
Abstract
Answering the question of whether life ever existed on Mars is a key goal of both NASA's and ESA's imminent Mars rover missions. The obfuscatory effects of oxidizing salts, such as perchlorates and sulfates, on organic matter during thermal decomposition analysis techniques are well established. Less well studied are the transformative effects of iron oxides and (oxy)hydroxides, which are present in great abundances in the martian regolith. We examined the products of flash pyrolysis–gas chromatography–mass spectrometry (a technique analogous to the thermal techniques employed by past, current, and future landed Mars missions) which form when the cyanobacteria Arthrospira platensis are heated in the presence of a variety of Mars-relevant iron-bearing minerals. We found that iron oxides/(oxy)hydroxides have transformative effects on the pyrolytic products of cyanobacterial biomolecules. Both the abundance and variety of molecular species detected were decreased as iron substrates transformed biomolecules, by both oxidative and reductive processes, into lower fidelity alkanes, aromatic and aryl-bonded hydrocarbons. Despite the loss of fidelity, a suite that contains mid-length alkanes and polyaromatic hydrocarbons and/or aryl-bonded molecules in iron-rich samples subjected to pyrolysis may allude to the transformation of cyanobacterially derived mid-long chain length fatty acids (particularly unsaturated fatty acids) originally present in the sample. Hematite was found to be the iron oxide with the lowest transformation potential, and because this iron oxide has a high affinity for codeposition of organic matter and preservation over geological timescales, sampling at Mars should target sediments/strata that have undergone a diagenetic history encouraging the dehydration, dihydroxylation, and oxidation of more reactive iron-bearing phases to hematite by looking for (mineralogical) evidence of the activity of oxidizing, acidic/neutral, and either hot or long-lived fluids.
1. Introduction
The search for evidence of past life on Mars is a key goal of both NASA's Mars2020 (Grant et al., 2018) and ESA's ExoMars 2022 (Vago et al., 2015) imminent missions.
During the late Noachian and early Hesperian (∼3.8–3.5 Ga) period of Mars, surface conditions are believed to have been much more benign than any other time in the history of the planet (Squyres and Kasting, 1994; Grotzinger et al., 2014). Volcanoes and hydrothermal processes released CO2, H2O, SO2, and other gases (Halevy and Head III, 2014) to maintain the atmosphere. These planetary gases were shielded from the stripping effects of ionizing solar radiation by the planet's magnetic field, generated by its (present-day defunct) internal dynamo (Acuña et al., 1998). In turn, the more dense atmosphere provided greenhouse warming effects and, at least intermittently, liquid surface waters (Wordsworth et al., 2013; Halevy and Head III, 2014).
Lake deposits and fluvial channels show that water, a prerequisite for all known life, was present at the martian surface at least sporadically (Grotzinger et al., 2015). The chemistry of the lake deposits in craters such as Gale shows that water was moderate in its pH and salinity and contained a potential microbial energy source in the form of iron and sulfur in multiple oxidation states (Hurowitz et al., 2017). A more dense atmosphere would have also acted to shelter surface life from the harmful effects of ultraviolet (UV) rays and ionizing radiation (Ehresmann et al., 2011). Thus, a warmer and wetter martian climate may have led to a temporal window of habitability when life could have evolved and colonized the surface environment at around the same time that life was emerging on the early Earth (Schulze-Makuch et al., 2008).
Cyanobacteria are a relatively old form of life on Earth (Walsh, 1992; Schopf, 2006; Schirrmeister et al., 2015). As such, cyanobacteria are hypothesized to be representative of the type of life (single-celled photosynthesizing protists) that may have existed on early Mars (Warren-Rhodes et al., 2006; Böttger et al., 2012), and so have been used as model organisms in numerous Mars analog studies (Warren-Rhodes et al., 2006; Böttger et al., 2012; de Vera et al., 2014; Smith et al., 2014). The remains of ancient martian communities of mat-forming microorganisms, from a more habitable period, may be preserved as stromatolites in lacustrine sedimentary layers (Cady et al., 2003).
On Earth, photosynthetic cyanobacteria are extremely stress tolerant (Navarro-Gonzalez et al., 2003; Warren-Rhodes et al., 2006; Fagliarone et al., 2020) and if martian protists have survived in the near-surface to the present, they may survive in cryptic special environmental niches (Wynn-Williams and Edwards, 2000). Microhabitats, including iron-oxide-rich sediments, pertinent to this study, may preferentially absorb UV while transmitting enough light to allow photosynthesizers to survive even to the present (Sagan and Pollack, 1974; Cockell and Raven, 2004), and high enough water activity for biological processes may still sporadically occur in isolated microhabitats due to the actions of deliquescing minerals (Davila et al., 2010, 2013), allowing for the survival of endolithic communities inside porous rocks or evaporitic deposits (Palmer and Friedmann, 1990; Cockell and Raven, 2004; Davila et al., 2010; Smith et al., 2014).
On Earth, cyanobacteria have been found to be highly adaptive microorganisms, capable of light-independent energy generation (Stal, 2012). They have even colonized the deep subsurface (detected down to 620 m) in perpetually dark, endolithic, and hypolithic environments where they use molecular hydrogen as energy (Puente-Sánchez et al., 2018; Sánchez-García et al., 2020). Similar hydrogenotrophic adaptations could have allowed communities of martian microbes, similar to cyanobacteria, and descended from ancient surface-dwelling photosynthesizers, to survive underground to the present (Nixon et al., 2013; McMahon et al., 2016).
Alongside other potential biosignatures such as stable isotopes, atmospheric gases, biosedimentary structures, and macrotextures (Summons et al., 2011), organic biomarker compounds are a target for both past habitability (e.g., as sought by Mars Science Laboratory) and life-detection (e.g., as sought by ExoMars 2022) efforts (Des Marais et al., 2008; Vago et al., 2017). The radiative, oxidative, and extremely arid martian near-surface environment is problematic for the preservation of organic matter over geological timescales (Sagan and Pollack, 1974; Cockell and Raven, 2004; ten Kate et al., 2005; ten Kate, 2010; Moores and Schuerger, 2012; Pavlov et al., 2012).
However, pertinent to this study, cyanobacterial biomarkers, especially UV-adsorbing pigments such as β-carotene, have been shown to survive well under the radiative and extremely arid conditions of the martian near-surface if shielded by host sediments (Baqué et al., 2016; Fagliarone et al., 2020), making these a pertinent target for study. The 2 m long drill on the forthcoming ExoMars Rosalind Franklin rover (Barnes et al., 2006) will allow depths to be reached that have been shielded from this harsh surface environment, hopefully removing this limiting factor.
Searches for life, and abiotic organic molecules, on Mars have so far been almost exclusively carried out by thermal extraction experiments, and have had little success (Klein et al., 1976; Biemann et al., 1977; Hecht et al., 2009; Cull et al., 2010; Blake et al., 2013; Kounaves et al., 2014), other than the detection of simple chlorinated molecules (Freissinet et al., 2015; Szopa et al., 2020), evidence of macromolecular material that appears abiological in nature (Eigenbrode et al., 2018), and recent detections of longer chained alkanes (Freissinet et al., 2019). Thermal decomposition techniques will also be applied to samples from up to 2 m below the martian surface by ExoMars (Barnes et al., 2006) as part of its life-detection strategy.
Difficulties in conclusively identifying the provenance of any surviving martian organic matter through this thermal extraction approach have been primarily attributed to the deleterious, obfuscating effects of oxidizing minerals on the analysis of martian materials, particularly perchlorates (Glavin et al., 2013; Montgomery et al., 2019) and possibly sulfates (Lewis et al., 2015). Pyrolytic products of martian organic matter are combusted, or at least partially oxidized, as they are heated in the presence of these strong oxidants (Navarro-González et al., 2010; Royle et al., 2018a). Oxidative degradation greatly reduces the structural information in the pyrolysis products detected leading to difficulty in identifying parent molecules, and thus the abiotic or biological provenance of organic structures (Freissinet et al., 2015; Eigenbrode et al., 2018; Guzman et al., 2018).
As well as the oxidizing salts, the crust of Mars has abundant iron (Taylor and McLennan, 2009), and its oxides are ubiquitous on the martian surface. Iron oxides promote pyrolytic reactions in industrial processes (Minicò et al., 2000; Gooßen et al., 2011; Bukur et al., 2016), and previous work has shown that they also can have transformatory effects on the pyrolytic products of Mars analog samples (Royle et al., 2021; Tan et al., 2021).
The nanophase, hydrated iron oxide ferrihydrite (5Fe2O3·9H2O) is believed to be widespread on the martian surface as a major component of the iron-rich amorphous soil (Dehouck et al., 2017), along with the more crystalline oxyhydroxide goethite [FeO(OH)] and the iron (II, III) and iron (III) oxides magnetite (Fe3O4) and hematite (Fe2O3). These inorganic species have been detected both in martian meteorites (Steele et al., 2007; Muttik et al., 2014) and in situ at Mars (Evans and Adams, 1980; Bell et al., 1993; Klingelhöfer et al., 2005; Morris et al., 2006a, 2006b; Bish et al., 2013; Blake et al., 2013; Treiman et al., 2016), and their relative abundances in both the martian strata and overlying regolith will depend on the stability of each phase under historic local geological conditions.
Iron oxides can preserve biosignatures through a variety of mechanisms. These include the entombment of bacteria by iron oxide crystals (Ferris et al., 1989; Peng et al., 2013), the formation of textural biosignatures by iron oxide encrustation (Potter-McIntyre et al., 2014; Williams et al., 2015b, 2017), the formation of organometallic complexes (Lalonde et al., 2012), and the preservation of organic molecular biosignatures through adsorption onto the surface of iron oxides and oxyhydroxides (Parenteau et al., 2014; Tan et al., 2018).
The end results of these preservative mechanisms can be clearly seen in a variety of aqueous environments on Earth, with almost a quarter of organic carbon in sedimentary deposits directly bound to reactive iron phases (Lalonde et al., 2012). This “rusty sink” effect promotes preservation over geological timescales (Lalonde et al., 2012). Many of these terrestrial, iron-rich settings are analogous to environments present on ancient Mars (Allen et al., 2000, 2004; Schelble et al., 2004; Fernández-Remolar et al., 2005); hence, iron-oxide-rich sediments are potential targets for future life-detection missions. It is therefore important to understand the potentiality of various iron oxides to affect the results of these life-detection efforts to allow for effective site selection.
Previously, we have shown that sediments rich in iron oxides have obfuscating effects both on the detection of organic molecular pyrolysis products in natural Mars analog samples (Tan et al., 2021) and on fatty acid standards (Royle et al., 2021). It is necessary to carry out analog experiments to determine the effect of Mars-relevant minerals on the thermal decomposition of Mars-relevant organisms as this will inform what a successful detection of life, by similar techniques, in situ at Mars could look like.
Arthrospira platensis is a well-studied and categorized cyanobacterium owing to its potential as feed-stock to produce bio-oils. Pyrolysis products of cyanobacteria have been shown to be significantly altered in the presence of various industrial catalysts (Chagas et al., 2016; Zhao et al., 2019). Here, we pyrolyze the cyanobacteria A. platensis in the presence of a range of Mars-relevant iron oxide/(oxy)hydroxide species to examine the modification of pyrolysis products, and therefore what could be expected to be detected by in situ thermal decomposition experiments at Mars.
2. Materials and Methods
2.1. Sample preparation
Organic dried A. platensis powder was obtained from Naturya. Quartz sand (US Silica F-35) was used as a relatively inert substrate. Magnetite, hematite, and goethite were obtained from Sigma-Aldrich. Due to its instability, ferrihydrite had to be produced in the laboratory.
Ferrihydrite was synthesized following the method detailed in Cornell and Schwertmann (2003). Eight grams of Fe(NO3)·9H2O (suitable for cell culture; Sigma-Aldrich) was dissolved in 100 mL of deionized water and stirred with a magnetic stirrer. To this solution, 66 mL of 1 M potassium hydroxide (KOH) solution was continuously added to bring the pH of the resulting mixture to 7.5. The final volume of KOH solution was added dropwise, and the pH continuously monitored. Dissolved salts were removed by adding deionized water, centrifuging and decanting the supernatant until the conductivity was <10 μS/cm as measured with a total dissolved solids probe. The synthesized ferrihydrite was then freeze-dried, and ground gently in a solvent-cleaned agate pestle and mortar.
X-ray diffraction (XRD) analysis [presented in Royle et al. (2021)] confirmed the produced ferrihydrite to be of the amorphous/least crystalline 2-line ferrihydrite, as opposed to the more (but still poorly) crystalline 6-line ferrihydrite, based on the number of broad peaks (2 or 6) exhibited in the XRD pattern (Cornell and Schwertmann, 2003; Kukkadapu et al., 2003). As ferrihydrite is known to recrystallize at room temperature, synthetic samples were wrapped in aluminum foil and stored at 4°C.
Quartz sand was cleaned by furnacing overnight at 450°C to remove any organic matter contamination. Due to their susceptibility to temperature, the iron oxides magnetite, hematite, and goethite had to be cleaned through solvent extraction rather than heating. Cleaning was carried out by using the standard liquid extraction protocol (Emmerton et al., 2012) as follows: 1–3 g of powdered sample was suspended in a 93:7 v/v DCM/MeOH solution and agitated in a sonic bath, the resulting suspension was then centrifuged, and the supernatant solution pipetted off. This process was repeated three times, and the materials were dried under a stream of nitrogen. As the ferrihydrite was produced in the laboratory by using clean materials and methods and subsequently stored clean, it was deemed unnecessary to clean this highly reactive mineral further.
The cleaned substrate minerals were dry mixed with the A. platensis powder to make samples that were 0.5, 1, 5, and 10 wt % A. platensis powder. Samples were stirred to ensure mixtures were homogeneous. Pure A. platensis and unspiked substrate minerals were also analyzed as control samples.
2.2. Pyrolysis gas chromatography–flame ionization detection and gas chromatography–mass spectrometry analysis
Compound identification was carried out with pyrolysis–gas chromatography–mass spectrometry (Py-GC-MS). Quantification of produced hydrocarbons was carried out by pyrolysis–gas chromatography–flame ionization detection (Py-GC-FID).
For Py-GC-MS analysis, biomass/mineral mixtures were used with concentrations of A. platensis (with total mass in parentheses) at 0.5% (40 ± 4 mg), 1% (40 ± 4 mg), 5% (10 ± 0.1 mg), 10% (5 ± 0.1 mg), and 100% (0.5 ± 0.1 mg). For Py-GC-FID, due to the increased sensitivity of the detector (compared with GC-MS) and improved quantitative comparability between samples, all mixtures were loaded at total masses of 5–10 mg. All samples were run in replicate.
For both Py-GC-MS and Py-GC-FID, individual samples were placed inside the platinum coil of a CDS Analytical 2000 Pyroprobe under helium carrier gas, and heated at 20°C/ms to 650°C and held for 15 s. Separation of products was achieved by using an Agilent Technologies 6890 gas chromatograph coupled through a CDS 1500 interface held at 300°C. The GC inlet was held at 270°C and operated in split mode (10:1) with a column flow rate of 1.0 mL/min. Separation was performed on a J&W DB-5 column (30 m × 250 μm × 0.25 μm). The GC oven was held for 2 min at 40°C and then ramped at a rate of 5°C/min to 310°C where it was held for 10 min.
For qualitative analysis and compound identification, the GC was coupled to an Agilent Technologies 5973 MS. Mass spectra were acquired over a range of m/z 45–550. Identification of peaks in the produced chromatograms was carried out by using the Agilent MSD Chemstation software package, and peak identification was achieved by comparison with the NIST database, published literature and standards. For semiquantitative analysis, the FID detector was used instead of the MS; other conditions of the method were kept identical. Py-GC-FID peaks were identified through comparison with the corresponding Py-GC-MS data.
3. Results
3.1. Pyrolysis–gas chromatography–mass spectrometry
Figure 1 shows the total ion current (TIC) chromatograms produced by pyrolyzing 0.5 mg of dried A. platensis at 650°C.

TIC chromatogram of 0.5 mg of dried Arthrospira platensis powder pyrolyzed at 650°C. TIC = total ion current.
The main identifiable compounds produced from the pyrolysis of A. platensis were consistent with other studies (Salter et al., submitted; Chagas et al., 2016). Aromatic hydrocarbons toluene and styrene (along with n-alkylbenzenes up to butylbenzene); nitrogen-heterocycles and other nitrogen-bearing moieties (including pyridine, pyrrole, benzonitrile, nitriles, and amides); phenols and other nonaromatic compounds: dominated by heptadecane, 2,6,10-trimethyl,14-ethylene-14-pentadecane (neophytadiene), and the fatty acids n-hexadecanoic acid and 9,12 octadecadienoic acid (tentative identification of double bond positions).
Figures 2–5 show the TICs of A. platensis pyrolyzed on quartz and the iron oxide/(oxy)hydroxide substrates at varying concentrations, with the detectable pyrolysis products summarized in Table 1.

TIC chromatograms of Arthrospira platensis mixed into various substrates at 10 wt % and pyrolyzed at 650°C. Red molecules indicate pyrolysis products not detected from A. platensis pyrolyzed by itself (alteration products). Color images are available online.

TIC chromatograms of Arthrospira platensis mixed into various substrates at 0.5 wt % and pyrolyzed at 650°C. Red molecules indicate pyrolysis products not detected from A. platensis pyrolyzed by itself (alteration products). Color images are available online.
Summary of Identifiable Compounds from Arthrospira platensis Pyrolyzed on Various Substrates at Differing Concentrations
PAH = polyaromatic hydrocarbon.
At 10 wt % A. platensis (Fig. 2) pyrolysis products were similar from all substrates to the products of 100% A. platensis. This suggests that 10 wt % A. platensis provided enough organic matter to overwhelm any surface effects from the mineral substrates. Exceptions to this, evidencing some mineral surface effects, were the production of new aromatic products from organic transformations induced by goethite, magnetite, and ferrihydrite. These new products were dominated by the two-ring structures naphthalene and biphenyl with minor acetophenone. There were also tentative detections of up to four-ring structures from goethite. Heptadecane was notably reduced, relative to the other products, in ferrihydrite while
At 5 wt % A. platensis (Fig. 3) pyrolysis products were still superficially similar to the products of 100% A. platensis. No major products were gained or lost from A. platensis pyrolyzed at 5 wt % on the quartz substrate. Only minor changes were observed in the products on hematite; notably alkane production was increased, and the new aromatics naphthalene and biphenyl were now produced in detectable amounts. The limited transformational effects of hematite on organic detection by Py-GC-MS are consistent with previous work, which showed that the presence of this iron oxide in sediments, from Mars analog environments, did not appear detrimental to organic matter detection by pyrolytic techniques (Lewis et al., 2018).
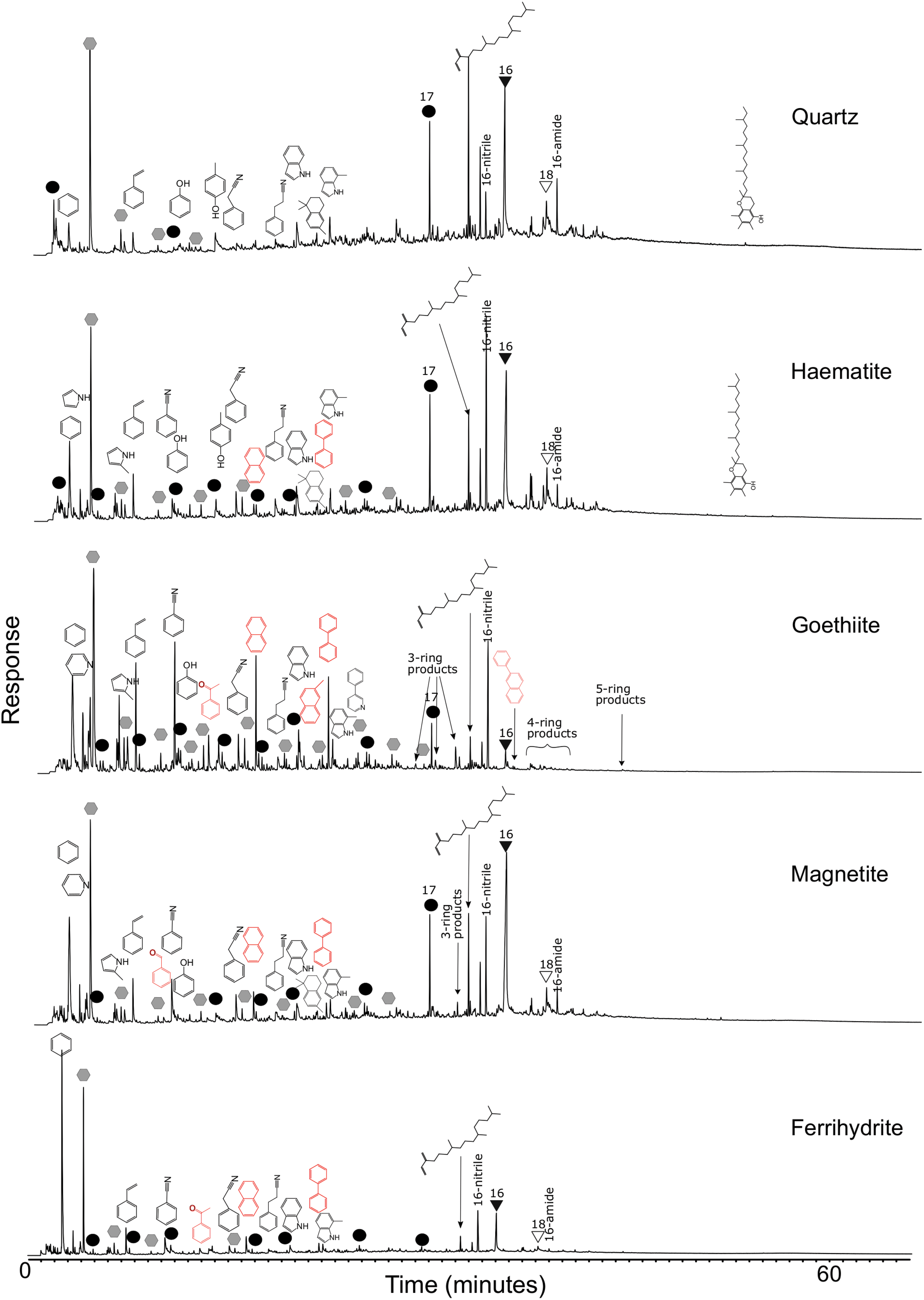
TIC chromatograms of Arthrospira platensis mixed into various substrates at 5 wt % and pyrolyzed at 650°C. Red molecules indicate pyrolysis products not detected from A. platensis pyrolyzed by itself (alteration products). Color images are available online.
A. platensis on goethite exhibited the greatest deviation from A. platensis pyrolyzed alone. There was a major reduction in longer chain products, particularly fatty acids, while aromatic moieties were significantly increased with polyaromatic hydrocarbons (PAHs) with up to five rings tentatively detected. This detrimental effect of goethite is also in line with the findings of the aforementioned work by Lewis et al. (2018) who reported a dominance of aromatic moieties in the pyrolytic products of goethite-rich sediments, although they inferred this to be an effect of preservation rather than alteration during analysis. All products appeared reduced for A. platensis on ferrihydrite with the effect more pronounced with the longer chain length compounds.
At 1 wt % A. platensis (Fig. 4) pyrolysis products were little changed with quartz or hematite substrates, although the lowest abundance compounds (e.g.,

TIC chromatograms of Arthrospira platensis mixed into various substrates at 1 wt % and pyrolyzed at 650°C. Red molecules indicate pyrolysis products not detected from A. platensis pyrolyzed by itself (alteration products). Color images are available online.
Trends observed from 5 wt % to 1 wt % were generally continued down to 0.5 wt % A. platensis, with a reduction in variability and abundance of products from hematite, most notably a loss of the fatty acids and few products detectable on goethite, magnetite, or ferrihydrite.
3.2. Pyrolysis–gas chromatography–flame ionization detection
Py-GC-FID analysis allows the direct comparison of the abundances of different pyrolysis products, with a linear response over a large range for all hydrocarbon species and no detection preferences (in contrast to GC-MS), allowing for semiquantitative analysis. As the pyrolysis and GC conditions were identical, the Py-GC-FID chromatograms produced were highly similar to Py-GC-MS TIC chromatograms (although responses were greater due to the increased sensitivity of the GC-FID) so are not shown. To allow quantitative comparison between Py-GC-FID data peak areas were normalized to the weight of dried A. platensis that would have been available to be pyrolyzed in each sample:
The plots in Fig. 6 show the normalized peak area of Py-GC-FID responses (average values from replicates) from selected products of interest formed when A. platensis was pyrolyzed at varying concentrations on quartz and the iron oxide/(oxy)hydroxide substrates.
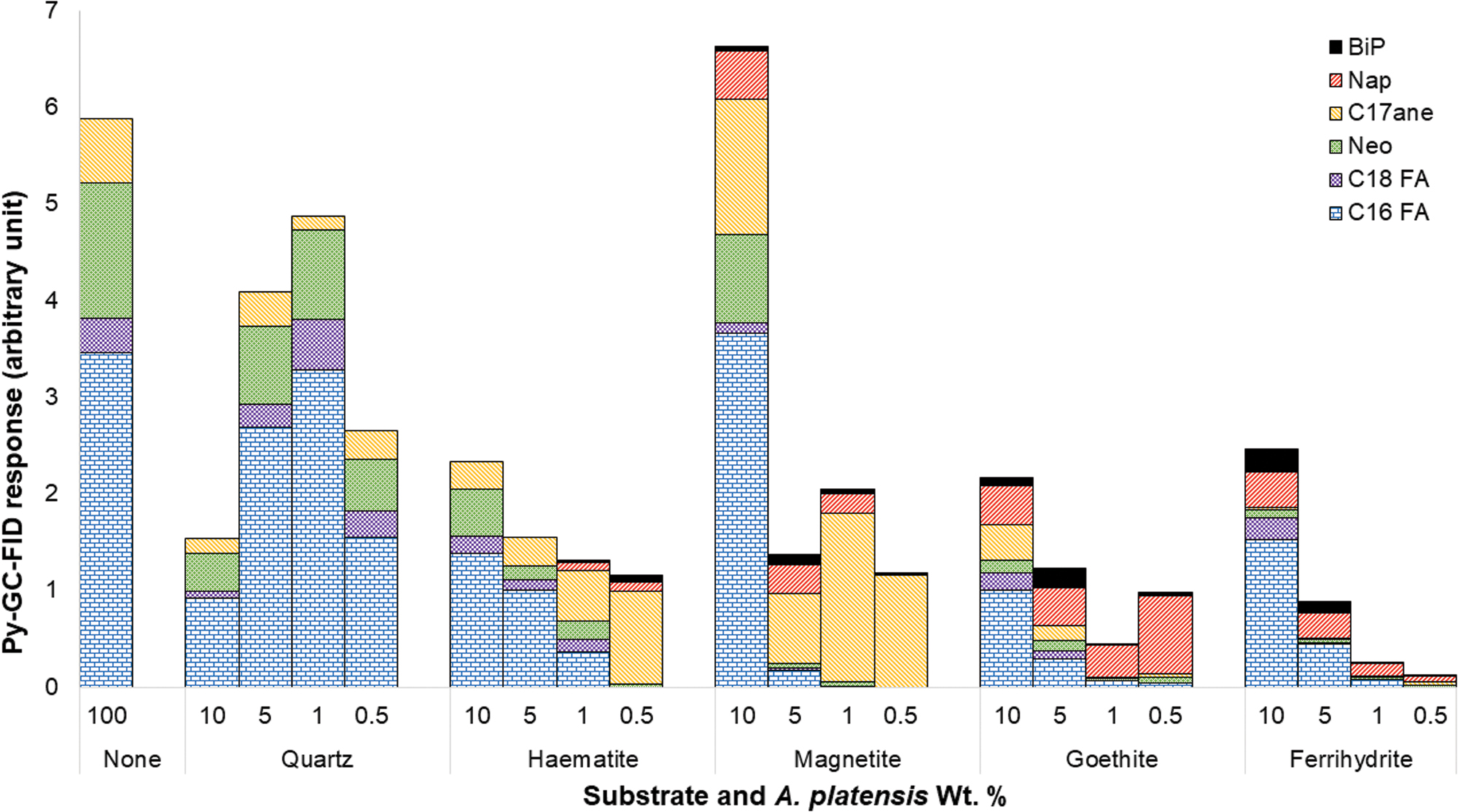
Py-GC-FID responses from peaks of interest, peaks identified using retention times from Py-GC-MS data collected under identical GC conditions. Peak areas normalized to both sample weight and Arthrospira platensis content of sample. Bip = biphenyl; C16 FA = hexadecanoic acid; C17ane = heptadecane; C18 FA = 9,12 octadecadienoic acid; Nap = naphthalene; Neo = neophytane; Py-GC-FID = pyrolysis–gas chromatography–flame ionization detection; Py-GC-MS = pyrolysis–gas chromatography–mass spectrometry. Color images are available online.
The data show that the relative abundances of products varied with differing substrates, and in most cases concentrations, even after normalization for the amount of A. platensis that was available for pyrolysis in the different samples. No trends were observed for any of the pyrolysis products of A. platensis on the quartz substrates as A. platensis concentration was decreased. Relative abundances of the pyrolysis products on quartz were equal to those produced from pure A. platensis. This suggests that a quartz substrate has little measurable effect on the breakdown of A. platensis and the detectability of produced compounds.
In contrast, all four of the iron oxide/(oxy)hydroxide substrates tested exhibited a general trend of decreasing total products with decreasing A. platensis concentration; primarily resulting from a loss of the fatty acids 9,12 octadecadienoic acid and hexadecanoic acid and, to a lesser extent, neophytadiene. While substrate specific, the relative, and in some cases overall, abundances of naphthalene and biphenyl increased with decreasing A. platensis content. Heptadecane abundance trends appear to be much more substrate specific with similar behaviors in hematite/magnetite and goethite/ferrihydrite pairings.
At 10 wt % A. platensis, a hematite substrate did not have a notable effect on the pyrolysis products (compared with pure A. platensis or a quartz substrate), and at 5 wt % no new products were observed; however, there was a relative increase in heptadecane at the expense of the other products. This trend of increasing heptadecane response (relative to the two fatty acids and neophytadiene) was further enhanced as biomass was decreased, until heptadecane was by far the main product. At 1 wt %, the aromatic hydrocarbons naphthalene and biphenyl began to be produced in detectable quantities, and their relative abundances increased down to 0.5 wt % A. platensis. At 0.5 wt % fatty acid abundances also dropped below detection limits, and neophytadiene was much reduced. The pyrolysis products from A. platensis pyrolyzed on magnetite exhibited a continuation of the same trends of alteration processes as those that occurred on hematite. Specifically, abundances of heptadecane and aromatic moieties were increased, at the expense of fatty acids and neophytadiene, as biomass was decreased. Naphthalene and biphenyl were already detectable in significant quantities at 10 wt % A. platensis on magnetite, and these aromatic hydrocarbons, along with heptadecane, already dominated by 5 wt %.
Pyrolysis products of A. platensis on both goethite and ferrihydrite displayed a greatly increased dominance of the aromatic species naphthalene and biphenyl at the expense of the nonaromatic products. The increase of aromatic moieties at the expense of nonaromatics increased with decreasing A. platensis content. Naphthalene production at the lowest A. platensis concentrations on goethite was particularly enhanced.
Of note is that heptadecane is the only pyrolysis product formed from pure A. platensis (and A. platensis on quartz), which was proportionally increased by the presence of iron oxides. This effect was greatest on magnetite, where little else was detectable at the lowest concentrations (0.5 and 1 wt % A. platensis). Data for the hematite substrate also display an increased response for heptadecane with poor responses for other compounds at 0.5 wt % A. platensis. The relative increase of heptadecane to fatty acid production, with decreasing A. platensis concentration, was less pronounced on a goethite substrate as the increases of naphthalene and biphenyl dominated. Ferrihydrite was the only substrate where significant decreases in the relative heptadecane abundance were observed.
4. Discussion
4.1. Pyrolysis products of A. platensis as potential biomarkers
The sources of the main pyrolytic breakdown products of A. platensis and other cyanobacteria have been well established by previous studies as shown in Table 2.
Biomolecules and Their Expected Pyrolytic Breakdown Products
Many of these detected compounds are of no use as biomarkers as they may form abiotically, and so cannot be unambiguously ascribed to a biological source, for example, aromatic compounds and nitrogen heterocycles.
However, isoprenoid hydrocarbons, such as diterpenoid neophytadiene and
Fatty acids may also be useful biosignatures as they are also relatively chemically recalcitrant, persisting over geological timescales (Summons et al., 2008) even under harsh, Mars-like conditions if sufficiently shielded from UV (Stalport et al., 2010). In terrestrial biosignatures, fatty acids with a distribution around C16-C17 are generally taken to be indicative of a microbial source, C18-C22 of algae and >C23 of higher plants (Cranwell, 1974; Tissot and Welte, 1978; Perry et al., 1979; Bianchi, 1995). Biotic n-fatty acids generally display a strong even-over-odd carbon number due to their extension through the addition of two methylene (C2H4 or CH2-CH2) units (Hartgers et al., 2000; Summons et al., 2008; Georgiou and Deamer, 2014).
Unsaturated longer chain fatty acids may indicate the presence of recent (extant) life (O'Leary, 1962; Volkman et al., 1989; Alfaro et al., 2006) but they are highly labile, undergoing reactions at the sites of their double carbon bonds (through microbial degradation, autoxidation with the atmosphere or other diagenetic processes) to quickly hydrogenate to saturated fatty acids, which are more stable over geological timescales (Parker and Leo, 1965; Matsuda and Koyama, 1977; Van Vleet and Quinn, 1979; Haddad et al., 1992). Chain length distribution patterns, however, are more resistant to alteration (Duda et al., 2018). Further to this, abundances of specific fatty acids may be indicative of certain taxa (O'Reilly et al., 2017). The C18:2 fatty acids, as detected here from A. platensis (along with other states of polyunsaturation), are commonly used as a biomarker for cyanobacteria in modern terrestrial sediments (Allen et al., 2010; Willers et al., 2015; Sánchez-García et al., 2020).
4.2. Effect of various mineral substrates on detectability and transformation of organic compounds
Pyrolyzing A. platensis on a quartz sand substrate had no observable effect compared with pyrolyzing A. platensis by itself. There were no notable changes in relative proportions of products, nor a decreasing trend of product total abundance (as concentration of A. platensis was decreased). The relative inactivity of quartz is consistent with the findings of previous work where individual fatty acids were pyrolyzed on a quartz substrate (Royle et al., 2021). However, the less volatile fatty acids and high molecular weight compounds were inferred to recondense onto the quartz surfaces, lowering detectability and thus increasing the relative abundance of lower molecular weight compounds (Royle et al., 2021), which we observe no clear evidence here.
When A. platensis was pyrolyzed in decreasing concentration on any iron oxide/(oxy)hydroxide substrate, we observed that (i) total abundance of products was decreased (even after accounting for the variable biomass available for pyrolysis within each sample); (ii) the relative distribution of pyrolysis products was altered; and (iii) there was an increasing relative abundance of new “alteration/transformation” products. The increase of these effects with decreasing A. platensis is likely due to the higher availability of reactive surfaces for released pyrolysis products to encounter and undergo mineral surface effects.
Under the experimental pyrolysis conditions, when heated in the presence of organic carbon, all of the iron oxide/(oxy)hydroxides used in this study are themselves altered. XRD data demonstrating this are shown and discussed at length in Royle et al. (2021). In short, hematite is reduced to magnetite; magnetite is oxidized to hematite; goethite is dehydroxylated to hematite, which is further partially reduced to magnetite; ferrihydrite is dehydrated to hematite with further minor reduction to magnetite (Carlson and Schwertmann, 1981; Gualtieri and Venturelli, 1999; Cornell and Schwertmann, 2003; Cudennec and Lecerf, 2006).
Dehydration, dehydroxylation, and recrystallization of these phases to hematite and magnetite would have produced many free radical H• and OH• ions as water and hydroxyl ions react with free Fe atoms on the crystal surface and dissociate (Stanjek and Weidler, 1992; Cornell and Schwertmann, 2003). The water gas shift reaction, also catalyzed by iron oxide surfaces, is a further source of H• radicals and OH• ions (Bukur et al., 2016). These radicals would have increased the rate of bond scission and hydrogenation in hydrocarbon chains, leading to the decreased total abundance of detectable molecules, loss of functional groups and increased saturation state of products (i.e., loss of fatty acids), and increased relative abundance of lower molecular weight products. These effects all increase as A. platensis concentration is decreased on the iron oxide/(oxy)hydroxide substrates. Free radical generation in this manner, during the reduction of iron oxides/(oxy)hydroxides, is recognized as an astrobiologically relevant method of organic matter oxidation in Fenton-like reactions (Foustoukos and Stern, 2012; Shuai et al., 2019).
The moieties most affected by the presence of the iron oxide(oxy)hydroxide substrates are the fatty acids, as these go from being from some of the most relatively abundant pyrolysis products to complete or almost complete loss as A. platensis content is decreased. Previous work by this group (Royle et al., 2021; Tan et al., 2021) has demonstrated that the presence of the same iron oxide/(oxy)hydroxide mineral substrates used here greatly promoted the cyclization, aromatization, and oxidative polymerization of fatty acids. This suggests that the main alteration products detected from A. platensis pyrolyzed on the iron oxide/(oxy)hydroxide substrates originate from fatty acid parent molecules.
Aromatic hydrocarbons (such as naphthalene and tentative detections of up to five-ring moieties) and aryl-aryl–bonded moieties biphenyl (and tentative phenylnaphthalene) are known iron oxide/(oxy)hydroxide transformation products of fatty acids (Royle et al., 2021). Heptadecane is also likely a decomposition product of C18 fatty acid(s), and it appears that its pathway of formation is promoted by the presence of certain iron oxides as its relative abundance was observed to increase as surviving fatty acids decreased. These three products are formed from the fatty acids through different alteration processes, and Fig. 7 shows that different iron oxide/(oxy)hydroxide substrates promote these to differing extents. Figure 8 illustrates a cartoon reaction series of the breakdown of fatty acids over reactive iron surfaces through the various processes.

Tri-plots of total fatty acid and the main secondary breakdown products of fatty acids (naphthalene, biphenyl, and heptadecane) produced from Arthrospira platensis pyrolyzed on the range of substrates to show varying contributions of different alteration/breakdown processes (aromatization, oxidative coupling, and decarboxylation) on different mineral substrates. Color images are available online.
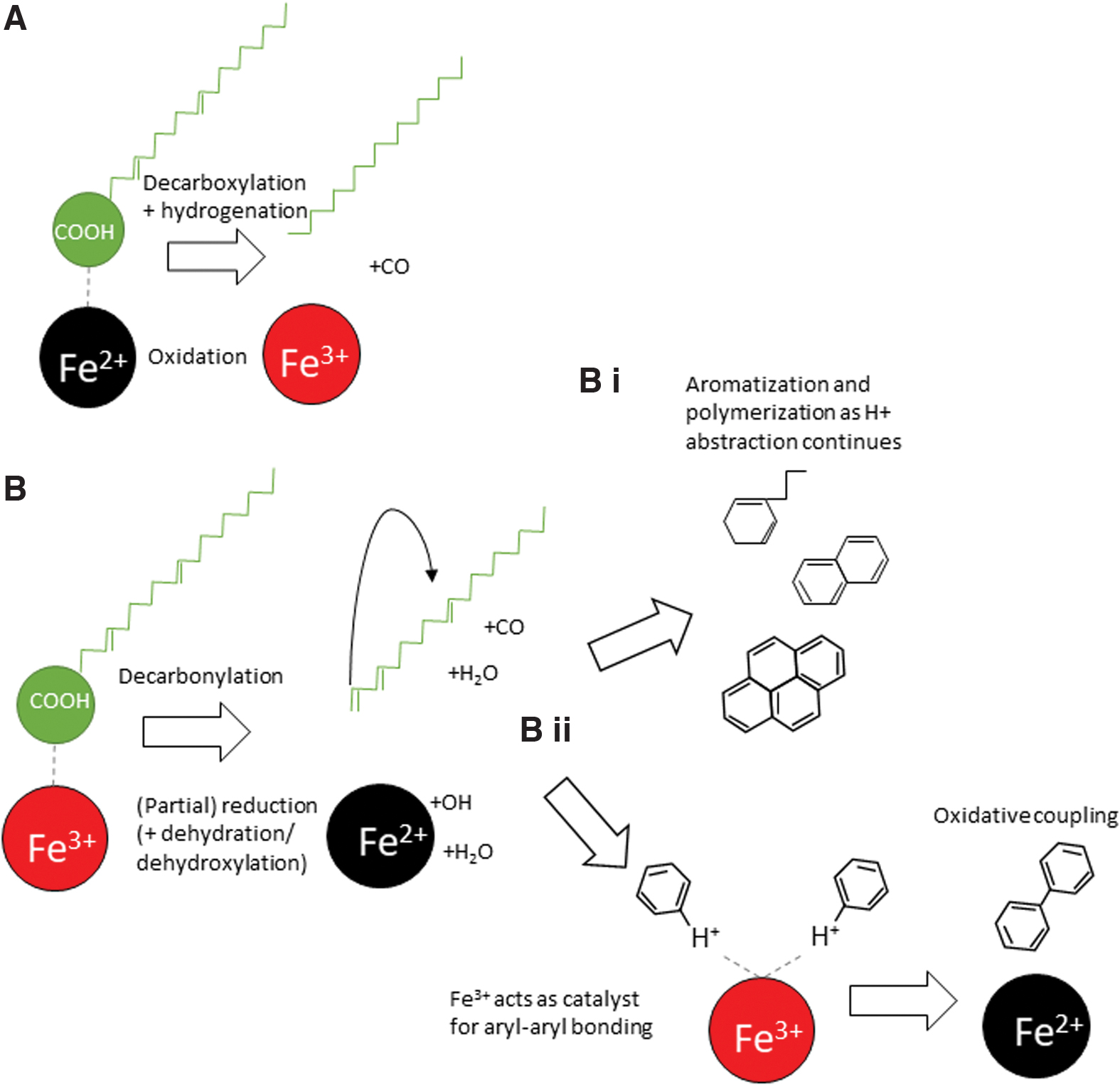
Cartoon showing proposed reaction schemes for main fatty acid alteration products on the iron oxide(oxy)hydroxide substrates. (
Fatty acids readily lose their functional group by decarbonylation/decarboxylation to produce a C n -1 alkene/alkane. This is promoted by the presence of iron oxide surfaces as fatty acids may act as negatively charged ions as the polar carboxyl group is attracted to the positive charge of free Fe atoms at the crystal surface (Wainipee et al., 2010; Chernyshova et al., 2011) and so is likely the first step in all observed iron-catalyzed transformation of the fatty acids. Decarbonylation involves the loss of a carbon and oxygen and tends to only occur at lower temperatures; decarboxylation is the loss of the carboxyl group (forming CO2) and the addition of a proton to the remaining alkene/alkane (Hartgers et al., 1995). Decarbonylation produces a terminal alkene, while decarboxylation leaves a hydrocarbon with a saturated terminal carbon. Terminal alkenes may be secondarily hydrogenated to alkanes from water–gas shift and hydrogenation reactions (Lopez-Ruiz and Davis, 2014). The observed heptadecane may be formed by decarboxylation of C18 fatty acids and subsequent hydrogenation of internal double bonds (Asomaning et al., 2014a, 2014b); this is the dominant process on the magnetite substrate (Figs. 7 and 8A). The Fe2+ oxidizes to Fe3+, forming hematite (Royle et al., 2021) as it provides H+ ions to promote both chain scission and hydrogenation. Heptadecane is not necessarily only the product of C18 fatty acids, but it may also be produced from C16 fatty acids, which have lost their carboxyl group by decarbonylation, generating a terminal alkene and alkyl radicals and subsequently undergoing C–C radical coupling until termination by hydrogen saturation (Huang et al., 2020).
A reduction of heptadecane on the goethite and ferrihydrite substrates suggests that any long chain alkenes or alkanes produced from fatty acids are further altered by subsequent transformative processes. Due to the produced terminal alkene and alkyl radicals, oxidatively decarbonylated fatty acids cyclize and wrap up on themselves. This is especially promoted if the parent fatty acid is (poly)unsaturated, as the double bonds attach to other double bonds; so the C18:2 fatty acids are especially susceptible (Hartgers et al., 1995; Royle et al., 2021). This process of cyclization and aromatization forms the observed alkylbenzenes and progresses to form benzene rings, which polymerize to naphthalene and PAHs (Fig. 8B-i). Goethite has the strongest effect on aromatization, followed by ferrihydrite (Fig. 7); this suggests that aromatization is promoted both by Fe3+ scavenging H and by the production of OH• (from the dehydration/dehydroxylation of the iron (oxy)hydroxide), which enhances chain scission.
Biphenyl and other aryl-bonded moieties are formed by oxidative coupling (Fig. 8B-ii). In this process, Fe3+ surface ions act as electron acceptors, forming aryl free radicals, which are then able to react with other molecules to form dimers (e.g., biphenyl) and other higher molecular weight polymers through oxidative coupling reactions (Grzybowski et al., 2013; Watson and Sephton, 2015).
The initial oxidation of organic matter by ferric iron is simply a contact–contact interaction; either solid, liquid, or gas phase organic compounds would be oxidized and aromatized by the process (Olcese et al., 2013). However, oxidative coupling is observed as a secondary process to aromatization on goethite, ferrihydrite, and hematite (Fig. 7), as oxidative coupling must occur at the crystal surface, whereas aromatization may also occur in the gas phase (if promoted by OH, H2O release).
4.3. Pyrolysis products of A. platensis as potential biomarkers in the presence of iron oxides
The cyanobacterial biomarker C18:2 fatty acid(s) is too readily altered to be of any use as a diagnostic biomarker in the presence of iron oxides/(oxy)hydroxides. The saturated hexadecanoic acid has a greater survivability; it is originally present at higher concentrations in the pyrolytic breakdown products of A. platensis, and saturated fatty acids are less susceptible to iron-oxide-promoted aromatization (Hartgers et al., 1995). A solo detection of a C16 fatty acid is highly indicative of a microbial source, C16 is the most common fatty acid in the biosphere, and it is statistically improbable that abiotic processes could produce this isomer in isolation (Sephton et al., 2018). Abiotic (e.g., hydrothermal) processes yield primarily shorter length molecules and decreasing amounts of fatty acids of longer chain lengths to produce an Anderson–Schulz–Flory type distribution, typical of polymerization processes, where an alkyl chain grows probabilistically by bonding with additional methylene groups verses chain terminating moieties (McCollom and Seewald, 2007; Smith et al., 2013).
Despite this, a solo fatty acid is not a biomarker in the strictest sense as it cannot be definitively identified as having a biological source, although it may be a useful biosignature. The increased loss of unsaturated fatty acids from the products of A. platensis also has the effect of artificially “maturing” the sample. Unsaturated fatty acids indicate the presence of recent (extant) life (O'Leary, 1962; Volkman et al., 1989; Alfaro et al., 2006), and under normal diagenetic conditions they will hydrogenate to saturated fatty acids (through microbial degradation, autoxidation with the atmosphere, or other diagenetic processes), which are more stable over geological timescales (Parker and Leo, 1965; Matsuda and Koyama, 1977; Van Vleet and Quinn, 1979; Haddad et al., 1992). Iron oxide-enhanced hydrogenation on pyrolysis thus has the effect of mimicking this biomarker “fossilization” process, even for extant microbial populations.
As with the solo C16 fatty acid, a detection of heptadecane without a corresponding series of shorter chain alkanes is a strong indicator of a biological process selecting for a specific carbon number (Sephton et al., 2018), yet cannot be described as a true biomarker. Heptadecane is in the range of aliphatic hydrocarbons that could be indicative of algal/microbial activity and has been used to indicate cyanobacterial activity by other studies (Dembitsky et al., 2000).
The main alteration products naphthalene and biphenyl are highly ambiguous. They retain no structural information about their parent molecule (believed to be C16 and C18 fatty acids in this case) and are formed abiotically by numerous processes. Naphthalene, biphenyl, and higher order PAHs and aryl-aryl–bonded polymers are quite representative of macromolecular, kerogen-like, organic matter. This may form by multiple condensation and polymerization processes, from a variety of abiotic precursors, with no biological input, due to hydrothermal, magmatic, radiative, or pressure-induced processes (Cherchneff et al., 2011; Chanyshev et al., 2017; Steele et al., 2018) which are promoted by iron oxides (Watson and Sephton, 2015). This is quite representative of the material that makes up the insoluble fraction of meteoritic organic carbon (meteoritic macromolecular carbon), which is completely abiotic (Sephton et al., 2004; Pizzarello, 2006; Matthewman et al., 2013).
Despite these ambiguities, a suite containing mid-length alkanes (such as heptadecane) and PAHs and/or aryl-bonded molecules in pyrolyzed samples, which also have a high iron oxide content, may allude to the transformation of cyanobacterially derived mid-long chain length fatty acids (particularly unsaturated fatty acids) originally present in the sample. This “transformation” series would not be enough for a conclusive detection of life, but it may be enough to justify further geochemical analysis of the sampled target with more time/energy/resource-intensive techniques, for example, carefully targeted heating steps or derivatization pyrolysis-GC-MS (see 4.4).
Neophytadiene, and by extension other isoprenoid hydrocarbons for other photosynthetic microbial taxa, may be the best option for biomarker detection in iron-oxide-rich substrates. These seem to have the greatest survivability (compared with other potential biomarker molecules) being detectable in all samples except magnetite and ferrihydrite ≤0.5 wt % and goethite ≤1 wt % A. platensis. They also have low ambiguity, with neophytadiene being a biomarker for photosynthetic organisms and other isoprenoid hydrocarbons indicative of other biological processes (Dembitsky et al., 2000; Brocks and Pearson, 2005; Peters et al., 2007).
Pigments such as β-carotene are produced for UV protection, and so are likely to be important for the survival strategy of any near-surface-dwelling (and therefore accessible to any Mars life-detection attempts) organisms (Wynn-Williams and Edwards, 2000). Therefore, this class of cyanobacterially produced biomolecules have been identified as a specific target for life-detection efforts at Mars (Wynn-Williams and Edwards, 2000; Böttger et al., 2012; Rull et al., 2017). They are highly recalcitrant and could survive well under certain martian near-surface conditions, including in the presence of iron oxides (Baqué et al., 2016).
Therefore, the detection of the diagnostic pyrolysis product of β-carotene, α-ionene, down to A. platensis concentrations of 0.5 wt % in quartz and hematite and 1 wt % in magnetite substrates is highly promising for thermal decomposition-based life-detection efforts at Mars. Over time through diagenetic processes, the original precursor biomolecules may undergo reduction, elimination, and aromatization to lose functional groups and become saturated. However, the resultant hydrocarbon skeletons tend to be both geologically stable and indicative of specific biological pathways, that is, good biomarkers (Brocks and Pearson, 2005). This combination of recalcitrance over time and resistance to iron oxide/(oxy)hydroxide-mediated transformation during analysis by thermal degradation makes isoprenoid hydrocarbons a promising target even in high iron oxide substrates at Mars.
4.4. How iron oxides may be affecting the search for life on Mars
A microbial population surviving in the (accessible) near-surface environment on Mars up to the present day is not outside the realms of possibility. Cryptic environmental niches (Klein, 1998) may still be occupiable by highly stress tolerant, “cyanobacteria-like” microorganisms. Populations of cyanobacteria persist on Earth even in the harshest desert environments. Alongside evolutionary stress-response adaptations to UV, desiccation, extremes of temperature, pH, and high salinity (Navarro-Gonzalez et al., 2003; Tamaru et al., 2005; de Vera et al., 2014; Biller et al., 2015; Singh, 2018), microbial communities have further increased their tolerance to extreme environments by retreating to endolithic microhabitats.
Communities of photosynthetic extremophiles (including cyanobacteria) have been found inhabiting grain boundary surfaces inside porous sandstones (Palmer and Friedmann, 1990), inside weathered volcanic glasses (Herrera et al., 2009; Cavalazzi et al., 2012), impact-shocked and altered crystalline rocks (Cockell et al., 2002), and inside evaporitic rocks—including under salt crusts (Davila et al., 2010; Finstad et al., 2017) and even inside fluid inclusions in salt crystals (Conner and Benison, 2013). With particular relevance to this study, it may be the UV-adsorbing properties of Fe-rich minerals that allow some of these localities to be habitable (Cockell and Raven, 2004).
All of the aforementioned endolithic environments certainly exist at Mars, providing a wide range of potentially habitable (although not necessarily inhabited) (Cockell et al., 2012) niches, accessible to current rover life-detection sampling technology. For example, weathered sandstones are common among the aeolian (Blake et al., 2013; Silvestro et al., 2021) and fluviodeltaic (Grotzinger et al., 2014, 2015) sedimentary deposits of Mars. Weathered volcanic basalts and their alteration products are widespread at the surface of Mars (Wyatt and McSween, 2002; Ruff and Christensen, 2007; Horgan and Bell, 2012). Extensive evaporitic layers, a predicted product of evaporating martian lacustrine environments (e.g., crater lakes) (Newsom et al., 2001), have not yet been detected, although at the small scale, evaporite minerals (such as halite) have been detected and interpreted as cements in bedrock (Thomas et al., 2019).
The water necessary for life to persist in these putative martian endolithic communities may be sourced from adsorbed water onto aluminosilicates (Zent et al., 2001; Jänchen et al., 2006, 2007, 2009; Nikolakakos and Whiteway, 2018), deliquescent salts (Schulze-Makuch et al., 2008; Davila et al., 2010, 2013; Martín-Torres et al., 2015), or by the porous rock surfaces themselves (Palmer and Friedmann, 1990). In the near-subsurface, the presence of highly deliquescent oxychlorine salts may greatly increase the length of time that water is meta-stable under martian near-surface conditions (Gough et al., 2011; Martín-Torres et al., 2015), especially if in contact with subsurface ice (Fischer et al., 2014), and this could be crucial for the long-term survival of any microbial communities (Oren et al., 2014; Jones, 2018).
Also, Paleoarchean molecular fossils on Earth give some hope that, if shielded from the degradative effects of solar and cosmic radiation, fossil organic lipid biosignatures from putative martian microbes, extinct since the more hospitable late Noachian-early Hesperian period, may persist to the present day. If these exist, they are likely to concentrate in lacustrine sedimentary layers (such as those identified/inferred in the ancient crater lakes at the Gale (Grotzinger et al., 2014), Jezero (Schon et al., 2012), and Oxia Planum (Quantin-Nataf et al., 2021) rover landing sites) and be associated with stromatolite deposits (Cady et al., 2003). With the lack of recent plate tectonics (Sleep, 1994), and a lower thermal gradient on Mars (Borlina et al., 2015), preservation in buried (and thereby shielded) sediments could be greatly enhanced (Royle et al., 2018b; Tan et al., 2021), therefore the detectability of biomarkers in the presence of Mars-relevant minerals is of particular interest.
However, this study has ascertained that a larger microbial population/biomass is necessary in iron-oxide/(oxy)hydroxide-rich sediments to be able to ascertain a biological source due to the iron-promoted alteration/transformation of biomolecules. To put the samples analyzed into context, A. platensis has a total organic carbon (TOC) of 40–60% of its dry weight, so the mineral-dried A. platensis mixtures analyzed have TOCs ranging from 4% to 6% down to 0.2–0.3%. In both the core of the Atacama Desert and the McMurdo Dry Valleys in the Antarctic, soil TOCs are generally 0.01–0.1% (Lester et al., 2007; Matsumoto et al., 2010; Lynch et al., 2012; Schulze-Makuch et al., 2018). These hyperarid environments are both frequently discussed as Mars analogues for both astrobiological and geochemical purposes (Navarro-Gonzalez et al., 2003; Stroble et al., 2013). Thus, the lower end of these measurements may be taken as an absolute maximum for a putative martian biomass. This is still likely a gross overestimation as any microbial life in near-surface environments on present-day Mars would likely occupy microhabitats. These microhabitats will be few and far between and potentially only transiently habitable due to the instability of liquid water under martian surface conditions (Davila et al., 2010; Martín-Torres et al., 2015; Jones, 2018), therefore the sustainability of a large biomass, which would be detectable in the presence of the more troublesome iron oxides analyzed here (e.g., goethite or ferrihydrite), is unlikely.
This iron oxide transformation of biological material to more ambiguous hydrocarbon species during analysis by pyrolysis is clearly potentially problematic for Mars life detection, or organic matter characterization, as the main techniques for this rely on thermal decomposition. Longer chain alkanes detected on Mars have been suggested to originate from degraded fatty acids (Freissinet et al., 2019). As all sediments analyzed on Mars so far have contained iron oxides of some form, it may be that the decarboxylation (and potentially hydrogenation) of these fatty acids occurred during analysis rather than as a martian in situ diagenetic process. It is also plausible that some of the nonchlorinated, particularly the aromatic, hydrocarbons detected in the lower Murray mudstone (Eigenbrode et al., 2018) may be the iron-oxide-transformed alteration products of fatty acids rather than from a kerogen-like macromolecular material as hypothesized (Eigenbrode et al., 2018).
The minimal alteration of the pyrolysis products between pure A. platensis and A. platensis pyrolyzed on quartz sand demonstrates that, in comparison with iron-oxide/(oxy)hydroxide-rich units, sediments rich in quartz will be less destructive to biosignature detectability when using classical pyrolysis techniques on Mars. This is in line with other work that has shown the amenability of silica-rich sediments to characterization of associated organic matter (Reinhardt et al., 2020).
Hydrated silica (opal) has been detected at the ExoMars designated landing site of Oxia Planum (Carter et al., 2016), and there are reports of >3.5 Ga old organic molecules being preserved in terrestrial sedimentary hydrated silica (chert) deposits (Duda et al., 2018), potentially making these deposits an ideal target. However, quartz sands appear to have a far lower affinity for, and preservation potential of, microbial lipids, such as fatty acids and their diagenetic products compared with iron oxides and hydroxides (Tan et al., 2018).
Thus, with iron oxides/(oxy)hydroxides being ubiquitous across the martian surface (Evans and Adams, 1980; Bell et al., 1993; Klingelhöfer et al., 2005; Morris et al., 2006a, 2006b; Steele et al., 2007; Bish et al., 2013; Blake et al., 2013; Muttik et al., 2014; Treiman et al., 2016; Dehouck et al., 2017), and with their potentiality to be codeposited with and aid preservation of organic matter (Schelble et al., 2004; Oudemans et al., 2007; Colín-García et al., 2011; Lalonde et al., 2012; Parenteau et al., 2014, 2016; Tan et al., 2018), it would neither be possible, nor advisable to avoid them completely, so strategic sampling, to target hematite-rich deposits, must be devised.
Oxidizing and dry conditions at the surface of Mars mean that in the uppermost regolith, hematite will be the most stable form of iron. Based on the results of this study and Royle et al. (2021), this is inferred to be good news due to the relative amenability of this iron oxide to organic matter detection. However, exposed martian surface conditions are not amenable to present-day habitability nor the survivability of “fossil” organic matter due to the harsh radiative environment (Moores and Schuerger, 2012).
On Earth, goethite is by far the most common iron oxide in soils and sediments due to its high thermodynamic stability (Schwertmann and Taylor, 1989). This is likely also the case in the subsurface martian regolith, out of reach of oxidizing effects of UV radiation (Pollack et al., 1970), and ferrihydrite is likely to increase in concentration beneath the surface for the same reason. Based on the results of this study, this is likely to have a negative effect on life-detection and organic matter characterization efforts in drilled regolith subsurface samples by the future ExoMars 2022 Rosalind Franklin and Mars2020 Perseverance rovers.
Therefore, we need to target localities that have both been shielded from the organic matter degrading effects of solar and cosmic radiation (either subsurface or recently exposed) (Montgomery et al., 2016) and have undergone a diagenetic history encouraging the dehydration, dehydroxylation, and oxidation of more reactive iron-bearing phases to hematite.
Ferrihydrite, magnetite, and goethite all tend to recrystallize to hematite in extended oxidizing conditions regardless of pH or temperature; however, higher temperatures (>200°C) or more neutral/acidic fluids will speed up this process (Gualtieri and Venturelli, 1999; Walter et al., 2001; Lagoeiro, 2004; Cudennec and Lecerf, 2006; Gialanella et al., 2010; Das et al., 2011; Usman et al., 2013). Thus, hematite will be the most stable of the iron oxides in settings that have been subjected to longer term, high temperature, oxidizing, and neutral/acidic fluid flow.
While experimental work has shown that ferrihydrite can survive the short-term, low temperature (<40°C), nonacidic aqueous alteration that is typical of the sediments of Aeolis Palus at Gale Crater, and ferrihydrite conversion to hematite may be retarded by the presence of amorphous silica even under high temperature hydrothermal conditions (Dehouck et al., 2017). Even very brief, strongly acidic episodes of aqueous alteration will rapidly convert ferrihydrite into hematite or goethite; this is suggested to account for the lack of nanophase ferric oxides in the Burns formation at Meridiani Planum (Dehouck et al., 2017), where other phases such as jarosite are evidence for the presence of strongly acidic fluids (Klingelhöfer et al., 2004). As discussed above, the longer these conditions prevailed for, the more the intermediate goethite phase would have been replaced by hematite (Das et al., 2011). The presence of oxidizing and acidic brines is also evidenced from the mineral (and organic) assemblages in the Tissint meteorite (Chen et al., 2015; Steele et al., 2018; Jaramillo et al., 2019). Brine water activities may have persisted all the way from Mars' early history to the present day (Chen et al., 2015), potentially providing a wide range of these environments to explore of varying age, in a suite of different host rocks representing a range of environmental conditions.
Vera Rubin Ridge (VRR) is a unit of fine-grained lacustrine sediments at the base of Mount Sharp, Gale Crater, which have been explored by MSL/Curiosity. Orbital observations, ground-truthed by the rover, have shown extensive enrichment in hematite in both VRR and 100s of meters of the stratigraphically underlying Murray Mudstone formation (Sutter et al., 2017; David et al., 2020; Fraeman et al., 2020; Frydenvang et al., 2020; Rampe et al., 2020; Thompson et al., 2020). It has been inferred that the VRR was created by large-scale and long-term subsurface diagenesis through the actions of warm, oxic waters (Fraeman et al., 2020). Iron precipitated under these conditions as hematite may have been sourced from goethite- or ferrihydrite-rich deposits remobilized from the ancient lakebed (Fraeman et al., 2013). The continuation of predominantly lacustrine sedimentation recorded within VRR (Edgar et al., 2020) demonstrates that habitable lakes persisted in Gale Crater even longer than previously reported (Grotzinger et al., 2014; Fraeman et al., 2020) and, following on from this, multiple, early- and late-stage diagenetic fluids may have provided a refugia for life continuing late into the Hesperian (Fraeman et al., 2020), if originally surface-dwelling communities were able to retreat into the subsurface.
With the combination of potentials for long-term habitability, “rusty sink” preservation of organic matter over geological timescales, and relative amenability for biomarker detection, hematite-rich localities with similar geological histories to the VRR are promising targets for life-detection missions. Orbital observations of the proposed ExoMars 2022 Rosalind Franklin rover landing site at Oxia Planum suggest that the sedimentary units have undergone regional-scale aqueous alteration to enrich the clay units with hematite, while it appears as though more reducing conditions may have been experienced at Mawrth Vallis, a previously proposed landing site, to produce Fe2+-rich layers (Carter et al., 2019). The results of this study suggest that Oxia Planum would thus be a preferential site for ExoMars' planned thermal degradation-based life-detection attempts over Mawrth Vallis.
Furthermore, similar oxic hydrothermal systems, coupled with the presence of reducing organic matter, may have provided a habitable environment for hydrogenotrophic/chemolithotropic microbes (including cyanobacteria) with energy generation from brine–mineral interactions and the oxidation of iron (and sulfate, chlorate, etc.) bearing minerals up to the present (Fernández-Remolar et al., 2008; Nixon et al., 2013; McMahon et al., 2016; Steele et al., 2018). It has also been suggested that concentrated regions of hematite-rich layers on Mars may be evidence of past or present large-scale active iron oxidation. On Earth, this is a process that is extensively mediated by chemolithotropic microorganisms (Weber et al., 2006; Fraeman et al., 2013) and so could be a prime target for future life-detection missions.
Experiments focusing on the degradation of soluble organic matter in ferrihydrite-rich environments suggest that ferrihydrite–magnetite recrystallization has little to no effect on the rate of degradation of organics (Tan, 2020). While these conclusions cannot simply be expanded to all organic matter sources or other iron mineral transformations, it does provide some insight on organic degradation during recrystallization processes.
As well as targeting more deposits with more amenable iron chemistries, it may be possible to optimize pyrolysis techniques or GC-MS operating conditions to target the molecules that are most likely to survive both diagenetic processes and transformation during analysis (e.g., isoprenoid hydrocarbons).
The negative impacts of oxidizing species may be reduced somewhat by ramped or stepped heating experiments, under these conditions (such as where Sample Analysis at Mars [SAM] uses a ramped heating rate of 35°C/min) (Glavin et al., 2013) not all phases will thermally decompose coevally. This contrasts with the “worst case scenario” of flash pyrolysis, which forces the coincidence of iron oxide/(oxy)hydroxide transformation and pyrolytic breakdown of organic molecules, maximizing the alteration/destruction of products (Royle et al., 2018a).
Using a carefully targeted heating program to decompose organic matter and oxidizing minerals at different pyrolysis steps may lead to improvements in analyzing samples rich in iron oxides (and other problematic species) at Mars. The most labile/volatile phases are likely to be affected by surface effects and the release of water and free radicals from ferrihydrite (≥25°C) (Cudennec and Lecerf, 2006; Das et al., 2011) and goethite (≥200°C) (Gualtieri and Venturelli, 1999; Gialanella et al., 2010) dehydration and dehydroxylation, whereas macromolecular phases may be too refractory to thermally decompose until after these most reactive phases have recrystallized. This may be evidenced by the detection of the macromolecular matter in the high temperature evolved gas analysis (EGA) cut by Eigenbrode et al. (2018). In these samples, the high, 500–820°C, temperatures of release of organic molecules would mean that any goethite and ferrihydrite would have already dehydrated/dehydroxylated, and so only hematite (or magnetite if present) would still be available to catalyze reactions.
Future analysis on Mars may have an advantage over the traditional pyrolysis techniques employed so far as both SAM and Mars Organic Molecule Analyzer (MOMA) have wet chemistry capabilities with the ability to carry out derivatized Py-GC-MS. In these techniques, a derivatization agent is used to transform astrobiologically relevant organic molecules (including fatty acids) into species that are sufficiently volatile and amenable to GC column chromatography (Mahaffy et al., 2012).
MOMA will have the further advantage of having two low temperature derivatization techniques N-methyl-N-(tert-butyldimethylsilyl) trifluoroacetamide/dimethylformamide (MTBSTFA/DMF) (≈250°C) and N,N-dimethylformamide dimethylacetal (DMF-DMA) (≈140°C) (Goesmann et al., 2017). The iron oxide/oxy(hydroxide)-catalyzed reactions described here are likely to be slow at these low temperatures, especially with the DMF-DMA technique, which should be within the stability range of all but ferrihydrite dehydration. Experiments in which iron-rich Mars analog sediments and organic standards were used suggest that this technique is effective at avoiding mineral matrix effects from both iron oxides and oxychlorines (Williams et al., 2015a).
However, these derivatization techniques are water sensitive, and so the derivatization agent may be ineffective if hydrated minerals (such as ferrihydrite) are present, also some derivatized compounds may be deactivated by iron oxides (Stalport et al., 2012). The presence of derivatization agents may also add a source of uncertainty to the provenance of pyrolysis products as liquid reagents have the potential to leak and themselves be a source of transformed products and detectable artifacts. This has been a serious problem with determining the source of organic compounds detected by both EGA and Py-GC-MS for the SAM instrument (Glavin et al., 2013; Freissinet et al., 2015; Eigenbrode et al., 2018).
A detection of the potential “transformation” suite of mid-chain length alkanes and aromatic hydrocarbons by “normal” single-shot pyrolysis techniques may thus be useful for sample triage. This could provide a precondition for running more resource-intensive derivatization pyrolysis or controlled multistep/ramped heating techniques on a martian sample.
Another promising way of avoiding negative mineral matrix effects associated with coeval thermal decomposition of organic and inorganic phases may be to use nondestructive techniques. Raman spectroscopy has been shown to be able to detect a range of biomolecules, including cyanobacterial biomarkers, in situ in a range of Mars-relevant sediment and mineral matrices (Wynn-Williams and Edwards, 2000; Böttger et al., 2012; Megevand et al., 2021; Razzell Hollis et al., 2021). This technique appears effective at detecting low concentrations of organics even when oxidizing minerals such as iron oxides and oxychlorines are present (Baqué et al., 2016; Razzell Hollis et al., 2021). A Raman spectrometer is now present on Mars as part of the SHERLOC instrumental suite on NASA's Perseverance rover (Beegle et al., 2015), and another is due to be landed on Mars in 2022 on ESA's Rosalind Franklin rover (Rull et al., 2017). While this technique does not have the resolving power of GC-MS, it should be well suited to identifying suites of biologically relevant compounds in problematic martian samples (Ellery and Wynn-Williams, 2003).
5. Conclusions
Iron oxides/(oxy)hydroxides have transformative effects on the pyrolytic products of the cyanobacteria A. platensis. As concentration of organic matter on the iron-bearing substrate decreases, both the abundance and variety of molecular species detected are decreased. Iron-bearing substrates transform biomolecules, by both oxidative and reductive processes, into highly ambiguous alkanes, aromatic and aryl-bonded hydrocarbons. This would increase the difficulty in ascertaining a biological source even with an extant microbial population.
Despite these ambiguities, a suite containing mid-length alkanes (such as heptadecane) and PAHs and/or aryl-bonded molecules in pyrolyzed samples, which also have a high iron oxide content, may allude to the transformation of cyanobacterially derived mid-long chain length fatty acids (particularly unsaturated fatty acids) originally present in the sample. Isoprenoid hydrocarbons appear to have the greatest survivability of any of the cyanobacterial biomarkers, when pyrolyzed in the presence of iron oxides, and these are also highly recalcitrant over geological time.
Quartz substrates have a minimal effect on the transformation of cyanobacterial biomolecules during pyrolysis. However, hematite, the iron oxide with the lowest transformation potential, has a greater potential for codeposition of organic matter and preservation over geological timescales, so iron-oxide-rich substrates should not be avoided.
For organic matter detection efforts utilizing thermal techniques, sampling should target sediments/strata that have undergone a diagenetic history encouraging the dehydration, dehydroxylation, and oxidation of more reactive iron-bearing phases to hematite by looking for (mineralogical) evidence of the activity of oxidizing, acidic/neutral, and either hot or long-lived fluids.
In martian iron-oxide/(oxy)hydroxide-rich samples, a detection of the potential “transformation” suite of mid-chain length alkanes and aromatic hydrocarbons by “normal” thermal decomposition techniques may be useful as a precondition for running more resource-intensive pyrolysis techniques, including using carefully targeted temperature ramp or derivatization techniques on a drilled sample.
While we have shown here the strong negative effects iron oxides may have on the detectability of modern biological matter, the pristine nature of this cellular material may exacerbate the transformative effects of the mineral substrate during thermal degradation. Further experiments are needed to ascertain the effects of iron oxides on ancient organic matter of biological heritage as this may have more direct application to martian samples.
Footnotes
Acknowledgments
We thank the two anonymous reviewers whose detailed and thoughtful suggestions significantly improved the scope of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Science and Technology Funding Council/UK Space Agency grant ST/N000560/1.
Abbreviations Used
Associate Editor: Kathleen Campbell