
Abstract
The possibility of biological transfer between planetary bodies is seldom factored into life detection strategies, although the actuality of such an event would have profound implications for how we interpret potential biosignatures found on other worlds. This article addresses the possibility of life on Mars in the context of a biological transfer and an independent genesis of life. The phylogenetic tree of life on Earth is used as a blueprint to interpret evidence of life and as a guideline to determine the likelihood that potential biosignatures could be expressed by martian organisms. Several transfer scenarios are considered, depending on the timing of transfer with respect to the evolution of life on Earth. The implications of each transfer scenario and an independent genesis of life on the biochemical nature of the resulting martian organisms are discussed. The analysis highlights how conceding the possibility of a biological transfer has practical implications for how we search for evidence of life, both in terms of the quality of potential biosignatures and the likelihood that certain biosignatures might be expressed. It is concluded that a degree of uncertainty on the origin of martian organisms might be unavoidable, particularly in the absence of a biochemical context.
1. Introduction
The next decades of planetary exploration will witness a surge in missions that search for evidence of life in the solar system. In the mid-to-late 2020s, two missions will search for evidence of life in ancient martian sedimentary deposits, either in situ (Vago et al., 2015) or in samples returned to Earth (Mustard et al., 2013). In the mid 2030s, the Dragonfly mission will search for evidence of prebiotic chemistry and possible biosignatures on Saturn's moon Titan (Turtle et al., 2018). Mission concepts exist to search for evidence of life on Saturn's moon Enceladus (Reh et al., 2016; Eigenbrode et al., 2018; MacKenzie et al., 2021) and Jupiter's moon Europa (Hand et al., 2017) in the late 2030s and into the 2040s.
The search for evidence of life on other worlds is a means to address questions in biology that cannot otherwise be answered based solely on the study of terrestrial life (Lederberg, 1960; Dick, 2012): What is life when considered in the universal context? Is there a general theory of living systems? How does life originate, and what is the role of chance and necessity in the origin of life? Is life a common cosmic phenomenon or an improbability of cosmic dimensions? Exobiology research is grounded on the premise that these questions can only be answered through the study of forms of life that are unrelated to Earth life—the result of an independent genesis of life (McKay, 2004, 2010). Implicit in this view is the assumption that finding evidence of life beyond Earth would constitute evidence of an independent origin of life event.
However, it is important to recognize that planetary bodies are not biologically isolated, and meteoritic exchange could have been an effective mechanism for transferring life within the solar system (Melosh, 1988; Mileikowsky et al., 2000; Worth et al., 2013). In addition, the possibility cannot be discarded a priori of interstellar panspermia (Napier, 2004). Both scenarios could lead to forms of life that do not share a common planet but do share a common origin.
Assuming an origin of life on Earth, >300,000 objects could potentially have transferred viable organisms to Mars in the past 3.5 billion years. Assuming an origin of life on Mars and later transfer to Earth, the estimated number of viable transfers rises to >20,000,000 (Worth et al., 2013). A viable transfer represents an unsterilized rock fragment that is at least 3 m in diameter, sufficient to protect dormant organisms from ionizing radiation damages for up to 10 million years (Mileikowsky et al., 2000; Worth et al., 2013). These estimates apply to post-Late Heavy Bombardment (LHB) times, and as many transfers could have occurred during the LHB (4.1–3.8 Gyr ago) (Fig. 1). While the probability of a biological transfer between the inner and the outer solar system is much lower, it is not zero (Worth et al., 2013).
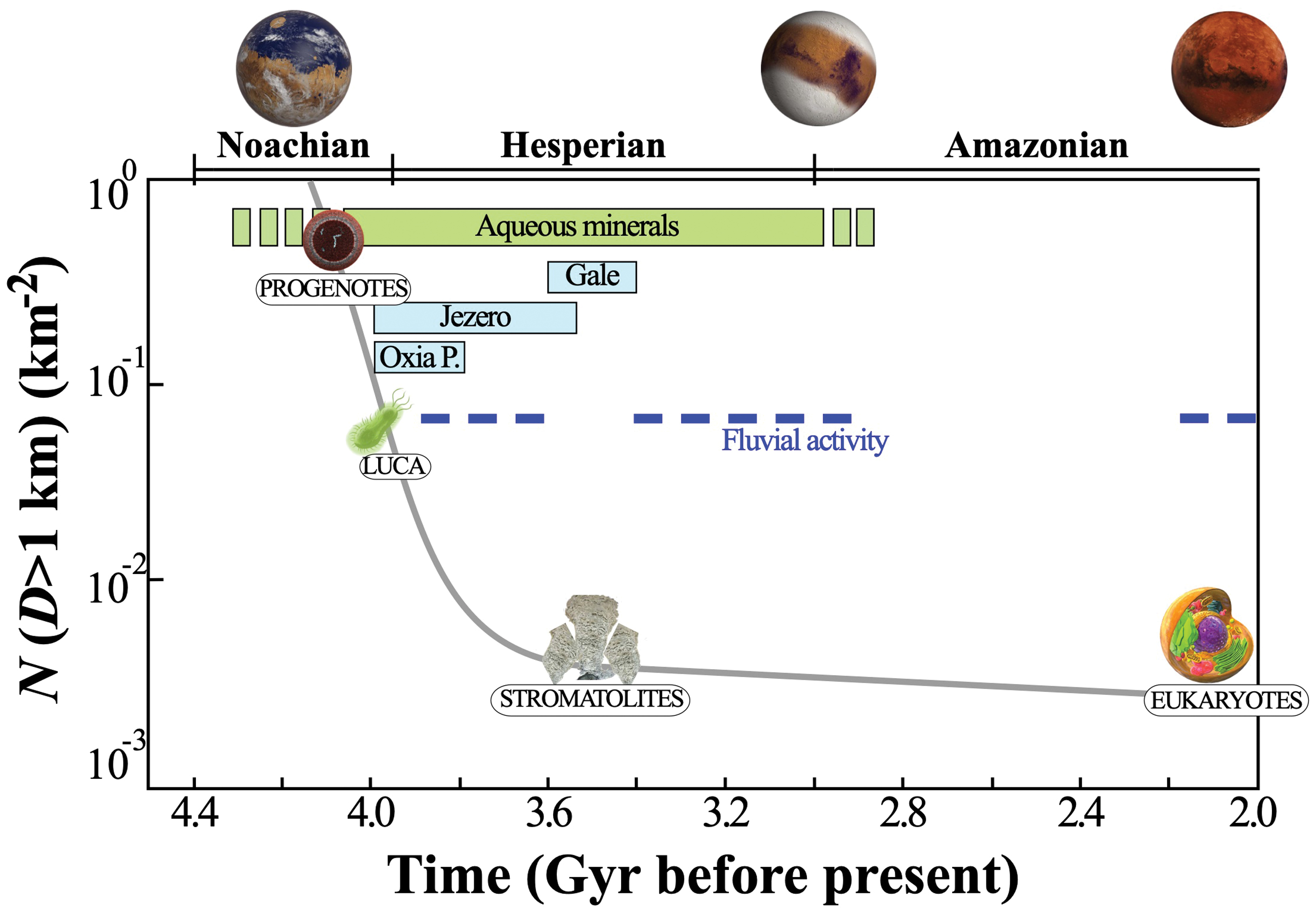
Terrestrial impact rates with major evolutionary events (gray line) and martian habitability conditions (indicated by fluvial activity and aqueous minerals) during the first 1.5 Gyr. The likelihood of a biological transfer from Earth to Mars decreased exponentially during the first billion years, along with the rate of large impacts. The end of the high-impact rate period coincides with the oldest fossil stromatolites found in the Earth's Archean sedimentary record. A biological transfer to Mars during that period could have been in the form of progenotes or early prokaryotes (archaea and eubacteria). Transferred progenotes or prokaryotes would have encountered a habitable planet with diverse aquatic environments, and widespread habitable conditions, based on the martian fluvial and sedimentary record. A biological transfer could have been recorded in sedimentary deposits that are being investigated by NASA's Curiosity rover (Gale crater), or that will be investigated by NASA's Perseverance rover (Jezero crater) and ESA's Rosalind Franklin rover (Oxia Planum). A transfer of eukaryotes could have occurred in the last 2 Gyr, but the transferred organisms would have encountered a hyperarid and hypothermal desert. Impact rates for bodies >1 km are based on Bottke and Norman (2017). Color images are available online.
The possibility of biological transfer between planetary bodies is seldom factored into life detection strategies, although it could have profound implications for how we interpret biosignatures found on other worlds. The selection of potential biosignatures in life detection missions is often based on the likelihood that a given biological trait might be expressed in the target planetary body. This likelihood is typically estimated based on the biosignature's reliability (how likely is a potential biosignature to have been produced by life), its survivability (how likely is a biosignature to have been preserved or to have persisted in its environment), and its detectability (how likely is the biosignature to be observed or measured), although additional criteria can also be implemented (Neveu et al., 2018). Seldom, if ever, the selection of target biosignatures is based on their power to discriminate between an independent genesis and a biological transfer.
This article addresses the search for evidence of life beyond Earth in the context of a biological transfer and an independent genesis. The analysis presented focuses on Mars because it is an immediate target for life detection missions, and because of the relatively high likelihood of biological exchange with Earth. It is assumed that martian life, regardless of its origin, would have endured until the present—although confined to cryptic refuges near the surface or in the deep subsurface with little detectable impact on the environment. This assumption allows for exploration of the full range of potential biosignatures that could be discovered, from signatures of an extant biosphere to signatures of extinct life that might have formed billions of years ago and could still survive in the geologic record. Model estimates predict an even higher likelihood of biological exchange between Earth and Venus (Worth et al., 2013). However, while Venus could have been habitable early in its geologic history (Grinspoon and Bullock, 2007), it is not currently considered a priority target for life detection missions; its surface is currently uninhabitable and it has been heavily modified by volcanic processes (Strom et al., 1994). On the contrary, subsurface oceans in some icy moons of the outer solar system appear to be habitable (McKay et al., 2008), but the likelihood of biological exchange between Earth and those icy moons is orders of magnitude lower than that of Earth and planets of the inner solar system (Worth et al., 2013). Regardless, many of the arguments and conclusions presented here are not planet specific and would also apply to the search for evidence of life on Venus and on icy moons.
2. The Evolutionary Tree: A Blueprint to Interpret Evidence of Life Beyond Earth
Conceptually, a biological transfer between Earth and Mars implies that life on both planets would share a common ancestor. Operationally, a biological transfer implies that martian life would map in the evolutionary tree of terrestrial life. The evolutionary tree is a visual representation of the phylogenetic relationships between extant and extinct organisms (Fig. 2A). The tree encompasses three major domains (Eubacteria, Archaea, and Eukarya). Multiple lines of evidence suggest that all living organisms within the three domains are descendants of a population of organisms that existed ∼3.9 Gyr ago (Betts et al., 2018)—commonly referred to as the Last Universal Common Ancestors (LUCA). Earlier depictions of the tree anchored the three domains to one common node. However, more modern versions of the tree provide support for two primary domains, the Eubacteria and the Archaea, with the Eukaryotes arising through partnerships with them (Williams et al., 2013). In an evolutionary tree with two primary domains, the LUCA were prokaryotes, or closely related ancestors with a prokaryote-like cell architecture. The analysis presented in the present study assumes an evolutionary tree with two primary domains, although the main conclusions would also apply to an evolutionary tree with three primary domains.
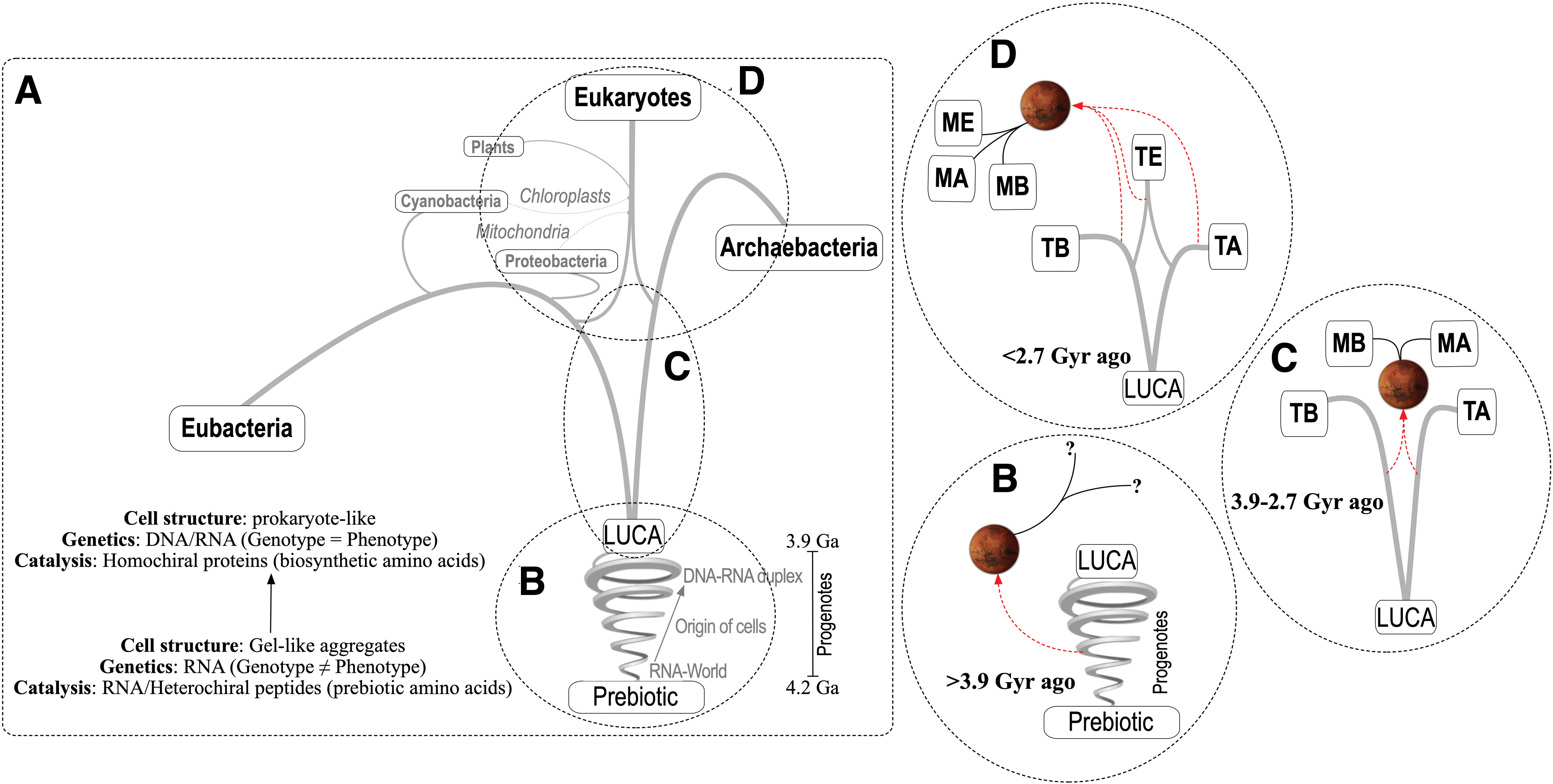
Before the LUCA there must have been a period of intense biochemical evolution, initiated by the first self-replicating evolving system we call life. The proposed scenario for this time period is called the era of progenotes (Damer and Deamer, 2020). Progenotes are conceptualizations of the first self-replicating and evolving systems that emerged from the prebiotic world, and whose chemical evolution eventually led to the first cells. Earlier progenotes have been described as gel-like aggregates that passively coalesced and disaggregated, shared a collective pool of monomers and simple polymers, and were capable of minimal metabolic functions (Woese, 1998).
Biochemistry during the progenote era was messy, with each protocell representing a natural chemical experiment, slowly evolving through a gradual process of combinatorial selection of favorable traits. Information would have been swapped largely via horizontal transfer, and evolution would have operated mostly through mechanisms of group selection of progenote populations (Arnoldt et al., 2015). The most basic and common biochemical underpinnings of the LUCA and of all extant terrestrial organisms “crystallized” during the era of the progenotes (Section 3.1.1) (Woese, 1998). Ultimately, this evolutionary process led to increasing cellularization and, eventually, to the LUCA, which might be considered both the last progenotes and the first prokaryotes. The estimated age of the LUCA implies a relatively early and short period of progenote evolution, likely within a window of 300–400 million years after planetary differentiation.
The evolutionary tree does not capture events that occurred in the progenote era, because before the LUCA, genetic lineages could not be easily established in the absence of a universal genetic code and of reliable DNA replication and RNA translation systems. Hence, while partial reconstructions of the LUCA are possible based on conserved biochemical traits in extant organisms (Weiss et al., 2016), the biochemical nature of progenotes becomes more difficult to recreate with certainty. This makes the LUCA a “biochemical event horizon” beyond which it is difficult to extract accurate biochemical and physiological information.
An independent genesis of life implies the emergence of a separate phylogenetic tree, with no hereditary links to the domains Eubacteria, Archaea, or Eukarya, the LUCA, or their progenote ancestors. Conversely, a transfer of life between Earth and Mars implies that martian organisms would map within the phylogenetic tree, either as part of one of the existing domains, or as a separate domain entirely. In a transfer scenario, terrestrial and martian life would share a common ancestor, but the biochemical nature of the resulting martian populations would have depended on the timing of the transfer. In the next section, different scenarios for life on Mars are discussed in the context of a biological transfer and an independent genesis, along with the possible implications of each scenario for the biochemical nature of the resulting organisms.
3. Scenarios for Life on Mars
3.1. Biological transfer scenario
Mars had a global magnetic field until ∼3.9 Gyr ago (Acuna et al., 1998) and a relatively thick atmosphere at least as far back as ∼3.5 Gyr (Wordsworth, 2016). Widespread habitable conditions, defined as the coexistence of liquid water, nutrients, sources of energy, and physicochemical conditions (e.g., temperature, pH, salinity) compatible with the stability of covalently bound biomolecules, might have persisted near the surface until ∼3 Gyr ago (Grotzinger et al., 2014) and until much later in the deep subsurface. Arguably, microorganisms transferred from Earth during the first 1.5 Gyr would have likely found a suitable niche on Mars.
A biological transfer between Earth and Mars could have occurred anytime between 4.3 Gyr ago—the current upper time boundary for the origin of life event—and the present, but the likelihood of a viable transfer decreases with time, as impact rates lowered and as Mars became increasingly uninhabitable (Fig. 1). The current earliest evidence of life on Earth (3.9 Gyr ago) (Tashiro et al., 2017) places an upper limit for a viable transfer from Mars to Earth. A viable transfer from Earth to Mars could theoretically have occurred at any point in the past. For the purpose of this analysis, the direction of transfer is not important—for simplicity a biological transfer from Earth to Mars is assumed; what matters is the timing of the transfer with respect to the LUCA. Two end-member transfer scenarios can be envisioned: (1) progenote transfers and (2) post-LUCA transfers.
3.1.1. Progenote transfers
Multiple transfers of terrestrial progenote populations to Mars could have occurred between 4.3 and 3.9 Gyr ago, a time frame that is bracketed by the process of planetary differentiation and the appearance of the LUCA (this time bracket might shift with new discoveries that better constrain the actual age of the LUCA). This time period overlaps with the LHB (Fig. 1). Models predict that as many as 1.2 × 1011 kg of material could have been transferred from Earth to Mars during this time span, under conditions compatible with the survival of known microorganisms (Worth et al., 2013). More transfers might have occurred under conditions that lie beyond the survival capabilities of known microorganisms.
As discussed below, the concept of progenote is not a unitary entity, but encompasses a sliding scale of biological complexity. Progenotes more closely related to the LUCA must have been fairly similar to that entity and, therefore, similar to fully formed and fully functional prokaryotes. A transfer to Mars of such evolved progenotes is better considered in the next section. For the purpose of this scenario, a progenote transfer refers to an earlier event, when the boundaries between biochemistry and prebiotic chemistry were less clearly defined. In this scenario, life on Earth and on Mars would share a common biochemical foundation (Fig. 2A), but given that, before the LUCA, genetic lineages could not have been firmly established, it might not be technically accurate to speak of progenote ancestors.
The biochemistry of the earlier progenotes was likely profoundly different from modern terrestrial organisms (Woese, 1998), possibly including an early emergence of phosphate-free biochemistry (Goldford et al., 2017). Based on the RNA-peptide world hypothesis (Gilbert, 1986), earlier progenotes lacked DNA and likely had an incomplete set of proteinogenic amino acids, which appear to have been incorporated sequentially in several stages during the progenote era, with a unique order: Gly&Ala<Val&Asp<Pro<Ser<Glu&Leu<others (“<” denotes earlier than) (Trifonov, 2000). Accordingly, earlier progenotes must also have lacked complex proteins. Instead, short peptides that formed abiotically from a feedstock of prebiotically available amino acids would have acted primarily as RNA cofactors, performing basic functions such as nucleic acid-binding and catalysis (Piette and Heddle, 2020).
During the earlier stages of progenote evolution, cellular architecture must have been rudimentary in every aspect, likely lacking a cell wall (Kandler, 1995). The unsophisticated biochemistry of earlier progenotes means they were highly dependent on the geochemical environment they inhabited. For example, progenotes lacking cell walls and complex transmembrane proteins could not generate ion gradients for energy conservation and must also have lacked complex soluble proteins to catalyze substrate-level phosphorylation reactions. Therefore, early progenotes might have been forced to harness an external, geochemically created proton-motive force for energy transduction, before evolving the tools to generate their own (Lane et al., 2010). As a result, early progenotes likely had an intimate relationship with their environment, to the point that biochemistry and geochemistry were inseparable.
It has been proposed that during the earlier stages of progenote evolution, biochemistry might have been contained not within organic membranes, but within mineral spaces in alkaline vents, where far-from-equilibrium conditions provided a continuously reactive chemical environment that could be exploited for organic synthesis, and where inorganic reactions between H2, CO2, and Fe(Ni)S minerals might have served as template for the evolution of the first core pathways in a carbon and energy metabolism (Martin and Russell, 2007; Martin et al., 2008; Lane et al., 2010). While alternative scenarios have been proposed for the evolution of early progenotes (Mulkidjanian et al., 2012; Damer and Deamer, 2020), this example serves to illustrate the necessary tight-coupling between early progenotes and their environment. This must be taken into account when considering the likelihood of a successful transfer to Mars, as discussed below.
As evolution progressed, the biochemistry and cellular structure of progenotes became increasingly more complex. DNA emerged as a more stable solution for information storage. Cell membranes became more complex, impermeable, and selective, and were populated with transmembrane proteins and complexes capable of generating, maintaining, and exploiting ion gradients for energy conservation. Cell walls must have evolved during this later stage of progenote evolution. Greater biochemical complexity led to the first fully contained pathways of carbon and energy metabolism, although metabolic diversity among progenotes was likely minimal. Metabolic reconstructions of the LUCA suggest that H2-dependent acetogenesis and methanogenesis evolved during the era of progenotes (Weiss et al., 2016). Anaerobic fermentation might also have evolved during that time (Adam et al., 2018). Other major pathways for carbon and energy metabolism require more complex enzymatic capabilities and appear to postdate the LUCA (Moore et al., 2017).
Progenotes transferred to early Mars would likely have encountered a habitable environment not too dissimilar from early Earth (Section 3.2). However, the concept of habitability has a different meaning with regard to the progenote era, and this remains an important gap in models of planetary biological transfer, which typically consider biological survival requirements based on biochemically sophisticated organisms (Mileikowsky et al., 2000; Worth et al., 2013).
A biochemically mature organism might be equipped to survive planetary transfer in a dormant state and to adapt and grow in the new environment at the destination. However, the “survival” requirements for early progenotes would have been different. A successful transfer of early progenotes would have required conditions that minimized damage to “naked” biomolecules such as RNA oligomers and short peptides during interplanetary cruise (an unlikely event given the fragility of RNA even under ideal conditions of preservation). In addition, the chemical cargo would have had to land in a spot that was geochemically very similar to the point of departure—here is where the intimate relationship between early progenotes and their environment becomes an important factor. While the most common environments referenced in the literature for the origin and evolution of early progenotes on Earth likely coexisted on Mars (Section 3.2), the likelihood of molecular survival during transfer, and of precise landing in the right environment, must have been significantly lower for early progenotes than for more sophisticated and autonomous post-LUCA organisms (Section 3.1.2).
Rather than biological seeds, planetary transfers during the progenote era might have acted as chemical seeds, potentially contributing to the organic inventory and chemical complexity of early Mars. Taking model predictions of the number of viable transfers during the LHB as an upper limit (∼360,000 transfers, equivalent to 1.2 × 1011 kg) (Worth et al., 2013) and assuming an average progenote mass fraction in the ejected material of a few percent (equivalent to the average organic content of typical carbonaceous chondrites), as much as a few kilograms of progenote “biomass” could have been transferred from Earth to Mars every year during the LHB period. For reference, the flux of unaltered (nonterrestrial) meteoritic carbon onto the martian surface during this early period could have been as high as 105–108 kg/yr (Flynn, 1996), but only a fraction of this meteoritic carbon (<10–4) would have been in the form of biochemical building blocks such as amino acids.
Therefore, the transfer of progenote “biomass” from Earth to Mars during the LHB might have been sufficiently high to steer organic chemical evolution on Mars along an Earth-like path, by contributing a significant fraction of relatively complex prebiotic intermediaries such as nucleotides, oligopeptides, or membrane-forming lipids (Table 1). Notably, the estimated rates of viable transfers from Mars to Earth, and from Earth to Venus, are an order of magnitude higher (Worth et al., 2013), and so, the exchange of complex prebiotic compounds between the terrestrial planets might have been prolific during the LHB.
Possible Manifestations of Fundamental Biochemical Traits in Martian Life in the Context of a Biological Transfer and a Second Genesis
3.1.2. A post-LUCA transfer
The ideal window for a successful transfer of life from Earth to Mars might have occurred between 3.9 and 3.4 Gyr ago, due to the confluence of three factors: (1) a relatively high rate of transfer events; (2) the existence of complex forms of life potentially capable of surviving interplanetary travel; and (3) widespread habitability conditions on the surface of Mars. Estimated rates of successful transfer events indicate a peak during this period (∼500–1000 events/million years, Worth et al., 2013), but they suggest a decrease in events thereafter due to lowering impact rates and impact sizes (Fig. 1).
Transferred populations would have been biochemically and physiologically sophisticated and ecologically diverse. Conserved biochemical pathways and sequence comparisons of nucleic acids and proteins in extant terrestrial organisms suggest that the LUCA (3.9 Gyr ago) already had an almost fully formed cellular architecture including a complex cell membrane and a cell wall, a chemiosmotic system for energy conservation, a fully developed genetic code, a rudimentary DNA-based replication system, a translation system that was as advanced or very similar to the modern version, and ribosomal protein biosynthesis based on 20 genetically encoded α-
The new martian populations might have been capable of sulfur reduction, sulfate reduction, methanogenesis, and anoxygenic photosynthesis, although other metabolisms such as oxygenic photosynthesis, methane oxidation, nitrogen fixation, nitrification, and denitrification, which only appeared on Earth after the initial buildup of oxygen (Moore et al., 2017), would have had to evolve independently on Mars. The geologic and mineralogic record on Mars suggests that prokaryote populations successfully transferred during this period could have found suitable habitats. For example, sedimentary deposits investigated by NASA's Curiosity rover at Gale crater were laid down during this period and are indicative of a protracted aquatic environment with low salinity, neutral to alkaline pH, and anoxic conditions that contained all the basic requirements to sustain life (Grotzinger et al., 2014; Hurowitz et al., 2017). Similar aquatic environments were likely widespread on Mars during this period (Carr and Head, 2010; Ehlmann and Edwards, 2014), including the purported lake that occupied Jezero crater, the target destination of NASA's Perseverance rover to search for potential signatures of an early martian biosphere.
A later transfer postdating the appearance of Eukaryotes (<2.7 Gyr ago) also deserves consideration (Fig. 2D). Rates of successful transfer events during that time would have been lower by an order of magnitude (<100 events/million years, Worth et al., 2013). This timing would have coincided with the early Amazonian period, when the surface of Mars was already a hyperarid and hypothermal desert (Carr and Head, 2010), and therefore, transferred microorganisms would have had to withstand water and radiation stress along with globally freezing temperatures—the hallmarks of land-dwelling microorganisms. While far less likely, such a late transfer cannot be discounted a priori, given that the so-called Terrabacteria, which includes members of desert-dwelling cyanobacteria, the Gram-positive phyla, and the Chloroflexi and Deinococcus-Thermus, might have emerged on Earth as early as ∼3.5 Gyr ago (Battistuzzi and Hedges, 2009).
Arguably, land-adapted microorganisms capable of surviving the transfer event might also have been able to survive exposure to martian near-surface conditions. However, the rates of biological proliferation and adaptation in the ensuing martian populations would have been impacted by the general decline in habitability conditions, at least near the surface. Populations of microorganisms transferred during this later period might have had a better chance of survival in the deep subsurface in locations shielded from harmful radiation where geothermal heat could still sustain deep-seated liquid water aquifers and water/rock reactions could generate chemical sources of energy. Notably, such favorable conditions might have persisted in the subsurface of Mars until the present (Stamenković et al., 2019).
Irrespectively of the exact timing of the transfer event, evolution by natural selection would have acted upon the terrestrial and the martian biospheres independently and at rates and intensities modulated by the respective planetary environments. Therefore, a certain degree of biochemical divergence through time ought to be expected between the terrestrial and martian “populations,” even in fundamental biochemical traits. For example, the genetic code, which is one of the most universally conserved biochemical traits of life on Earth, still has a degree of evolvability as reflected by the recruitment of nonstandard amino acids (Koonin and Novozhilov, 2009) and singular events of codon reassignment (Sengupta and Higgs, 2015).
However, biochemical divergence between terrestrial and martian life after a post-LUCA transfer would likely be limited because the rates of random mutations and genetic drift tend to be slow even over geologic timescales. This is because while mutations can be advantageous, neutral, or harmful, most with fitness effects are deleterious. Also, fitness-increasing beneficial mutations (i.e., selectable mutations) constitute only a small fraction of all possible mutations (Eyre-Walker and Keightley, 2007). As a result, higher mutation rates tend to have diminishing returns (Sprouffske et al., 2018). In addition, random mutations and genetic drift operate on base sequences that are then translated into changes in peptide sequences, and while the resulting phenotypic changes can be significant, these genetic alterations do not alter the underlying translation machinery, which, for post-LUCA organisms, already involved mRNA/tRNA, the ribosome, and the set of 20 proteinogenic amino acids (with the rare exceptions of new amino acid recruitment and codon reassignment mentioned above). The fact that, on Earth, the genetic code and the underlying translation machinery have remained practically unchanged for >3 Gyr (Koonin and Novozhilov, 2009) suggests that martian descendants of post-LUCA populations would likely share many of the same biochemical hallmarks of extant terrestrial organisms, even after several billion years of independent evolution (Table 1).
3.2. Second genesis scenario
The possible nature of an independent genesis of life has been discussed in the literature, particularly with regard to its biochemistry (Ward and Benner, 2007). The analysis below assumes a water-based biochemistry with C-H as the basic molecular scaffolding. For clarity, the origin of life is defined here as the appearance of a self-replicating and evolving chemical system. In the case of Earth, the simplest expression of such a system commonly referenced in the literature is catalytic RNA (i.e., a ribozyme). Consequently, an independent genesis of life on Mars implies the appearance of an equivalent molecule with similar capabilities. Similar to the case of life on Earth, it is further assumed that an original self-replicating and evolving chemical system on Mars (the Ur-molecule) would have to have evolved in biochemical complexity to survive until the present.
Considering fundamental chemistry principles and the range of organic chemical compounds that could theoretically exist (Meringer et al., 2013), the biochemical space available for martian life in the context of an independent genesis could be almost unlimited and difficult to constrain. However, this vast biochemical space is significantly reduced with two considerations. First, the theory of organic chemical evolution states that life originates from a sequence of prebiotic chemical steps. This implies that certain biochemical traits must be reminiscent of the prebiotic world whence life originates. This prebiotic endowment is manifested in terrestrial biochemistry, where 50% of the proteinogenic amino acids are common products of prebiotic chemical reactions. Similarly, prebiotic chemistry would have shaped, at least in part, the biochemistry of an independent genesis of life on Mars. Second, a certain amount of biochemical overlap could be expected between populations that originate independently, but in a similar prebiotic environment (Davila and McKay, 2014). Therefore, a key question when considering the biochemical nature of an independent genesis of life is how similar were prebiotic Earth and early Mars?
The two most common environments referenced in the literature for the origin of life on Earth are alkaline hydrothermal vents in the ocean bottom (Russell and Hall, 1997) and subaerial hot spring pools (Damer and Deamer, 2020). There is tantalizing evidence that both types of environments existed on Mars during the Noachian and Hesperian periods (Ruff and Farmer, 2016; Michalski et al., 2017). Both aquatic environments would have been in contact with an atmosphere likely dominated by CO2, N2, and H2O (McKay and Stoker, 1989). Punctuated volcanic emissions of SO2, and H2 and CH4 emissions from surface and subsurface water/rock interactions, would have been a source of reducing power. Similar to the case of Earth, large impacts could also have delivered transient, although large, amounts of reducing power (Benner et al., 2020). Volcanic lightning could have been an endogenous source of hydrogen cyanide (HCN) (Segura and Navarro-González, 2005), and photochemical reactions in the atmosphere could have formed other reduced organic compounds that are considered primary precursors of prebiotic building blocks (e.g., H2NCN, HCCCN, NCCN, and NCHO) (Benner et al., 2020).
The relatively small water inventory of early Mars compared with early Earth would also have created favorable conditions to support the networks of prebiotic chemical reactions that could have led to the abiotic formation of RNA-like replicators (Benner and Kim, 2015). Exogenous sources (i.e., carbonaceous chondrites, comets) of organic matter, including prebiotic building blocks such as nucleobases and amino acids, would have been the same for early Earth and early Mars (Burton et al., 2012). Therefore, it is conceivable that early Mars was very similar to early Earth in terms of aqueous environments, prebiotic chemistry, and availability of prebiotic building blocks.
Based on the above, a certain degree of biochemical overlap could be expected between life on Earth and on Mars even in the context of an independent genesis, at least in part, due to similar sources of prebiotic organic matter (e.g., carbonaceous chondrites, comets) (Weber and Miller, 1981; Davila and MacKay, 2014). The degree of overlap would have been highest during the earlier stages of biological evolution (e.g., the protocell or progenote period) and decreased with time as evolutionary processes acted independently upon both populations. Due to the stochastic nature of evolutionary processes, the biochemical and cellular nature of more evolved forms of life cannot be anticipated with certainty. However, certain predictions might still be possible regarding the biochemistry of an independent genesis of life on Mars, particularly at the level of essential heteroatoms and molecular building blocks (Table 1).
Biomolecules with a C-H scaffolding would likely have been decorated with nitrogen, oxygen, and sulfur heteroatoms, all of which would have been abundant in early Mars, but phosphorous could theoretically have been substituted (Hoehler et al., 2020). If amino acids played a biochemical role in early martian forms of life, then prebiotically abundant amino acids such as glycine (Gly) and alanine (Ala) would likely have been utilized (these amino acids were assimilated first by terrestrial progenotes), but not the biosynthetic ones such as tryptophan (Trp) and tyrosine (Tyr) (these amino acids were assimilated last by terrestrial progenotes, after the evolution of a more complex enzymatic apparatus). Therefore, while the set of proteinogenic amino acids in a martian second genesis would likely be different from the terrestrial set, a certain overlap would still be expected, perhaps as much as 50–75% (Weber and Miller, 1981).
Exotic deviations in the use of molecular building blocks, such as the use of N-methyl amino acids, cannot be discounted a priori (Breslow and Cheng, 2009), although a plausible prebiotic scenario for the formation of such building blocks in high yields, or their accumulation to levels comparable with the more common α-amino acids, is still lacking (Bada, 2009). Similar arguments could be made for other molecular building blocks such as nucleobases, sugars, and carboxylic acids, the precursors of membrane lipids.
Larger biochemical differences ought to be expected between independent terrestrial and martian populations at higher order levels, such as in the structures of biopolymers assembled from the simpler units (e.g., proteins, nucleic acids, membrane lipids). This is because abiotic polymerization reactions are highly dependent on physicochemical parameters such as temperature, pressure, pH, water activity, redox state, and radiation environment, and the chemistry of solutions. Different physicochemical conditions in early Earth and early Mars would have had a different impact on the structure and composition of those larger molecules, affecting the mechanisms through which those molecules assembled and in which they participated.
Inferred biochemical traits of early terrestrial progenotes provide additional clues of other fundamental aspects where the biochemistry of martian life could diverge from that of terrestrial life, in the context of an independent genesis. Although an unlikely solution for long-term survival, the entire biochemical apparatus could be based on a single functional biopolymer that acted both as a genetic and catalytic molecule, analogous to the RNA-world, although the exact nature of that polymer cannot be anticipated. Other possible divergent traits would include the structure and composition of membrane lipids, the genetic code, and the nature of the translation machinery (if one existed) (Table 1). While speculations of the biochemical nature of martian life are an intriguing exercise, a more immediate question is whether it will be possible to discriminate between an independent genesis of life and a biological transfer, with data obtained by future life detection missions. This question is addressed in the next section. While the probability of biological transfer diminishes with distance from Earth, it is never zero (Napier, 2004; Worth et al., 2013). Therefore, many of the arguments exposed in the next section for the case of Mars would also apply to the search for evidence of life in the outer solar system.
4. Implications for the Search for Evidence of Life on Mars
Finding any evidence of life on Mars would be a momentous result. In that regard, the distinction between an independent genesis and a biological transfer might be dismissed as merely academic. However, the scientific implications of finding evidence of an independent genesis or a biological transfer would, in fact, be radically different. Given the age and size of the Universe, and the number and diversity of worlds, evidence of an independent genesis would imply that life is a common cosmic phenomenon.
A separate biology would also help to derive universal biological principles necessary to support a theory of living systems and elucidate those processes that lead from prebiotic chemical reactions to the appearance of self-replicating, evolving chemical systems. On the contrary, evidence of a biological transfer would validate theories of planetary panspermia (or perhaps even cometary panspermia) and could revolutionize our understanding of the biochemistry and evolutionary processes, but it would still leave us ignorant about how widespread life is in the Universe and with an Earth-centric view of biology. In that regard, not all evidence of life can be considered of equal value.
Conceding the possibility of a biological transfer has practical implications for how we search for evidence of life. First, it implies that not all biosignatures are of the same quality (Table 2). The ancestral relations captured in the phylogenetic tree are based on (1) comparisons of molecules whose sequence changes randomly in time such as proteins and nucleic acids or (2) the composition of cell membranes and cell walls. A biological transfer or an independent genesis can only be established or refuted through comparisons of similar biochemical traits. If the origin of life event is defined as the appearance of a self-replicating and evolving chemical system, it follows that the most unambiguous evidence of an independent genesis would be a genetic molecule that is not a precursor to, or derivative from, RNA. Other chemical biosignatures such as life's preference for certain isomers (e.g., L/D enantiomers) or the choice of monomer building blocks (e.g., the specific set of proteinogenic amino acids) could add additional discriminatory power to separate an independent genesis from a biological transfer, although there might be limitations, as noted below.
Examples of Life Detection Measurements and Their Relevance As Potential Signatures of Life and As Indicators of an Independent Genesis (Assuming Prokaryotic-Like Life)
The assessment is based on “life as we know it,” not on theoretical forms of life strongly divergent or unlike Earth-life.
++, strongly relevant; +, relevant; −, not relevant.
However, other classes of potential biosignatures, notably those that are not based on biochemical traits such as morphological features, inorganic disequilibria, or certain elemental or isotopic ratios (or even the types of biosignatures sought by the twin Viking landers, Klein et al., 1972), could be indicative of life, but would not discriminate between an independent genesis and a biological transfer.
This issue might be particularly relevant when searching for evidence of forms of life that might have existed billions of years ago, such as those that will be targeted by NASA's Perseverance or ESA's Rosalind Franklin rovers. While results from NASA's Curiosity rover demonstrate that organic matter can survive in sedimentary deposits of a similar age (Eigenbrode et al., 2018), the types of biomolecules that record phylogenetic information would likely not have survived intact for billions of years, particularly near the surface (Blanco et al., 2018). The discovery of chemical (e.g., isotopic, elemental) or structural (e.g., biofabrics, microscopic cell-like structures) biosignatures in such ancient martian sedimentary deposits would be a significant accomplishment but, in the absence of biochemical context, would afford no insight regarding the exact origin of the organisms that produced them. Such determinations might only be possible after considering multiple independent lines of evidence based on certain biochemical traits (Table 1).
Second, the possibility of a biological transfer highlights the importance of placing potential biosignatures in the right evolutionary context. In that regard, the evolutionary tree represents not only a blueprint to interpret evidence of life but also a guideline to determine the likelihood that certain biosignatures could be expressed by martian organisms. The terrestrial sequence: prebiotic chemistry→progenotes→LUCA, suggests a hierarchy of potential biosignatures based on their timing of appearance. Biological traits that appeared early in the evolutionary sequence could be said to be fundamental and more likely to be universal. Biological traits that appeared later in the sequence could be said to reflect evolutionary events on Earth, which are more likely to be idiosyncratic.
As a practical example of the 20 proteinogenic amino acids found in extant terrestrial life, those amino acids used by earlier progenotes by virtue of their prebiotic abundance (Gly, Ala, Val, Asp, Pro, Ser, Glu, Leu) would be more likely to also be used by martian organisms, irrespectively of their origin. On the contrary, biosynthetic amino acids incorporated later by terrestrial life (Arg, Asn, Cys, His, Phe, etc.) would only be expected in martian organisms in the context of a post-LUCA biological transfer. Future missions to search for evidence of life on Mars would be advised to consider such evolutionary aspects of potential biosignatures, in addition to their reliability, survivability, and detectability.
Yet, the very existence and nature of early terrestrial progenotes represent a cautionary tale for our hopes of discerning an independent genesis of life on Mars from a biological transfer. As explained in Section 3.1.1, the transfer of early progenotes could potentially have steered the organic chemical evolution on Mars along an earth-like path by contributing a still incomplete set of proteinogenic amino acids, primitive membranes, an underdeveloped translation apparatus, or an incipient genetic code. Forms of life evolving on Mars from such a rudimentary biochemical foundation could have quickly diverged, eventually leading to more evolved organisms lacking some of the universal traits common to all life on Earth. Such forms of life, if discovered today, might not even map in the phylogenetic tree of life and, therefore, might be wrongly interpreted as evidence of an independent genesis. This represents an intriguing gray area that points to a fundamental limitation in our ability to discriminate between an independent genesis of life and life that is ultimately related to us.
Footnotes
Acknowledgments
The author would like to thank Chris McKay, Dale Andersen, Marc Ditzler, Craig Everroad and two anonymous referees for their helpful comments and suggestions during the preparation of this article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Writing of this article was supported with funding from the NASA Internal Scientist Funding Model (ISFM).