
Abstract
The documentation of biosignatures in Precambrian rocks is an important requirement in the search for evidence of life on other ancient planetary surfaces. Three major kinds of biosignatures are crucially important: primary microbial sedimentary textures, diagenetic organomineral assemblages, and stable isotope compositions. This study presents new petrographic, mineralogical, and organic geochemical analyses of biosignatures in dolomitic stromatolites from the Pethei Group (N.W.T., Canada) and the Kasegalik Formation of the Belcher Group (Nunavut, Canada). Both are approximately contemporary late Paleoproterozoic stromatolite-bearing dolomitic units deposited after the Great Oxidation Event. Micro-Raman and optical microscopy are used to identify and characterize possible diagenetic biosignatures, which include close spatial association of diagenetic materials (such as ferric-ferrous oxide and anatase) with disseminated organic matter (OM), dolomitic groundmass textures, and mineralized balls. Many of these petrographic relationships point to the oxidation of OM either biotically or abiotically in association with iron reduction and chemically oscillating reactions. Oxidation of OM in these stromatolites is consistent with the widespread oxidation of biomass during the late Paleoproterozoic Shunga-Francevillian Event. Biosignatures identified in this study are also compared with possible carbonate outcrops on Mars, and thereby contribute a basis for comparison with potential biosignatures in ancient martian terrains. Similarities are drawn between the paleoenvironments of the studied units to the Isidis and Chryse planitia as locations for potential extraterrestrial dolomitic stromatolites.
1. Introduction
The ability to distinguish between abiotic and biotic features in carbonate sedimentary rocks is notoriously difficult (Grotzinger and Rothman, 1996; Foucher et al., 2015), and remains an important goal for geobiological and astrobiological research (Brasier et al., 2015). Precambrian stromatolites have been widely studied in an effort to find fossils of the oldest microbial life-forms and communities on Earth (Nutman et al., 2016; Homann, 2019). However, uncertainty remains regarding whether many stromatolitic structures assumed to be a result of microbial mats are actually biotic in origin (Grotzinger and Knoll, 1999) and not the result of structural deformation (Allwood et al., 2018), or of some abiotic process such as chemically oscillating reactions (Papineau, 2020).
Studies on stromatolites have often relied on paleoenvironmental analysis or comparisons with younger analogues to argue a biotic origin (Allwood et al., 2007; Nutman et al., 2016). The development of reliable petrographic biosignatures in stromatolitic dolomite will assist in reducing such ambiguity, especially when paired with preserved organic matter (OM) inside carbonate minerals. Indigenous OM is not a “smoking gun” (unequivocally definitive) for life in and of itself, but it can be an important biosignature when considered alongside specific sedimentological, diagenetic, and mineralogical features (Cady et al., 2003). We specify here that the term “biosignatures” is used in this work to mean “possible signatures of life” to account for possible abiotic explanations for each type of biosignature. Carbonate stromatolites often lack obvious microscopic textural indicators for a biotic origin (e.g., microfossils) as the crystallization of carbonate minerals usually destroys cell-sized structures (Altermann and Ka
In this study, late Paleoproterozoic stromatolite samples from the Belcher and Pethei groups were studied in the context of the Shunga-Francevillian Event (SFE), an interval that featured the oxidative recycling of OM sequestered during earlier stages of the Great Oxidation Event (GOE) (Kump et al., 2011). The studied units were deposited contemporaneously with the SFE, which can be identified globally as a negative δ13C excursion ∼2.06 Ga (Martin et al., 2013). Postdepositional diagenetic oxidation of OM has been proposed as an important process during this event, and may have produced abiotic botryoidal or concretionary textures as seen in cherty stromatolites previously identified in the Belcher sequence (Gabriel et al., 2021), similarly to apatite rosettes in stromatolitic phosphorite (Papineau et al., 2016), and granules in chert (Papineau et al., 2017; Dodd et al., 2018). Diagenetic oxidation of OM has also recently been suggested to participate in the formation of botryoids of malachite in association with biotic OM (Papineau, 2020).
The identification of biosignatures in carbonate stromatolites has substantial implications for the search for extraterrestrial life, notably on Mars where bodies of liquid water once existed. The newest generation of rovers set to explore the martian surface feature sophisticated, miniaturized instruments that allow for remotely operated geochemical analysis (Bhartia et al., 2021). These include Raman spectrometers capable of sensitive in situ detection of OM and associated minerals, and microfocusing X-ray fluorescence spectrometers to confirm detected mineralogies (Vago et al., 2015). Such extraterrestrial analyses require an assortment of Earth analogues to use for comparative study (Gibson et al., 2001). Stromatolites are an important candidate biosignature to consider because they are often macroscopic, there is extensive literature regarding their formation and inhabiting microbial communities, and they have been used as proxies both for paleoenvironment analysis and for the evolution of life on the early Earth.
This study aims to identify biosignatures in well-preserved late Paleoproterozoic dolomitic stromatolites by imaging the spatial association of OM with minerals and microstructures. We hypothesize that the original distribution of OM in ancient stromatolites governs the spatial distribution of diagenetic minerals and textures in stromatolites. Thus, diagenetic sedimentological features may act as proxy perhaps, biosignatures for the influence of microbial communities on the deposition of carbonate lithologies. As such, this study attempts to answer the following: What microscopic biosignatures exist in stromatolitic dolomite, and how do they spatially relate to OM?
2. Geological Setting
Dolomitic stromatolite samples from two late Paleoproterozoic units were studied in this work: the Kasegalik Formation (Fm) part of the Belchers Gp (Nunavut, Canada), and the Douglas, McLean, Wildbread, and Hearne formations as part of the Pethei Group (Gr) (North West Territory, Canada) (Hoffman, 1988; Sami and James, 1994; Pope and Grotzinger, 2000) (Fig. 1). Both sequences are known to host a range of dolomitic stromatolite morphologies within miogeoclinal settings, present in two different tectonic settings during the assembly of the supercontinent Nuna (Corrigan et al., 2009). The specific stromatolitic dolomite samples investigated in this work are listed in Table 1.
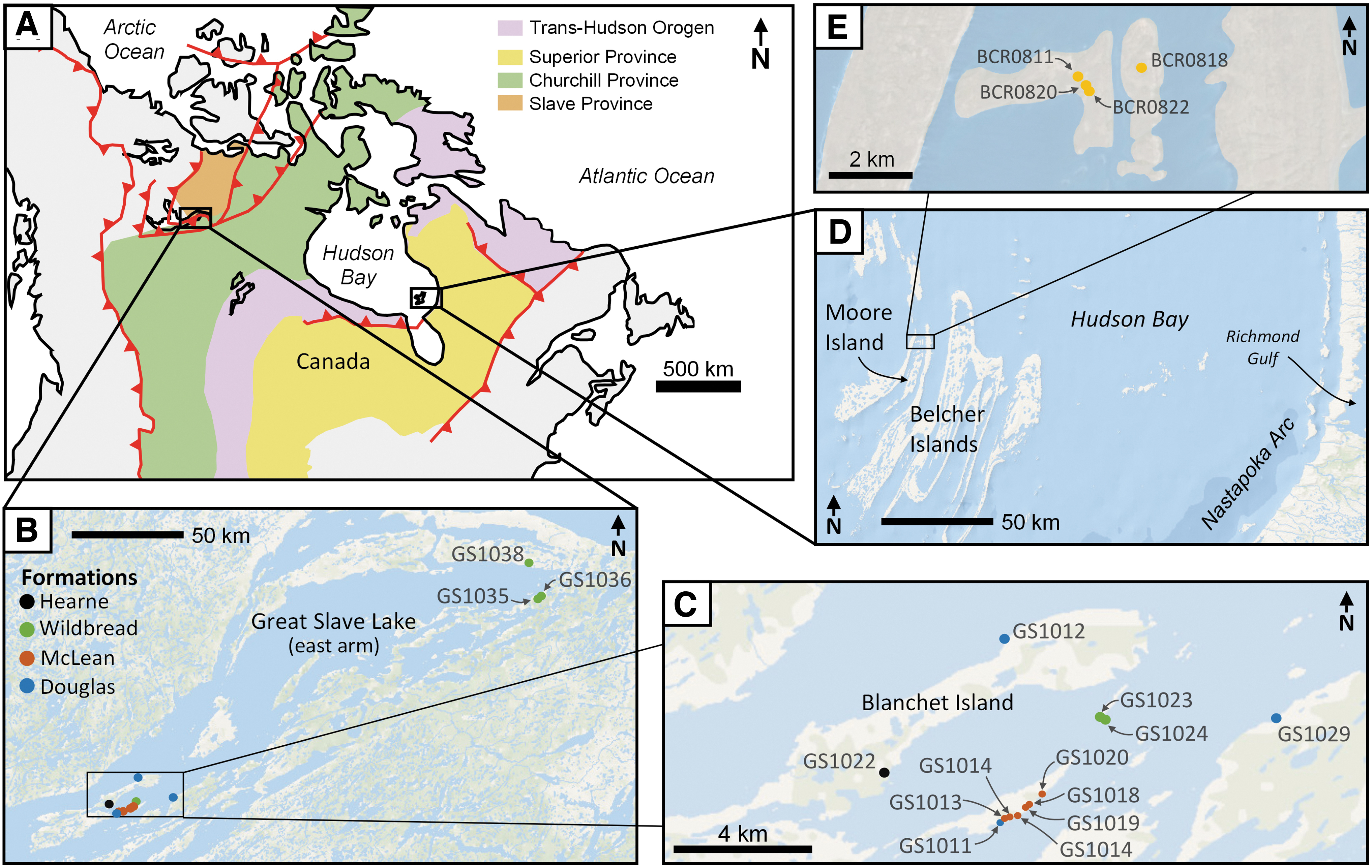
Geological map of Canada
The 26 Studied Samples with Associated Coordinates and Lithologies
Terminology based on Walter (1972).
Samples from the Kasegalik all come from the same section.
2.1. The geological setting of the Pethei Group
The Pethei Gp is a late Paleoproterozoic ∼0.45 km thick foreland-basin carbonate-ramp succession situated on the east arm of the Great Slave Lake (N.W.T., Canada) (Hoffman, 1988; Sami and James, 1996; Pope and Grotzinger, 2000) (Fig. 1A–C). The Pethei Gp is part of the Great Slave Lake Supergroup and is noted for multiple prolific stromatolitic horizons that display a diverse range of morphologies, superposed on a predominantly shoaling-upward marine sequence (Sami and James, 1996) (Fig. 2). This sequence is representative of two ramp successions (Sami and James, 1993) estimated to have been at a paleolatitude of ∼18°N, and facing northward (Hoffman, 1988; Mitchell et al., 2010). The Pethei Gp formed ∼1.88 Ga based on U-Pb zircon dating of correlated laccolith intrusions in the Wopmay orogen (Hoffman and Bowring, 1984).
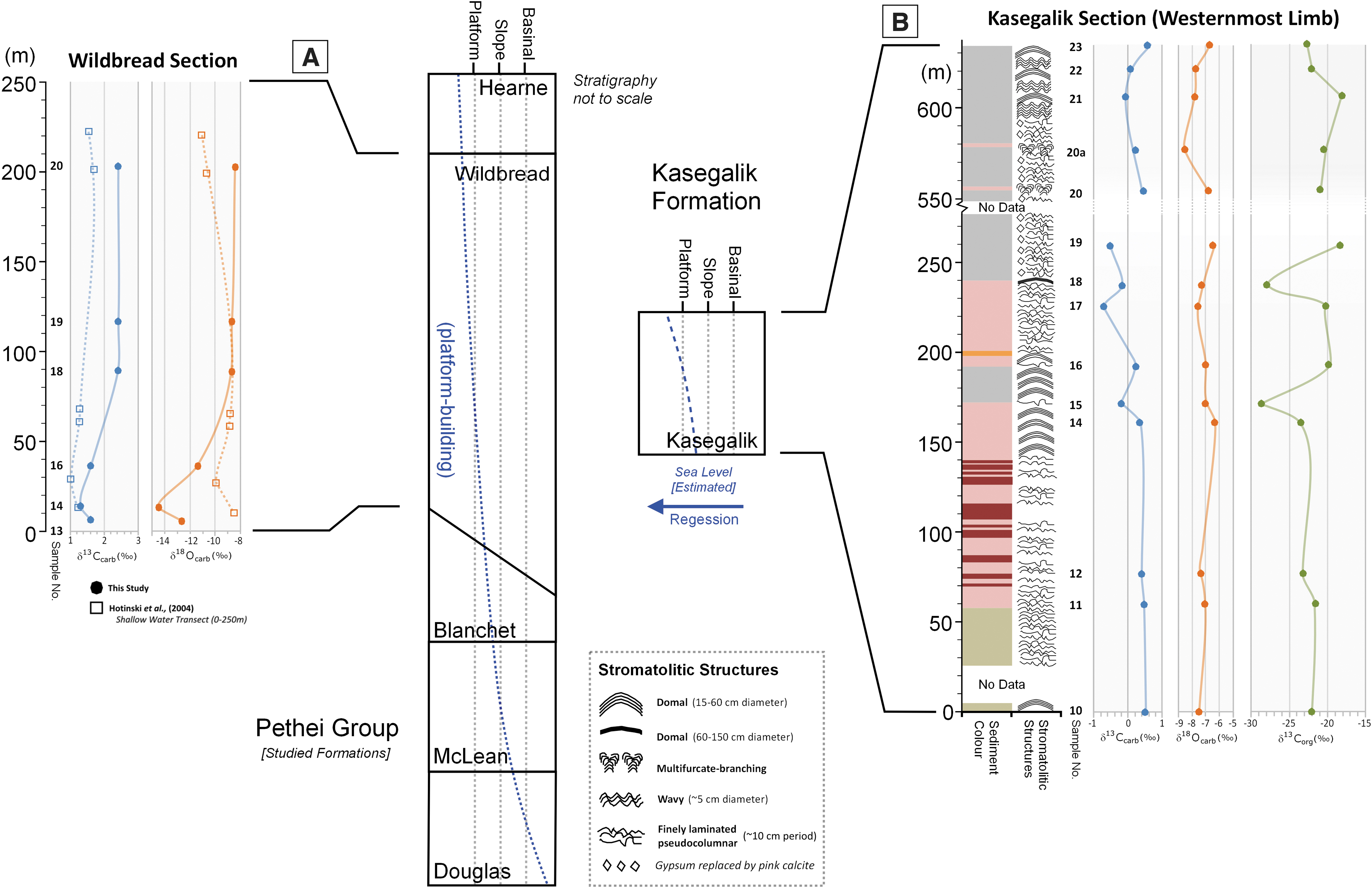
Sections of the studied formations, with schematic indicating inferred water depth during deposition (adapted from Jackson, 1960; Hoffman, 1988; Corrigan et al., 2009). The regional stratigraphy is correlated with two sections with δ13C and δ18O isotope records for
The Pethei Gp initially represents the filling of a foreland basin along the southern margin of the Slave Craton. Deposition is proposed to have occurred syn- and post-thrusting during the short-lived Wopmay orogen (Hoffman and Bowring, 1984; Hoffman, 1988), which involved oblique collision between the Slave and Churchill provinces ∼1.92–1.97 Ga (Hoffman et al., 1989; Sami and James, 1994), followed by postorogenic relaxation. The notably linear character of the structurally controlled rift basin (Chandler, 1980) is thought to represent the failed arm of a triple-point junction (the Athapuscow aulacogen), which created significant accommodation for the margin-to-basinal deposition of the Pethei succession (Hoffman, 1973). Thanks to high rates of tectonic subsidence, the Wildbread–Hearne carbonate platform lacked significant transgression (Sami and James, 1996). Gabbro dykes from the Mackenzie dyke swarm ∼1.085 Ga crosscut and locally caused contact metamorphism in some exposures of the Pethei Gp (Roscoe et al., 1987).
The stratigraphic sequence of the Pethei Gp is composed of four prominent formations. The Douglas Fm represents a basal deep-marine marlstone facies characteristic of sedimentation before the development of a carbonate ramp (Sami and James, 1993). It is overlain by the McLean Fm, which is composed of turbidites interbedded with columnar and anastomosed stromatolitic marlstones (Fig. 3A–C), indicative of marginal slope facies during ramp development (Hoffman et al., 1977). Both units feature deep-water sediments, which become more calcareous upslope (Roscoe et al., 1987). Overlying these facies is a shallowing-upward succession composed of the Wildbread and Hearne formations, representative of ramp-top environments (Sami and James, 1994). Both are composed of domal to bulbous stromatolitic dolomite formed in shallow water within low-energy back-reef environments (Fig. 3D–I). The Hearne Fm represents a transition toward a basinal marine setting (Sami and James, 1996) and marks the top of the Pethei Gp (Fig. 2).

Field images of dolomitic stromatolites in the Pethei Gp:
2.2. The geological setting of the Kasegalik Formation
The Kasegalik Fm outcrops on the Belcher Islands, a small archipelago located south of the Hudson Bay, Nunavut, Canada (Fig. 1A, D, E). The islands are dominantly composed of folded dolomitic and siliciclastic sedimentary rocks grouped into the 6–9 km thick Belcher Group (Hofmann, 1976). The sequence includes mixed volcanic rocks, siliciclastics, and cherty dolomite indicative of a Paleoproterozoic tidal carbonate-platform environment, punctuated by episodes of subaerial and subaqueous volcanism evidenced by the presence of columnar and pillow basalt. Early sediments were deposited within the remnants of the Proterozoic Aphebian Trans-Hudson orogenic belt ∼1.7–1.9 Ga (Ueda et al., 1991), with the basal Kasegalik Fm presumably overlying >2.5 Ga old Archean basement although a contact has yet to be observed (Hofmann, 1976). Uranium-lead dating of zircons from volcanic horizons in the overlying Eskimo Fm (Chauvel et al., 1987) as well as from tuffs in the lowest and uppermost parts of the Kasegalik Fm yield a depositional age at 2015 ± 2.1 Ma and 2018 ± 2.6 Ma, respectively, with a maximum deposition of age of 2120.6 ± 2.5 Ma evaluated from detrital zircons from sandstones in the middle Kasegalik (Hodgskiss et al., 2019).
The Belcher Islands are located slightly south of the geometric center of the Nastapoka Arc (east coast of Hudson Bay), a perfectly semicircular geomorphological feature with a diameter of ∼450 km (Fig. 1A, D) present along the unconformity between underlying Archean cratonic material and overlying gently dipping Proterozoic mixed sediments (Hynes, 1991). The Nastapoka Arc may represent the front edge of an east-verging foreland thrust-and-fold belt, developed during the Trans-Hudson orogen ∼1.8 Ga (Darbyshire and Eaton, 2010; Pawlak et al., 2012). Later postorogenic collapse led to reactivation of the underlying foreland thrust-and-fold belt, forming a cratonic margin. Tight, kilometer-scale, isoclinal upright folding on the Belcher Islands (Hynes, 1991) suggests a later postdepositional deformation episode involving thin-skinned tectonics and activation of near-surface decollements (Hynes, 1991). The possibility that the Nastapoka Arc represents an “astrobleme” was hypothesized during initial studies (Hofmann, 1976); however, no compelling evidence for an impact has been published. The Kasegalik Fm underwent subgreenschist facies regional metamorphism ∼1.6–1.7 Ga (Donaldson, 1970).
The ∼1.2 km thick Kasegalik Fm outcrops locally within the Kasegalik Lake anticline in the westernmost limb of the Belcher Islands (Fryer, 1972; Donaldson and Ricketts, 1979) (Fig. 1). Sedimentation forms a miogeoclinal sequence, with carbonate deposition on a passive-margin continental shelf in a sabkha to shallow-subtidal setting (Legault, 1994; Hodgskiss et al., 2019). Much of the formation consists of dolomite containing stromatolitic horizons (Fig. 2). Near the base, laminated mudstones are present with evidence of wave-ripples, mudcracks, and channel fills (Hofmann, 1976), which collectively suggest deposition on a shallow supratidal to subtidal marine platform (Ueda et al., 1991). Decimeter- to meter-size dolomitic stromatolite structures are common throughout the formation (Fig. 4A), featuring large 2–5 m domes (Donaldson and Ricketts, 1979) (Fig. 4B) and some smaller bioherms with multifurcate, turbinate columns a few millimeters in size (Fig. 4C). These structures are preserved in very-fine-grained to crystalline wrinkly laminated dolomite (Fig. 4D), interbedded with red beds (Fig. 4E) and black chert, which also contain exceptionally preserved microfossil assemblages (Hofmann, 1976).

Field images of stromatolites in the Kasegalik Fm:
3. Materials and Methods
3.1. Materials
A total of 26 thin sections are studied, and include 17 from the Pethei Gp and 9 from the Kasegalik Fm (Table 1). Of these 26 thin sections, the dozen that have the most potential for reliable biosignatures are investigated in detail with micro-Raman spectroscopic imaging (Fig. 5). The samples are collected from exposures that consist of dolomitic stromatolite with variable amount of chert, hematite, pyrite, and siliciclastics. In the Kasegalik Fm, dolomitic stromatolite laminations are generally light gray to light pink, whereas in the Pethei Gp stromatolitic dolomite is red, pink, or light brown, and exhibits wrinkled to domal laminations richer in hematitic siliciclastic material, for instance in the Douglas and McLean formations (Fig. 5).
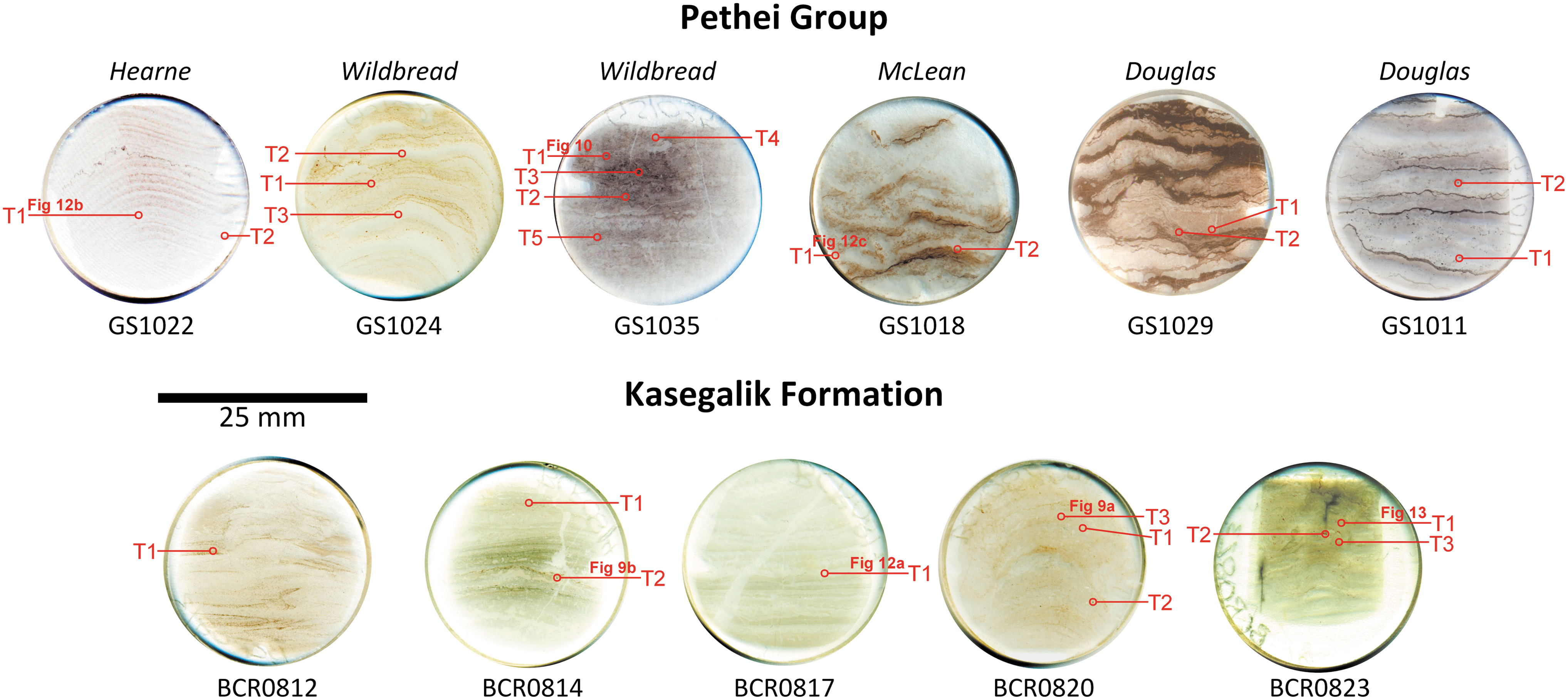
Eleven thin sections studied using Raman spectroscopy, showing sample maps identifying the target locations for Raman scans. Thin sections were cut orthogonal to laminations to provide a cross-section through textures and microstructures.
3.2. Analytical methods
3.2.1. Optical and micro-Raman microscopy
Raman spectroscopy is an analytical technique useful to distinguish the structure of OM (Schopf and Kudryavtsev, 2005). It is a nondestructive analytical method with submicron-scale spatial resolution, and is able to analyze both inorganic and organic substances (Steele et al., 2020; Allwood et al., 2006). Laser Raman spectroscopy is already a favored technique used in laboratories studying martian meteorites, as well as being a key analytical method in paleobiology (Edwards et al., 2003, 2007; Ellery et al., 2004; Clarke and Stoker, 2013).
For this study, samples are first analyzed through optical microscopy under transmitted, crosspolarized, and reflected light (RL). Optical microscopy is performed with an Olympus BX51 microscope using 5 × , 10 × , 20 × , 50 × , and 100 × objectives on polished 30 μm thin sections to locate target biosignatures. Petrographic imaging is documented in RL and plane-polarized light (PPL), although crossnicols and dark field were also used in the research to highlight features that can be compared with expected images from future automated martian laboratories (rovers).
Micro-Raman imaging is then performed with a WITec α300 confocal Raman imaging system located in the Geological Spectroscopy Laboratory of the Department of Earth Sciences at University College London. The 532 nm laser power is set at 7–8 mW, and then focused at 500 × and 1000 × magnification to scan small areas. A 50 μm diameter optic fiber is used to collect the Raman spectrum at a confocal depth ∼0.5 μm below the polished surface, and a grating of 600 lines/mm is used to generate a large bandwidth and a spectral resolution of 4 cm−1. Data processing is completed on the WITec “Project Four Plus” software. All presented average Raman spectra are corrected with a background subtraction by using polynomial fits of order 5 or 7 following cosmic ray removal (filter size: 2 pixels, dynamic factor: 8 × CCD counts).
Raman hyperspectral images that depict mineral associations are created by mapping the peak intensities for the strongest mineral-specific peaks at each pixel. Unless otherwise noted, minerals in Raman hyperspectral images are colored with respect to the following peaks: quartz = 465 cm−1 (blue), dolomite = ca. 1099 cm−1 (green), feldspar = 512 cm−1 (orange), hematite = 1320 cm−1 (purple), pyrite = 373 cm−1 (turquoise), anatase = 141 cm−1 (yellow), fluorapatite = 965 cm−1 (yellow), rutile = 488 cm−1 (turquoise), ferric-ferrous oxide = 670 cm−1 (turquoise), and OM = 1600 cm−1 (red). This color scheme is selected to best highlight contrast between different phases, which are also distinguished with annotations. This is performed with the WITec “Project Four Plus” processing software, and Raman peak positions are read from average spectra calculated from pixels with high signal-to-noise ratio.
3.2.2. The Raman spectra of OM preserved in dolomitic stromatolites
The Raman spectra of OM can be broadly divided into first-order (1100–1800 cm−1) and second-order (2200–3400 cm−1) regions (Ferrari and Robertson, 2000; Beyssac et al., 2002a). Second-order peaks are often the result of combination scattering (Nemanich and Solin, 1979; Beyssac et al., 2002a), with characteristic peaks formed ∼2700–2900 cm−1. OM found in the studied carbonate stromatolites always has two distinguishable peaks: the G peak (∼1580 cm−1) and the D1 peak (∼1350 cm−1) (Beyssac et al., 2002b). Other peaks such as the D2, D3, and D4 are not resolved in any spectrum, but are used for spectral modeling (Fig. 6A).

Spectral characteristics of the first-order OM Raman band:
Raman spectra of preserved OM can be used to estimate the peak metamorphic temperatures of the studied units as OM becomes more ordered upon exposure to higher temperatures (Marshall et al., 2010) (Fig. 6B). This ordering is displayed in the relative intensity (≈height) of the D1 and G Raman peaks in the first-order spectrum for OM. Heating OM results in the irreversible reorganization and polymerization of aromatic structures. The changing shape of these two peaks in OM spectra records peak metamorphic temperatures independent of later retrograde effects (Beyssac et al., 2002a; Lahfid et al., 2010). Thus, correct modeling of spectral characteristic can be used as a metamorphic peak-temperature indicator and a useful index for metamorphic grade in low-pressure regimes (Beyssac et al., 2002b, 2003; Lahfid et al., 2010).
In this work, the maximum exposure temperatures of OM are estimated by using the geothermometer of Lahfid et al. (2010), which covers the prehnite-pumpellyite to lower greenschist facies temperature range. The empirical relationship shows that the intensity of the D1 peak increases at the expense of the D4 peak at a temperature range of 230–300°C. The symmetry of the G peak also changes, preferring to skew toward higher relative wavenumbers (Raman shifts) with increasing pregraphitization temperatures (Lahfid et al., 2010) (Fig. 6B). Peak fitting is modeled by using numerical methods on averaged, background-subtracted spectra. Peak areas are derived from the integration of Lorenz functions as linear combinations of the five bands (D1–D4 and G) over a spectral range of 1000–2000 cm−1. To ensure accurate results, only spectra of OM not associated with hematite are selected for this analysis.
3.2.3. Stable isotope analyses of carbonate, OM, and pyrite
Stable isotope compositions of carbonate and OM are measured in bulk powders. Rock powders are prepared from cuboid rock fragments cut with a water-cooled diamond saw and grinded to remove saw marks. Cleaned fragments are crushed with a steel mortar and pestle cleaned with methanol, and rinsed with deionized water between each sample and with intercalated crushates of pure quartz powders muffled in air at 600°C for 2 h to minimize cross-sample contamination. Bulk analyses of carbon isotopes were then performed at the Carnegie Institution for Science and UCL by using previously used techniques (Papineau et al., 2013). These include a Gas Bench for bulk dolomitic carbonate and an Elemental Analyzer for OM in HCl-dissolved dolomitic powder; both linked to a Thermo-Delta V isotope ratio mass spectrometer. Reproducibility (1σ) on analyses of procedural internal standards for δ13Ccarb is better than 0.1‰, δ13Corg is better than 0.2‰, for δ18OPDB is lower than 0.3‰.
Sulfur isotope analyses (δ34S) are performed with a CAMECA NanoSIMS 50L at the Carnegie Institution for Science according to previously described methods (Hauri et al., 2016). Pyrite is identified by RL microscopy, and four grains separated from BCR0818 are mounted in Indium and subsequently polished with 0.25 μm Al2O3. Precision with the nanoSIMS for δ34S is better than 0.02‰ and the 2σ reproducibility for δ34S is ±0.53‰, based on the analysis Balmat pyrite standards (n = 10) and Trout Lake chalcopyrite standards (n = 4).
4. Results
4.1. Stable isotope compositions of carbonate, OM, and pyrite
Geochemical analyses reveal homogeneous δ13C values for dolomite in the McLean Fm, where it varies between +1.6‰ and +2.4‰ (n = 6), and in the Kasegalik Fm, where it varies between −0.7‰ and +0.5‰ (n = 14) (Table 2), consistent with published data from the work of Hotinski et al. (2004) and Hodgskiss et al. (2019), respectively. Values of δ18OPDB for bulk carbonate are between −14.5‰ and −8.4‰ (n = 6) for the McLean Fm and between −8.5‰ and −6.3‰ (n = 14) for the Kasegalik (Table 2). Carbonate δ13C and δ18O isotope data are shown in their generalized stratigraphic context where available (Fig. 2). For OM, δ13Corg values in the Kasegalik Fm are between −18.1‰ and −28.6‰. Finally, δ34S values for submillimeter-size euhedral pyrite in dolomitic stromatolite from Kasegalik Fm are homogeneous and between +30.7‰ and +32.5‰ (n = 12) (Table 3).
Carbon and Oxygen Isotope Data in Two Measured Sections in the McLean and Kasegalik Formations, Depicted Graphically in Fig. 2
NanoSIMS Sulfur Isotope Data for Three Pyrite Grains Recovered from Sample BCR0818
4.2. Diagenetic mineral assemblages associated with OM in dolomitic stromatolites
Through the analysis of 26 Raman scans (Fig. 5), the crystallinity and distribution of OM are characterized in the different formations. The collected data show a broad affinity toward predominantly intergranular OM in the Pethei, and disseminated poikilotopic OM within the carbonate groundmass of the Kasegalik Fm. Raman spectra for OM in the Kasegalik Fm and Pethei Gp depict highly disordered OM, with broad D1 bands typically peaking at ∼1330 cm−1 (full width at half maximum between 90 and 140 cm−1) and a superimposed G+D2 peak occurring at ∼1610 cm−1 (full width at half maximum ∼55 cm−1) (Fig. 6B). A selection of OM spectra from three samples in the Kasegalik Fm and in the Pethei Group is used to estimate peak metamorphic temperature based on the Lahfid et al. (2010) geothermometer (Table 4).
Summary of the Paleotemperature Analyses on Selected Samples Using the Lahfid et al. Paleothermometer Model A1 and A2 Equations
The spectral parameter “R” is given by Lahfid et al. (2010), as [D1/(G + D2)] × 100, such that increasing R correlates with increasing peak metamorphic temperature pregraphitization.
The bold values correspond to the values seen alongside the plots in Figure 6B.
Ferric-ferrous oxide is a common accessory material in many samples, but its Raman spectrum does not allow identification as a specific mineral. Hence, this phase is referred to as ferric-ferrous oxide because it contains both the distinguishing Raman peaks for hematite (∼1320 cm−1) and for magnetite (∼664 cm−1). In the Pethei Gp, ferric-ferrous oxide typically occurs as fine (∼10 μm) grains disseminated throughout laminations, whereas in the Kasegalik Fm stromatolites grains are fewer, coarser, and often constrained to clearly defined laminated horizons (Fig. 7A, B). In the Pethei Gp, ferric-ferrous oxide grains sizes vary from micron-size (1–10 μm) disseminations up to coarse (50–100 μm) subhedral to euhedral grains (Fig. 8A). Ferric-ferrous oxide shows preferential mineralization within certain laminae-parallel horizons (Figs. 7A and 8B), most notably in the domed Kasegalik stromatolites (Fig. 8A). Variable concentrations of ferric-ferrous oxide mineralization often form weak laminations, which also mirror alternations in the concentration of disseminated hematite. Ferric-ferrous oxide typically displays a close association, or is in direct contact with areas of significant OM concentration in dolomite (Fig. 9A–E). This is a similar relationship to that seen with subhedral pyrite grains (Fig. 9A, B).
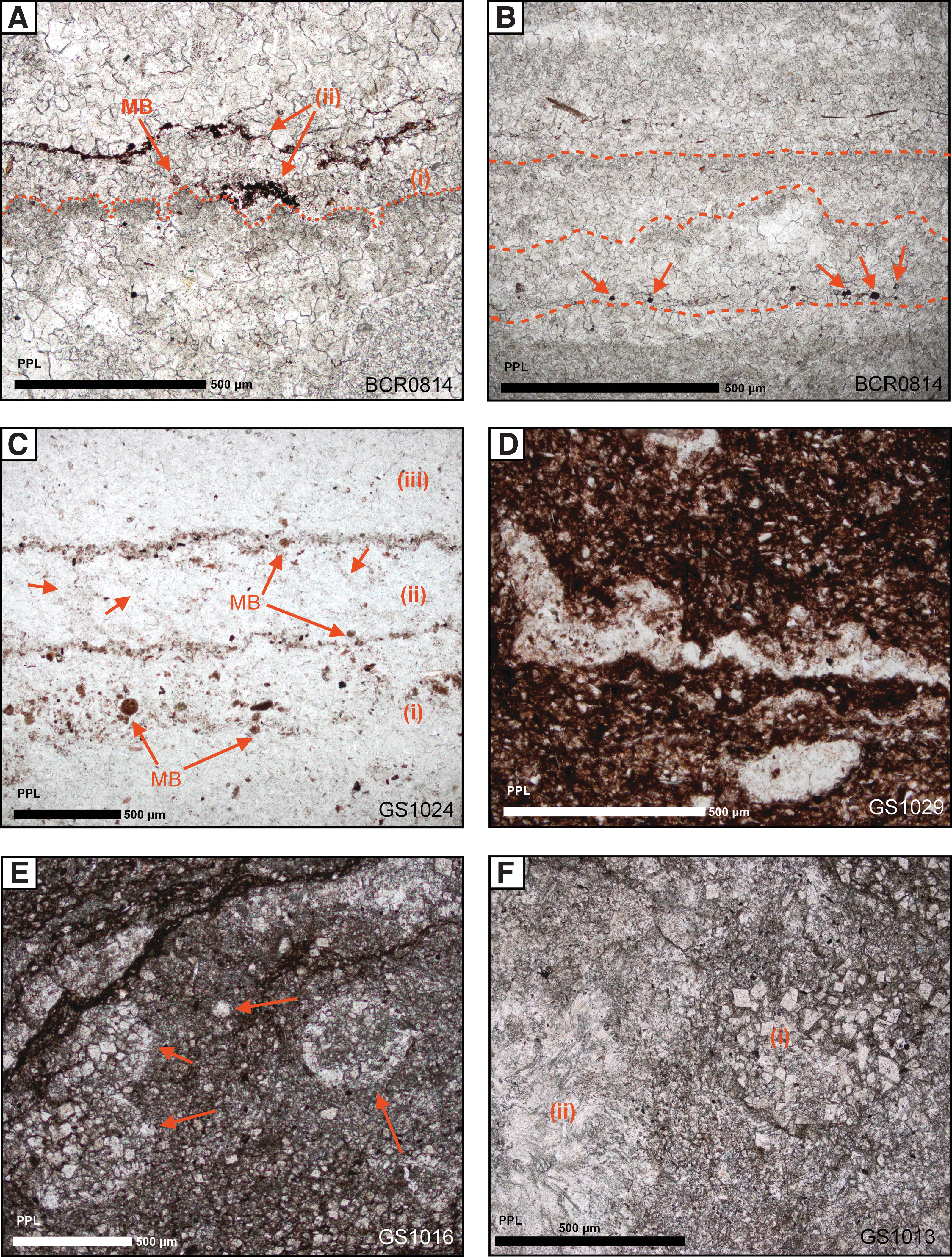
Photomicrographs of sedimentary laminations and textures in plane-polarized light:

Photomicrographs of thin sections, RL:

Micro-Raman scans of two targets
Anatase is an accessory mineral that is ubiquitous throughout the Pethei Gp dolomitic stromatolites and some Kasegalik Fm stromatolites. Nearly all Raman scans show the presence of accessory anatase in close spatial association with OM (Fig. 10). Anatase most often occurs as euhedral crystals, typically 1–15 μm in size, although it also congregates as fine acicular needles forming amorphous masses up to 30 μm in size (Fig. 10B). Authigenic micron-size anatase grains are preferentially concentrated in detrital-rich laminations, often in spatial association with authigenic pyrite grains. Although TiO2 can form a number of polymorphs in sedimentary rocks, anatase is the predominant diagenetic phase detected. Brookite was not observed in any of the samples, and only minor fine (<5 μm) accessory rutile is sometimes present (Fig. 8E, F). Unlike the Pethei Gp where both ferric-ferrous oxide and anatase are preferentially found in laminations alongside OM, the Kasegalik Fm stromatolites have more disseminated OM throughout the carbonate groundmass independent of anatase (Figs. 8E, 8F, and 9B). In the Pethei Gp, anatase concentrations are directly related to OM with more numerous and larger (up to 10 μm) anatase grains in OM-rich areas. Anatase grains are fewer and larger in the Kasegalik Fm than in the Pethei Gp dolomitic stromatolites.

Dissemination of OM in diagenetic phases from the Douglas Fm:
4.3. Microscopic laminations and carbonate sedimentology of Paleoproterozoic dolomitic stromatolites
A key defining characteristic for the classification of stromatolites at the microscopic scale is the nature of their laminations. No classification scheme has been universally accepted for stromatolite microstructures except to characterize the nature of sparry and fine-grained laminae carbonate crusts (Riding, 2008). In the studied stromatolites, laminations typically vary according to their detrital material content (Fig. 8B), alternating carbonate grain size (Fig. 7B), disseminated hematite mineralization, concentration of diagenetic mineralization (Fig. 7C), or a variable combination of these factors. In this study, a lamination is regarded as a planar to parallel sedimentological unit of a thickness <10 mm. Micrite is defined as microcrystalline carbonate with a grain size <5 μm, microsparite 5–30 μm, and sparite >30 μm. Dolomite and calcite can be differentiated by their main Raman peak at 1095–1104 cm−1 and ∼1085 cm−1, respectively.
In the wrinkly to wavy laminated, domal, and columnar stromatolites of the Pethei Gp (Fig. 3E–I), submillimeter laminations generally occur within coarse sparite that alternates with thin bands of detrital- and hematite-enriched microsparite. In the basal units of the Douglas and McLean formations, laminations are predominantly the result of grain size changes, creating rhythmic sparry laminae. Variation in grain size becomes less significant toward the top of the Pethei Gp where detrital material (including quartz and feldspar) becomes more abundant in the Hearne Fm (Fig. 8B). Within the Douglas and McLean, laminations consist of dense concentrations of OM, pelitic material, and/or hematite, which form high-opacity red, muddy laminations (Fig. 7D). These laminae commonly occur in clotted fabrics with internal convolutions around intraclasts and large grains up to 300 μm, creating undulose to pseudocolumnar laminae morphologies. Such contorted laminae contain greater concentrations of allochthonous grains (both siliciclastics and carbonate clasts) in <1 mm bands (Fig. 8B) with a clotted or ornamented fabric. In the Douglas Fm, individual hematite-enriched laminae have internal structure linking laminations—with rounded micritic filaments that connect laminar horizons (Fig. 7C).
Higher in the stratigraphy of the Pethei Gp, laminations are predominantly rhythmic alternations in detrital content (Fig. 8B), and they correlate with the facies shift from an off-ramp to ramp-top environment. Clotted and silty laminations (Fig. 7D, E) are associated with finely laminated wrinkled and columnar stromatolites in the Douglas and McLean formations (Fig. 3B, C), in contrast to more homogeneous rhythmic laminations seen in the linked domal stromatolites of the Hearne Fm, repeating at decimeter intervals (Fig. 3E). Hearne Fm stromatolitic laminations are distinguished by alternations in the concentration of noncarbonate detrital grains (Fig. 8B). Such detrital grains often correlate well with later diagenetic minerals preferentially crystallized within detrital-enriched horizons. Disseminated hematite also acts to record submillimeter alternations. Through the transition from the basal facies of the Douglas to the platform facies of the Wildbread and Hearne formations, clotted muddy laminations (Fig. 8D) become rarer, and there is a change to rhythmic laminae-interlaminae alternations with generally higher micrite content (Fig. 8C).
The sedimentary textures of the Kasegalik Fm show similarities to the Wildbread Fm. For instance, both share a similar grain size and regular lamination intervals. The columnar layered to partly linked domal stromatolites of the Kasegalik Fm (Fig. 4A, G, H) features laminations distinguishable by alternations in carbonate groundmass grain size by an order of magnitude or more (typically between coarse and microcrystalline sparite) (Fig. 7B), either as an original fabric or from later diagenetic overprinting. Laminations in the domed decimeter-scale Kasegalik stromatolites commonly display roughening-upward textures where stacked laminations become increasingly more wrinkled to pseudocolumnar, before resetting to flat laminations (Fig. 7A, B). Higher in the stratigraphy, stromatolites show a broad trend toward wavy laminations and branching domes (Fig. 2), with samples preserving significant amounts of detrital siliciclastic phases, including feldspar and quartz.
4.4. Petrography and mineralogy of Paleoproterozoic dolomitic stromatolites
4.4.1. Stromatolitic dolomite groundmass characteristics and morphologies
Visually identifying the difference between dolomite and calcite is often difficult within the various formations, except where dolomite grains have a characteristic rhombic habit (Fig. 7E, F). Unlike the Pethei Gp stromatolites, whose groundmass generally consists of mosaics of calcite and dolomite, the Kasegalik Fm stromatolites have horizons that alternate between dominantly calcite or dolomite rich. Two morphological end-members for the groundmass mosaic texture of sedimentary dolomite exist, in agreement with previous temperature studies completed by Gregg and Sibley (1984) and more recently by Huang et al. (2014): (1) an idiotopic texture comprised of planar euhedral crystals and (2) a xenotopic texture that consists in nonplanar anhedral crystals with irregular grain contacts. Idiotopic euhedral dolomite is often found preferentially within and surrounding hematite-rich laminations, predominantly near the base of the Pethei Gp. The McLean Fm displays the most diversity in dolomite crystal morphologies, with idiotopic dolomite preserving structural textures, including subspherical masses ∼500 μm in size, undulating laminations, and clotted microcolumnar features (Fig. 7E, F). Euhedral dolomite grains are often 10–20 μm in length and dirtied by dense, ultrafine inclusions (Fig. 7E). Near the top of the Pethei Gp, dolomite is almost exclusively xenotopic in nature and correlates with a general decrease in groundmass grain size higher in the group.
4.4.2. Character and distribution of mineralized balls
“Mineralized balls” are a variety of subspherical to spherical hematite-rich sedimentological features that range in size from 20 to 200 μm (Fig. 11 and Table 5). They are enriched in finely crystalline (∼4–10 μm) noncarbonate detrital minerals such as hematite, feldspar, and rutile, as well as OM and anatase (Fig. 12A–F). These minerals are often distributed throughout the mineralized balls homogeneously (Fig. 12A–D), but can form internal concentrations (Fig. 12E, F). Balls often feature high-contrast hematite coating, but hematite density is highly variable and often related to the stratigraphic horizon as well as sphere composition—where mineralized balls with greater detrital mineral content generally contain proportionally more hematite. Balls typically cluster to form sublaminar bands parallel to laminar alternations in groundmass grain size and detrital mineral horizons (Fig. 8D).
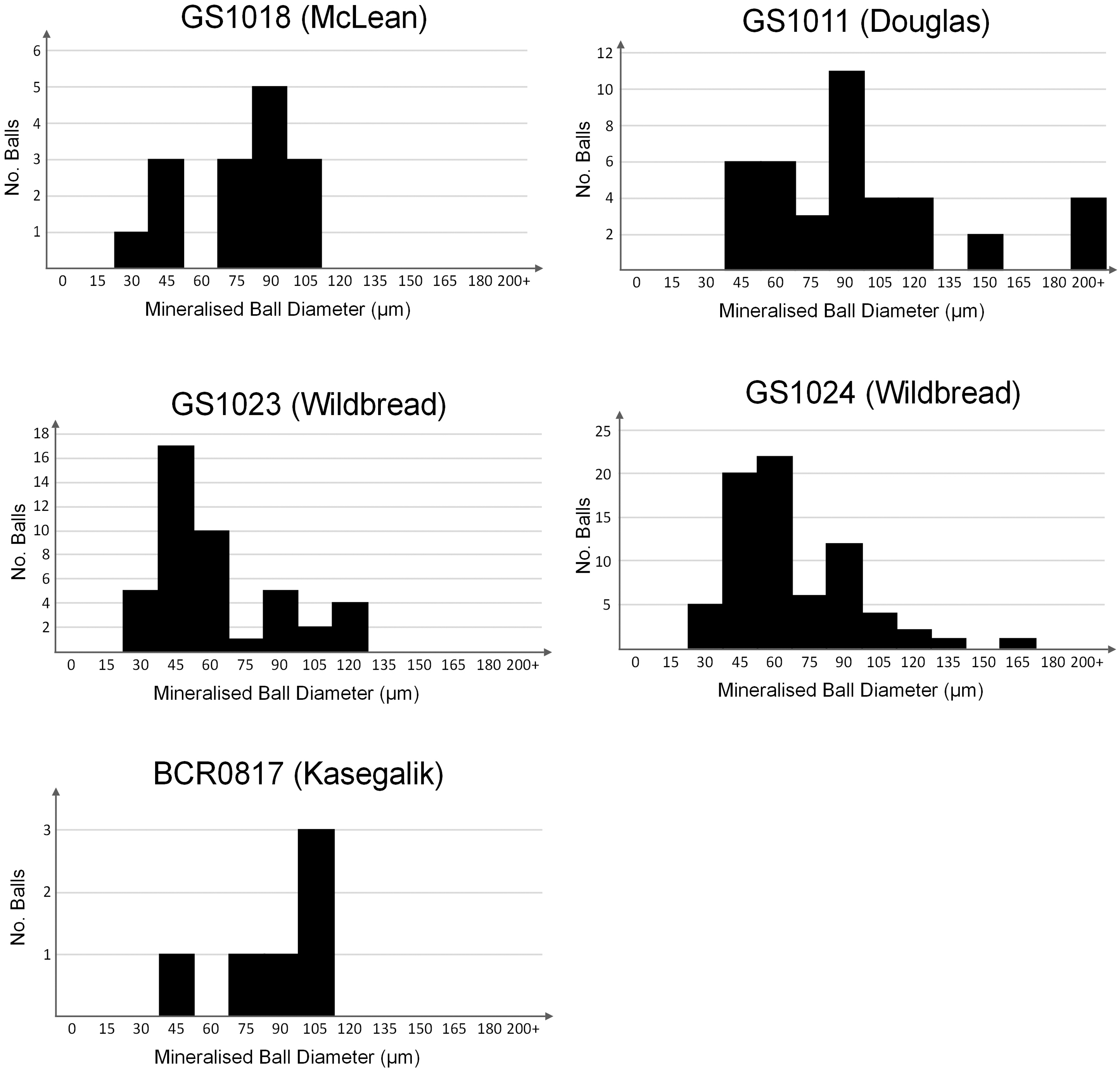
Histograms showing the distribution of binned mineralized ball diameters for five different thin sections (Table 3). There is a broad distribution of diameters over one order of magnitude from 20 to 200 μm, with an under-representation of balls with a diameter of 60–75 μm.

Photomicrographs and hyperspectral images of MB in stromatolitic dolomite. MB occur as rounded masses of detrital minerals (often quartz) in association with OM and TiO2 polymorphs;
Data Collected for Mineralized Ball Size Distributions from Five Thin Sections
See Fig. 11 for a graphical representation.
Balls do not feature concentric laminations that could be indicative of ooids or of any other pisolite (Fig. 12A–F). The concentration of noncarbonate minerals varies and is often difficult to determine due to excessive fluorescence during Raman imaging. Such fluorescence occurs in a broad peak ∼1400 cm−1, corresponding to the first-order region for OM (Fig. 12G). Mineralized balls are present in both the Pethei Gp and Kasegalik Fm, and are most common within platform-margin samples. Their size distributions show a weak bimodal trend in sphere diameter between 20 and 200 μm (Table 5) with a deficit in spheres with a diameter 60–75 μm (Fig. 11). This size distribution is consistent across the Pethei Gp, but is difficult to ascertain for the Kasegalik Fm due to their lower abundance.
4.4.3. Siliceous botryoids associated with Fe-oxides and OM
A sample from a multifurcate columnar stromatolite in the uppermost Kasegalik Fm (Fig. 4C) features replacement and/or infilling of banded OM-rich micrite laminations by chert (Fig. 13A). Quartz infills of formerly vuggy porosity exhibit botryoidal textures, with concentric growth of radial, acicular quartz (Fig. 13B–D) around masses of OM in their geometric centers (Fig. 13H). These botryoids have a radius of ∼200 μm and fill pore spaces, with remaining voids infilled by isopachous acicular quartz grains. Coarse idiotopic dolomite rhombs up to 300 μm in size overprint some textures (Fig. 13A). Raman images of botryoids reveal very fine clusters of crystals of dolomite and ferric-ferrous oxide in conjunction with OM in their centers (Fig. 13I). Microlaminations within botryoids are perfect circular sections nearly equally spaced at ∼20 μm, and they consist of clean acicular quartz progressively colored by nanoscopic impurities toward the geometric center (Fig. 13C, G).

Silicified botryoids in Kasegalik Fm sample BCR0823:
5. Discussion
5.1. Metamorphic facies and paleotemperature indicators in the studied units
As both the Pethei Gp and Kasegalik Fm are predominantly calcareous units, estimates of the metamorphic grade are challenging. In samples where pelitic material is present (e.g., the Douglas Fm), prograde minerals such as muscovite constrain peak metamorphic conditions to a minimum of prehnite-pumpellyite facies (∼250°C), which includes a weaker retrograde metamorphic overprint in the form of disseminated chlorite replacement of hematite. Kasegalik dolomitic stromatolites are notably enriched in siliciclastic minerals in detrital-rich layers such that accessory metamorphic muscovite is more common.
Carbonate twinning can also be used as an approximate paleotemperature indicator in relation to deformation episodes at small (but ≥15 MPa) stresses. Calcite twin lamellae form and evolve as a function of deformation temperature, with a positive correlation between twin width and peak metamorphic temperature (Burkhard, 1993; Ferrill et al., 2004). In the studied units, carbonate twinning is often limited to type I and type II (thin and thick tabular twins, respectively), which independently suggest a peak temperature of 150–300°C. However, the patchy appearance of type III (lensoid and curved thick twins) in the Kasegalik suggests greater postdepositional strain occurred in this unit, syn-tectonically with recrystallization >200°C.
Finally, Raman spectra for OM in both the Kasegalik Fm and Pethei Gp represent highly disordered OM (Schopf et al., 2005) (Fig. 6B). Using the Lahfid model (Lahfid et al., 2010), we calculated the Kasegalik peak metamorphic temperature to be ∼242 ± 30°C and the Pethei at ∼198 ± 30°C (Table 4). There exists a small distribution in temperatures, both between stratigraphic horizons as well as between targets within the thin sections (Table 4). The combination of three independent paleotemperature results, namely metamorphic accessory minerals, calcite twinning, and Raman spectra of OM, all yield a consistent interpretation for the peak temperature of the Kasegalik Fm and the Pethei Gp occurring within prehnite-pumpellyite facies metamorphism.
5.2. Biosignatures in late Paleoproterozoic dolomitic stromatolites
A compilation of identified biosignatures and their occurrence in the studied thin sections is summarized in Table 6. Although the presence of OM may act as a potential biosignature in and of itself (Horita, 2005), the spatial association of OM with key mineralogical and sedimentological biosignatures is required to bolster confidence in interpreting a biotic origin for such features. All thin sections show a similar degree of biotic influence due to the extensive occurrence of microbial structures (both macro- and microscopic), association of OM with diagenetic minerals, and isotopic biosignatures (Table 6). In this study, a range of possible biosignatures are identified, but we emphasize it is incomplete. Although many biosignatures can be created through known abiotic processes, the cooccurrence of multiple signatures (Table 6) might still suggest a shared, biotic origin. Two confidence levels for biological influence, “Medium” and “High,” have been assigned based upon the total number of individual biosignatures irrespective of the density of such features (Table 6).
Summary of the Presence of Biosignatures in the Studied Thin Sections and Overall Confidence in Biological Influence
Confidence in biological influence is based on the variation of different types of biosignatures present—where samples with a greater variety of biosignatures have the most confidence in their biological influence. Here, “Medium” confidence ≥3 biosignatures and “High” ≥5 biosignatures. Stromatolitic structures seen in field outcrops (Figs. 3 and 4) are stated as “Primary microbially derived structures” in the table. Blanks = not observed.
ND = no data; OM = organic matter.
In the Pethei Gp and Kasegalik Fm, a number of possible primary textual and diagenetic mineralogical features have been interpreted as characteristics influenced by the existence of microbial mats. Primary carbonate sedimentary structures include clotted textures, wavy laminations to irregular wrinkly ripples (Figs. 3B, 3E, and 4F–H), and sediment roughening sets (Fig. 7A, B). Such structures often correlate spatially with laminar OM concentrations in the carbonate groundmass (Fig. 8E, F). Micro-Raman images have been used to identify not only the presence of OM in the carbonate stromatolites (Figs. 8E, 8F, 9A, 9B, and 10A) but also a number of mineral associations created by the postdepositional oxidation of OM—many of which appear preferentially constrained to stromatolitic laminae. This section identifies and discusses three major kinds of possible biosignatures: (1) diagenetic organomineral assemblages, (2) C and S isotope fractionation, and (3) primary sedimentary textures.
5.2.1. Diagenetic organomineral biosignatures
In the Pethei Gp dolomites, disseminated ferric-ferrous oxide generally shows the strongest association with OM. Within sparry interlaminations, authigenic ferric-ferrous oxide grains are often correlated with detrital-enriched laminations in the basal Pethei Group, and may share a dual origin between biotic influences and/or abiotic diagenetic mineral reactions, including the oxidation of pyrite. In the Kasegalik stromatolites, small (1–2 μm) pockets of poikilotopic OM occur in euhedral ferric-ferrous oxide crystals (Fig. 9A, B), especially in areas of spongy overgrowth. Some Kasegalik stromatolitic laminations feature high concentrations of ferric-ferrous oxide that often cap clotted fabrics, formed of coarse dolomite stained with finely disseminated OM (Fig. 7A, B). Such horizons may represent mat-like structures, which have later been buried and acted as sites for diagenetic sulfate reduction and the production of pyrite. Postdepositional diagenetic oxidation of OM may have occurred in association with ferric iron reduction, which results in the precipitation of carbonate as well as ferric-ferrous oxide minerals.
In the Kasegalik domal cherty stromatolites (Fig. 4H), subhedral crystalline pyrite is identifiable as isolated crystals or within laminae-parallel bands (Fig. 7B). Pyrite shows close spatial associations with OM, fluorapatite, and ferric-ferrous oxide. The spectra of pyrite (Fig. 10C) and ferric-ferrous oxide (Fig. 9E) in association with OM suggest that OM was diagenetically oxidized with ferric Fe, a process known to produce ferrous Fe in abiotic spheroids and microbial precipitates (Köhler et al., 2013; Picard et al., 2015). This reaction could have been abiotic (although the expected spheroidal concretionary morphology was not produced) or biotic from heterotrophic ferric Fe reduction. Pyrite is known to weakly absorb later forming organics onto crystal faces (Lindgren et al., 2011). Hence, rare OM coatings on pyrite grains (Fig. 10A–C) may indicate insignificant remobilization of OM following deposition.
Anatase is commonly associated with OM in the Pethei Gp, especially in branching multifurcate columnar stromatolites of the Wildbread Fm (Fig. 3F, G), but less common in the Kasegalik Fm. OM often occurs in diagenetic mineral relationships with ferric-ferrous oxide and anatase, both of which form fine-grained disseminations through the dolomite groundmass. The common presence of anatase and minor rutile in the Kasegalik Fm is comparable with observations in stratigraphically overlying stromatolites from the McLeary Fm (Gabriel et al., 2021). Biotic weathering of ilmenite (FeTiO3) is known to result in the mineralization of hematite and anatase (Schroeder et al., 2004; Bower et al., 2015), which might explain higher anatase concentration in the siltier, detrital-rich Pethei Gp carbonates. Environmental studies on TiO2 nanoparticles found mobilized TiO2 preferentially coalesced and adsorbed onto OM (Danielsson et al., 2017; Luo et al., 2018). Although this process was only tested in aqueous solutions, it is possible that early forming diagenetic nanocrystalline anatase could have been mobilized in pore waters and preferentially congregated (and later mineralized) around existing OM concentrations. Similarly, studies also show that amino and nucleic acids will preferentially bind to rutile (TiO2) surfaces (Jonsson et al., 2009; Cleaves et al., 2010). Results may differ in anatase, however, which has been shown to have a photocatalytic bactericidal effect in aqueous oxygenic settings (Dunlop et al., 2002; Glamoclija et al., 2009).
The interpretation for diagenetic mineralization of anatase on pre-existing OM is supported by observations of anatase commonly associated with phosphatized microfossils in the ∼1 Ga Torridon Group, Scotland (Sirantoine et al., 2021). In the Torridon Gp, anatase was interpreted to nucleate within cell complexes during postmortem breakdown, with crystal growth fueled by mobilization of Ti from surrounding Ti-enriched labile OM during very early diagenesis. A similar process may have occurred in the Pethei Gp stromatolites and as such, the distribution of anatase may match the original location of microbial cells. This suggestion is supported by clouds of acicular anatase in spatial association with OM (Fig. 10A), similar to TiO2 needles in McLeary stromatolites (Gabriel et al., 2021), as well as spherical micron-sized grains on anatase (Fig. 10B), which may relate to original cellular material.
Fluorapatite was only rarely seen as an accessory mineral in Kasegalik cherty domal stromatolites. The association between apatite and OM as derived from biomass has been documented in detail in stromatolitic phosphorite (Papineau et al., 2016) and in banded iron formations (Dodd et al., 2019). While there are abiotic occurrences of apatite with OM in mantle rocks, as well as in martian meteorites (e.g., McKay et al., 1996), this mineral association in the Kasegalik stromatolitic dolomite suggests that the occurrence of OM associated with ferric-ferrous oxide likely originated from biomass oxidation.
5.2.2. Isotope fractionation as a potential biosignature
Carbon and oxygen isotopes from carbonate in both the Kasegalik Fm and Pethei Gp (Table 2) suggest an equilibrium in the carbon cycle balanced by CO2-fixing oxygenic phototrophs during the deposition of the carbonate ramp sequence, after the Lomagundi-Jatuli Event (Karhu and Holland, 1996). In agreement with similar δ13Ccarb data collected by Hotinski et al. (2004), the δ13Ccarb values in the Wildbread dolomite vary from +1.3‰ to +2.4‰ within the upward-shallowing carbonate sequence (Fig. 2A and Table 2). We agree with their interpretation that this suggests some biological pumping and variable, low levels of primary productivity. OM in Kasegalik stromatolites have a large range of δ13Corg values (Table 2), which are broadly consistent with a number of C-fixation pathways, including CO2-fixing oxygenic phototrophs (House et al., 2003), whereas the δ13C and δ18O values of dolomite indicate a rather normal C-cycle with a minor overprint from diagenesis and metamorphism. Similarly, the near-zero δ13Ccarb of the studied stromatolitic dolomite (Fig. 2A, B) indicates deposition during a period of relative stability of the carbon cycle. The small +2‰ δ13Ccarb excursion in the Pethei and near-zero δ13Ccarb in the Kasegalik (Fig. 2B and Table 2) do not preserve isotopic evidence for biotic OM oxidation. However, oxidation is suggested sedimentologically by diagenetic mineral associations created by oxidation reactions, including ferric-ferrous oxides and pyrite associated with OM (Figs. 9E and 10C). Low metamorphic grades means there has been insignificant alteration of δ13C values, although systematically low δ18Ocarb values (Fig. 2) suggest some diagenetic fractionation of oxygen isotopes by meteoritic water.
The highly fractionated and 34S-enriched pyrite in Kasegalik domal stromatolites (Table 3) indicates microbial sulfate reduction under low sulfate concentrations, likely <200 μM (Habicht et al., 2002). Such heavy δ34S values are consistent with some data collected from euhedral pyrite from stromatolitic dolomite in the Rantamaa Fm, a slightly older Paleoproterozoic unit from the Fennoscandian shield dominated by 13C-enriched stromatolitic dolomite (Papineau et al., 2005). The possibility of abiotic Rayleigh-type distillation isotope fractionation process during evaporative precipitation is not favored over microbial sulfate reduction, as the former does not explain the common association of OM with pyrite and ferric-ferrous oxides in Kasegalik stromatolites (Figs. 9A–E and 10C). The correlation of idiotopic dolomite with OM-enriched horizons and diagenetic minerals including pyrite and ferric-ferrous oxide (Fig. 9C, D) indicates that surface dolomite mineralization may have resulted from a number of possible processes including bacterial sulfate reduction under low seawater sulfate conditions—as suggested by the 34S-enriched pyrite (Table 3). Biological protodolomite precipitation in surface environments (i.e., <50°C) is thought to be a result of microbial activity sustaining high pH and alkalinity required for crystallization, with sulfur reducing bacteria hypothesized to be capable of mediating dolomite formation (Van Lith et al., 2003; Liu et al., 2020).
5.2.3. Sedimentary structures and textures as potential biosignatures
Patchy metamorphic recrystallization of dolomite in the studied units means that areas of original primary dolomite have been preserved (Fig. 7E, F). At low surface temperatures, primary dolomite grows to form euhedral crystals and an idiotopic texture up to a “critical roughening temperature” ∼50°C (Gregg and Sibley, 1984; Huang et al., 2014), above which crystal growth produces nonplanar anhedral crystal shapes and a xenotopic groundmass texture. As such, idiotopic masses may be used to identify original depositional features formed within primary protodolomite—confirming the origin of primary ornamented and clotted sedimentary microstructures (Fig. 7E, F). The Pethei Gp displays a diverse range of stromatolitic dolomite morphologies, particularly within the columnar stromatolites of the McLean Fm (Fig. 3A–C). Idiotopic mosaics form sublaminar horizons parallel to stromatolitic laminations, preferentially associated with alternations enriched in detrital siliciclastics and hematite (Fig. 7F). In comparison with amorphous xenotopic dolomite overprints, idiotopic dolomite masses form laminar, subspherical, and rounded sedimentary structures (Fig. 7E). The Kasegalik Fm is notably different with dolomite that typically forms mosaics of anhedral microsparite hosting dense poikilotopic OM inclusions (Fig. 8E, F).
The microtexture of Kasegalik stromatolites is primarily controlled by laminar alternations in the concentration of micrite versus cherty dolomitic microsparite (Fig. 7B). Unlike the Pethei Gp, the Kasegalik Fm features much higher porosity, notably in the form of anastomosing columns (Fig. 4C, D) and centimeter-size euhedral gypsum crystals parallel to stromatolite beds (Fig. 4G). The latter independently confirms the availability of sulfate in this environment for bacterial sulfate reduction. Micritic dolomite in the Kasegalik commonly features sediment roughening between laminations (Fig. 7B), involving progressive convolution of laminae to produce clotted structures, suggestive of trapping and binding through microbial mat growth (Gebelein, 1972). Microbial mats in the Kasegalik Fm often feature bands of detrital minerals in interlaminations along with convoluted structures and ferric-ferrous oxide laminations (Fig. 7A, B), possibly formed as a result of sulfate reducing bacteria.
Mineralized balls (Fig. 12A–F) are a potential diagenetic biosignature. Ferric-ferrous oxide phases and OM are often associated with, and commonly occur within, mineralized balls (Figs. 8C, 8D, and 12B). Mineralized balls may represent possible abiotic biomorphs that could have formed due to the diagenetic oxidation of biotic OM, which has been proposed to create spheroidal diagenetic mineral structures that can be associated with anatase, ferric-ferrous oxide, and OM (Papineau et al., 2017). However, similar spherical features can also form from a variety of other abiotic processes, including gas bubbles (Bosak et al., 2010) or as rounded intraclasts. This intraclast interpretation is supported by their distribution and clast-like character (Fig. 8C), including apparent fracturing and disintegration of some spheres, consistent with allochthonous intraclastic grains rather than concretionary structures (Figs. 8C, 8D, and 12C). In the Kasegalik Fm, however, mineralized balls most commonly appear as amorphous blobs without clearly defined edges (Fig. 12A, B, E, F). While it is possible that microbially derived OM could have helped to bind detrital minerals, it remains unclear why the observed mineralized ball size distribution is weakly bimodal (Fig. 11), but a size variation spanning one order of magnitude is certainly inconsistent with an origin as microfossils.
Siliceous botryoids present within multifurcate and columnar cherty stromatolites in the Kasegalik (Fig. 13) provide further evidence consistent with the diagenetic oxidation of OM. Equidistant and circularly concentric fine laminations occur within radially aligned acicular quartz (Fig. 13D), centered around concentrations of OM, ferric-ferrous oxide, and micron-size dolomite (Fig. 13H). Such combination of geometric patterns (Fig. 13B–D, G) and mineralogical chemical composition (reactants and products) is suggestive of chemically oscillating reactions during precipitation—with identical morphology of concentric laminations to those developed by the Belousov-Zhabotinsky (B-Z) reaction (Papineau, 2020) (Fig. 13B, C, E). The B-Z reaction produces circularly concentric, radially expanding chemical waves (Fig. 13E) after the addition of a ferroin redox indicator, which catalyzes the oxidation of carboxylic acids (Papineau et al., 2017). The coexistence of ferric-ferrous oxide in association with dolomite and OM in the Kasegalik botryoidal centers (Fig. 13H, I) suggests that abiotic decarboxylation of biotic OM has occurred in association with the reduction of ferric iron during diagenesis (Dodd et al., 2018). Silica botryoids may thus be an example of a biosignature (biotic OM) located inside an abiotic biomorph (botryoid). This type of signature is comparable with opaline silica cement laminations preserved in the late Paleoproterozoic Sokoman Fm, which has also been interpreted to be early silica precipitation around grains, contributing to the preservation of microfossils within fenestrae (Maliva et al., 2005).
5.3. Widespread oxidation of OM during the late Paleoproterozoic
Both the Pethei Gp and the Kasegalik Fm feature stromatolitic dolomite that was deposited contemporaneously with the SFE, a negative excursion in δ13C in OM and some carbonate that occur alongside massive contemporary deposits of OM (Mancuso et al., 1989; Melezhik et al., 2005; Kump et al., 2011). Atmospheric oxygen reached high concentrations during this period, which resulted in the oxidation of formerly deposited OM and pyrite in the aftermath of the GOE (Ouyang et al., 2020). Biomass oxidation was widespread in Gabon, northwest Russia, and may represent a global mass oxidation event at this time (Kump et al., 2011). The near-zero δ13Ccarb values of dolomite in both the Kasegalik Fm and Pethei Gp (Table 2) are consistent with their deposition after the GOE when the δ13C value of seawater returned to normal near-zero values after 2.06 Ga (Karhu and Holland, 1996; Martin et al., 2013). Red beds in the Kasegalik Fm are consistent with increased environmental oxygen at this time (Fig. 4E), as well as evidence of abiotic OM oxidation in the form of siliceous botryoids (Fig. 13H, I).
5.4. Application of biosignatures in stromatolitic dolomite to martian carbonates
Multiple lines of evidence in the form of morphological similarities to outcrop-scale sedimentary structures, evidence for the occurrence of OM, and the existence of biosignatures (Table 6) are needed to confidently infer the biotic origin of stromatolitic horizons—both on Earth and on other planetary bodies. Since microscopic stromatolitic sedimentary textures and mineral relationships are identifiable through combined approach of RL microscopy with micro-Raman spectroscopy (Figs. 8E, 8F, 9A–E, and 10A–C), we suggest that similar inferences can be achieved if similar datasets are generated by robotic instruments, for instance by the SHERLOC instrument on the NASA Perseverance rover (Wiens et al., 2017). Inclusion of micro-X-Ray Fluorescence data would further complement comparisons with the datasets produced on Precambrian specimens and by martian rovers, hence this represents a future direction of this work. Regions such as Isidis and Chryse planitia (chosen as landing sites for ESA and NASA rovers) are good candidates for past martian carbonate deposits based on their geographical location. Both basins occur near the martian highland/lowlands boundary, interpreted as a paleoshoreline of a past martian northern ocean (Niles et al., 2013). There is significant evidence for past liquid water preserved in the martian geological record (Boston et al., 1992), and it has been hypothesized that microbial mats could have thrived on a wet martian surface ∼3.8–3.1 Ga (Ellery et al., 2004). This time bracket for a watery surface matches the formation ages of the Isidis Planitia “shoreline” 3.5 Ga (Citron et al., 2018) and Chryse Planitia mass-flow deposits before 3.6 Ga (Adler et al., 2019). Both Isidis and Chryse planitia contain deltaic types of paleoenvironments that include eroded siliciclastic and volcanically derived clastic deposits on a sedimentary ramp with the potential for intermittent carbonate environments (Mustard et al., 2009; Adler et al., 2019). Basement outcrops from crater lake paleoshorelines are some of the best targets to find stromatolites on Mars. Mg-rich carbonate has already been identified as a bedrock lithology in exposures adjacent to Isidis basin (Ehlmann et al., 2008).
The Pethei Gp was deposited syn-kinematically with the Wopmay orogeny ∼1.92–1.97 Ga within the Athapuscow aulacogen, which forms a continental margin-type environment. Similarly, Isidis Planitia exhibits a potential shoreline geomorphology along the martian highlands contact, including deltaic deposits in the Jezero crater region, valley networks, and single channels—suggesting past water drainage into the basin from highlands to the south (Scott et al., 1995). Cliffs exist at the boundary between the Libya Montes, with evidence of terraces that suggest wave-cut action (Erkeling et al., 2012). Curvilinear ridges within the Isidis basin have been compared with barrier ridges and spits (Ghatan and Zimbelman, 2006). Furthermore, alluvial fans have been identified on the Isidis basin floor, although it is uncertain whether these were subaqueous during deposition (Erkeling et al., 2012).
The Isidis basin originated as an impact crater (Mustard et al., 2009), leaving an arcuate embayment that shows similarities to the Nastapoka Arc region in Québec, Canada (Fig. 1A, D), a feature that has been hypothesized as an astrobleme (Beals, 1968; Hofmann, 1976). Although lithological, mineralogical, and structural indicators of shock metamorphism have yet to be reported, the perfect circularity of the Nastapoka Arc may be the result of an older Archean impact structure later modified by deep weathering and by the Trans-Hudson orogeny (Goodings and Brookfield, 1992). The Kasegalik Fm could thus represent a deep basin facies comparable with water filled impact craters on Mars. The paleocurrent direction in the neighboring ∼2.025 Ga Pachi Fm (Nastapoka Gp, Richmond Gulf Supergroup) suggests an embayment within a coastline remarkably similar to the present Nastapoka shoreline (Chandler, 1984; Chandler and Parrish, 1989). The Belcher Gp paleoenvironment was a shallow marine environment offshore from the mouth of the Richmond Gulf with estuarine input of nutrient-rich water sourced from eroded granitoids of underlying Archean basement. This paleogeography is comparable with paleoshore locations around Isidis basin and Chryse Planitia, both of which contain fan-shaped deposits hypothesized to be an extensive deltaic environment (Erkeling et al., 2012; Adler et al., 2019).
Although widespread carbonate lithologies are generally missing from the martian geological record (Bibring et al., 2005; Niles et al., 2012), the correlation of deltaic activity with minor bedrock carbonate in the Jezero Crater (Ehlmann et al., 2008) and with subaqueous sedimentary structures on Meridiani Planum (Squyres et al., 2004) suggests that intermittent carbonate deposition might have taken place, and that such paleoenvironments could have been inhabited by stromatolite-forming microbial communities. These martian locations share paleoenvironmental and mineralogical similarities with the studied late Paleoproterozoic localities, and we suggest that the biosignatures identified in this work (Table 6) are applicable for the search for evidence of life on early Mars in these locations and beyond.
6. Conclusions
This study reports a number of biosignatures in dolomitic stromatolites from a carbonate platform and a basal ramp setting. Both units have been weakly metamorphosed around the prehnite-pumpellyite facies (Pethei Gp ∼198 ± 30°C vs. Kasegalik Fm ∼242 ± 30°C). Micro-Raman images of sedimentological features therein reveal the presence of OM associated with key diagenetic minerals in both the Kasegalik Fm and Pethei Gp stromatolitic dolomite. In both units, OM displays a systematic correlation with diagenetic pyrite, ferric-ferrous oxides, and anatase. These mineral associations often occur in complex, clotted sedimentary microstructures that independently suggest a biotic origin. A number of potential microscopic biosignatures are identified and described. Alone, each biosignature provides a low level of confidence for a biotic origin, but in the context of well-preserved dolomitic stromatolites, multiple biosignatures can be combined to yield a medium-to-high level of confidence in the biotic origin of these rocks.
In this study, three categories of biosignatures are identified. First, microbial sedimentary textures occur as (1) outcrop-scale and microscopic primary microbially derived structures, (2) idiotopic dolomite textures in association with OM, (3) mineralized balls, and (4) botryoidal silica in association with OM. A second class of biosignatures exists in the form of disseminated OM with diagenetic minerals, including (5) TiO2 polymorphs, (6) ferric-ferrous phases, (7) apatite, and (8) pyrite. Finally, isotopic signatures can also be biosignatures in the form of (9) C-isotope fractionation and (10) S-isotope fractionation. Each of these biosignatures has been shown in different context to be potentially formed by abiotic processes; hence, they remain possible signs of life. However, the spatial relationships of OM with microscopic sedimentary structures and minerals in their petrological and paleoenvironmental context (i.e., in field outcrops) considerably improve confidence in the biotic origin of the studied dolomitic stromatolites.
Combined with microscopic observations and the geological context of outcrops, C- and S-isotope compositions yield useful criteria to infer the presence of microbial metabolisms such as oxygenic photosynthesis and sulfate reduction in these late Paleoproterozoic carbonates. All studied samples were identified as biotic in origin with either a “medium” or “high” confidence level based upon the total number of different, independent biosignatures present. Hence, we imply the past formation of stromatolite structures by microbial communities in these carbonate ramp environments. Mineralogical and sedimentological evidence for OM oxidation in some studied thin sections is also consistent with the contemporaneous SFE.
The 10 types of biosignatures identified may prove useful in future studies for comparisons with other late Paleoproterozoic successions, as well as for various locations on Mars where similar dolomitic units may occur (such as Isidis and Chryse Planitia). Raman spectroscopy is a useful method in conjunction with optical microscopy to map the spatial distribution of OM associated with sedimentary structures and diagenetic minerals, which is something currently achievable through robotic missions. This combined strategy can yield high levels of confidence in concluding a biological influence of microbial mats in ancient lithologies, including extraterrestrial rock outcrops.
Footnotes
Acknowledgments
W. Bleeker, R. Mitchell, and D.A.D. Evans are thanked for discussions and facilitating access to the Great Slave Lake field localities. A.-L. Jourdan is thanked for her work on the carbonate isotope data from the Great Slave Lake. Three reviewers are also thanked for their constructive comments that helped improve this paper. The late E. Hauri is acknowledged for his help with the NanoSIMS analyses performed in this project.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
D.P. acknowledges the Carnegie Institution for Science and the Carnegie of Canada for financial support to visit the Great Slave Lake and the specific Belcher Islands locality described in this work, as well as the University College London for supporting analytical work.