
Abstract
The degradation of glycine (Gly), proline (Pro), and tryptophan (Trp) was studied under simulated Mars conditions during UV-driven production of oxychlorines and compared under Mars ambient and humid conditions, as films, and with addition of sodium chloride (NaCl), sodium chlorate (NaClO3), and sodium perchlorate (NaClO4) salts. It was shown that glycine sustained no significant destruction in either of the non-salt samples under Mars ambient or humid conditions. However, its degradation increased in the presence of any of the three salts and under both conditions though more under humid conditions. Proline degradation followed the order No Salt > NaCl > NaClO3 > NaClO4 under Mars ambient conditions but the reverse order under Mars humid conditions. A mechanism is proposed to explain how water and silica participate in these degradation reactions and how it is strongly influenced by the identity of the salt and its ability to promote deliquescence. No difference was observed for tryptophan between Mars ambient and humid conditions, or for the different salts, suggesting its degradation mechanism is different compared to glycine and proline. The results reported here will help to better understand the survival of amino acids in the presence of oxychlorines and UV on Mars and thus provide new insights for the detection of organic compounds on future Mars missions.
1. Introduction
Extraterrestrial organic compounds have been detected in comets (Kissel and Krueger, 1987), chondritic meteorites (Hahn et al., 1988; Sephton et al., 1998; Ehrenfreund et al., 2000), and interplanetary dust particles (Flynn, 1996). It has been estimated that long-term bombardment of Mars by such objects has delivered up to ∼104 g/m 2 yr of material to its surface (Flynn and McKay, 1990) including ∼108 g/yr of reduced carbon (Benner et al., 2000); thus organic compounds should be abundant on its surface (Bland and Smith, 2000; Tomkins et al., 2019). However, no definitive evidence of organic compounds was found by the 1976 Viking landers, which included a gas chromatograph–mass spectrometer capable of detecting them (Biemann et al., 1977). It was suggested that this might be due to such reasons as strong oxidants, metal oxides, oxygen radicals, degradation by UV or other radiation, or that the Viking gas chromatograph–mass spectrometers failed to detect them (Zent and McKay, 1994; Stoker et al., 1997; Yen et al., 2000; ten Kate, 2010). The situation drastically changed when the Sample Analysis at Mars (SAM) instrument suite on board the Mars Science Laboratory first detected 150–300 ppb of indigenous chlorobenzene, with lesser amounts of C2–C4 dichloroalkanes (Freissinet et al., 2015), and soon afterward a variety of pyrolysis products, including thiophenic, aromatic, and aliphatic compounds (Eigenbrode et al. 2018).
In 2008, the first wet chemical analysis of martian soil by the Wet Chemistry Laboratory on board the Phoenix Mars lander (Kounaves et al., 2009) revealed the presence of ∼0.6 wt % perchlorate (ClO4 -) (Hecht et al., 2009; Kounaves et al., 2010, 2014a). The widespread occurrence of ClO4 - and chlorate (ClO3 -) on Mars has subsequently been confirmed by their detection in the martian meteorites EETA79001 and Tissint (Kounaves et al., 2014b, Jaramillo et al., 2019) and by the SAM instrument on Curiosity at Gale crater (Glavin et al., 2013; Leshin et al., 2013), and by detection of chlorinated hydrocarbons (Ming et al., 2014; Freissinet et al., 2015; Eigenbrode et al., 2018).
Degradation of organic molecules on Mars' surface is likely the result of a combination of processes. Radiation in various modes, directly or indirectly, is likely the dominant factor in destroying organic molecules. Even though much higher levels of solar energetic particles and galactic cosmic rays can reach Mars' surface (Kuhn and Atreya, 1979; Cheptsov et al., 2018), their total energy flux is ∼104 times less than the solar UV flux (Hassler et al., 2014). Although the UV at the martian surface penetrates only millimeters into the regolith (Moores et al., 2007; Schuerger et al., 2012), its destructive power is likely felt more widely due to such processes as cryoturbation (Levy et al., 2010), aeolian transport, and impact gardening (Hartmann et al., 2001), which allow regolith to be circulated and exposed to it. However, organic compounds or evidence of life could still be present where it has been sheltered from exposure to radiation (Pavlov et al., 2012; Fornaro et al., 2018; Stalport et al., 2019; Tan and Sephton, 2019).
It has been demonstrated that degradation of amino acids by UV radiation can occur rapidly at timescales of days or weeks but can extend to millions of years. It can also vary drastically depending on a variety of factors such as the chemical environment, reaction pathways, degree of protection, molecular weight/structure, and the UV wavelength (ten Kate et al., 2005, 2006; Bertrand et al., 2015; Laurent et al., 2019; Rowe et al., 2019; Stalport et al., 2019). For example, it has been shown that a thin film of glycine has a half-life of 22 ± 5 h when irradiated under UV between 120 and 180 nm at room temperature, but modelling and experiments have shown that its half-life can be extended on the order of 107 years when protected by inert martian regolith (ten Kate et al., 2005). When absorbed on minerals such as forsterite, antigorite, spinel, or pyrite, the half-life of glycine degradation was shown to be between 0.5 and 2 h. This relatively rapid degradation of glycine indicates that these minerals on Mars may be involved in photo-Fenton catalytic reactions (Fornaro et al., 2018; Poggiali et al., 2020). The destructive effect of UV radiation seems to involve more than just the photon energy of the different wavelengths. Using tryptophan and tryptophan analogs, investigators have shown that UVB (280–315 nm) has greater destructive power than the higher-energy UVC (100–280 nm) (Rowe et al., 2019).
Under Mars ambient conditions both ClO4 - and ClO3 - can be photochemically produced on Cl-bearing mineral surfaces, most likely due to silicate (SiO2) and/or metal-oxides acting as photocatalysts to generate radicals such as O2 -, which can then react with chloride (Yen et al., 2000; Carrier and Kounaves, 2015). During this process, several oxychlorine intermediates such as hypochlorite (ClO-), chlorite (ClO2 -), chlorate (ClO3 -), and chlorine dioxide (ClO2) gas, as well as radicals such as •OCl, •Cl, O2 -, and •OH are also likely generated (Catling et al., 2010). Recently, it was confirmed that the photochemical perchlorate formation pathway can exist without ozone (O3) and that a highly oxidizing oxychlorine gaseous intermediate, most likely ClO2 gas, was present in this pathway (Jain et al., 2017; Liu and Kounaves, 2019).
Here, we compared the degradation of three amino acids—glycine (Gly), proline (Pro), and tryptophan (Trp) (Fig. 1)—selected in order to investigate how structure and size influences their destruction by UV photons under Mars conditions. Both glycine and proline have been found in meteorites (Kvenvolden et al., 1971). Tryptophan was selected due to its aromatic ring structure. The overall aim of the reported effort was to investigate the UV degradation of glycine, proline, and tryptophan in the presence of NaCl, NaClO2, NaClO3, and NaClO4 under Mars conditions.
Structures of glycine (Gly), proline (Pro), and tryptophan (Trp).
Based on the data obtained in this study, we propose different degradation mechanisms for glycine, proline, and tryptophan (under simulated Mars conditions) that are related to their chemical structures as well as the role of oxychlorine salts in these reactions. These results will help us to better understand the survival of amino acids in the presence of oxychlorines and UV light on Mars and provide important insights for the detection of organic compounds on future Mars missions.
2. Materials and Methods
2.1. Reagents
The salts used were sodium chlorite (NaClO2, Sigma-Aldrich technical grade); sodium chlorate (NaClO3, ACS reagent, ≥99.0%); sodium perchlorate (NaClO4, ACS reagent, ≥99.0%); chlorite ion (ClO2 -, 1000 ppm IC standard, >99.9%); and chlorate ion (ClO3 -, 1000 ppm IC standard, >99%). Silica sand (SiO2, 50–70 mesh calcined quartz, with ≤0.05% HCl-soluble substances, Sigma-Aldrich 1075360250) was used as the supporting matrix. The amino acids glycine, DL-proline, and L-tryptophan were purchased from Sigma-Aldrich (ReagentPlus, ≥99% HPLC grade). A 1000 ppm amino acid stock was prepared in Nanopure 18.2 MΩ cm deionized water and kept at ∼4°C until use. The amino acid solutions were sterilized by filtration with a 0.2 μm syringe filter (Thermo Scientific 7239920). It has been shown that there is no difference between the D and L forms of amino acids with respect to UV photolysis (Bertrand et al., 2015); thus the chirality of the amino acids is not of any consequence in these studies.
2.2. Mars ambient and humid simulation chamber
The Tufts Mars Simulation Chamber (MSC) has been previously described in detail (Liu and Kounaves, 2019). It consists of a stainless-steel cylindrical chamber (∼5 × 104 cm3) with a cold plate at the bottom maintained at -15°C and is continually monitored by a temperature data logger (EL-USB-2, Lascar, PA, USA). The simulated Mars ambient atmosphere is composed of 95.3% CO2, 2.8% N2, 1.8% Ar, 0.10% O2, <0.1% H2O (Airgas USA), and maintained at a pressure of 10 torr. The UV light is provided by a 450 W xenon lamp through a 22 cm fused silica port and monitored with a UV spectrophotometer probe (BLUE-Wave, StellarNet). Heating of the sample by the UV lamp was effectively eliminated by passing the UV light beam through an NIR water filter (Model 6123NS, Newport, USA). Since UV radiation on Mars of wavelengths <200 nm is attenuated by the CO2 atmosphere (Kuhn and Atreya, 1979), the experiments described here used UV at wavelengths >200 nm. Mars humid conditions were roughly controlled by placing or removing a 9 cm diameter Petri dish filled with 50 mL of deionized water ice on the cold plate ∼30 cm from the sample, creating water vapor with ∼1 torr partial pressure at -15°C.
2.3. Sample preparation
Amino acid films were prepared by adding 100 μL of 1000 ppm amino acid stock to custom-made glass plates (∼25 mm diameter) cut from the bottom ∼8 mm of 20 mL borosilicate glass scintillation vials (Fisher Scientific) and then oven-dried overnight at 60°C. All glass plates were cleaned before use with Piranha solution (2.5:1.0 v/v concentrated H2SO4:H2O2) to ensure a hydrophilic surface. The custom-made plates with the dried amino acid film were placed symmetrically under the UV light spot in the Mars chamber.
The sand + amino acid sample stocks were prepared by combining 10 g silica sand, 2.0 mL of 1000 ppm amino acid, and 5 mL water, then oven-dried overnight at 60°C to avoid degrading the amino acid. After drying, the stock was homogenized with a mortar and pestle, and its homogeneity was verified with electrospray ionization mass spectrometry (ESI-MS). For each experiment, four replicates (0.5 g each) from the stock were used as samples and four as controls. For the samples with salt, an extra 0.60 mL of 1 × 105 ppm NaCl, NaClO3, or NaClO4 was added to the sample stock before drying at 60°C overnight.
The amino acid degradation in each sample with the added salt was confirmed by comparing with room temperature controls that showed no significant degradation after drying. A set of recovery tests were conducted for each amino acid + salt combination, and no significant loss was observed during sample preparation. The amino acid to salt ratio was optimized based on the ESI-MS signal and the amount of amino acid degraded in order to ensure sufficient amino acid ionization in the ESI-MS while maintaining 10 times more oxychlorine salt than the molar amount of amino acid (Constantopoulos et al., 1999). The detection limit for each sample combination was determined to ensure it was <10% of the starting amino acid concentration. A 0.5 g sample was placed in each vial during the experiment with a resulting sample layer thickness of ∼1 mm. The samples were configured as previously described (Liu and Kounaves, 2019).
Sodium chlorite was not used for the amino acid + salt degradation test because it would cause amino acid degradation during sample preparation. This is due to the decomposition temperature of sodium chlorite decreasing as the moisture content increases, making the chlorite more reactive toward organic compounds during drying (White et al., 1942). This process was evidenced by the presence of a yellowish discoloration (see Supplementary Fig. S2).
2.4. Calibration and recovery test
Calibration curves for each amino acid and internal standard in the presence of each salt were obtained during method optimization by using ESI-MS. It was confirmed that the ratio of the amino acid to the internal standard was linear within the range of 10–200% of the starting amino acid concentration (10–200 ppm).
2.5. Electrospray ionization mass spectrometry analysis
Electrospray ionization mass spectrometry (ESI-MS), instead of gas chromatography, was used to determine the concentration of the amino acids, because oxychlorine salts will cause thermal decomposition of organic compounds at the high temperatures typically found in gas chromatography analyses. The amino acids were leached by adding 10 mL of an optimized solvent consisting of a 1:1 (v/v) mixture of 1% acetic acid : methanol, stirred for 1 h and then filtered through a 0.2 μm filter. The internal standards were added prior to leaching. The ESI-MS was cleaned prior to use by running a 1:1:1 v/v mixture of water/methanol/acetonitrile (cleaning solvent) and then individually methanol and water at 30 μL/min for 5 min. In addition, the ESI-MS was also rinsed with the cleaning solvent between samples to minimize interference.
Individual ESI-MS direct injection methods were developed for each type of sample (No Salt, NaCl, NaClO3, NaClO4) by Xcalibur 2.0 built-in calibration using proline m/z = 116 peak with flow rate at 10 μL/min. Internal standards were used for each amino acid to ensure signal reproducibility and to account for the adsorption of the amino acids on the silica sand surface during leaching (glycine-213C for glycine, alpha-methyl-L-proline for proline, and 5-methyl-DL-tryptophan for tryptophan). The internal standards for each amino acid were selected to be chemically similar to the target amino acid. All the amino acids and internal standards were calibrated and verified before use.
The peaks were selected based on reference data from MassBank 1 and NIST Chemistry WebBook 2 . The peak at m/z = 76 was monitored for glycine and at m/z = 78 for glycine-2-13C internal standard. The peak at m/z = 116 was monitored for proline and at m/z = 130 for alpha-methyl-L-proline internal standard. Peak m/z = 205 was monitored for tryptophan, and m/z = 219 was monitored for 5-methyl-DL-tryptophan internal standard. The presence of the salts in the sample solution led to lower target peak intensity due to lowered ionization efficiency of the target amino acid. This was especially the case in the glycine+NaCl samples. Thus, precise peak positions instead of nominal peak positions were used in these samples for glycine (m/z = 76.45) and glycine-2-13C internal standard (m/z = 78.45).
Different solvent additives such as ammonia and ammonium acetate were also evaluated due to concerns about UV activated oxychlorine salts causing additional amino degradation in acidic solutions during storage. However, the acetic acid additive yielded the best signal stability as well as peak intensity. Tests were also conducted for all sand + salt + amino acid combinations to demonstrate that maintaining the sample leachates at 7°C for up to 4 days would not cause noticeable degradation of the amino acids. All the samples were analyzed within 48 h after leaching.
3. Results
3.1. Amino acid films
Glycine, proline, and tryptophan in the form of dried films were directly exposed to UV irradiation (200–400 nm) for ∼24 h under both Mars ambient and humid conditions. For Mars ambient samples, ESI-MS analysis showed that 100% ± 2% of the glycine remained in respect to the control sample. In contrast only 77% ± 5% of the proline and 48% ± 7% of the tryptophan remained in comparison to the controls (Fig. 2), with the tryptophan film also showing a yellow discoloration after irradiation similar to that when reacted with NaClO2 with increased moisture. In comparison, under Mars humid conditions no degradation of glycine or proline was observed, with 100% ± 2% of both remaining, while the amount of tryptophan remaining increased to 70% ± 4% (Fig. 2).
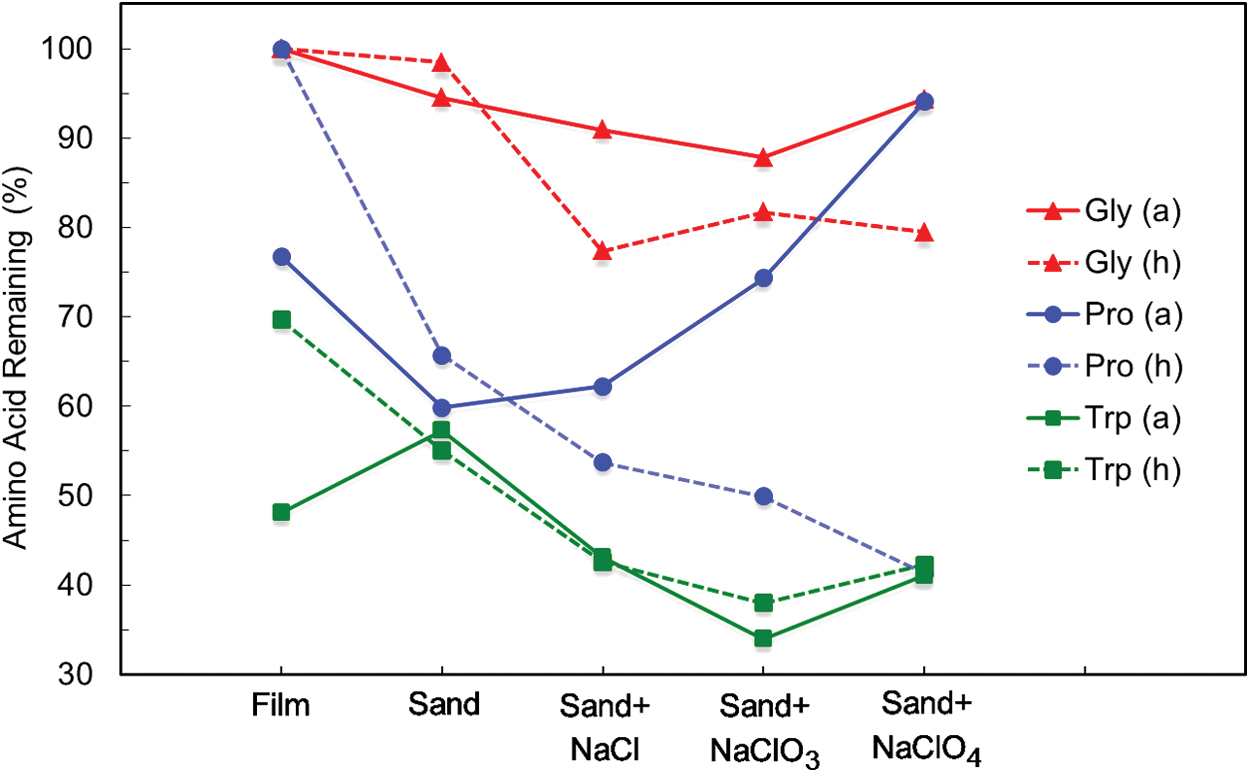
Amino acid degradation as a film and in the presence of NaCl, NaClO3, or NaClO4 under Mars ambient (a) or humid (h) conditions. The average shown for each sample group was normalized to 100% in respect to the control group. (For numerical data and error analyses, see Supplementary Table S1.) Color images are available online.
3.2. Amino acids combined with sand and sand+salt
Unlike the films, when the amino acids were mixed with silica sand and irradiated, degradation was observed at varying degrees in all the samples. As shown in Fig. 2, for the glycine+sand sample, 95% and 99% of the glycine still remained under Mars ambient and humid conditions, respectively. Under Mars ambient conditions the addition of NaCl, NaClO3, or NaClO4 to glycine+sand led to a small but significant change in glycine degradation, with ∼94–88% of the glycine remaining. However, under humid conditions the addition of a chlorine-bearing salt resulted in an additional ∼10% increase in degradation, with only 82–77% of glycine remaining.
The proline+sand samples under both ambient and humid conditions showed a ∼40% decline in proline. However, the addition of NaCl, NaClO3, or NaClO4 under Mars ambient conditions led to a drastic decrease in degradation from 62% to 74% to 94% of proline remaining, respectively. Yet under Mars humid conditions, the addition of NaCl, NaClO3, or NaClO4 had the opposite effect, accelerating its degradation process with only 54%, 50%, and 41% remaining, respectively.
Overall, tryptophan showed the greatest degradation of the three amino acids when irradiated. The tryptophan+sand samples showed a similar level of degradation as the tryptophan film (48–57%). Addition of NaCl, NaClO3, or NaClO4 under either Mars ambient or humid conditions resulted in ∼43%, ∼36%, and ∼41% tryptophan remaining, respectively.
4. Discussion
4.1. Amino acid films
Because the amino acid films were translucent and allowed some transmission of UV through the layer, their degradation did not strictly correlate with the total amount of UV absorbed. However, the molar absorptivity of each amino acid (Fig. 3), calculated by using Beer's law, was found to be consistent with the NIST standard spectra as well as the theoretical estimation based on functional group values (Tannenbaum et al., 1953).
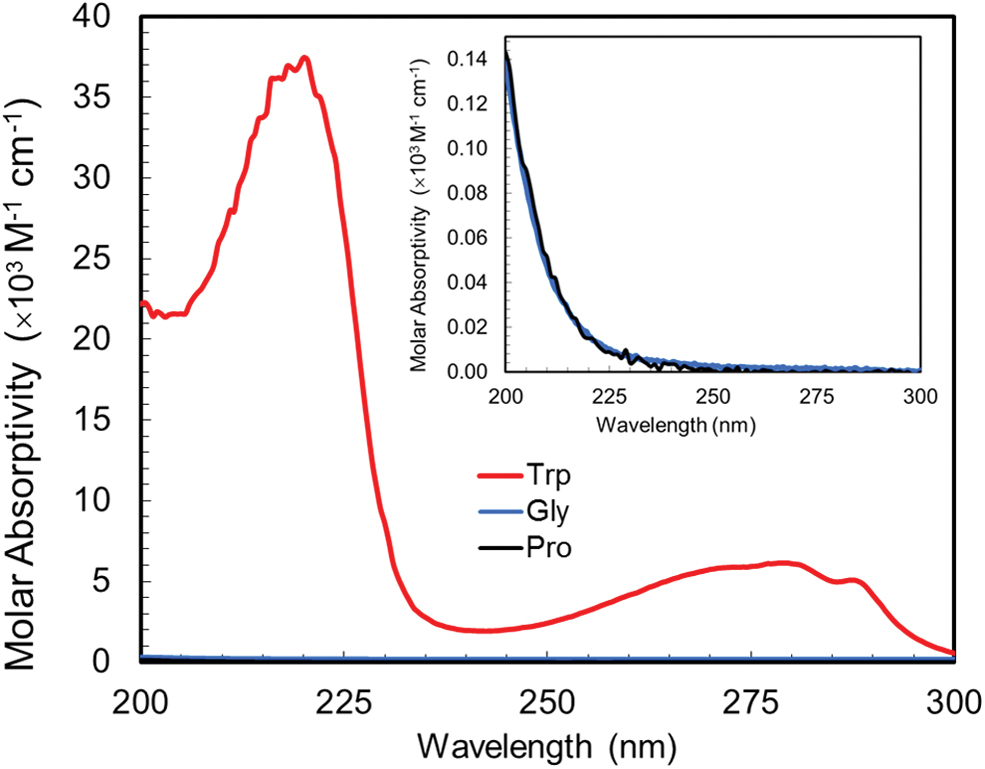
Molar absorptivity (M−1 cm−1) between 200 and 300 nm for 10 ppm tryptophan and 100 ppm of glycine and proline in water. Inset: Molar absorptivity axis × 300. Color images are available online.
The tryptophan film for both ambient and humid Mars conditions showed the most degradation and developed a visible yellowish discoloration. This is likely due to its higher and broader UV molar absorptivity in the 200–300 nm range compared to proline and glycine (Fig. 3). However, the discoloration of the tryptophan film also blocked the UV and thus afforded some protection from degradation. This was seen in preliminary tests where using a high-concentration tryptophan film resulted in little overall tryptophan degradation even though the film showed significant discoloration (see Supplementary Fig. S1).
Aromatic amino acids, such as tryptophan, in the free state are rapidly destroyed when irradiated with UV in the presence of oxygen (O2), but less so when O2 is absent (Vladimirov et al., 1970). The rapid degradation of tryptophan is due to the presence of the aromatic ring; thus UV irradiation below 275 nm (>4.5 eV) can drive it to the first excited singlet state. One possible mechanism for its photochemical degradation is the removal of one hydrogen from the nitrogen of the indole ring to produce a tryptophan radical (Vladimirov et al., 1970).
As can be seen in Fig. 3 (inset), the molar absorptivity of glycine and proline are almost identical in the 200–300 nm range. This is consistent with the absorption data for primary amines and pyrrolidine accounting for those in glycine and proline, respectively (Tannenbaum et al., 1953; Getoff and Schwörer, 1970). The increased degradation of proline under Mars ambient conditions cannot be explained solely by the UV absorptivity but is also likely caused by the presence of the pyrrolidine ring being more susceptible under UV radiation.
Aliphatic amino acid degradation can be induced by photo-excited molecules or by free radical precursors (Bonifačić et al., 1998). Glycine has only a weak UV absorption below 250 nm, which increases as the wavelength decreases. Decarboxylation free radical generation is the starting point for this reaction. For glycine, under γ radiation in basic aqueous solution, the •OH radicals created from H2O radiolysis induced a degradation reaction for glycine. Thus, the increase of humidity in this case could provide more radicals for degradation. These reactions generate CO2, CO, and HCN, which are some of the end products as the result of decarboxylation (Ehrenfreund et al., 2001; Bertrand et al., 2015). In our amino acid film tests, glycine films showed no significant degradation after 24 h under Mars ambient UV levels. This is consistent with the data from amino acid experiments on board the International Space Station (Bertrand et al., 2015).
It has been shown that glycine degradation under 200–300 nm UV irradiation depends on the concentration of O2. After 109 h of UV irradiation at room temperature, glycine was almost completely degraded in a pure 100 equivalents O2 atmosphere (stoichiometrically 100 times the amount of O2 needed to degrade all the glycine) yet showed no degradation in the absence of O2 (Oro and Holzer, 1979). Considering the concentration of O2 in the Mars gas (0.1% O2), this is consistent with our observation. Furthermore, it has also been shown that UV radiation >300 nm is not likely to contribute to amino acid degradation (Orzechowska et al., 2007).
Ultraviolet-visible absorption data for proline indicates that it also absorbs in the 200–250 nm range but with a high molar absorptivity that increases exponentially as the wavelength decreases. One key difference of proline compared to glycine and tryptophan is that proline does not have a -NH2 terminal but rather a secondary amine attached to an α-carbon, and thus in this case HCN is not likely to be the product of proline degradation. It has been shown that proline is an effective quencher for singlet oxygen due to the cyclic secondary amine and has a low ionization potential (Alia et al., 2001). Additionally, secondary radicals formed from proline are more stable than primary radicals formed from glycine; thus the proline degradation is much faster.
The absence of significant degradation for proline under Mars humid conditions was likely caused by the opaque frost that formed on the surface of the films. It has been observed that snow in Antarctica can reflect 96–98% of UV and visible radiation (Grenfell et al., 1994). Similarly, the frost on the films reflected most of the UV radiation, shielding the amino acid and thus leading to a decrease in degradation. The frost formation was unavoidable due to the increased water vapor present under higher humidity. No frost formation was observed on any of the sand-based samples.
4.2. Amino acids combined with sand and sand+salt
Through confirmatory tests, it was found that generally >80% recovery of amino acids is possible when making amino acid + sand with/without salt stock compared to a direct amino acid addition. Thus, the amount of amino acid present in the amino acid + sand samples was slightly less than the amount of amino acid in the film form. This does not present a problem since comparisons are made in terms of % differences.
4.2.1. Glycine with sand and sand+salt
Glycine+sand displayed increased degradation in all the samples under both Mars ambient and humid conditions compared to the glycine film. This is likely the result of silica sand particles providing a larger surface area and therefore more reactive sites (Carrier and Kounaves, 2015). It has been shown that the presence of water (usually 10–1000 ppm in the atmosphere) is necessary for amino acid degradation under Mars ambient conditions, while the absolute level of water does not seem to affect the reaction rate significantly (ten Kate et al., 2006). The absorption of UV by water is negligible, although UV radiation at 242 nm wavelength has been shown to cause H2O dissociation to produce •H and •OH radicals (Okabe, 1978).
There was only a slight increase in degradation observed for glycine+sand under both Mars ambient and humid conditions. However, in the presence of NaCl, NaClO3, or NaClO4, the degradation increased for Mars humid compared to Mars ambient conditions, although no significant difference between NaCl, NaClO3, or NaClO4 was observed. This indicates that at Mars ambient conditions with the salt concentrations used here (6000 ppm in each 0.5 g amino acid + sand + salt sample), the amount of oxidizing intermediates generated after 24 h of UV irradiation was not sufficient to cause significant degradation of glycine.
4.2.2. Proline with sand and sand+salt
For proline+sand without any salts, there was increased degradation under both Mars ambient and humid conditions compared to the proline film sample. This trend is similar to what was noted in the previous section with glycine+sand. For proline+sand with NaCl, NaClO3, or NaClO4, it appears that the salts were protecting proline from degradation under Mars ambient conditions and accelerating it under Mars humid conditions following this trend: No Salt < NaCl < NaClO3 < NaClO4. This difference was especially obvious for the degradation of proline mixed with sand+NaClO4 where it was only ∼5% under Mars ambient conditions yet ∼60% under Mars humid conditions. This difference cannot be explained solely by either oxidizing intermediates or absorption of UV by the salts because NaClO3 has a higher and broader UV absorption and is expected to be more reactive than NaClO4 in both cases. This again confirms that the degradation of proline was not driven by the oxidizing power of the oxychlorine salts or the intermediates produced under UV irradiation.
It has been suggested that in an O2 saturated aqueous solution irradiated with γ-rays, proline would be converted to hydroxyproline, initiated by intermediary •OH radicals and peroxide (Kopoldová and Voráček-Hübsch, 1975). On the other hand, both decarboxylation and addition of CO2 - to proline radicals to form dicarboxylic acid were observed in a nitrogen atmosphere. However, as discussed above, •OH generation in this case was not likely to be significant due to the UV wavelength used.
Another possible reaction pathway would be initiated by reacting singlet oxygen with proline, an effective singlet oxygen quencher in aqueous solution, though the exact mechanism for this reaction is unclear (Alia et al., 2001). It has also been shown that UV radiation can induce photodissociation of molecular oxygen and generate an oxygen radical, ozone, as well as singlet oxygen (Brasseur and Solomon, 2006), as shown in the reactions below:
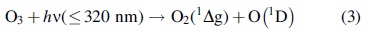
Due to the low oxygen content of the Mars simulation gas (<0.1%) and the significant difference between glycine and proline absorption, it is not likely that any of the above mechanisms occurred with proline under Mars ambient conditions. If oxygen radicals or singlet oxygen are key for proline degradation, its degradation under Mars humid conditions should decrease compared to Mars ambient conditions, as an increased H2O level would reduce the amount of reactive oxygen species available to proline.
Given the above, it is proposed that the absorption of a UV photon by proline under Mars ambient conditions generates unstable radicals (Fig. 4). For photolysis of secondary cyclic amines, it has been shown that breaking the N-H bond is likely the dominant dissociation process in nonpolar solvents and that breaking the C-N bond is more likely in polar solvents (Cohen et al., 1973). The adsorbed H2O and -OH on the silica surface could then react with the proline radicals and convert them to the two products shown in Fig. 4. It has been shown that different forms of amorphous silica adsorb the same amount of H2O under the same water partial pressure (Iler, 1979). The reacted H2O and -OH groups can then be regenerated with the H2O vapor present in the Mars gas (H2O < 0.1%). Under Mars ambient conditions, when NaCl, NaClO3, and NaClO4 are present they will compete for the H2O and -OH groups; thus the degradation of proline would be hindered. Under Mars humid conditions, the H2O is in excess, and the salts will then aid in increasing the adsorbed H2O on the silica sand and thus accelerate the degradation. The presence of water could also leach the adsorbed amino acid from the sand, making it more susceptible to degradation (Fornaro et al., 2018). The ability of the salt to promote degradation depends on its solubility in H2O. For NaCl, NaClO3, and NaClO4, their solubility at 25°C is 36, 100, and 209 g/100 g of H2O, respectively (Lide, 2004), and their deliquescence power follows with NaClO4 > NaClO3 > NaCl. These salts are likely to behave similarly with the glycine+sand samples. However, the effects were not as obvious as with the proline+sand samples due to the structural stability of glycine as discussed above.

Proposed mechanism for the degradation of proline by UV under Mars ambient conditions. The H2O and -OH on the SiO2 are regenerated with the <0.1% H2O present in the simulated Mars gas.
4.2.3. Tryptophan with sand and sand+salt
Irradiation of tryptophan as a film or with any sand or salt resulted in a yellowish product, in stark contrast to glycine and proline. To explain this discoloration, it is proposed that the aromatic indole ring in tryptophan enables it to absorb much more UV energy than glycine or proline. Thus, the bond breakage in tryptophan occurs mainly on the indole ring under Mars ambient conditions and is more significant but independent of the humidity level (Creed, 1984).
Although identification of this yellow product was beyond the scope of this research, one possible candidate is β-carboline formed via UV-driven decarboxylation. This has previously been shown to be the product of thermally decomposed tryptophan (Cuq and Cheftel, 1983). Another possible candidate is 1H-indole-3-carboxaldehyde, which was determined to likely be the dominant fragment of tryptophan photodissociation under 265 nm UV (Talbot et al., 2005). Whatever this degradation product is, the salient accompanying effect under Mars ambient conditions is that it provides some protection from UV and thus further degradation of the tryptophan.
5. Conclusions
The direct UV degradation of glycine, proline, and tryptophan was studied under Mars ambient and humid conditions, as a film or with addition of NaCl, NaClO3, or NaClO4. These amino acids demonstrated different degradation behaviors. Glycine, being the most stable amino acid tested, showed no significant destruction in any of the samples under Mars ambient conditions. However, the degradation of glycine increased in the presence of NaCl, NaClO3, and NaClO4 but with no significant difference between the three salts. The degradation of proline was significantly greater than that observed for glycine, even though the molar absorptivity of glycine and proline are similar for UV between 200 and 400 nm. In addition, the degradation order for proline was No Salt > NaCl > NaClO3 > NaClO4 under Mars ambient conditions, but under Mars humid conditions, the order of degradation for proline was reversed to NaClO4 > NaClO3 > NaCl > No Salt. This difference is likely due to the difference in the affinity of each salt toward water.
For tryptophan, no difference was observed between Mars ambient and humid conditions. This suggests that tryptophan possesses a different degradation mechanism compared to glycine or proline. This is likely the result of tryptophan's strong broad UV absorption, which allows significantly more UV energy for degradation compared to glycine and proline. The opaque products resulting from the tryptophan degradation can also act as a UV screen for the amino acids immediately below and protect them from further photochemical degradation.
Addition of NaCl, NaClO2, NaClO3, or NaClO4 to the amino acid+sand samples resulted in different behaviors for glycine, proline, and tryptophan. Under Mars ambient conditions, degradation generally increased for glycine but not for proline. The salts in proline+sand appear to be protecting proline from UV degradation, with the proline+sand+perchlorate sample showing only 6% degradation.
Footnotes
Acknowledgments
We would like to thank Dr. Minghua Zhuo for helping with the possible mechanisms for amino acid degradation and Robert Bolhofner for his contribution to some of the preliminary work at the beginning of this project. We also would like to thank Dr. Neveda Naz for proofreading this paper and the reviewers for their helpful suggestions.
Author Disclosure Statement
The authors have no competing or personal financial interests.
Funding Information
This work was carried out in the Chemistry Department of Tufts University, supported by the National Aeronautics and Space Administration (NASA) through the Mars Fundamental Research Program (MFRP) under grant NNX14AG40G and the Solar System Workings (SSW) Program under grant 80NSSC20K0460. The ESI-MS data were acquired with an instrument procured with funding from the National Science Foundation (NSF) under grant CHE-0320783.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.