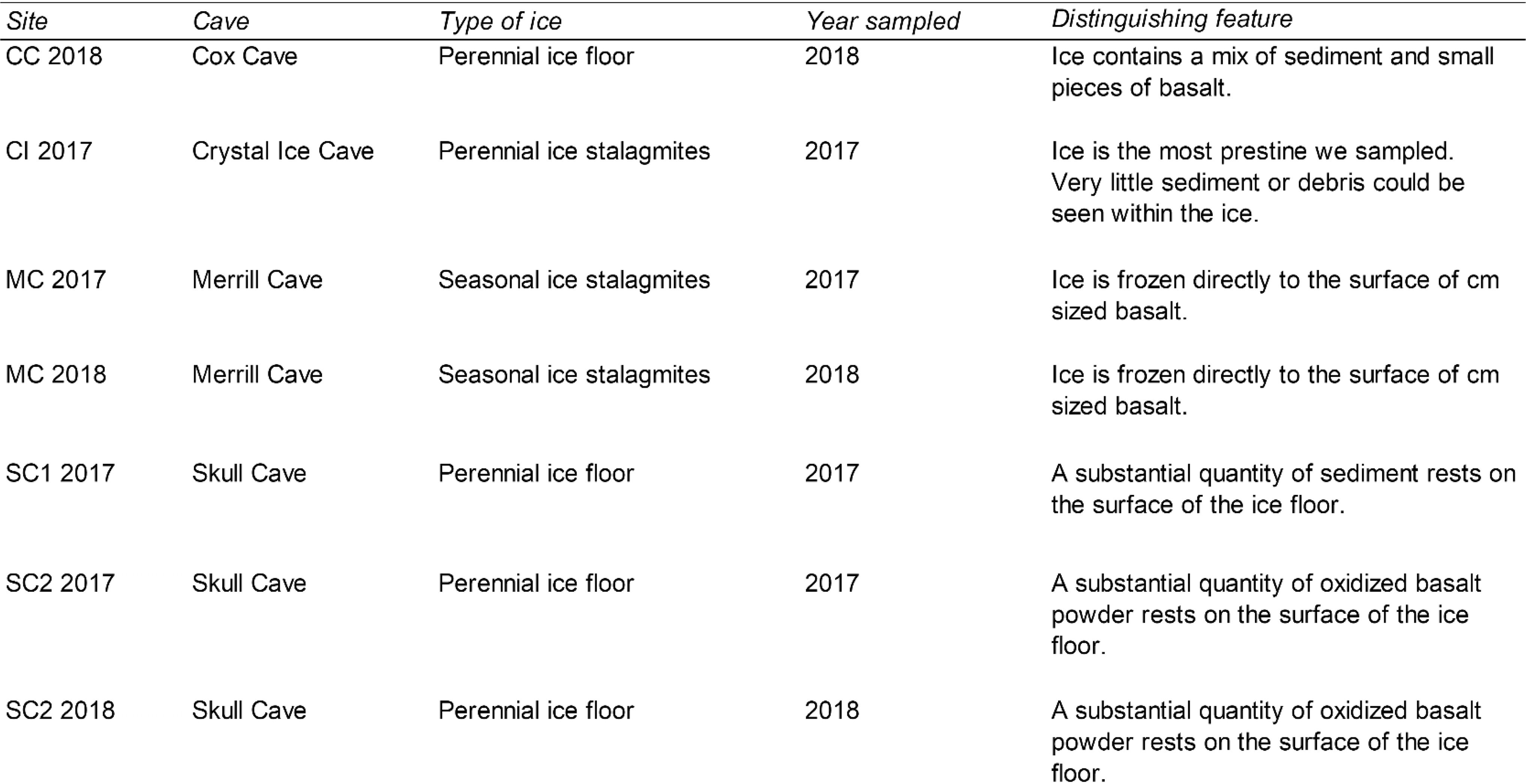
LABE, whose climate is classified as a semi-arid desert, is located in Northern California (41° 42′ 50″ N, 121° 30′ 30″ W) on the northeastern flank of the Medicine Lake Volcano. This area contains the largest concentration of lava tubes in North America, with >800 discovered to date (personal communication with park staff). Many of these caves contain large seasonal or perennial ice deposits (Fig. 1). Within LABE, the average annual air temperature fluctuates between −0.5°C in the winter and 15.8°C in the summer; the mean annual precipitation varies between 12.2 cm in the winter and 4.8 cm in the summer (Adamus et al., 2013).

Location and pictures of sampling sites.
Ice samples were collected during the summer of 2017 and 2018 at sites chosen based on the different types of ice they contained (i.e., perennial vs. seasonal and clear vs. sediment containing). Unfortunately, due to site restrictions, not all caves and sites could be sampled both years. A description of each sampling site can be found in Table 1. Samples were obtained with flame and ethanol-sterilized chisels and collected in sterile Whirl-Pak bags, which were kept frozen at −20°C during transport and storage at McGill University to await further analysis.
Descriptions of Lava Beds National Monument Cave Ice Samples
2.2. Physicochemical analyses
The pH of the ice samples was measured in the field by submerging pH strips in melted surface ice. The temperature of the ice was determined by either placing a handheld temperature probe directly on the ice surface or placing the probe ∼4 cm into the ice by chiseling a small divot. Processing of ice samples for further physicochemical analyses was conducted in the laboratory by melting and filtering the ice at room temperature through a 0.45 μm nitrocellulose filter.
Total organic carbon (TOC) was then measured by the UV/persulfate oxidation method on a Sievers Innovox TOC Analyzer (General Electric Power and Water, Water and Process Technologies, Boulder, CO) following standard protocols. Analyses of nitrate, ammonia, and phosphate concentrations were prepared following the manufacturer's instructions and measured colorimetrically on a multichannel Lachat autoanalyzer (Lachat Instruments, Milwaukee, WI).
2.3. Cell enumeration, isolation, cultivation, and characterization
To determine the total number of viable microorganisms present within our samples, colony-forming units (CFUs) were enumerated on Reasoner's 2A (R2A) agar as follows: melted ice was diluted to 10−2, 10−3, and 10−4 in 0.1% Na4P2O7 buffer, pH 7, and plated on agar. To determine the effect of growth temperature on the total CFU, three replicates of each dilution were incubated at three temperatures (5°C for 28 days, 23°C for 14 days, and 37°C for 7 days) before counting the total CFU.
To determine whether the viable microbial population was cold adapted, we isolated those CFU that presented unique morphologies on plates incubated at 5°C and 23°C from previous analyses. The ability of each isolate to grow at a range of temperatures was further tested on R2A agar at 37°C, 23°C, 5°C, 0°C, and at −5°C on R2A+7% sucrose to prevent the agar from freezing. The ability of each isolate to grow in 0%, 3%, 6%, 9%, 12%, 15%, 18%, and 21% NaCl was also tested on R2A agar at 25°C except those isolates that could not grow at 25°C, which were grown at 5°C.
Sanger sequencing was performed to determine the identity of each isolate. First, isolated colonies were lysed by mixing with 250 μL of deionized H2O and heating in a microwave for 3 min. Polymerase chain reaction (PCR) amplification of the 16S rRNA gene was performed with primers 27F (5′-AGRGTTTGATCMTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTACGACTT-3′), which amplify nearly the full length of the gene (∼1500 bp). The 16S rRNA PCR cycling conditions were as follows: (1) 95°C for 7 min, (2) 94°C for 45 s, (3) 55°C for 45 s, (4) 72°C for 1 min (where steps 2–4 were repeated 30 times), (5) 72°C for 10 min.
Amplicon DNA was sent to the Plate-forme d'Analyses Génomiques de l'Université Laval (Quebec City, Quebec, Canada) for sequencing. Low-quality sequences were trimmed, and miscalled bases corrected, with Sequencher version 5.4.6 (Ann Arbor, MI). Sequences were then compared against the GenBank database by using BLASTn to look for the most similar sequence to the query sequence.
2.4. Nucleic acid extraction and 16S rRNA gene sequencing
Five replicate extractions were performed on each ice sample by using the Qiagen DNeasy PowerLyzer PowerSoil DNA Extraction Kit (Qiagen, Venlo, Netherlands), following the manufacturer's instructions but with slight variations in the final elution step: DNA was eluted in 50 μL (20 μL for samples from Crystal Ice Cave) of molecular grade H2O rather than elution buffer. Sample replicate extractions were then pooled together, and DNA concentrations were then checked on a NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific, Waltham, MA) and concentrated using a ThermoSavant DNA 110 SpeedVac (Thermo Fisher Scientific) if the concentration was found to be too low for sequencing library preparation.
Sequencing libraries were prepared following Illumina's 16S metagenomic sequencing library preparation protocol. Primer pair 515F-Y (5′-GTGYCAGCMGCCGCGGTAA) and 926R (5′-CCGYCAATTYMTTTRAGTTT) (Parada et al., 2016), targeting the V4–V5 hypervariable region of the prokaryotic 16S rRNA gene, was used to amplify DNA for sequencing. Sequencing libraries were constructed from the amplified PCR product by using the Illumina Nextera XT Library Preparation Kit. Libraries were loaded on an Illumina MiSeq sequencer with MiSeq Reagent Kit v3 (Illumina, CA), and the sequencing run was carried out for 500 cycles. Sequence data was submitted to NCBI under BioProject accession number: PRJNA682127.
2.5. Sequence analysis
After sequencing, reads were grouped into amplicon sequencing variants (ASVs) with the DADA2 package for R (Callahan et al., 2016). Settings and parameters for sequence analysis were chosen by following the DADA2 Pipeline Tutorial (1.12) (
Alpha and beta diversity indices were calculated in R v 4.0.2 (R Core Team, 2020). The “vegan” package (Oksanen et al., 2019) was employed to calculate the Shannon diversity index (here referred to as H′-tax) and Bray–Curtis distances on ASV distribution tables; “ape” (Paradis and Schliep, 2019) and “phangorn” (Schliep, 2011) were employed to create a neighbor joining (NJ) tree (best parsimony results) with “GTR” nucleotide substitution model on ASV fasta sequences, previously aligned using Clustal Omega software (Sievers et al., 2011); the “picante” package (Kembel et al., 2010) was employed to calculate Faith's phylogenetic diversity (Faith's PD) using the NJ tree and ASV distribution tables, whereas “GUniFrac” (Chen, 2018) was employed to calculate UniFrac distances using these same parameters.
To determine the functional potential of lava tube ice microbial communities, 16S rRNA ASVs obtained from the lava tube ice were analyzed with the functional prediction pipeline Piphillin (Iwai et al., 2016), employing a 99% sequence identity cutoff against the May 2020 release of the Kyoto Encyclopedia of Genes and Genomes database (Kanehisa and Goto, 2000).
2.6. Community-level physiological profiling
To assess microbial activity and heterotrophic carbon substrate utilization among the microbial communities of LABE ice, samples were incubated in Biolog EcoPlates at both 5°C and −5°C. Melted ice samples were diluted 10−2 in 0.1% Na4P2O7 buffer, pH 7, both to remove sediment particles, which could interfere with spectrophotometric readings, and to lengthen the time required to detect substrate utilization. One hundred fifty microliters of diluted sample was then added to the wells and incubated in the dark at 5°C and −5°C. We believe that the dye, buffer, and substrates contained within the wells reduced the freezing point of water, which prevented freezing of plates incubated at −5°C (see Supplementary Fig. S1 for the EcoPlate experimental setup).
Absorbance changes were monitored weekly at a wavelength of 595 nm with a SpectraMax M2e Multi-Mode Microplate Reader (Molecular Devices, San Jose, CA). Wells were considered to be positive when their OD595 value reached above 0.25 (Garland, 1996, 1997). The average well-color development (AWCD) (time to reach plateau phase) (Garland and Mills, 1991; Iţcuş et al., 2016), substrate richness (R), Shannon–Weaver diversity index (here, referred to as H′-clpp), and Shannon substrate evenness (E) (Garland, 1997; Iţcuş et al., 2016) were calculated in triplicate once the AWCD hit the plateau phase (Pessi et al., 2012), corresponding to 53 days at 5°C and 102 days at −5°C. The AWCD = ∑ODi/31, R = number of metabolized substrates, H′-clpp = −∑pi(lnpi), where pi = ODi/∑ODi and E = (H′-clpp)/lnR.
A two-tailed t-test was performed on the data to determine whether substrate utilization differed significantly between incubation temperatures. Bray–Curtis distances were calculated on the Biolog EcoPlate substrate absorbance data (standardized and log-normalized) found for each sample, together with physical and chemical factors measured on these same samples. This distance matrix was calculated and visualized by using nonmetric multidimensional scaling (NMDS) with R package “vegan” (Oksanen et al., 2019).
2.7. Measurement of microbial activity using a radiorespiration assay
Radiorespiration assays were configured according to the protocol developed by Steven et al. (2007b). Microcosms were set up in triplicate with 5 mL of melted ice; the assay was then incubated at both 5°C and −5°C. Sterilized negative controls (autoclaved twice with a 24-h interval) were also included in triplicate for all samples at both temperatures. Each microcosm was spiked with 20 μL of 0.041 μCi (∼90,000 dpm) of 1,2-14C acetic acid and 20 μL of cold acetic acid to a final concentration of 15 mM acetic acid in a total volume of 40 μL. To create the CO2 trap, 500 μL of 1 M KOH was added to an open top vial within the microcosms. Measurements of CO2 were taken periodically over a period of 483 days (−5°C incubations) and 447 days (5°C incubations) and checked on a Perkin Elmer Tri-Carb 4810 TR Scintillation Counter (Waltham, MA), measuring 14C counts for 1 min.
In the −5°C incubations, to determine whether CO2 was trapped in the ice and avoiding detection, after 483 days, we warmed these incubations, to 5°C and mixed them with 37% formaldehyde to prevent detection of new actively respiring microorganisms after thaw. Results were calculated by subtracting values from sterilized controls from the average radioactivity of replicates and expressed as the percentage of starting 1,2-14C acetic acid respired by the microbial community over time.
3. Results
3.1. Ice characteristics
In situ measurements showed that the temperature of all ice samples was always between 0.5°C and −0.5°C, while pH values ranged between 6 and 7.5 (Table 2). Values for TOC, nitrogen (NO3 − and NH3), and phosphorus (PO4 3−) varied greatly among sites and year (Table 2). Samples SC1 2017 and SC2 2017 had the highest levels of TOC (176.4 and 302.7 mg/L, respectively), whereas the samples from Crystal Ice Cave (CI 2017) and Merrill Cave (MC 2018) had the lowest levels of TOC (30.6 and 8.0 mg/L, respectively). No related trends were observed between nutrient concentration and biomass.
Lava Tube Ice Biomass and Physicochemical Characteristics
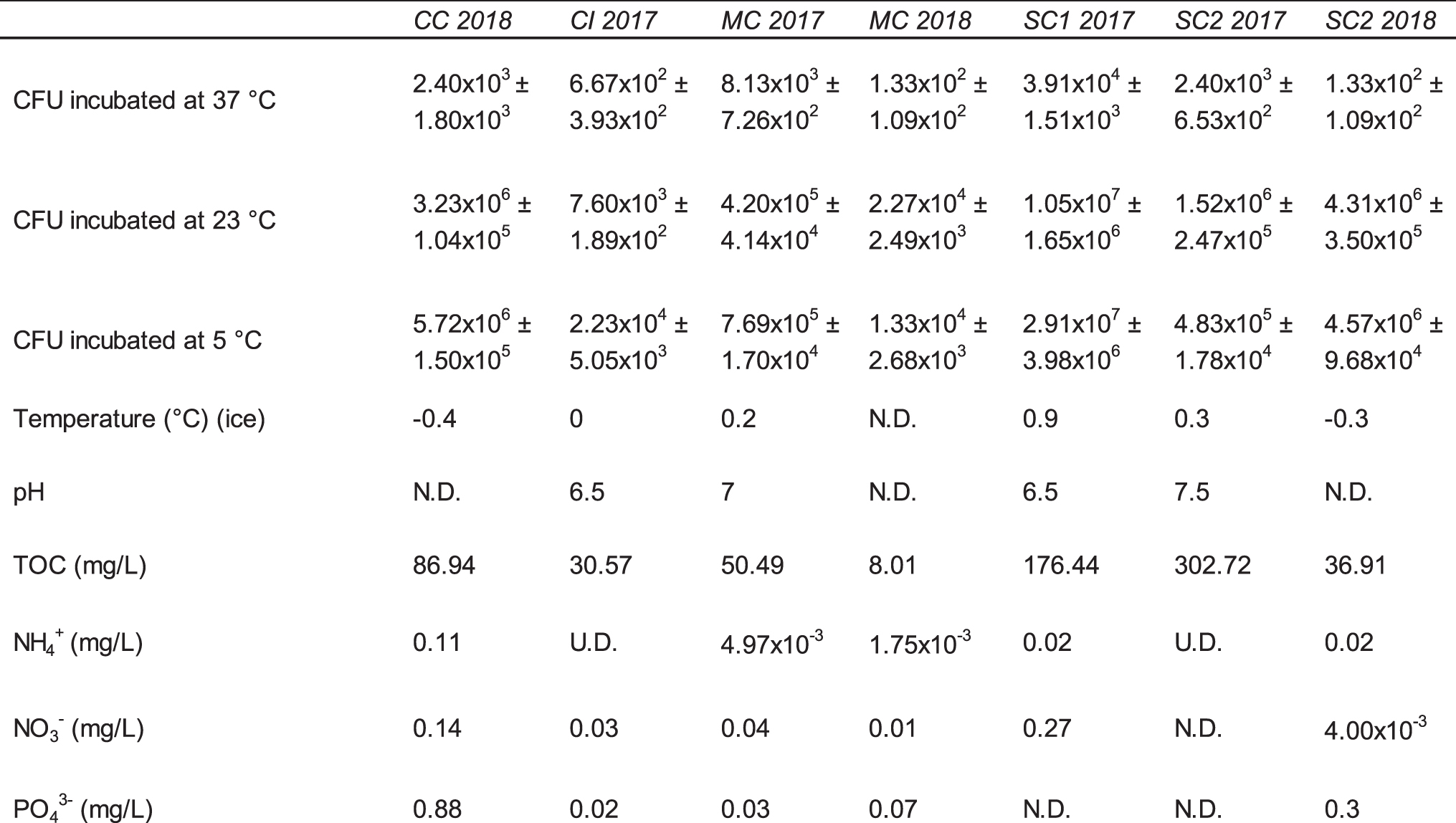
For CFU, standard error is included as a (±) value.
CFU = colony-forming units; N.D. = no data; TOC = total organic carbon; U.D. = undetectable.
3.2. Microbial cell enumeration, isolation, and characterization
The majority of CFUs incubated at 5°C, 23°C, and 37°C were observed on plates incubated at 5°C (23–74%). The total number of CFU determined at this temperature ranged from 1.3 × 104 CFU/mL in sample MC 2018 to 2.9 × 107 CFU/mL in sample SC1 2017 (Table 2). Based on morphology, 38 unique colonies obtained from the 2017 samples were isolated from plates incubated at 5°C and 23°C and chosen for further identification and characterization. Of the 38 isolates, 37 grew at 5°C and 24 grew at −5°C. Twenty-nine isolates grew in concentrations of 3% NaCl, while 13 isolates grew in concentrations of 9% NaCl and none grew in concentrations of 12–21% (Table 3).
Characterization of Lava Tube Ice Cultured Isolates
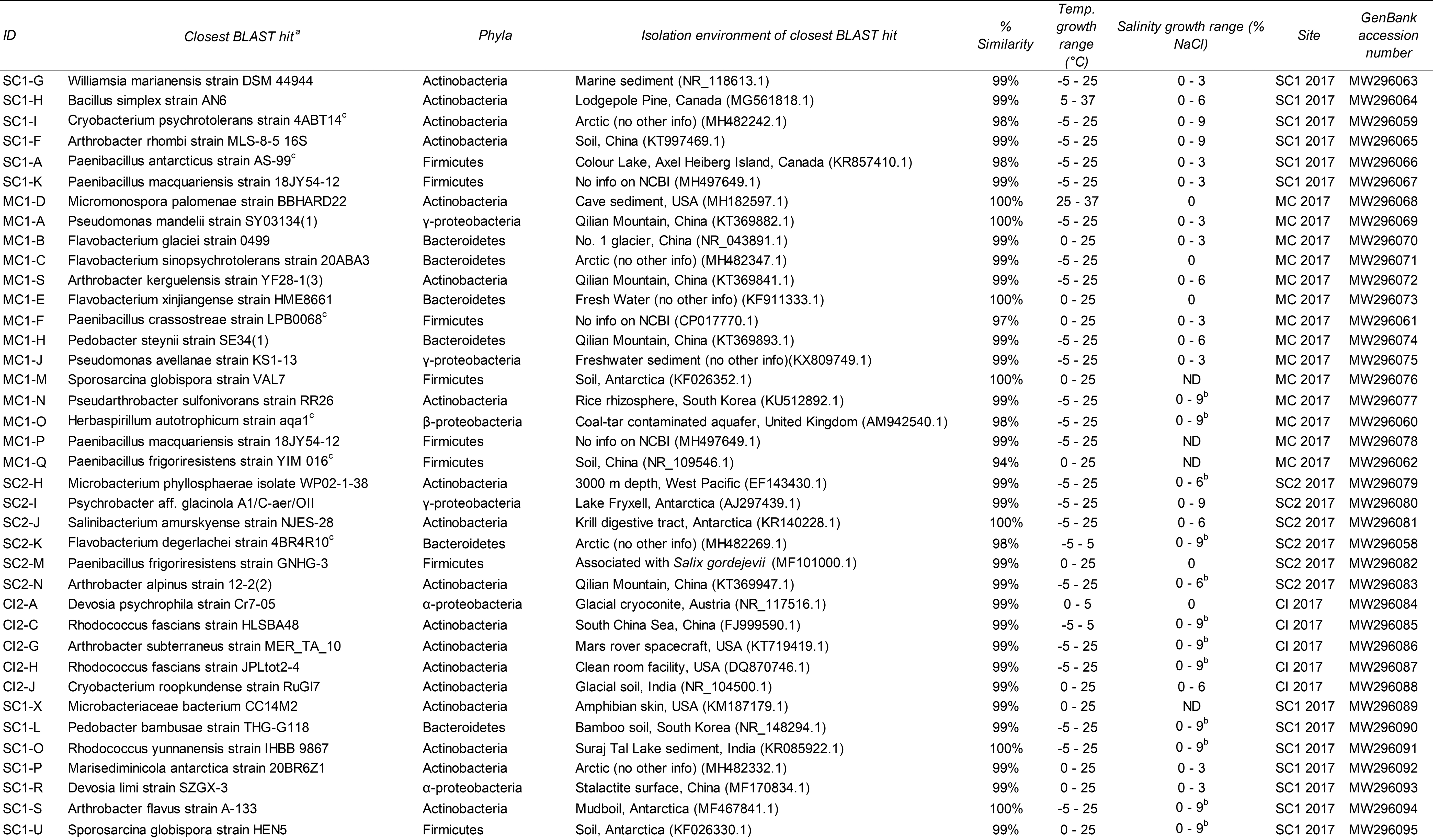
The closest BLAST hit to a cultured isolate was chosen for inclusion in the table.
Indicates poor growth.
Indicates a putatively novel species based on a 16S rRNA gene sequence similarity cutoff of ≥98%.
BLAST = Basic Local Alignment Search Tool; ND = not determined; Temp = temperature.
Sanger sequencing of the 16S rRNA gene of these isolates revealed that all isolates belonged to the phyla Actinobacteria (17), Firmicutes (9), Bacteroidetes (6), and Proteobacteria (6, specifically the Alpha, Beta, and Gamma classes) (Supplementary Fig. S2). According to the NCBI database, we determined at least 14 of our isolates shared closest sequence similarity with bacteria isolated from various cryophilic environments, including the Arctic, Antarctica, and various glaciers (Table 3). Only two isolates showed closest similarity to sequences obtained from a cave (Micromonospora palomenae strain BBHARD22 and Devosia limi strain SZGX-3). Additionally, another isolate isolated from Merrill Cave in 2017 may be a candidate novel genus since it showed a 94% sequence similarity to Paenibacillus frigoriresistens strain YIM 016 as the highest hit.
3.3. Prokaryotic community profiling
Illumina amplicon sequencing of the 16S rRNA V4–V5 gene region of microbial communities from LABE ice samples generated, after processing, 838,591 reads grouped into 11,475 unique ASVs. Generally, the majority of ASVs were distributed in two or more sites (Supplementary Fig. S3), except ASVs from MC 2017 and MC 2018 where most of them (6528 or 85%) were unique to that cave (Supplementary Figs. S3 and S4).
The community from all samples was composed almost exclusively of bacteria (>99.97% of total sequences), with very few archaeal sequences (<0.02%) detected. The dominant bacterial phyla included the Actinobacteria (30% of total sequences), Proteobacteria (30%), Bacteroidetes (19%), Firmicutes (5%), and Chloroflexi (5%) (Fig. 2). The most abundant genera within LABE ice were Flavobacterium (7% of total sequences) (Bacteroidetes), Arthrobacter (4%) (Actinobacteria), Sphingomonas (4%) (Proteobacteria), Nocardioides (3%) (Actinobacteria), and Pseudomonas (3%) (Proteobacteria), but with a high abundance of unclassified sequences at this taxonomic rank (25%). Of the few archaeal sequences recovered, most belonged to either the phyla Euryarchaeota or Thaumarchaeota.
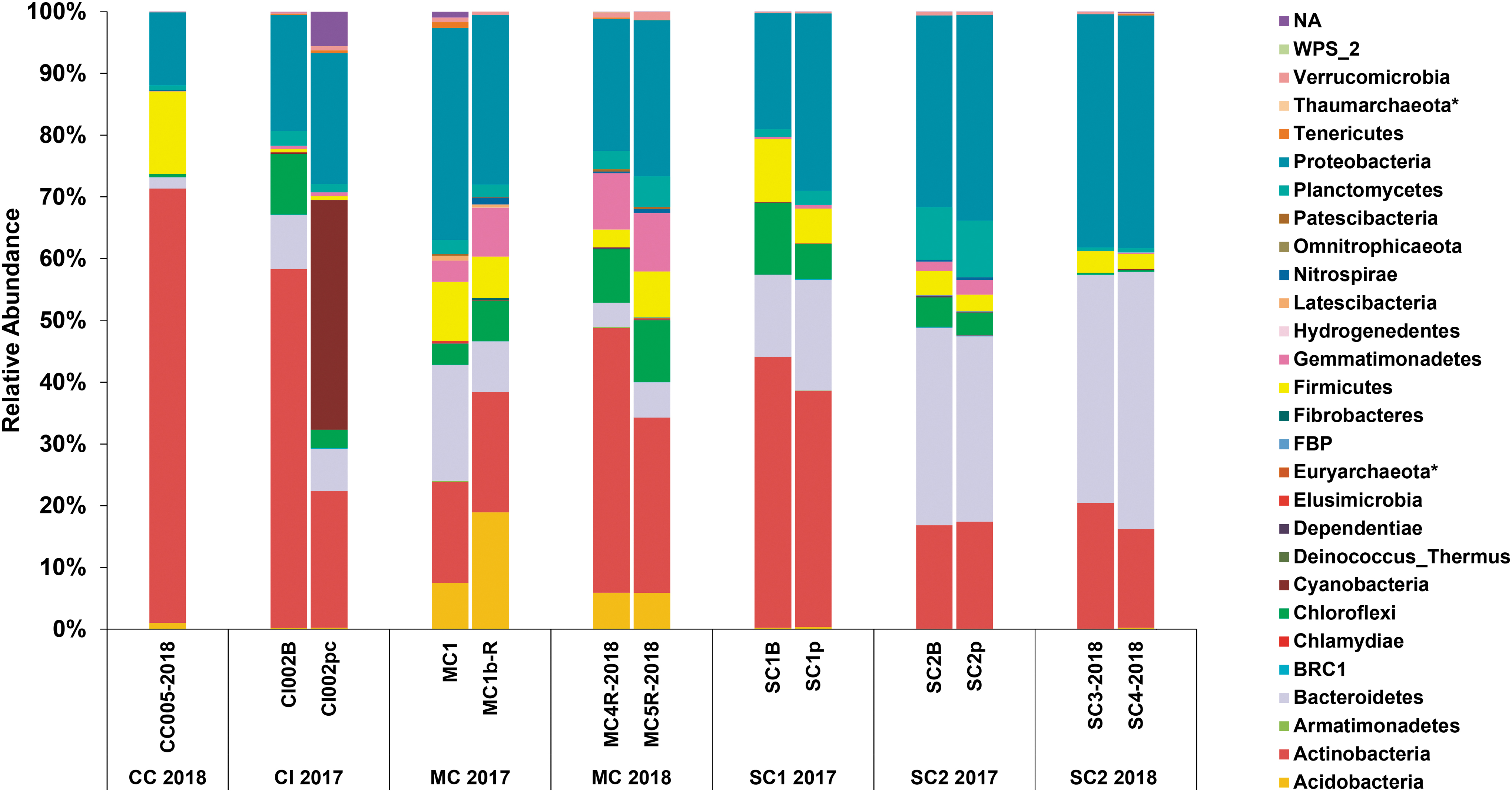
Bacterial and archaeal 16S rRNA community profile of LABE ice. Relative abundance is calculated from sequence read counts. Archaeal phyla are denoted in the legend by an asterisk (*). LABE, Lava Beds National Monument.
Of the 20 most abundant ASVs recovered from our sites, 16 shared closest blast similarity to other cryophilic environment-derived sequences and none with cave- or subterranean-derived sequences (Table 4). Alpha diversity measures (Shannon and Faith's PD) revealed sample MC 2017 to be the most diverse, whereas beta diversity measures (Bray–Curtis and UniFrac distances) revealed samples CC 2018 and CI 2017 to be the most dissimilar among all samples (Supplementary Fig. S5).
Most Abundant Amplicon Sequence Variants Within Lava Tube Ice
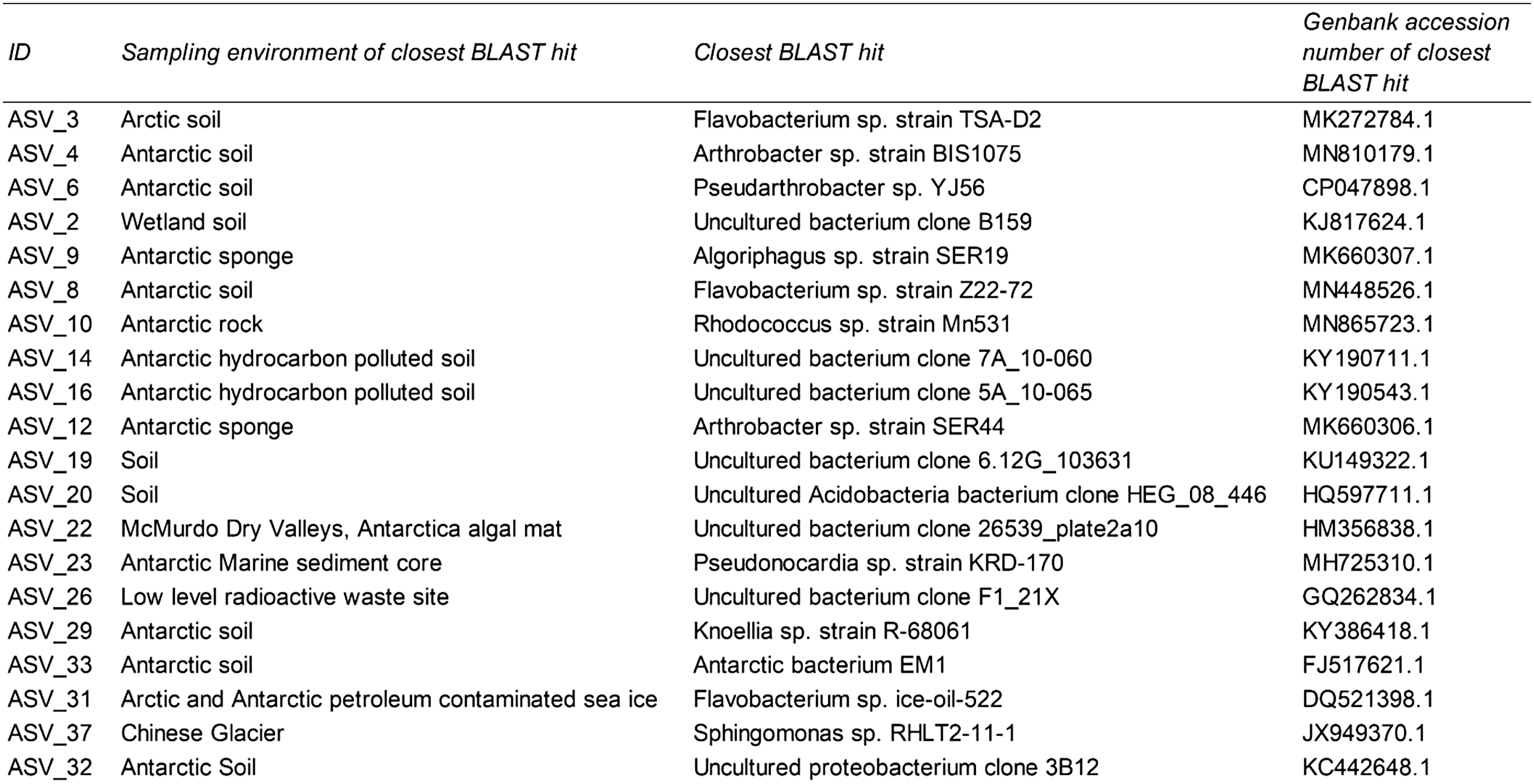
The 20 most abundant ASVs within our data set were BLAST against the NCBI nucleotide database. ID refers to the identifier of the ASV in our data set.
ASV = amplicon sequencing variant; BLAST = Basic Local Alignment Search Tool.
Piphillin predicted the LABE ice microbial communities to contain complete or almost complete heterotrophic and lithoautotrophic pathways. Lithoautotrophic pathways include C1 compound metabolism (methanotrophy and methanogenesis), sulfur cycling (thiosulfate oxidation and sulfate reduction), nitrogen cycling (nitrogen fixation, nitrification, complete nitrification, denitrification, and dissimilatory nitrate reduction), as well as others related to phototrophy, and manganese oxidation (Supplementary Fig. S6 and Supplementary Table S2).
3.4. Community-level physiological profiling of LABE ice microorganisms
A plateau in the AWCD was observed after 53 and 102 days for 5°C and −5°C incubations, respectively. Biolog EcoPlate data indicate higher overall substrate utilization and functional diversity in plates incubated at 5°C when compared with plates incubated at −5°C (Supplementary Table S2). Microbial communities from site CC 2018 showed the highest substrate utilization, being able to metabolize an average of 26 ± 2.20 and 23.33 ± 2.43 of the 31 substrates at 5°C and −5°C, respectively. This sample also showed the highest functional diversity (H′-clpp), with average values of 3.18 ± 0.07 at 5°C and 3.14 ± 0.05 at −5°C (Supplementary Table S1).
The only sample in which both substrate utilization and functional diversity were higher at −5°C was in CI 2017. This same microbial community utilized Tween 80, glycogen, 2-hydroxybenzoic acid, and glycyl-L-glutamic acid only at −5°C and D-mannitol, N-acetyl-D-glucosamine, and L-arginine substrates to a much higher degree at −5°C than 5°C (Fig. 3). No microbial community showed the capacity to metabolize glucose-1-phosphate, D,L-α-glycerol phosphate, or α-ketobutyric acid either at −5°C or at 5°C. In addition, no microbial community was capable of metabolizing itaconic acid or D-malic acid at −5°C (Fig. 3).

Heat map depicting EcoPlate substrate utilization by LABE ice microbial communities incubated at −5°C and 5°C. The average absorbance of three triplicate wells with blanks subtracted was used to calculate the data. Data used to calculate the average absorbance were collected once the AWCD plateaued, corresponding to 53 days for plates incubated at 5°C and 102 days for plates incubated at −5°C. AWCD, average well-color development.
NMDS ordination of the Biolog EcoPlate results (Supplementary Fig. S7) showed the location of the samples in the ordination mainly depended on the cave they came from (stress = 0.046). Incubations from MC 2017, MC 2018 (except for incubation MC 2018 + 5°C), and CI 2017 were distantly located from the main group, showing a higher influence of ice temperature on them than on the rest of the samples.
3.5. Heterotrophic microbial activity measured by a radiorespiration assay
In vitro heterotrophic microbial activity was detected above control levels in all samples by day 77 of the incubation at 5°C (Fig. 4). By day 447, between 6% (MC 2018) and 12% (SC1 2017) of initial acetate was mineralized. Activity was much lower in samples incubated at −5°C (Fig. 4). In this case, microbial radiorespiration was only detected above sterile control levels from samples CC 2018 and SC2 2018. By day 483, CC 2018 and SC2 2018 communities had mineralized 1% and 0.5%, respectively, of the initial acetate, compared with control values of 0.6% and 0.3%, respectively. In both samples, control and experimental assays were significantly different until day 447 for SC2 2018 when error bars began to overlap.

Microbial activity measured at −5°C and 5°C by the detection of biomineralized 14C acetate. Data are displayed as the percent of 14C respired as CO2 of the initially provided 14C. The assay was stopped after 447 days for 5°C incubations and after 483 days for −5°C incubations. Dotted lines refer to control samples and thus abiotic release of CO2. Error bars are calculated as the standard error based on three replicates. Note the differences in scale of the vertical axis.
4. Discussion
4.1. LABE ice harbors microorganisms adapted to cold temperatures
Several lines of evidence suggest that LABE ice microbial communities are primarily cold adapted. The strongest evidence being that the greatest number of CFUs we recovered was from plates incubated at 5°C as opposed to 23°C or 37°C. Of all the CFU we isolated for further characterization, almost all grew at 0°C or −5°C. Using a single media may only offer a narrow view of the culturable microbial community. However, the R2A media employed in this study has previously resulted in the highest viable plate counts from other cryoenvironments (Christner et al., 2000; Miteva et al., 2004; Hansen et al., 2007; Steven et al., 2007a; Bottos et al., 2008) and, therefore, is a good indicator of the viable lava tube ice microbial community.
Among both cultured isolates and the 20 most abundant ASVs recovered from our samples, most shared closest sequence similarity with sequences recovered from other cryoenvironments such as from the Arctic, Antarctic, and mid-latitude glaciers. Extensive study of microbial communities inhabiting Scarisoara Ice Cave, a karstic cave, has also suggested a predominantly cold-adapted microbial community (Hillebrand-Voiculescu et al., 2015; Iţcuş et al., 2016; Brad et al., 2018; Itcus et al., 2018; Paun et al., 2019).
Our results add to the evidence that cave ice deposits may constitute reservoirs of novel cold-adapted microorganisms. Based on a full-length 16S rRNA gene sequence similarity cutoff of <98.7% as proposed in the work of Kim et al. (2014), we determined six of our cultured isolates to be putatively novel species, all of which grew at −5°C and/or 0°C. At the rank of family and genera, 10% and 25%, respectively, of our ASVs were unassigned. Similarly, study of Scarisoara cave has revealed a large proportion of unassigned and putatively novel operational taxonomic units among bacteria, archaea, and fungi (Brad et al., 2018; Itcus et al., 2018). Therefore, future study of these novel microorganisms may help us understand cryophilic biology and evolutionary adaptations, which may allow life to persist on other planets.
Actinobacteria, Proteobacteria, Bacteroidetes, Firmicutes, and Chloroflexi revealed by 16S rRNA gene sequencing to be dominant at LABE were also detected in other cave ice environments including Scarisoara Cave (Itcus et al., 2018; Paun et al., 2019) and Hawaiian lava tubes on the side of the Mauna Loa Volcano (Teehera et al., 2017). Proteobacteria and Chloroflexi also dominated sediment from three volcanic ice caves from Mount Erebus in Antarctica, with a lesser presence of Actinobacteria and Firmicutes (Tebo et al., 2015). Together, these studies suggest that, at higher taxonomic classifications, microbial communities remain generally consistent among ice containing and subzero cave habitats.
Although phyla recovered from our samples such as Actinobacteria and Proteobacteria are ubiquitous across many environments, the appearance of Firmicutes in LABE ice may be due in part to the ability of members of this phylum to persist in cryophilic environments (Shivaji et al., 2014). The family Planococcaceae, comprising the largest proportion of Firmicutes from our sites, has routinely been isolated from cryophilic environments around the world (Shivaji et al., 2014). In fact, Planococcus halocryophilus, isolated from Arctic permafrost (Mykytczuk et al., 2012), can grow at temperatures of −15°C and metabolize to temperatures as low as −25°C (Mykytczuk et al., 2013).
Many genera of the Planococcaceae can also form endospores (Shivaji et al., 2014), protective structures that allow the bacterium to enter dormancy, protecting it from nutrient limitation, extreme temperatures, UV radiation, and desiccation. Within our own data set, we found sequences belonging to numerous Firmicutes endospore formers such as Oxobacter, Anoxybacillus, Syntrophomonas, Anaerobacillus, Coxiella, Bacillus, Paenibacillus, Paenisporosarcina, and Sporosarcina.
Of note, we found abundant sequences from our sites showing high sequence similarity to endospore formers, Paenisporosarcina macmurdoensis and Bacillus subtilis; the first is a psychrophilic bacterium originally isolated from the McMurdo Dry Valleys in Antarctica and the second is a model spore former. Bacillus subtilis spores have even survived years in the harsh conditions of space (Horneck et al., 1994), leading to the suggestion that should endospore formers have evolved when Mars' climate was milder then they may still persist on the planet today (Yung et al., 2007).
Given the average surface temperatures of present-day Mars are well below 0°C and recent evidence suggests that early climatic conditions on Mars may have been predominantly cold with surficial water stored as snow and ice, Grau Galofre et al. (2020) suggests that extinct or extant martian life would need to be adapted to cold temperatures. This theory also applies to microorganisms inhabiting martian lava tubes given modeling of atmospheric temperatures within this environment are also well below 0°C (Williams et al., 2010).
Our results depict a community composed of psychrophilic, psychrotrophic, and endospore-forming microorganisms more closely related to communities inhabiting other polar cryoenvironments, as opposed to those communities inhabiting temperate cave and subsurface environments. Such an insight points to the strong selective pressure, which ice and cold temperatures exert on microbial communities.
4.2. Lava Beds ice supports active microbial communities
We believe this to be the first study to directly measure metabolic activity of lava tube ice-associated microorganisms and the first time Biolog EcoPlates have been used to measure subzero microbial metabolic activity. Our results imply that the heterotrophic microbial communities of LABE ice are just as capable of metabolic activity at subzero temperatures as at above zero temperatures. In every sample, tested substrates were metabolized at −5°C and, in the majority of samples, there were no significant differences in substrate richness or diversity between the two incubation temperatures (Supplementary Table S2).
Although microbial communities inhabiting cryophilic environments are assumed to be adapted to life at cold temperatures, the influence of incubation temperature on carbon substrate utilization is still poorly understood. Reports from cryophilic environments such as ice containing caves and Antarctic soil have shown substrate utilization richness and functional diversity increases with decreasing temperature (Pessi et al., 2012; Iţcuş et al., 2016); however, the opposite result was reported from Arctic permafrost (Ernakovich and Wallenstein, 2015). Our data do not fully support either result but do extend the range these results can be applied, to including subzero temperatures.
Exploratory ordination analysis (NMDS, Supplementary Fig. S6) of our sites suggests that higher organic substrate abundance and diversity promotes the development of more diverse microbial communities capable of metabolizing those substrates, no matter the temperature of the site. Therefore, the cave sites which contained higher concentrations of organics also contain more functionally diverse microbial communities, similar to what has been found for other similar settings as Scarisoara Cave (Iţcuş et al., 2016). Such results suggest that at both above and subzero temperatures, organic carbon content, and presumably substrate richness, may predict heterotrophic functional diversity.
Among sites MC 2017, MC 2018, and CI 2017, many substrates were found to be metabolized preferentially at −5°C but not 5°C (Fig. 3), thus hinting at potential strategies used by active microbial communities to thrive in lava tube ice. Such substrates that may play a role in cold adaptation include D-mannitol, γ-aminobutyric acid, Tween 40, and Tween 80. D-mannitol is a well-known compatible solute, capable of providing cryopreservation, and osmoprotection to psychrophiles in addition to carbon, nitrogen, and energy (Weinstein et al., 1997; Tribelli and López 2018). γ-Aminobutyric acid is a compatible solute produced by plants to withstand various stresses such as cold temperatures (Shelp et al., 1999; Mazzucotelli et al., 2006), and given our results, may do the same in bacteria.
Extensive metabolism of Tween 40 and Tween 80 has been reported by cryophilic microbial communities in environments such as Antarctic and Arctic seawater (Sala et al., 2005, 2008), as well as in Antarctic soils (Pessi et al., 2012). It has been proposed that polyols such as Tweens are metabolized by cold-adapted microbial communities to grow at cold temperatures and protect themselves from freeze-thaw events (Wynn-Williams, 1990; Robinson, 2001). Our results support this hypothesis. Tween 80 may specifically play a role in microbial cold tolerance by being incorporated into cell membranes and repressing fatty acid synthesis, thus preserving membrane fluidity (Reitermayer et al., 2018) at low temperatures.
It is surprising that microbial activity was not detected more readily in −5°C radiorespiration assays, given the sensitivity of the assay to detect respiration from cryophilic environments (Steven et al., 2007b), and that in situ microbial activity has been detected from ice and other cryophilic environments all over the world, including Arctic permafrost (Rivkina et al., 2000; Jakosky et al., 2003; Steven et al., 2008), sea ice (Junge et al., 2004), Antarctic snow (Carpenter et al., 2000), and Antarctic water tracks (Chan-Yam et al., 2019). Although radiolabeled carbon may have been trapped in the ice, analysis after thawing still did not show any significant increase in respired carbon, so perhaps our samples required a longer incubation period or a larger nutrient spike to detect greater activity.
This hypothesis could explain the observed disparity in metabolic activity between radiorespiration and EcoPlate assays. For example, substrates contained within EcoPlate wells are ∼100 times more concentrated than within our radiorespiration assays. While EcoPlates can indicate the potential of a microbial community to metabolize substrates at in situ temperatures, radiorespiration assays better mimic the actual in situ environment (Steven et al., 2007b). However, radiorespiration assays do not perfectly mimic the in situ environment nor do any other laboratory-based metabolic activity assays. Therefore, we suggest further investigation, employing techniques such as in situ stable isotope probing or in situ gas flux analyses should be explored to confirm our laboratory-based assays that demonstrated active microbial metabolic activity.
Despite the uncertainty in the extent that LABE ice microbial communities are metabolically active in situ, our results do point to microbial communities from this environment capable of metabolic activity at subzero temperatures. The continued investigation of these communities may increase our understanding of microbial adaptations to living in subzero temperatures. Moreover, such a finding raises the possibility that martian lava tubes may be a habitable environment for extant microbial life.
4.3. Potential metabolisms of Lava Beds ice microbial communities
Our sites are not considered oligotrophic, since oligotrophic caves should contain <2 mg/L of TOC (Tomczyk-Żak and Zielenkiewicz, 2016) and, although many caves meet this definition, lava tubes may be an exception (Northup and Lavoie, 2015). Given their proximity to the surface and the propensity of the host rock to fracture, organic carbon can readily enter into lava tubes by transport of meteoric water, plant roots and soil from the surface through cracks, and fissures in the walls and ceiling. Deposition of guano and other detritus by seasonal bat and rat populations can also occur (Northup et al., 2011).
We suspect most of the organic carbon from LABE caves originated from an allochthonous origin, having observed plant roots, animal deposition, and human alterations near our sampling sites. Such sources help explain the abundance of heterotrophic microorganisms found within our sites.
TOC concentrations measured in the martian surface are quite low (although likely higher in the subsurface) (Freissinet et al., 2015; Eigenbrode et al., 2018), suggesting that any heterotrophic microorganism in existence today within martian lava tubes would likely need to rely on primary production by chemolithoautotrophs, as has previously been observed in terrestrial cave habitats (Engel, 2012).
Complete or almost complete pathways for heterotrophic and lithoautotrophic metabolisms were predicted by Piphillin to exist within our samples. Furthermore, various Pseudomonas ASVs, all within the 100 most abundant ASVs within our data set, showed 99% 16S rRNA gene sequence similarity to a facultatively autotrophic iron oxidizing, Pseudomonas spp., recovered from the basalt/ice interface of a lava tube from the nearby Oregon Cascades (Popa et al., 2012).
Many Flavobacterium and Arthrobacter ASVs also contained within the 100 most abundant ASVs showed a 16S rRNA gene sequence similarity of ≥98% to manganese oxidizing bacteria isolated from caves of the upper Tennessee River basin (Carmichael et al., 2013). We suggest that these taxa may derive energy from reduced iron and manganese present within the basaltic host rock of LABE caves. Thus, these taxa may contribute to cave mineral alteration and dissolution (Northup and Lavoie, 2001).
Lithoautotrophic microorganisms, such as those recovered from our sites, likely persist in lava tubes owing to a variety of energy sources produced by water–rock interactions and radiolysis (Hays et al., 2017). These interactions may support, among others, iron oxidation, sulfate reduction, hydrogen oxidation, and methanogenesis (Stevens and McKinley, 2000; Bach and Edwards, 2003; Lin et al., 2006; Lever et al., 2013; Osburn et al., 2014; Simkus et al., 2016). Considering much of the martian surface is composed of basalt and ultramafic rock (Boynton et al., 2008; McSween et al., 2009), similar interactions also likely occur there.
Molecules such as NO3 − and SO4 2−, both observed on Mars (Stern et al., 2015; Rapin et al., 2019), may serve as electron acceptors in the absence of oxygen (Hays et al., 2017). Thus, the variety of both inorganic electron donors and acceptors potentially contained within martian lava tubes suggests primary production by lithoautotrophs, especially chemolithoautotrophs is favorable, should life have ever been present there in the first place.
5. Conclusions
Our results demonstrate that microbial communities inhabiting lava tube ice from LABE, analogues to martian lava tubes, are cold adapted and capable of metabolic activity at subzero temperatures. Furthermore, these communities resemble cryophilic microbial communities more than cave microbial communities. Lithoautotrophic metabolisms predicted to exist within our sites such as methanogenesis, sulfate reduction, nitrate reduction, and iron oxidation are believed to be plausible metabolisms for survival on Mars.
Our results raise the prospect that martian lava tubes may be a habitable environment and a refuge to martian microorganisms, as the planet's surface transitioned to the inhospitable environment observed today. In this regard, lava tube ice would likely provide relatively stable temperatures, protection from radiation, water from ice, and energy from basaltic minerals, of which lava tubes are composed. Our findings serve as a basis for further study of this unique environment.
Footnotes
Acknowledgments
We thank the LABE park staff and rangers for their tremendous support during fieldwork. All fieldwork was conducted under National Park Service research permit no. LABE-2016-SCI-0011. We thank Jenny Ni (McGill University) and Christopher Patterson (McGill University) for their help with sample collection while in the field and Hélène Lalande (McGill University) and Hisham Benslim (McGill University) for chemical analysis.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was financially supported by the Canadian Space Agency's Flights and Fieldwork for the Advancement of Science and Technology (FAST) program, the Natural Sciences and Engineering Research Council of Canada's (NSERC) Discovery program and Discovery Accelerator program, the Canada Research Chair (CRC) program, and the McGill Space Institute.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Table S1
Supplementary Table S2
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.