
Abstract
Finding evidence of life beyond Earth is the aim of future space missions to icy moons. Icy worlds with an ocean underlying the icy crust and in contact with a rocky subsurface have great astrobiological interest due to the potential for water-rock interactions that may provide a source of nutrients necessary to sustain life. Such water-rock interactions in icy moons can be indirectly investigated using analogous environments on the deep seafloor on Earth. Here, we investigate the presence of molecular and isotopic biomarkers in two submarine cold seep systems with intense rock-fluid interactions and carbon sink as carbonates with the aim of gaining understanding of potential carbon cycles in the icy worlds' oceans. Authigenic carbonates associated to cold seeps (a chimney from the Gulf of Cádiz and a clathrite from the Pacific Hydrate Ridge) were investigated for their mineralogical composition and lipid biomarker distribution. Molecular and compound-specific isotopic composition of lipid biomarkers allowed us to infer different carbonate origins in both carbonate scenarios: biogenic methane (clathrite) versus thermogenic methane together with allochthonous carbon (chimney). In the Pacific cold seep, carbonate precipitation of the clathrite was deduced to result from the anaerobic oxidation of methane by syntrophic action of methanotrophic archaea with sulfate-reducing bacteria. The distinct carbon sources (thermogenic methane, pelagic biomass, etc.) and sinks (gas clathrates, clathrite, chimney carbonates) were discussed in the light of potentially similar carbon cycling pathways in analogous icy-moon oceans. We show how the isotopic analysis of carbon may be crucial for detecting biosignatures in icy-world carbon sinks. These considerations may affect the strategy of searching for biosignatures in future space missions to the icy worlds.
1. Introduction
Icy moons of our solar system (e.g., Europa, Enceladus, and Titan) are potential targets for future space missions, due to the astrobiological interest of their internal oceans as potential habitable environments fulfilling some of the requisites for sustaining life (Hand et al., 2017; Taubner et al., 2018, 2020). The Cassini-Huygens mission revealed the presence of geological activity in the internal rock mantle—possibly formed by partially altered chondritic rocks according to its high relative bulk density (Sekine et al., 2015)—connected to the surface of Enceladus, where many jets of ice particles emerge from the south polar region (Porco et al., 2006). The chemical composition of the plumes was analyzed in situ with the Ion and Neutral Mass Spectrometer (INMS) on board the Cassini spacecraft and revealed the presence of water (H2O), methane (CH4), carbon dioxide (CO2), ammonia (NH3), molecular nitrogen (N2), and molecular hydrogen (H2) (Waite et al., 2017). A comparable activity may occur on Europa, according to observations from the Hubble Space Telescope and the Galileo spacecraft, indicating that plumes emerge currently from the equatorial and southern latitudes of the jovian satellite (Sparks et al., 2016, 2017; Jia et al., 2018).
Potential geochemical interactions between the liquid water and the rocky core in those moons are assumed to be at temperatures from low to moderate (<100°C) (Sekine et al., 2015; Waite et al., 2017). On Enceladus, the hydrogen and methane found in the jet plumes emerging from the surface could be generated in the satellite interior through serpentinization of the olivine present in the rocky core (Sleep et al., 2004; Kotlarz et al., 2020). In contrast, CO2 could derive from the seafloor degassing (Glein et al., 2015). If carbon sequestration occurs in Enceladus' ocean, the composition of carbon in the final sequestered form may help constrain its origin. Glein and Waite (2020) propose carbonation as a likely process to sequestrate CO2 as ferromagnesite through reactions in the serpentine-rich interior, while Bouquet et al. (2015) considered gas clathrate layers as possible intermediate carbon reservoirs.
Methane and to a lesser extent carbon dioxide are gases commonly released in submarine cold seeps on Earth. Here, we refer to cold seeps as any seepage at low-temperature conditions in oceanographic settings released from the rocky layer–ocean interface independent of the tectonic context. Methane leakage may be sourced from not only subsurface gas-bearing reservoirs (Le Bris et al., 2016) but also active low-temperature serpentinization of ultramafic rocks (Etiope and Sherwood-Lollar, 2013), such as those at Pomonte-Ogliera seeps in the Tuscan Archipelago (Italy) where abiotic methane-rich emissions related to Fischer-Tropsch type reactions originate from serpentinizing ophiolites (Sciarra et al., 2019). Upon favorable conditions of pressure and temperature, these gases may be trapped in the form of gas clathrates, as observed in the Cascadia continental margin (Elvert and Suess, 1999; Baumberger et al., 2018) or trapped in sediments as in shallower deposits of the Gulf of Cádiz (e.g., Mazurenko et al., 2003; Pinheiro et al., 2003). The dissociation of gas clathrates in turn may liberate carbon-bearing gasses that, upon saturation, may precipitate in the form of carbonates, where microorganisms may play a role. Riding and Virgone (2020) classified methane-derived authigenic carbonates as hybrid carbonates because their precipitation takes place through combinations of abiotic processes, bio-induced and even bio-controlled mineralization. In seafloor hydrocarbon seeps in Earth's oceans, biological activity is typically associated to chemolithotrophic metabolisms, whose energy source for cell biosynthesis and maintenance comes from the reduction of inorganic substances (e.g., sulfate, nitrate, or oxidized metals) (e.g., Beal et al., 2009; Himmler et al., 2018). In these systems, the production of biological methane (methanogenesis) is typically coupled to its consumption (methanotrophy) through anaerobic oxidation (e.g., Orphan et al., 2001; Elvert et al., 2003; Chevalier et al., 2011). A consortium of methanotrophic archaea and sulfate-reducing bacteria (SRB) mediates the process of anaerobic oxidation of methane (AOM) (Hoehler et al., 1994). The AOM activity produces bicarbonate that increases the water alkalinity and eventually induces the precipitation of authigenic carbonate (Ritger et al., 1987; Aloisi et al., 2002; Suess, 2014; Sun and Turchyn, 2014). These methane-derived authigenic carbonate deposits constitute geological archives retaining information about biogeochemical processes and associated metabolisms occurring in terrestrial cold seeps (Peckmann and Thiel, 2004; Campbell, 2006; Zwicker et al., 2018).
Molecules such as 2,6,11,15-tetramethylhexadecane (crocetane), 2,6,10,15,19-pentamethyleicosane (PMI), unsaturated C25 isoprenoids, archaeol, or sn-2-hydroxyarchaeol are characteristic biomarkers of methanotrophic and methanogenic archaea (Tornabene and Langworthy, 1979; Pancost et al., 2000; Thiel et al., 2001). Other compounds such as fatty acids with terminal branches (e.g., iso- and anteiso-C15) or monounsaturations (e.g., C16:1ω7) are rather linked to SRB (Thiel et al., 2001; Elvert et al., 2003). The compound-specific isotopic composition (δ13C) of all those molecular biomarkers may provide information about the metabolic pathway they are resulting from. As methane is highly depleted in 13C (δ13C of thermogenic methane ∼ -50‰, and δ13C of biogenic methane is from ∼ -60 to -110‰; Whiticar et al., 1986), its uptake by microorganisms involved in the precipitation of carbonates in cold seeps can be easily recognized in very depleted δ13C values. Thus, compound-specific isotopic analysis of diagnostic lipid biomarkers preserved in authigenic carbonates can serve as fingerprints of archaeal-bacterial consortia involved in sulfate-dependent AOM (Thiel et al., 2001; Peckmann and Thiel, 2004; Chevalier et al., 2011). Molecular and isotopic fingerprints of AOM have been explored in carbonates from cold seeps to trace present and past biological activity (e.g., Elvert and Suess, 1999; Peckmann and Thiel, 2004; Merinero et al., 2012; Prouty et al., 2020), with a major focus on determining the sources of methane-derived carbonates. Despite the astrobiological interest of submarine venting systems (Barbieri and Cavalazzi, 2004; Merinero, 2008), no studies have been devoted to interpreting molecular and isotopic tracks of carbon on a global cycling perspective with application to ocean environments in icy worlds so far.
Here, we investigate the biological fingerprints retained in methane-derived authigenic carbonates from two distinctive cold seeps venting methane-rich fluids situated offshore on the Pacific (Cascadia Margin, Oregon) and the Atlantic (Gulf of Cadiz, Spain) oceans. We applied a biogeochemical approach involving the use of lipid molecular biomarkers, compound-specific isotope analysis, bulk geochemistry, and mineralogy to characterize organically and inorganically the two forms of authigenic carbonates (chimney vs. clathrite). We wanted to elucidate the different carbon sources contributing to the eventual precipitation of carbonates (i.e., carbon sinks) and the role of gas clathrates in the two cold seeps, with the aim of understanding local carbon cycling in submarine gas venting systems. The stable carbon isotopic information retained in the two methane-derived authigenic carbonates allowed us to constrain different origins of the carbon sequestered. The ultimate aim is to apply this knowledge to elucidate how carbon cycling could be functioning in low-temperature seafloor venting systems on icy moons from the Solar System. Tracking the carbon sources and sinks in submarine methane-cold seeps on Earth may provide clues for the search for biosignatures in future space missions to the icy worlds.
2. Materials and Methods
2.1. Geological settings and sample collection
Methane-derived authigenic carbonates from two submarine cold seeps were provided and studied for their organic and mineralogical composition. A clathrite was collected from TV-grab 18 at the southern summit of Hydrate Ridge in the Cascadia Margin (Pacific Ocean, Oregon coast) in 1996 during cruise SO110-1 and a carbonate chimney from the TASYO field situated in the Gulf of Cadiz (Atlantic Ocean, SW of Spain) during the TASYO-2000 cruise (Fig. 1a). Both submarine environments are different in terms of geological context and source of methane seepage, resulting in two different carbonate structures (i.e., chimney versus clathrite).

Location of the two cold seep settings (
The Hydrate Ridge is located in the accretionary wedge of the Cascadia Subduction Zone, where the Juan de Fuca plate is subducting orthogonally beneath the North American plate (Fig. 1b). The accretionary prism is constituted by a fold and thrust belt that deforms a Plio-Quaternary sedimentary sequence composed of hemipelagic sediments interbedded with turbidites, volcanic ash, and debris (Flueh et al., 1998; Melgar, 2009). These deposits of continental origin are rich in phyllosilicates, such as detrital micas and chlorite-serpentines, derived mostly from the alteration of the Klamath ophiolites (Orr et al., 1992; Melgar, 2009; Alexander, 2014). One of its thrust ridges is known by the name of Hydrate Ridge (black square in Fig. 1b) because numerous gas hydrate deposits are found along its 25 km length and 15 km width (Greinert et al., 2001; Suess et al., 2001). The Hydrate Ridge is characterized by having two summits on the terminations of its north-south axis at 600 and 800 m depth, respectively (Fig. 1d). In these two summits, the greatest thicknesses of gas hydrate deposits are concentrated (Tréhu et al., 1999; Yu et al., 2006), and the major seepage activity is found. Methane ejections from both gas hydrate dissociation and gas trapped in the sediment are of biogenic origin (Heeschen et al., 2005). Unlike the mechanism of carbonate chimney formation, a clathrite forms just at the same site where gas hydrate dissociation takes places (Bohrmann and Torres, 2006).
The Gulf of Cadiz belongs to the Atlantic passive margin that comprises from Cape Säo Vicente in Portugal to the Rharb basin in Morocco (Fig. 1c). In this region, the vast majority of cold-seep systems are found in the middle section of low-gradient slopes where sandy contourite deposits and the underlying allochthonous unit of the Gulf of Cadiz acts as a gas reservoir (Llave, 2003; Medialdea, 2005). On the seafloor, methane venting is reflected in a variety of geomorphic structures such as pockmarks, mud volcanoes, mud diapirs, diapiric ridges, and authigenic carbonates that are also referred to as methane-derived authigenic carbonate (Vanneste et al., 2012). Methane-derived authigenic carbonate is composed of different carbonate minerals (calcite, aragonite, dolomite, and siderite) that show a range of morphologies (crusts, chimneys, irregular massive forms), but they all share the same internal structure. They are rocks with low porosity and clastic texture, where carbonate fraction constitutes micrite or sparite groundmass that contains a variable proportion of detrital grains. The detrital fraction consists of mineral grains mainly quartz and clays, bioclasts, peloids, and pellets (Magalhães et al., 2012; Wang et al., 2015). In addition, methane-derived authigenic carbonate frequently contains microcrystals of pyrite and iron (oxyhydr)oxides (Magalhães et al., 2012).
2.2. Mineralogical characterization
Mineralogy of both carbonate samples was analyzed with a Bruker X-ray diffractometer (AXS D8-Focus XRD). The dried and ground samples were scanned in the 2θ·diffraction angle from 5° to 70°, with a scanning step size of 0.01°, operated at 40 kV and 40 mA with a Cu X-ray source (Cu Kα1,2, λ = 1.54056 Å), similarly as described elsewhere (Carrizo et al., 2019). Infrared spectroscopy analysis was performed using diffuse reflectance infrared Fourier transform spectroscopy (DRIFTS) in a Thermo-Nicolet spectrometer. Spectra of 2 cm−1 of resolution and 64 scans were collected in the mid (4000–400 cm−1) and near (0.82–2.5 microns) infrared region, using a DTGS-KBr detector and a quartz beamsplitter.
In addition, thin sections were prepared from both samples, as well as a polished thicker section from the chimney sample, to study both carbonates under optical petrographic microscope with transmitted polarized (thin sections) or reflected (polished thick section) light.
2.3. Bulk isotopic geochemical characterization
Stable isotope analysis was conducted on carbon, both organic and inorganic (i.e., carbonates), and oxygen, by means of isotope-ratio mass spectrometry (IRMS). The isotopic composition of the organic carbon (δ13Corg) was measured at the Centro de Astrobiología facilities, following the USGS method (Révész et al., 2012). For δ13Corg, samples were homogenized by grinding with a mortar and pestle, decarbonated with HCl (3 N) and, after 24–48 h of equilibration, adjusted to neutral pH with ultrapure water. The decarbonated samples were then dried in an oven (50°C) until constant weight, then analyzed by a MAT 253 IRMS (Thermo Fisher Scientific, Waltham, Massachusetts, USA) and reported in the standard per mil notation (‰). Three certified standards (USGS41, IAEA-600, and USGS40) were used, providing an analytical precision for δ13Corg of 0.1‰. The isotopic composition of the samples for inorganic carbon δ13Cinorg and associated oxygen (δ18O) was determined at the Laboratorio de Isótopos Estables de la Universidad de Salamanca (Spain) according to their own protocols. Briefly, the bulk samples were subjected to reaction with 103% H3PO4, and the resulting CO2 was offline analyzed with a second dual-inlet SIRA-II mass spectrometer (McCrea, 1950; Craig, 1957). The content of total organic carbon (TOC %) was measured with an elemental analyzer (HT Flash, Thermo Fisher Scientific, Waltham Massachusetts, USA) during the δ13Corg analysis.
2.4. Lipid extraction, fractionation, and analysis
About 7 g of freeze-dried and ground samples of the carbonates were extracted with a mixture of dichloromethane/methanol (DCM/MeOH, 3:1, v/v) by ultrasound sonication (3 × 10 min cycles). Internal standards (tetracosane-D50, myristic acid-D27, 2-hexadecanol) were added prior to extraction. The concentrated and desulfured total lipid extract (e.g., Sánchez-García et al., 2018) was hydrolyzed overnight with methanolic KOH (6% w/w) at room temperature (Grimalt et al., 1992). The hydrolyzed total lipid extract was separated in two fractions by extracting with n-hexane (Hx), according to Grimalt et al. (1992). The neutral fraction was further separated into nonpolar (hydrocarbon) and polar (alkanols and sterols) fractions according to procedures extensively described elsewhere (Carrizo et al., 2019). The acidic and polar fractions were derivatized with BF3 in methanol (80°, 30 min) and BSTFA (80°, 60 min), respectively, prior to analysis by gas chromatography–mass spectrometry (GC-MS). The nonpolar fraction was directly analyzed by GC-MS.
The three lipidic fractions (i.e., nonpolar, acidic, and polar fraction) were analyzed using a 6850 GC system coupled to a 5975 VL MSD with a triple axis detector (Agilent Technologies) operating with electron ionization at 70 eV and scanning from m/z 50 to 650 (analytical details in Sánchez-García et al., 2020). Compound identification was based on retention times and the analysis of mass spectra against reference materials and compound databases (e.g., NIST v. 2.2). For quantification, we used external calibration curves of n-alkanes (C10 to C40), fatty acid methyl esters or FAMEs (C8 to C24), n-alkanols (C10, C14, C18, and C20), and branched isoprenoids (2,6,10-trimethyl-docosane, crocetane, pristane, phytane, squalane, and squalene), all from Sigma-Aldrich. Recoveries of the internal standards averaged 72 ± 23%.
2.5. Compound-specific isotope analysis of lipid biomarkers
Those lipid biomarkers detected in sufficient amount were analyzed for carbon isotopic composition by coupling a gas chromatograph (Trace GC 1310 ultra) to an IRMS system (MAT 253 IRMS, Thermo Fisher Scientific). For the GC analysis, the oven temperature program was set to gradually increase from 70°C to 130°C at 20°C·min−1 and then to 320°C at 10°C·min−1 (held for 30 min). Conditions for IRMS analysis were as follows: electron ionization of 100 eV, Faraday cup collectors of m/z 44, 45, and 46, and temperature of the CuO/NiO combustion interface at 1000°C. The samples were injected in a PTV injector in splitless mode, with an inlet temperature of 50°C, then a ramp temperature of 2.5°C s−1 until 320°C (held 2.5 min), using helium as carrier gas at constant flow of 1.1 mL·min−1. The δ13C value of individual lipids was calculated using CO2 spikes of known isotopic composition, introduced directly into the MS source, three times at the beginning and end of every run. Reference mixtures from the Indiana University (US) of known isotopic composition of n-alkanes (A6) and FAMEs (F8) were run every four samples to check the accuracy of the GC-IRMS determinations. For the carboxylic acids, δ13C was calculated from the value determined on the FAMEs, correcting them for the one carbon atom added in the methanolysis (Abrajano et al., 1994).
3. Results
3.1. Mineralogy and bulk isotopic geochemistry
X-ray diffraction (XRD) analysis indicated differences in the mineralogical composition of the two authigenic carbonates. The chimney-like carbonate consisted of low-magnesium calcite, high-magnesium calcite, Fe-rich dolomite, and quartz, whereas the clathrite was composed of aragonite, high-magnesium calcite, quartz, and chlorite-serpentine (Table 1). For the chimney carbonate, the mid-infrared (MIR) spectra allowed identification of two mineral groups (Fig. S1a). On the one hand, calcite exhibiting peaks at 725, 880, 1451, 1801, and 2518 cm−1 was the only representative mineral of carbonates. The first and second peaks corresponded to the CO3 2- symmetric bending ν4 and asymmetric bending ν2 modes, respectively (Jovanovski et al., 2005; Rodríguez-Blanco et al., 2011). A strong and broad band observed around 1451 cm−1 was interpreted as due to the CO3 2- asymmetric stretching ν3 mode (Bromiley et al., 2007; Comstock et al., 2019), while the sharp band at 1801 cm−1 was assigned to the CO3 2- symmetric stretching ν1 + ν4 mode (Rodríguez-Blanco et al., 2011). On the other hand, a number of bands with a peak at 800 cm−1 belonging to Si-O-Si symmetric stretching mode (Ojima, 2003) were attributed to the presence of quartz and illite-muscovite (Graeser et al., 2003; Ojima, 2003; Chukanov, 2014). In contrast, according to its MIR spectra, the clathrite (Fig. S1b) showed a predominantly aragonitic composition (Chakrabarty and Mahapatra, 1999), with the exception of the 880 cm−1 band of calcite.
Mineralogical Composition of the Two Authigenic Carbonates Based on X-Ray Diffraction (XRD) and Infrared Spectroscopy (MIR, NIR, and IMS)
This was confirmed by its near-infrared (NIR) spectrum (Fig. S2b) showing features characteristic of aragonite (Bishop et al., 2019). As for the chimney carbonate (Fig. S2a), a number of bands in its NIR spectrum were identified: 1.41 μm (OH stretching), 1.92 μm (H2O stretching + bending), 2.21 μm (Al-OH), and 2.34 (Mg-OH) μm, of which the second band is distinctive of montmorillonite (Loizeau et al., 2007; Gavin et al., 2010; Bishop et al., 2011).
The petrographic analysis of the samples showed differences between the two carbonates likely due to their different formation conditions (Fig. 2). The chimney carbonate exhibited reddish-brown color along its external and central hole surfaces and around scarce, larger pores (1–3 mm) (Figs. 1f and 2a). This is due to the precipitation of hematite, goethite, and (oxy)hydroxides. This sample showed an internal fabric characterized by an alternating mudstone/wackestone texture with quartz grains, bioclasts (e.g., foraminifera, gastropods, bivalve-fragments, or green seaweeds), peloids, intraclasts, and isolated glauconite and muscovite are found oriented according to main flux directions (Fig. 2b). Finally, the presence of pyritic framboids with an oxidized halo of goethite and hematite pseudomorphs of pyrite was also observed in the chimney carbonate, as infill of moldic porosity (Fig. 2b, 2c). In contrast, the clathrite showed a greater porosity (Fig. 1g) between its heterometric angular micrite clasts (Fig. 2d). Hence, it is a monogenetic intraformational breccia in which its intergranular porosity is filled partially or completely by radial-acicular aggregates of aragonite (Fig. 2d, 2e). In addition, aragonite crystals were also observed growing in contact with clasts or infilling fracture-like features or bioturbation-like structures, but they seem to have a botryoidal morphology more noticeable thanks to the opaque minerals that border them (Fig. 2e, 2f).

Thin sections of the two carbonate samples, of the chimney carbonate from the Gulf of Cadiz (a–c) and the other three of clathrite from the Hydrate Ridge (d–f) (scale bar: 100 μm). All the thin sections were taken in light transmitted under crossed polarized filters excepIc) that was taken under polarized reflected light. (
The TOC content in the two authigenic carbonates was measured to be as different as 0.5% in the chimney carbonate and 9% in the clathrite (Table 2). Their stable-carbon isotopic composition was also different, with the chimney sample showing a δ13Corg ratio of -22.6‰ and a δ13Cinorg ratio of -12.5‰, and the clathrite a δ13Corg of -49.1‰ and a δ13Cinorg of -44.8‰. In contrast, the stable isotopic composition of oxygen was rather similar in both carbonates, with δ18O ratios of 4.0‰ and 3.6‰ in the chimney and clathrite, respectively (Table 3).
Organic Geochemistry in the Two Authigenic Carbonates, with the Concentration of Total Organic Carbon (TOC, % of Dry Weight) and Molecular Lipid Biomarkers (ug·g−1 of Dry Weight)
n.d. stands for not detected.
PMI is 2,6,10,15,19-pentamethyleicosane.
Sum of monounsaturated isoprenoids with C25 chain length (two C25:1 isomers).
ACL is average chain length, calculated as Σ(iXi + … + nXn )/ ΣXi + … + Xn ), where X is concentration, i the smallest unit of carbon (i.e., 15), and n the largest (i.e., 29).
Sum of alkyl benzenes (n = 12; from Fig. S3).
Bulk Isotopic Composition (‰ Notation) of Oxygen (δ18O) and Organic and Inorganic Carbon (δ13C) in the Two Authigenic Carbonates, as Well as Compound-Specific Isotopic Composition of Carbon (δ13C) in Selected Lipid Biomarkers
n.d. stands for not detected.
Range of monounsaturated isoprenoids with C25 chain length (two C25:1 isomers).
3.2. Lipidic biomarkers and stable isotopic composition
A number of lipid biomarkers were found in the three polarity fractions from the authigenic carbonates (i.e., nonpolar, polar, and acidic), with distinct molecular patterns in the two samples. In the nonpolar fraction, the chimney carbonate exhibits a unimodal distribution of n-alkanes from C15 to C29, with dominance of the C18, C19, and C20 congeners (concentration of 0.42, 0.43, 0.41 ug·g−1, respectively), and lower presence (0.1 ug·g−1) of the isoprenoids pristane and phytane (Fig. 3a). A series of alkyl benzenes were also found in the nonpolar fraction (Fig. S3) at a total concentration of 0.32 ug·g−1. In contrast, in the clathrite, saturated isoprenoidal hydrocarbons such as crocetane (1.4 ug·g−1) and PMI (0.98 ug·g−1) were prevailing in the nonpolar fraction, whereas n-alkanes from C16 to C29 were a minority and similarly abundant among them. Pristane and two unsaturated hydrocarbons (C25:1) were only detected at low concentrations (Fig. 3b).

Total ion chromatogram of the hydrocarbon fractions extracted from the chimney carbonate (
In both carbonate samples, the acidic fraction was exclusively composed of low-molecular-weight (LMW) carboxylic acids. However, while the most abundant carboxylic acids in the chimney carbonate were the saturated, straight-chained 16:0, 14:0, and 12:0 moieties (Fig. 4a), in the clathrate, 16:0 and 18:0 were prevailing (Fig. 4b). In addition, both samples showed minority peaks of the terminally branched iso/anteiso-15:0, and the monounsaturated 16:1ω7 and 18:1ω9 congeners. In the clathrate, the amount of iso/anteiso-15:0, 16:1ω7, and 18:1ω9 were ∼7 and 30-fold those in the chimney carbonate (Table 2).
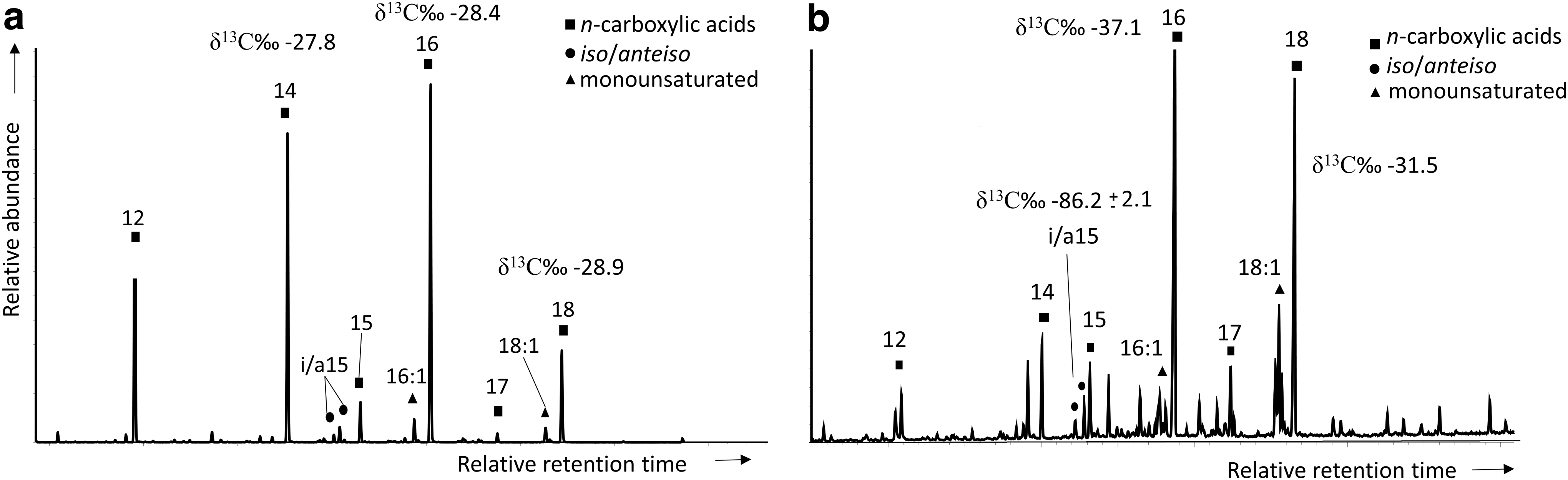
Total ion chromatogram of the acidic fraction extracted from the chimney carbonate (
The composition of the polar fraction was also different in the two carbonate samples. In the chimney carbonate, the LMW n-alkanols C14, C16, and C18 dominated the polar molecular distribution (range values 0.02 to 0.1 ug·g−1) together with dihydrophytol (0.34 ug·g−1) (Fig. 5a). In the clathrate in contrast, archaeol (11 ug·g−1), sn-2-hydroxyarchaeol (3.6 ug·g−1), and dihydrophytol (2.5 ug·g−1) were the prevailing polar compounds (Fig. 5b). Cholesterol was detected in both samples at relatively low concentration (Table 2).

Total ion chromatogram of the polar fraction extracted from the chimney carbonate (
The compound-specific isotope analysis of the lipid biomarkers from all fractions showed two groups of values: (i) one of relatively enriched δ13C ratios (i.e., from -24.9‰ to -37.1‰) generally measured in the chimney carbonate and in the LMW n-alkanes and n-carboxylic acids in the clathrate and (ii) the other of much more depleted δ13C values (i.e., from -59.1‰ to -117‰) all observed in the clathrate (Table 3).
4. Discussion
4.1. Carbonate precipitation scenarios and the role of gas clathrates
Results from mineralogy, petrography, and bulk geochemistry allowed us to reconstruct two different carbonate-precipitation scenarios in the studied cold seeps. First, the mineralogical composition of the two authigenic carbonates showed qualitative differences associable with distinct environmental chemistry in each venting system. The dominance of low-magnesium calcite and iron-rich dolomite in the chimney carbonate suggested an environment of relatively low concentration of sulfates (Greinert et al., 2001; Merinero, 2008). In contrast, the predominance of aragonite in the clathrite sample rather pointed to a relatively cooler and sulfate-richer environment, as the formation of this carbonate polymorph is favored in conditions of high Mg-over-Ca ratios derived from the dissociation of gas hydrates (Yang et al., 2007; Nöthen and Kasten, 2011).
Second, the proportion of autochthonous/allochthonous material in the carbonates appeared to be different in both cold seeps. The relative abundance of minerals such as quartz, montmorillonite, or illite-muscovite in the chimney carbonate (Table 1) indicated a relatively higher input of terrigenous sources in this relative to the clathrite sample. In particular, the origin of montmorillonite could be related to mud volcanoes of the TASYO field, as smectites are the typical mud minerals extruded from them (Martín-Puertas et al., 2007). The allochthonous origin of part of the mineral components in the chimney carbonate was supported by the greater presence of terrigenous material observed in its thin section (Fig. 2a, 2b), which revealed that the chimney was formed in a marine environment of relative high energy, such as that of continuous mixture of Mediterranean and Atlantic water masses in the TASYO field (Llave, 2003; Medialdea, 2005). So much that the reddish-brown tone in the chimney sample (Fig. 1f), as well as iron oxide minerals (Fig. 2a), was caused by oxidation after exposure to oxygenated waters by overlying sediment erosion (Descamps and Swennen, 2002; Magalhães et al., 2012). Additional signs of alteration in the chimney system were the glauconite grains and peloids (Fig. 2a, 2b) likely derived from the erosive impact of the relatively denser Mediterranean waters circulating in depth across the Gibraltar straight (Merinero et al., 2010; Tallobre et al., 2019). In contrast, the brecciation observed in the clathrite (Fig. 2d) is explained as a result of a process of in situ clathrate dissociation which causes the collapse of the clasts and methane expulsion, driving the oxidation of methane and the authigenic formation of the clathrite (Bohrmann et al., 2002). Thus, whereas the chimney carbonate appeared to contain carbon from different sources with considerable contribution of allochthonous organic matter (i.e., surrounding phytoplankton, foraminifers or other type of zooplankton), the clathrite was largely derived from autochthonous seeping methane.
This was supported by the different stable-carbon isotope signals measured in each sample (Table 3), as carbonates inherit the stable isotope composition of their carbon sources (Campbell et al., 2002). In the Pacific clathrite, the low δ13Cinorg value (-44.5‰) reflected the use of 13C-depleted forms of carbon such as methane, which typically shows δ13C values from -20‰ to -45‰ when it is thermogenic or lower than -55‰ when it is biogenic (Whiticar, 1999). In this cold seep, assimilation of 13C-depleted methane through AOM would have induced the precipitation of carbonate upon saturation of the resulting CO2 (Peckmann and Thiel, 2004). In the chimney system in contrast, the considerably enriched δ13Cinorg value measured in the carbonate (-12.5‰; Table 3) denoted mixing sources of carbon, involving marine biomass (from -8‰ to -20‰), marine bicarbonates (from +1‰ to -3‰), as well as abiotic (from 0‰ to -7‰) and, perhaps, thermogenic methane (Blumenberg, 2010 and references therein) and AOM activity (Díaz-del-Río et al., 2003), although likely in minor extent.
As for the oxygen isotopic composition, very similar δ18O values in the chimney (4.0‰) and clathrite (3.6‰) carbonates revealed a similar influence of deep masses of water in both cold seeps. Still, while the δ18O value in the chimney carbonate reflects the isotopic composition of the Gulf of Cádiz ambient seawater (Logvina et al., 2007), the similar δ18O value in the clathrite suggests compositional differences relative to the much more 18O depleted water in the Cascadia Margin (LeGrande and Schmidt, 2006). The relatively enriched δ18O in the clathrite could be the result of clathrate dissociation because clathrate tends to form its structure with 18O and deuterium water molecules (Maekawa and Imai, 2000; Hachikubo et al., 2007). In addition, the slightly enriched δ18O in the chimney carbonate could be related to the relatively higher abundance of dolomite and high-magnesium calcite in this system compared to the Pacific clathrite (Table 1), as dolomite typically shows δ18O values about 3‰ higher than coexisting calcite (δ18O of ca. +3.2‰) and magnesium-rich calcite has δ18O values in between those of dolomite and calcite (Anderson and Arthur, 1983). In sum, despite general similarities, methane-derived authigenic carbonate precipitation in the two cold seeps appeared to occur using different carbon sources.
4.2. Molecular and isotopic evidence of distinct carbon metabolic pathways in the two authigenic carbonates
The inventory of lipid biomarkers preserved in the two authigenic carbonates revealed biological fingerprints associated to different carbon metabolisms in cold seeps (e.g., Elvert et al., 2003; Chevalier et al., 2011). Lipids are cell membrane constituents relatively resistant to diagenetic influences that have specific chemical structures with potential to trace biological sources (Kaneda, 1991; Blumenberg, 2010). Analyzing lipid-derived molecules in combination with their compound-specific stable-carbon isotopic composition may provide information not only about organismic sources but also about environmental conditions, biogeochemical processes, metabolic modes (autotrophy vs. heterotrophy), carbon fixation pathways, or trophic interrelationships (Hayes, 2001; Summons et al., 2008).
Here, the preserved record of lipid biomarkers in the two authigenic carbonates reflected the use of distinct carbon sources in the two venting systems. In the clathrite cold seep, the precipitation of carbonates seemed largely influenced by AOM activity. On the one hand, the molecular distribution of the nonpolar and polar lipidic compounds in the clathrite showed a dominance of archaeal-specific isoprenoid hydrocarbons (crocetane or PMI; Fig. 3b) and dialkylglicerols (dihydrophytol, archaeol, or sn-2-hydroxyarchaeol; Fig. 5b), respectively. Archaeol and sn-2-hydroxyarchaeol are lipid biomarkers related to archaeal anaerobic methanotrophs (ANME) of the clades ANME-1 and ANME-2 (Blumenberg et al., 2004), whereas PMI and crocetane are derived from methanogenic and/or methanotrophic archaea (Tornabene et al., 1979; Robson and Rowland, 1993). On the other hand, the diagnostic archaeal biomarkers yielded very depleted compound-specific δ13C values (from -105‰ to -118‰; Table 3), consistent with the consumption of 13C-depleted methane.
In addition to the archaeal activity, the detection in the clathrite of carboxylic acids such as the iso/anteiso-C15:0 and the MUFA C16:1ω7 (Fig. 4b) denoted additional occurrence of bacterial sulfate reduction (Pancost et al., 2001; Thiel et al., 2001; Blumenberg et al., 2004). The depleted isotopic composition (δ13C from -59‰ to -88‰; Table 3) of the SRB biomarkers suggests a syntrophic association with the archaea (Boetius et al., 2000; Niemann and Elvert, 2008), such as that developed by SRB of the Desulfosarcina/Desulfococcus cluster with methanotrophic archaea from the ANME-1 and ANME-2 clades (Boetius et al., 2000) or by Desulfobulbus spp. SRB with ANME-3 (Niemann et al., 2006). Microbial syntrophy of archaea with SRB allows AOM to occur via reverse methanogenesis, whereby sulfate acts as electronic acceptor facilitating methane metabolism (Skennerton et al., 2017). At the Hydrate Ridge in the Cascadia convergent margin, hydrate destabilization supplies free methane sufficiently to exceed a threshold beyond which net methane oxidation is favored (Elvert and Suess, 1999), with eventually precipitating carbonates (e.g., clathrite) retaining an isotopically depleted, AOM biosignature.
In contrast, the chimney carbonate reflected a quite different biological signal, at both molecular and isotopic level. No evidence of archaeal biomarkers was found in the form of crocetane, PMI (Fig. 3), archaeol, or sn-2-hydroxyarchaeol (Fig. 5). Instead, a general dominance of unspecific bacterial biomarkers (straight chains of LMW) was observed in the three polarity fractions (Figs. 3–5). Still, some dihydrophytol was detected in the chimney carbonate polar fraction (Fig. 5a), but its origin was interpreted as different from that in the Pacific clathrite according to its much higher δ13C ratio (i.e., -31.9‰ versus -109‰). Overall, all lipid compounds in the chimney carbonate showed compound-specific δ13C values considerably enriched (from -24.9‰ to -31.9‰) relative to those in the clathrite (Table 3), which were associated with a prevailing autotrophic metabolism through the Calvin cycle (Hayes, 2001) rather than with the assimilation of methane. That was also the case of phytane, an isoprenoid that can also come from archaeal membrane lipids (i.e., archaeol) but was here related to algal and/or microbial chlorophylls and bacteriochlorophylls (Peters et al., 2005) due to its high δ13C (-24.9‰).
The occurrence in the chimney carbonate of sulfate reduction activity was also inferred from the detection of iso/anteiso-C15:0 and C16:1ω7 carboxylic acids (Fig. 4a) but at concentrations not high enough as to be measured for their δ13C ratio. This precluded us from assessing the syntrophic involvement of putative SRB in AOM in the chimney system, where SRB biomarkers would exhibit depleted δ13C ratios similar to those in the clathrite (Table 3). Still, despite not having such data, the participation of SRB in AOM in syntrophy with methanotrophic archaea was considered unlikely given the lack of detection of archaeal biomarkers in the chimney carbonate.
Some investigation in venting systems from the Gulf of Cádiz attributed to n-alkanes from C16 to C25 from sedimentary deposits (contourites) a thermogenic origin based on their δ13C values ranging from -20‰ to -37‰ (González et al., 2012). Other works in the Gulf of Cádiz described the implication of AOM in the formation of authigenic carbonates associated to methane vents, based on the detection of archaeal-specific lipid biomarkers and more or less depleted δ13C values (down to -82‰; Díaz-del-Río et al., 2003; Merinero et al., 2012; Wang et al., 2015; Feenstra et al., 2020). In some cases, the occurrence of AOM was clearly deduced from strongly depleted compound-specific δ13C (as low as -100‰) in archaeal membrane lipids (e.g., Feenstra et al., 2020). In other cases, the link between moderately depleted δ13C values (from -9‰ to -46‰) and AOM was not that clear and just based on bulk isotopic composition of carbonates, and factors such as thermal maturity and/or aerobic degradation were argued to have possibly played a role in increasing the original more depleted ratios (Díaz-del-Río et al., 2003; Merinero et al., 2012; Wang et al., 2015).
Here, our total lack of archaeal biomarkers together with the relatively high δ13C values (typical of Calvin carbon fixation; Hayes, 2001) measured in all the inorganic carbon (-12.5‰), total organic carbon (-22.6‰), and lipid biomarkers (from -24.9‰ to -31.9‰) precluded us from linking the formation of authigenic carbonate in the chimney system to AOM. We argue that, if AOM activity took place in the chimney system, the resulting carbonate and preserved molecular biomarkers should retain an isotopic signature reflecting a strong fractionation due to the consumption of methane (Blumenberg, 2010). Instead, both the bulk carbonate and the preserved lipidic compounds showed δ13C values enriched by ∼7–26‰ relative to the regional methane (δ13C from -39‰ to -42‰; Stadnitskaia et al., 2006). We interpreted these values in terms of mixed sources of carbon involving thermogenic methane (Blumenberg, 2010), as well as pelagic phytodetritus, marine phytoplankton (mostly cyanobacteria and microalgae), or zooplankton (e.g., Demopoulos et al., 2017). In such a scenario, SRB could exist independently of AOM activity, developing either autotrophically, fixing carbon through the reductive acetyl-CoA pathway (δ13C from -28‰ to -44‰; Preuss et al., 1989), or heterotrophically, growing on marine biomass (of δ13C typically higher than -25‰; Meyers, 1994). The observation of framboidal pyrite aggregates in the chimney carbonate (Fig. 2b, 2c) supported the activity of SRB (Berner et al., 1985; León et al., 2006) in this cold seep. However, in contrast to the clathrite, sulfate reduction in the chimney was not coupled to archaeal AOM but instead associated with lighter sources of carbon (i.e., of relatively less negative δ13C).
Altogether, mineralogical, molecular, and isotopic evidence recreated two different carbon (and sulfate)–cycling scenarios in the studied cold seeps, where distinct submarine environments with different water chemistry seem to condition the use of different sources and processing of carbon, thus resulting in the precipitation of different carbonate phases and textures (aragonite and high-magnesium calcite versus dolomite and low-magnesium calcite).
4.3. Implications for interpreting potential carbon cycling in the icy worlds
Our multidisciplinary re-creation of the different carbon sources resulting in the precipitation of methane-derived authigenic carbonates of distinct isotopic signatures has implications for elucidating potential carbon tracks in the icy worlds (Fig. 6). Gas seeping and carbon sequestration via clathration and carbonatation processes could affect the carbon cycle in ocean worlds despite the origin of methane. On Earth the main primary chemicals (O2, CO2, nitrate, or sulfate) involved in the carbon cycle are ultimate products of photosynthetic activity, which is not the case on icy worlds. On Enceladus or Europa, carbon could be released as carbon dioxide and/or methane during rock mantle degassing (Glein et al., 2015) or serpentinization (Hussmann et al., 2010) of olivine-rich core rocks involving Fischer-Tropsch reactions (Sleep et al., 2004; Etiope and Sherwood-Lollar, 2013), as initial input of the cycle. In the methane-derived authigenic carbonates studied here, carbon may be stocked in subsurface methane reservoirs (Greinert et al., 2010), as in the clathrite system. In the chimney system, methane that has passed through the sediments has been oxidized to carbon dioxide by oxygenated water and then precipitated as carbonates. Microbial methanogenic sources of carbon were deduced to be a minority in the chimney, and thermogenic methane and allochthonous mixed sources of carbon prevailed. Furthermore, although less common because there are few known geological sites, methane may also have an abiogenic origin related to serpentinization of ophiolites, as is thought to be the case for the methane expelled by the cold seeps near Elba Island (Sciarra et al., 2019).
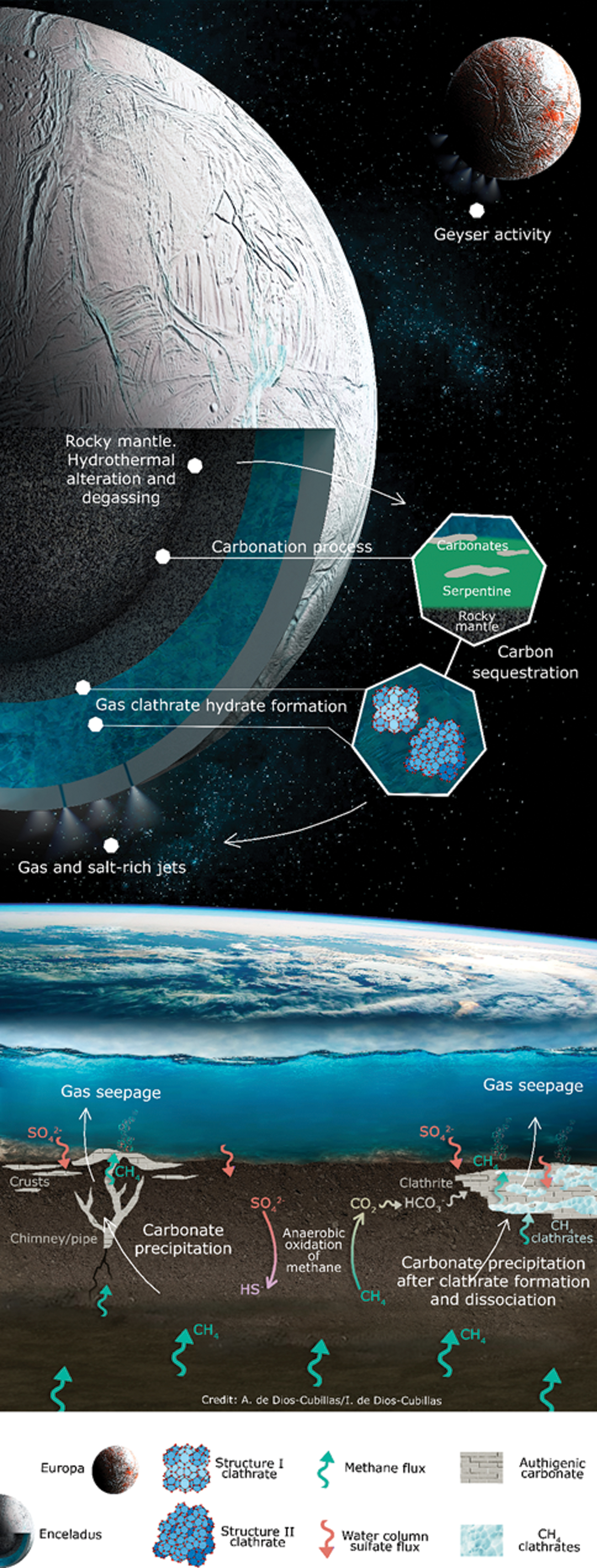
Sketch of comparable carbon pathways in terrestrial cold seep environments and analogous ocean worlds. An interior cross-section of Enceladus is shown at the top, with major carbon sources represented by mantle degassing and serpentinization mechanisms (CH4, CO2), and major carbon sinks by carbonation and clathration processes. At the bottom, a gas venting cross-section illustrates major carbon sources (biogenic or thermogenic methane) and sinks (carbonate precipitation) in terrestrial cold seeps. Color images are available online.
On Earth's continental margins and maybe on icy moons, there are regimes of high pressure and low temperature in which methane clathrate forms and stabilizes (Prieto-Ballesteros et al. 2005; Bouquet et al., 2015; Rajput and Thakur, 2016), sequestering carbon until the crystalline solids dissociate by changes in physicochemical parameters. In fact, Bouquet et al. (2015) found that Enceladus' ocean may have conditions quite favorable for methane clathrate formation. The destabilization of clathrate hydrates results into secondary releases of carbon. The liberated carbon-bearing gases may in turn end up precipitating upon saturation as authigenic carbonate if physical-chemical (e.g., alkalinity or P/T) conditions are favorable (Kennett and Fackler-Adams, 2000), thus constituting a new carbon sink (i.e., clathrite) (Fig. 6). On icy moons, a first carbonate process would take place when dissolved carbon dioxide reacts with the products of the aqueous alteration of silicate minerals—present here in the Pacific clathrate—in the rocky core (Glein and Waite, 2020), as well as through the oxidation of methane. In terrestrial cold seeps, the oxidation of methane takes place upon deficiency of oxygen by the syntrophic action of methanogenic/methanotrophic archaea with SRB acting as electron acceptors (Elvert and Suess, 1999). On Enceladus, it has been proposed that serpentinization processes increase its ocean alkalinity (Kotlarz et al., 2020), turning the environment favorable for chemolithoautotrophic life, such as methanotrophic archaea (McKay et al., 2008). Serpentinization reactions in Enceladus generate hydrogen that, under particular conditions, could supply chemical energy to support a hypothetical chemoautotrophic life on Enceladus.
If chemosynthetic life ever arose in the ocean of any icy world, its development could be comparable to that on deep-sea vents on Earth. In the latter systems, methanogenic archaea able to operate in reverse methanogenesis mode are the only known microorganisms capable of oxidizing methane in the absence of oxygen (Taubner et al., 2015, 2018) in consortium with SRB or metal oxides (see review by Timmers et al., 2017). The extremophilic methanotrophs can be found on Earth over a wide range of pH (4.5–10.2), temperatures (<0–122°C), and pressures (0.005–759 bar) (Taubner et al., 2015). Their distribution may be compatible with the proposed theoretical conditions in Enceladus' ocean, where temperature ranges from 0°C to 90°C, pressure from 40 to 100 bar, pH from 8.5 to 13.5, and salinity is similar to that in Earth's ocean (Glein et al., 2015; Hsu et al., 2015). If chemosynthetic life ever existed on Enceladus promoting biogeochemical cycles, it is reasonable to think that a biological signal should be somehow retained in the geological carbon reservoirs. Assuming carbonation and clathration as primary processes for carbon sequestration in cold seep environments (Fig. 6), analyzing the stable carbon isotope composition in such carbon sinks seems crucial for detecting biological fingerprints (Taubner et al., 2018). It is important to mention that the use of carbon isotopic fractionation as a tool for apportioning carbon source requires a deep knowledge of the different reservoirs that are involved in a biogeochemical cycle; thus this approach is more resource-intense than the molecular approaches. While the carbonate compensation depth could prevent the precipitation of carbonates at higher pressures, clathrates would primarily sink the carbon-bearing gasses under the planetary ocean lysocline. Gas retention in durable clathrate deposits is strongly dependent on the change of physical conditions such as pressure or temperature. The occurrence of the retention/release of volatiles by clathrate formation/dissociation due to the planetary dynamics and further clathrite precipitation would have some environmental effects such as constraint of the availability of essential elements, fractionation of the better guest molecules, modification of the acidity of the aqueous solutions, or concentration of nutrients if the water activity reduces locally. These factors should be considered when designing astrobiological missions to explore Enceladus or similar ocean moons.
5. Conclusion
Some icy worlds have great astrobiological potential due to the presumable habitability of their internal oceans. A biogeochemical approach was performed here on methane-derived authigenic carbonates from cold seep environments with the aim of investigating carbon routes that help on astrobiology exploration. Molecular and isotopic patterns of cell-membrane molecules (lipid biomarkers) preserved in the two carbonate scenarios (chimney and clathrite) revealed different origins for the parent methane of two authigenic carbonates. Precipitation of the clathrite most likely stemmed from the syntrophic AOM activity of methanotrophic archaea and SRB, whereas the chimney carbonate resulted from biomediated precipitation of carbonate coming from mixed sources, mainly including thermogenic methane and allochthonous carbon forms. Identifying carbon pathways, sources, and sinks in terrestrial cold seeps is essential for elucidating a hypothetical carbon cycle in the Solar System icy worlds. This work provides a baseline for interpreting molecular and isotopic signals in future exploration missions to the icy moons in the search for unequivocal biomarkers.
Footnotes
Author Contribution Statement
D. Carrizo led the manuscript building, interpretation, discussion, and writing. O. Prieto-Ballesteros conceived the original idea. D. Carrizo and A. de Dios processed the samples, analyzed the lipid biomarkers and their stable-carbon isotopic composition. L. Sánchez-García contributed to interpret the molecular and isotopic data. A de Dios and O. Prieto-Ballesteros performed and interpreted the mineralogical and geochemical analysis. L. Sánchez-García, O. Prieto-Ballesteros, and Iván López discussed and revised the manuscript.
Funding Statements
This study was partially funded by the Spanish Agencia Estatal de Investigación (AEI) and Fondo Europeo de Desarrollo Regional (FEDER), through the projects RYC-2014-19446 (D. Carrizo), PID2019-107442RB-C32 (O. Prieto-Ballesteros, D. Carrizo, and Iván López), RYC2018-023943-I (L. Sánchez-García), and MDM-2017-0737 Unidad de Excelencia “María de Maeztu”–Centro de Astrobiología (INTA-CSIC) by the Spanish Ministry of Science and Innovation/State Agency of Research MCIN/AEI/10.13039/501100011033 and by “ERDF A way of making Europe.” Ana de Dios is supported by the AEI pre-doctoral contract under the project “SOLAR SYSTEM” (MDM-2017-0737-19-1). Thanks to Jens Greinert/Kathy Cambell, and Luis Gago Duport for providing respectively the samples from the Hydrate Ridge and Cadiz Gulf.
Author Disclosure Statement
The author D. Carrizo, A de Dios, L. Sánchez-García, and O. Prieto-Ballesteros have research support from the Centro de Astrobiología, in Madrid.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Abbreviations Used
Associate Editor: Kathleen Campbell
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.